
Enhanced 3T3-L1 Differentiation into Adipocytes by Pioglitazone Pharmacological Activation of Peroxisome Proliferator Activated Receptor-Gamma (PPAR-γ)

 , , , , , and
, , , , , and 
Abstract
:Simple Summary
Abstract
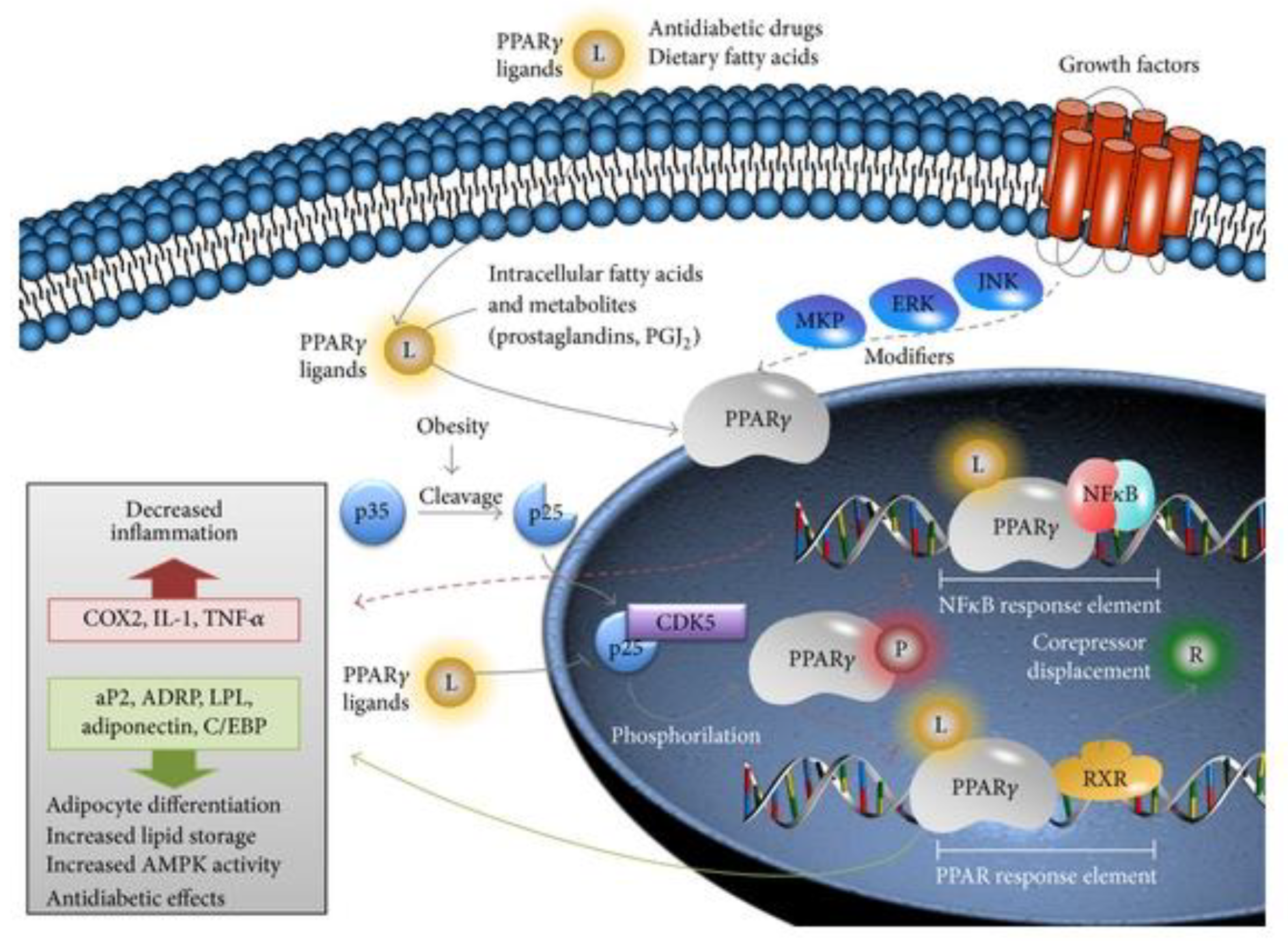
1. Introduction
2. Materials and Methods
2.1. 3T3-L1 Adipocytes Differentiation
2.2. Evaluation of Lipid Droplets Accumulation via the Oil-Red-O Technique
2.3. Adipocyte RNA Extraction and Gene Expression via RT-qPCR
2.4. Cell Cultures
2.5. Evaluation of the Adipocyte Secretome for the Cellular Viability (MTT) and Proliferation (BrdU) of the PC3 and RAW264.7 Cell Lines
2.6. Evaluation of the Adipocyte Secretome in the Inflammation of the PC3 and RAW264.7 Cell Lines
2.7. Statistical Analysis
3. Results
3.1. Pioglitazone Increases Lipid Droplet Accumulation in Differentiated Adipocytes
3.2. Pioglitazone Increases PC3 and Decreases RAW264.7 Mitochondrial Metabolic Activity and Cellular Proliferation
3.3. PC3 Revealed a Higher Expression of Anti-Inflammatory than Pro-Inflammatory Cytokine Production, While RAW264.7 Does the Opposite
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Spencer, M.; Yang, L.; Adu, A.; Finlin, B.S.; Zhu, B.; Shipp, L.R.; Rasouli, N.; Peterson, C.A.; Kern, P.A. Pioglitazone Treatment Reduces Adipose Tissue Inflammation through Reduction of Mast Cell and Macrophage Number and by Improving Vascularity. PLoS ONE 2014, 9, e102190. [Google Scholar] [CrossRef] [PubMed]
- Okada, K.; Hosooka, T.; Shinohara, M.; Ogawa, W. Modulation of Lipid Mediator Profile May Contribute to Amelioration of Chronic Inflammation in Adipose Tissue of Obese Mice by Pioglitazone. Biochem. Biophys. Res. Commun. 2018, 505, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Pereira, A.C.; Oliveira, R.; Castro, A.C.; Fernandes, R. Does Pro12Ala Polymorphism Enhance the Physiological Role of PPAR γ 2? PPAR Res. 2013, 2013, 7–9. [Google Scholar] [CrossRef] [Green Version]
- White, U.; Fitch, M.D.; Beyl, R.A.; Hellerstein, M.K.; Ravussin, E. Adipose Depot-Specific Effects of 16 Weeks of Pioglitazone on in Vivo Adipogenesis in Women with Obesity: A Randomised Controlled Trial. Diabetologia 2021, 64, 159–167. [Google Scholar] [CrossRef] [PubMed]
- Miyazawa, M. Pioglitazone Inhibits Periprostatic White Adipose Tissue Inflammation in Obese Mice. Physiol. Behav. 2017, 176, 139–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, M.; Mohapatra, J.; Malik, U.; Nagar, J.; Chatterjee, A.; Ramachandran, B.; Jain, M.R. Effect of Pioglitazone on Metabolic Features in Endotoxemia Model in Obese Diabetic Db/Db Mice. J. Diabetes 2017, 9, 613–621. [Google Scholar] [CrossRef]
- Cariou, B.; Charbonnel, B.; Staels, B. Thiazolidinediones and PPARγ Agonists: Time for a Reassessment. Trends Endocrinol. Metab. 2012, 23, 205–215. [Google Scholar] [CrossRef]
- DeFronzo, R.A.; Inzucchi, S.; Abdul-Ghani, M.; Nissen, S.E. Pioglitazone: The Forgotten, Cost-Effective Cardioprotective Drug for Type 2 Diabetes. Diabetes Vasc. Dis. Res. 2019, 16, 133–143. [Google Scholar] [CrossRef] [Green Version]
- Lian, Q.X.; Deng, H.Z.; Chen, K.Y.; Deng, H. Role of Peroxisome Proliferator-Activated Receptor (PPARγ) in Metabolic Disorders in SGA with Catch-Up Growth. Obesity 2018, 26, 88–93. [Google Scholar] [CrossRef] [Green Version]
- Ren, R.; Chen, Z.; Zhao, X.; Sun, T.; Zhang, Y.; Chen, J.; Lu, S.; Ma, W. A Possible Regulatory Link between Twist 1 and PPARγ Gene Regulation in 3T3-L1 Adipocytes. Lipids Health Dis. 2016, 15, 189. [Google Scholar] [CrossRef] [Green Version]
- Unno, Y.; Yamamoto, H.; Takatsuki, S.; Sato, Y.; Kuranaga, T.; Yazawa, K.; Ono, Y.; Wakimoto, T. Palmitoyl Lactic Acid Induces Adipogenesis and a Brown Fat-like Phenotype in 3T3-L1 Preadipocytes. Biochim. Biophys. Acta -Mol. Cell Biol. Lipids 2018, 1863, 772–782. [Google Scholar] [CrossRef] [PubMed]
- Fu, M.; Sun, T.; Bookout, A.L.; Downes, M.; Yu, R.T.; Evans, R.M.; Mangelsdorf, D.J. A Nuclear Receptor Atlas: 3T3-L1 Adipogenesis. Mol. Endocrinol. 2005, 19, 2437–2450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hallakou, S.; Doaré, L.; Foufelle, F.; Kergoat, M.; Guerre-Millo, M.; Berthault, M.F.; Dugail, I.; Morin, J.; Auwerx, J.; Ferré, P. Pioglitazone Induces in Vivo Adipocyte Differentiation in the Obese Zucker Fa/Fa Rat. Diabetes 1997, 46, 1393–1399. [Google Scholar] [CrossRef] [PubMed]
- Pereira, R.I.; Leitner, J.W.; Erickson, C.; Draznin, B. Pioglitazone Acutely Stimulates Adiponectin Secretion from Mouse and Human Adipocytes via Activation of the Phosphatidylinositol 3′-Kinase. Life Sci. 2008, 83, 638–643. [Google Scholar] [CrossRef]
- Ramalho, R.; Almeida, J.; Fernandes, R.; Costa, R.; Pirraco, A.; Guardão, L.; Delgado, L.; Moreira, A.; Soares, R. Neurokinin-1 Receptor, a New Modulator of Lymphangiogenesis in Obese-Asthma Phenotype. Life Sci. 2013, 93, 169–177. [Google Scholar] [CrossRef] [Green Version]
- Luis, C.; Duarte, F.; Faria, I.; Jarak, I.; Oliveira, P.F. Warburg E Ff Ect Inversion: Adiposity Shifts Central Primary Metabolism in MCF-7 Breast Cancer Cells. Life Sci. 2019, 223, 38–46. [Google Scholar] [CrossRef]
- Moreira, Â.; Pereira, S.S.; Costa, M.; Morais, T.; Pinto, A.; Fernandes, R.; Monteiro, M.P. Adipocyte Secreted Factors Enhance Aggressiveness of Prostate Carcinoma Cells. PLoS ONE 2015, 10, e0123217. [Google Scholar] [CrossRef] [Green Version]
- Gacci, M.; De Nunzio, C.; Sebastianelli, A.; Salvi, M.; Vignozzi, L.; Tubaro, A.; Morgia, G.; Serni, S. Meta-Analysis of Metabolic Syndrome and Prostate Cancer. Prostate Cancer Prostatic Dis. 2017, 20, 146–155. [Google Scholar] [CrossRef]
- McGrowder, D.A.; Jackson, L.A.; Crawford, T.V. Prostate Cancer and Metabolic Syndrome: Is There a Link? Asian Pac. J. Cancer Prev. 2012, 13, 1–13. [Google Scholar] [CrossRef]
- Castoldi, A.; De Souza, C.N.; Saraiva Câmara, N.O.; Moraes-Vieira, P.M. The Macrophage Switch in Obesity Development. Front. Immunol. 2016, 6, 637. [Google Scholar] [CrossRef] [Green Version]
- Iwasaki, Y.; Otsuka, H.; Yanagisawa, N.; Hisamitsu, H.; Manabe, A.; Nonaka, N.; Nakamura, M. In Situ Proliferation and Differentiation of Macrophages in Dental Pulp. Cell Tissue Res. 2011, 346, 99–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avgerinos, K.I.; Spyrou, N.; Mantzoros, C.S.; Dalamaga, M. Obesity and Cancer Risk: Emerging Biological Mechanisms and Perspectives. Metabolism 2019, 92, 121–135. [Google Scholar] [CrossRef] [PubMed]
- Tang, P.M.K.; Nikolic-Paterson, D.J.; Lan, H.Y. Macrophages: Versatile Players in Renal Inflammation and Fibrosis. Nat. Rev. Nephrol. 2019, 15, 144–158. [Google Scholar] [CrossRef] [PubMed]
- Marcadenti, A.; de Abreu-Silva, E.O. Different Adipose Tissue Depots: Metabolic Implications and Effects of Surgical Removal. Endocrinol. Nutr. 2015, 62, 458–464. [Google Scholar] [CrossRef] [PubMed]
- Roszer, T. Understanding the Mysterious M2 Macrophage through Activation Markers and Effector Mechanisms. Mediat. Inflamm. 2015, 2015, 16–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Forward | Primer Reverse | Probe |
|---|---|---|---|
| 18S | ACCGCAGCTAGGAATAATGGA | GCCTCAGTTCCGAAAACCA | CY5- ACCGCGGTTCTATTT -BHQ3 |
| PPARα | GCCGAAAGAAGCCCTTACAG | GCCTCAGGGTACCACTAC | JOE-ACATGCGTGAACTCCGT-BHQ1 |
| PPARδ | AGATCCGATCGCACTTCTCA | AGGCGGCAGCCTCAACAT | ROX-AAGGGCTTCTTCCGCC-BHQ2 |
| PPARγ | CCTGCATCTCCACCTTATTATTCTG | CCTTGCATCCTTCACAAGC | FAM-CCTCATGAAGAACCTTCTAACTCCCTCATGGC-BHQ1 |
| Gene | Primer Forward | Primer Reverse |
|---|---|---|
| 18S | ACCGCAGCTAGGAATAATGGA | GCCTCAGTTCCGAAAACCA |
| IL-1β | ACCTAGCTGTCAACGTGTGG | TCAAAGCAATGTGCTGGTGC |
| IL-4 | GCAGCTGATCCGATTCCTGA | TCCAACGTACTCTGGTTGGC |
| IL-6 | TGTGTGAAAGCAGCAAAGAGG | TTTTCACCAGGCAAGTCTCC |
| IL-10 | TGAAAACAAGAGCAAGGCCG | ATAGAGTCGCCACCCTGATG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Teixeira, C.; Sousa, A.P.; Santos, I.; Rocha, A.C.; Alencastre, I.; Pereira, A.C.; Martins-Mendes, D.; Barata, P.; Baylina, P.; Fernandes, R. Enhanced 3T3-L1 Differentiation into Adipocytes by Pioglitazone Pharmacological Activation of Peroxisome Proliferator Activated Receptor-Gamma (PPAR-γ). Biology 2022, 11, 806. https://doi.org/10.3390/biology11060806
Teixeira C, Sousa AP, Santos I, Rocha AC, Alencastre I, Pereira AC, Martins-Mendes D, Barata P, Baylina P, Fernandes R. Enhanced 3T3-L1 Differentiation into Adipocytes by Pioglitazone Pharmacological Activation of Peroxisome Proliferator Activated Receptor-Gamma (PPAR-γ). Biology. 2022; 11(6):806. https://doi.org/10.3390/biology11060806
Chicago/Turabian StyleTeixeira, Catarina, André P. Sousa, Inês Santos, Ana Catarina Rocha, Inês Alencastre, Ana Cláudia Pereira, Daniela Martins-Mendes, Pedro Barata, Pilar Baylina, and Rúben Fernandes. 2022. "Enhanced 3T3-L1 Differentiation into Adipocytes by Pioglitazone Pharmacological Activation of Peroxisome Proliferator Activated Receptor-Gamma (PPAR-γ)" Biology 11, no. 6: 806. https://doi.org/10.3390/biology11060806