
Predicting the Potential Distribution of Haloxylon ammodendron under Climate Change Scenarios Using Machine Learning of a Maximum Entropy Model


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Species-Occurrence Records
2.3. Environment Variables
2.4. Maxent Model
2.5. Division of the Potentially Suitable Area
3. Results
3.1. Model Performance and AUC
3.2. Analysis of Variable Contributions
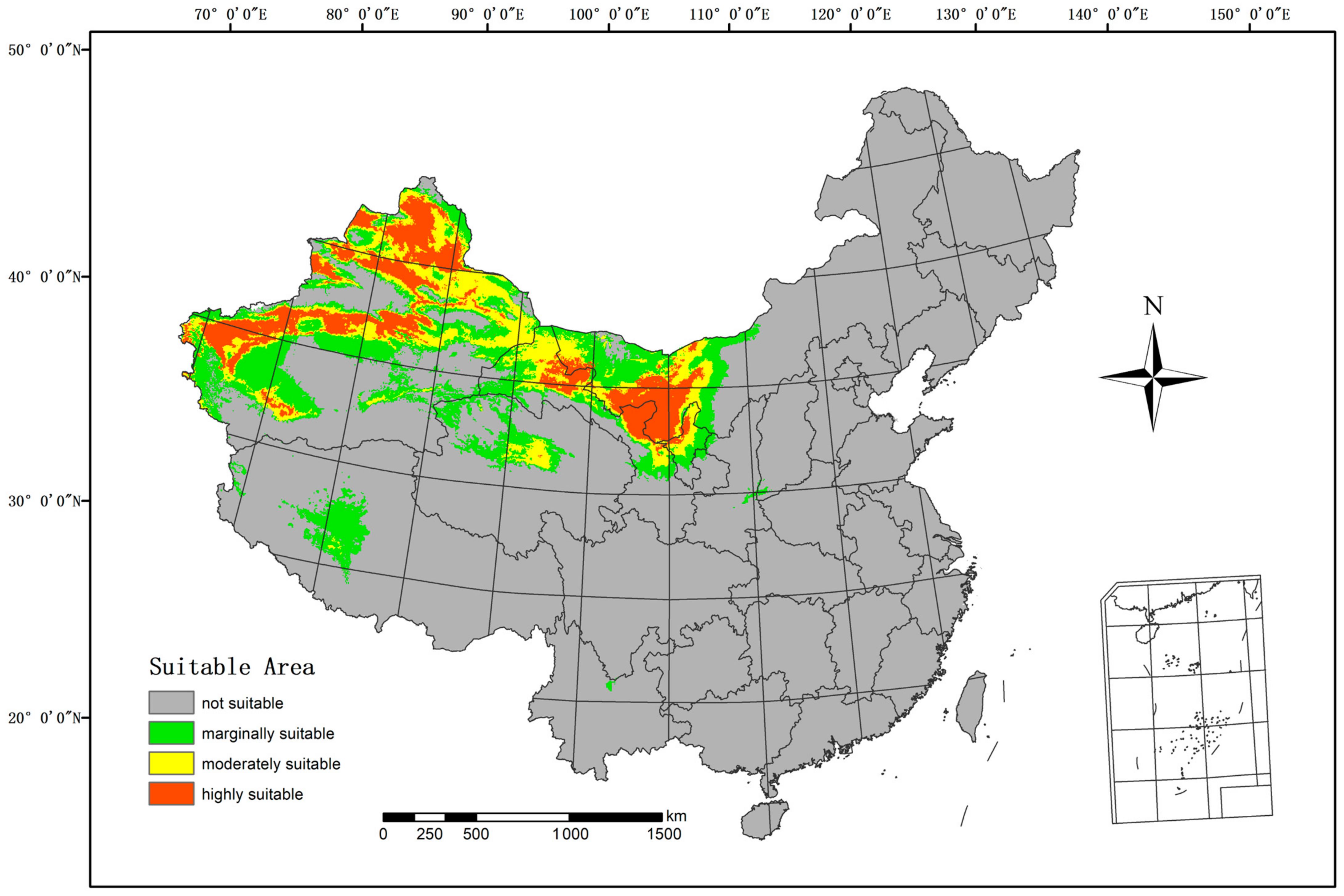
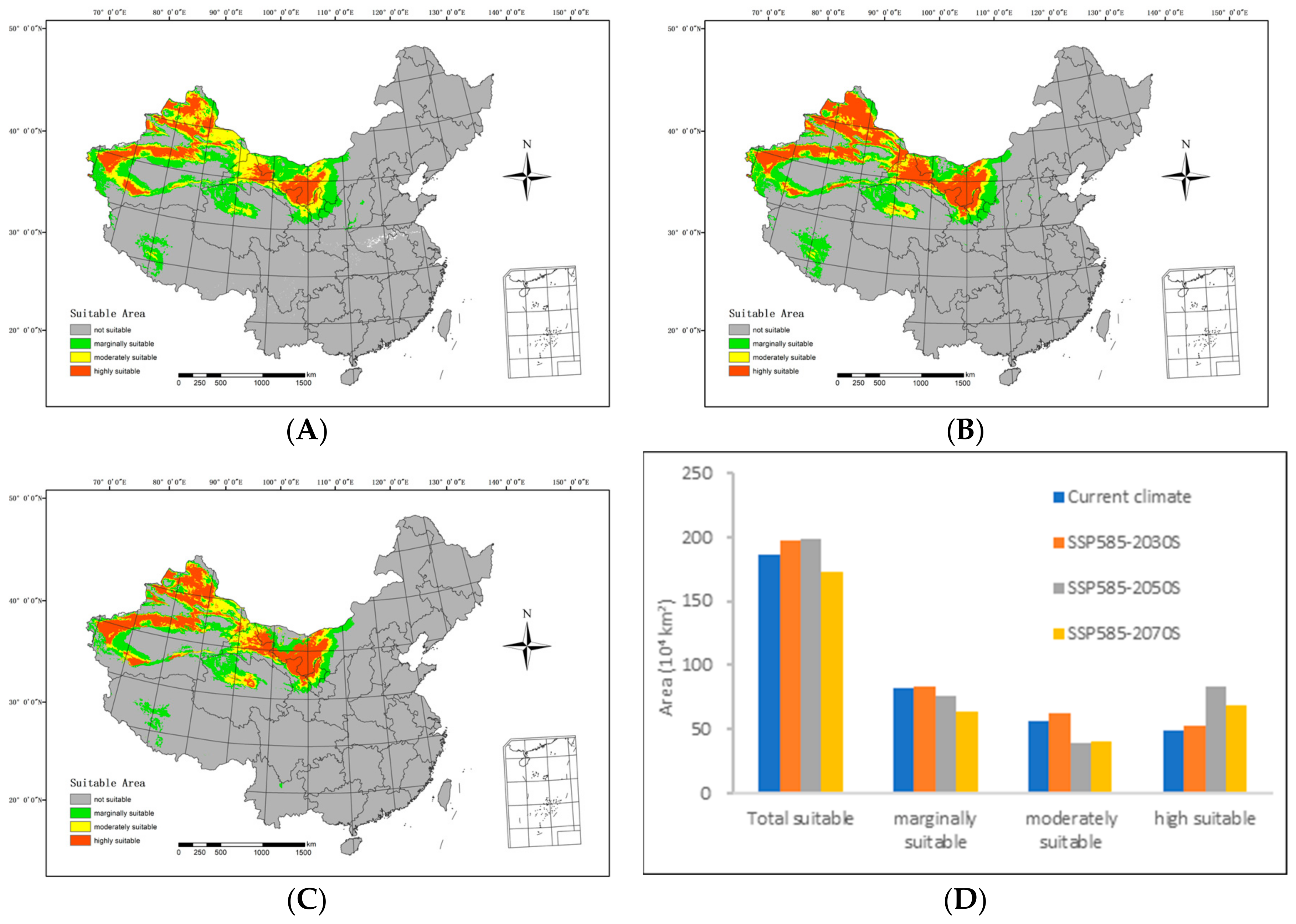
3.3. Potentially Suitable Area under Current and Future Scenarios
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- IPCC. 2022: Climate Change 2022: Impacts, Adaptation and Vulnerability. Contribution of Working Group II to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2022. [Google Scholar]
- Di Febbraro, M.; Bosso, L.; Fasola, M.; Santicchia, F.; Aloise, G.; Lioy, S.; Tricarico, E.; Ruggieri, L.; Bovero, S.; Mori, E.; et al. Different facets of the same niche: Integrating citizen science and scientific survey data to predict biological invasion risk under multiple global change drivers. Glob. Chang. Biol. 2023, 29, 5509–5523. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.T.; Grote, U.; Neubacher, F.; Do, M.H.; Paudel, G.P. Security risks from climate change and environmental degradation: Implications for sustainable land use transformation in the Global South. Curr. Opin. Environ. Sustain. 2023, 63, 101322. [Google Scholar] [CrossRef]
- Easterling, D.R.; Meehl, G.A.; Parmesan, C. Climate Extremes: Observations, Modeling, and Impacts. Science 2020, 289, 2068–2074. [Google Scholar] [CrossRef] [PubMed]
- Parmesan, C.; Yohe, G. A globally coherent fingerprint of climate change impacts across natural systems. Nature 2003, 421, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Tian, H.; Wang, Y. Vulnerability of 208 endemic or endangered species in China to the effects of climate change. Reg. Environ. Chang. 2012, 13, 843–852. [Google Scholar] [CrossRef]
- Mccarty, J.P. Ecological consequences of recent climate change. Conserv. Biol. 2001, 15, 320–331. [Google Scholar] [CrossRef]
- CMA Climate Change Center. Blue Book on Climate Change in China; Science Press: Beijing, China, 2023. [Google Scholar]
- Balfagón, D.; Terán, F.; de Oliveira, T.; Santa-Catarina, C.; Gómez-Cadenas, A. Citrus rootstocks modify scion antioxidant system under drought and heat stress combination. Plant Cell Rep. 2022, 41, 593–602. [Google Scholar] [CrossRef] [PubMed]
- You, J.; Qin, X.; Ranjitkar, S.; Lougheed, S.C.; Wang, M.; Zhou, W.; Ouyang, D.; Zhou, Y.; Xu, J.; Zhang, W. Response to climate change of montane herbaceous plants in the genus Rhodiola predicted by ecological niche modelling. Sci. Rep. 2018, 8, 5879. [Google Scholar] [CrossRef]
- Fujii, T.; Ueno, K.; Shirako, T.; Nakamura, M.; Minami, M. Identification of Lagopusmuta japonica food plant resources in the Northern Japan Alps using DNA metabarcoding. PLoS ONE 2022, 17, e0252632. [Google Scholar] [CrossRef]
- Zhuang, Y.L.; Zhao, W.Z. Dew formation and its variation in H. ammodendron plantations at the edge of a desert oasis, northwestern China. Agric. For. Meteorol. 2017, 247, 541–550. [Google Scholar] [CrossRef]
- Wang, J.H.; Ma, Q.L. Study on restoration strategies characteristics and status of degenerated artificial Haloxylon ammdendron communities at the edge of Minqin oasis. Acta Bot. Boreali-Occident. Sin. 2003, 23, 2017–2112. [Google Scholar]
- Xu, D.; Li, X.; Jin, Y.; Zhuo, Z.; Yang, H.; Hu, J.; Wang, R. Influence of climatic factors on the potential distribution of pest Heortia vitessoides Moore in China. Glob. Ecol. Conserv. 2020, 23, e01107. [Google Scholar] [CrossRef]
- Zhang, H.D. The forest type and its characteristics of the Holoxylon in Junggar Basin in Xinjiang. J. Des. Res. 1990, 10, 41–49. [Google Scholar]
- Zhang, J.B.; Wang, B.F.; Hao, Y.G. Geographical distribution, suitable environment and provenance variation of Haloxylon ammdendron (C.A.M) Bge in China. J. Arid Res. Environ. 2010, 24, 166–171. [Google Scholar]
- Zhang, L.Y. The H. ammodendron and H. persicum in Xinjiang Desert. Plant Mess. 2022, 5, 4–5. [Google Scholar]
- Ma, H.B.; Bao, G.X.; Ma, W.D.; Rong, Z.J.; Wang, X.Y.; Li, B. The resource, protection and utilization of H. ammodendron deserted grassland in Inner Mongolia. Prat. Sci. 2000, 17, 1–5. [Google Scholar]
- Tobe, K.; Li, X.; Omasa, K. Effects of sodium chloride on seed germination and growth of two Chinese desert shrubs, H. ammodendron and H. (Chenopodiaceae). Aus. J. Bot. 2000, 48, 455–460. [Google Scholar] [CrossRef]
- Guo, Q.S.; Wang, C.L.; Guo, Z.H.; Tan, D.Y.; Shi, Z.M. Geographic Distribution of Existing Haloxylon Desert Vegetation and Its Patch character in China. Sci. Sil. Sin. 2005, 41, 2–7. [Google Scholar]
- Ma, S.M.; Wei, B.; Li, X.C.; Luo, C.; Sun, F.F. The impacts of climate change on the potential distribution of H. ammodendron. Chin. J. Ecol. 2017, 36, 1243–1250. [Google Scholar]
- Zeng, X.L.; Liu, T.; Zhang, W.B.; Sun, Q.M.; Shen, X.Y.; Si, L.M. Variations in groundwater levels and quality and their effects on vegetation in the western Grurbantonggut Desert. Acta Ecol. Sin. 2012, 32, 1490–1501. [Google Scholar] [CrossRef]
- Zhang, R.; Liu, T. Plant species diversity and community classification in the southern Gurbantunggut Desert. Acta Ecol. Sin. 2012, 32, 6056–6066. [Google Scholar] [CrossRef]
- Editorial Committee of Chinese Vegetation Atlas, Chinese Academy of Sciences. Vegetation Atlas of China; Science Press: Beijing, China, 2001; p. 588. [Google Scholar]
- Department of Protected Areas and Species Management, Department of Nature Conservation, State Environmental Protection Administration. Protection and Research of Rare and Endangered Plants; China Environmental Science Press: Beijing, China, 1991; p. 157. [Google Scholar]
- Phillips, S.; Anderson, R.R.; Schapire, R. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Jayathilake, D.R.; Costello, M.J. A modelled global distribution of the kelp biome. Biol. Conserv. 2020, 252, 108815. [Google Scholar] [CrossRef]
- Qin, A.L.; Jin, K.; Batsaikhan, M.E. Predicting the current and future suitable habitats of the main dietary plants of the Gobi Bear using MaxEnt modeling. Glob. Ecol. Conserv. 2020, 22, e01032. [Google Scholar] [CrossRef]
- Yang, J.T.; Huang, Y.; Jiang, X. Potential geographical distribution of the edangred plant Isoetes under human activities using MaxEnt and GARP. Glob. Ecol. Conserv. 2022, 38, e02186. [Google Scholar] [CrossRef]
- Dong, P.B.; Wang, L.Y.; Wang, L.J.; Jia, Y.; Li, Z.H.; Bai, G.; Zhao, R.M.; Liang, W.; Wang, H.Y.; Guo, F.X.; et al. Distributional Response of the Rare and Endangered Tree Species Abies chensiensis to Climate Change in East Asia. Biology 2022, 11, 1659. [Google Scholar] [CrossRef] [PubMed]
- Bosso, L.; Mucedda, M.; Fichera, G.; Kiefer, A.; Russo, D. A gap analysis for threatened bat populations on Sardinia. Hystrix 2016, 27, 212. [Google Scholar]
- Wilkening, J.; Perason-Prestera, W.; Mungi, N.A.; Bhattacharyya, S. Endangered species management and climate change: When habitat conservation becomes a moving target. Wildl. Soc. Bull. 2019, 43, 11–20. [Google Scholar] [CrossRef]
- Yeh, H.T.; Cheah, H.Y.; Chiu, M.C.; Liao, J.R.; Ko, C.C. Assessment of potential invasion for six phytophagous quarantine pests in Taiwan. Sci. Rep. 2021, 11, 10666. [Google Scholar] [CrossRef]
- Xu, L.G.; Zhou, H.F.; Du, L.; Piao, Z.Y. Spatio-temporal change and trend of precipitation in northwest China during 1951–2008. J. Desert Res. 2015, 35, 724–732. [Google Scholar]
- Digger, P.J. Statistical Analysis of Spatial Point Patterns, 2nd ed.; Arnold: London, UK, 2003. [Google Scholar]
- Zhao, J.Q.; Shi, J. Prediction of the potential geographical distribution of Obolodiplosis robiniae in China based on a novel Maximum Entropy model. Sci. Silvae Sin. 2019, 55, 118–127. [Google Scholar]
- Phillips, S.J.; Anderson, R.P.; Dudík, M. Opening the black box: An open-source release of Maxent. Ecography 2017, 40, 887–893. [Google Scholar] [CrossRef]
- Zhuang, H.F.; Zhang, Y.B.; Wang, W. Optimized hot spot analysis for probability of species distribution under different spatial scales based on MaxEnt model: Manglietia insignis case. Biol. Sci. 2018, 26, 931–940. [Google Scholar] [CrossRef]
- Zhao, X.J.; Gong, J.X.; Zhao, S.S. Impact of sample size and spatial distribution on species distribution model. J. Lan. Univ. Nat. Sci. 2018, 54, 70–77. [Google Scholar]
- Huang, J.H.; Li, G.Q.; Li, J. Projecting the range shifts in climatically suitable habitat for Chinese sea buckthorn under climate change scenarios. Forests 2017, 9, 9. [Google Scholar] [CrossRef]
- Root, T.L.; Price, J.T.; Hall, K.R. Fingerprints of global warming on wild animals and plants. Nature 2003, 421, 57–60. [Google Scholar] [CrossRef]
- Zhang, Q.; Yang, J.H.; Wang, P.L. Progress and prospect on climate warming and humidification in Northwest China. Chin. Sci. Bull. 2023, 68, 1814–1828. [Google Scholar] [CrossRef]
- Xu, G.Q.; Li, Y. Rooting depth and leaf hydraulic conductance in the xeric tree Haloxyolon ammodendron growing at sites of contrasting soil texture. Funct. Plant Biol. 2008, 35, 1234–1242. [Google Scholar] [CrossRef]
- Xue, Y.; Han, P.; Feng, G.H. Change Trend of the Precipitation and Air Temperature in Xin jiang since recent 50 years. Arid Zone Res. 2003, 20, 127–130. [Google Scholar]
- Li, L.; Shen, H.Y.; Li, H.M.; Xiao, J.S. Regional differences of climate change in Qaidam Basin and its contributing factors. J. Nat. Res. 2015, 30, 641–650. [Google Scholar]
- Huang, P.Y.; Li, Q.J.; Yuan, Q.F. Effects of climate change on H. ammodendron community in southern edge of Zhunger Basin. Acta. Ecol. Sin. 2008, 28, 6051–6059. [Google Scholar]
- Liu, H.L.; Tao, Y.; Qiu, D.; Zhang, D.Y.; Zhang, Y.K. Effects of artificial sand fixing on community characteristics of a rare desert shrub. Conserv. Biol. 2013, 27, 1011–1019. [Google Scholar] [CrossRef] [PubMed]
- Ren, Z.X.; Yang, D.Y. Climate change and surface runoff change impact on desertification on the arid area of northwest China in recent 50 years. J. Arid Land Res. Environ. 2008, 22, 91–95. [Google Scholar]
- Shi, S.B.; Yang, Z.; Wu, Y.H.; Zuo, Z. Desert, Sandy Land and Psammophytes in China; China Agricultural Science and Technology Press: Beijing, China, 2013; pp. 238–239. [Google Scholar]
- Borcard, D.; Legendre, P.; Drapeau, P. Partialling out the spatial component of ecological variation. Ecology 1992, 73, 1045–1055. [Google Scholar] [CrossRef]
- Sillero, N. What does ecological modelling model? A proposed classification of ecological niche models based on their underlying methods. Ecol. Mod. 2011, 222, 1343–1346. [Google Scholar] [CrossRef]
- Chang, H.; Liu, T.; Wang, D.W.; Ji, X.R. Haloxylon ammodendron’s potential distribution under climate change in arid area of northwest China. J. Des. Res. 2019, 39, 110–118. [Google Scholar]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiao, F.; Liu, Q.; Qin, Y. Predicting the Potential Distribution of Haloxylon ammodendron under Climate Change Scenarios Using Machine Learning of a Maximum Entropy Model. Biology 2024, 13, 3. https://doi.org/10.3390/biology13010003
Xiao F, Liu Q, Qin Y. Predicting the Potential Distribution of Haloxylon ammodendron under Climate Change Scenarios Using Machine Learning of a Maximum Entropy Model. Biology. 2024; 13(1):3. https://doi.org/10.3390/biology13010003
Chicago/Turabian StyleXiao, Fengjin, Qiufeng Liu, and Yun Qin. 2024. "Predicting the Potential Distribution of Haloxylon ammodendron under Climate Change Scenarios Using Machine Learning of a Maximum Entropy Model" Biology 13, no. 1: 3. https://doi.org/10.3390/biology13010003
APA StyleXiao, F., Liu, Q., & Qin, Y. (2024). Predicting the Potential Distribution of Haloxylon ammodendron under Climate Change Scenarios Using Machine Learning of a Maximum Entropy Model. Biology, 13(1), 3. https://doi.org/10.3390/biology13010003