
Evaluation of the Cardiometabolic Disorders after Spinal Cord Injury in Mice

 ,
,  ,
,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Care and Procedure for SCI
2.2. Behavioral Assessment
2.3. Body Composition Analysis
2.4. Liver Ultrasound Image Acquisition and Analysis
2.5. Echocardiography Analysis
2.6. Glucose Metabolism
2.7. Triglyceride, Cholesterol, and Insulin Analysis
2.8. Histological Analysis of Heart and Liver Tissues
2.9. Tyrosine Hydroxylase (TH) Immunofluorescence Staining of the Heart and Liver
2.10. Histological Analysis of SC Injury
2.11. Blood Cytokines Measurements
2.12. Statistical Analysis
3. Results
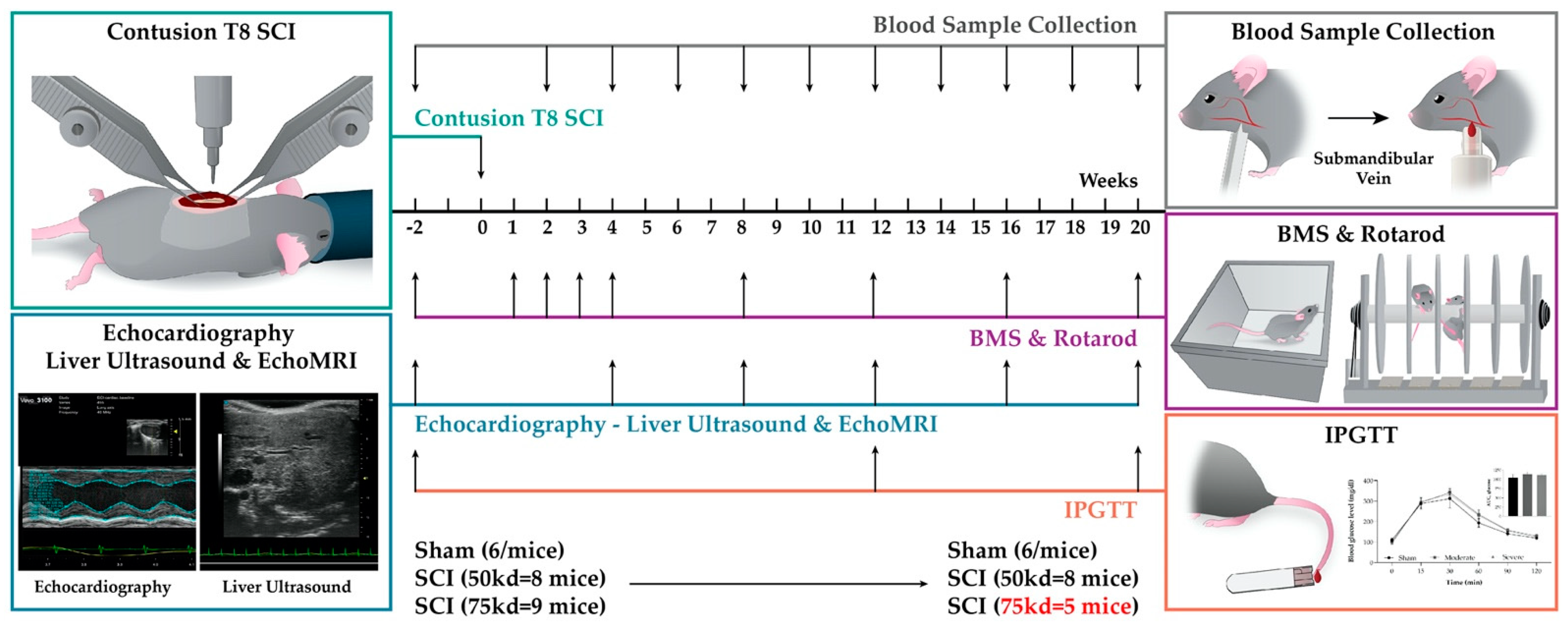
3.1. Experimental Design and Data Collection
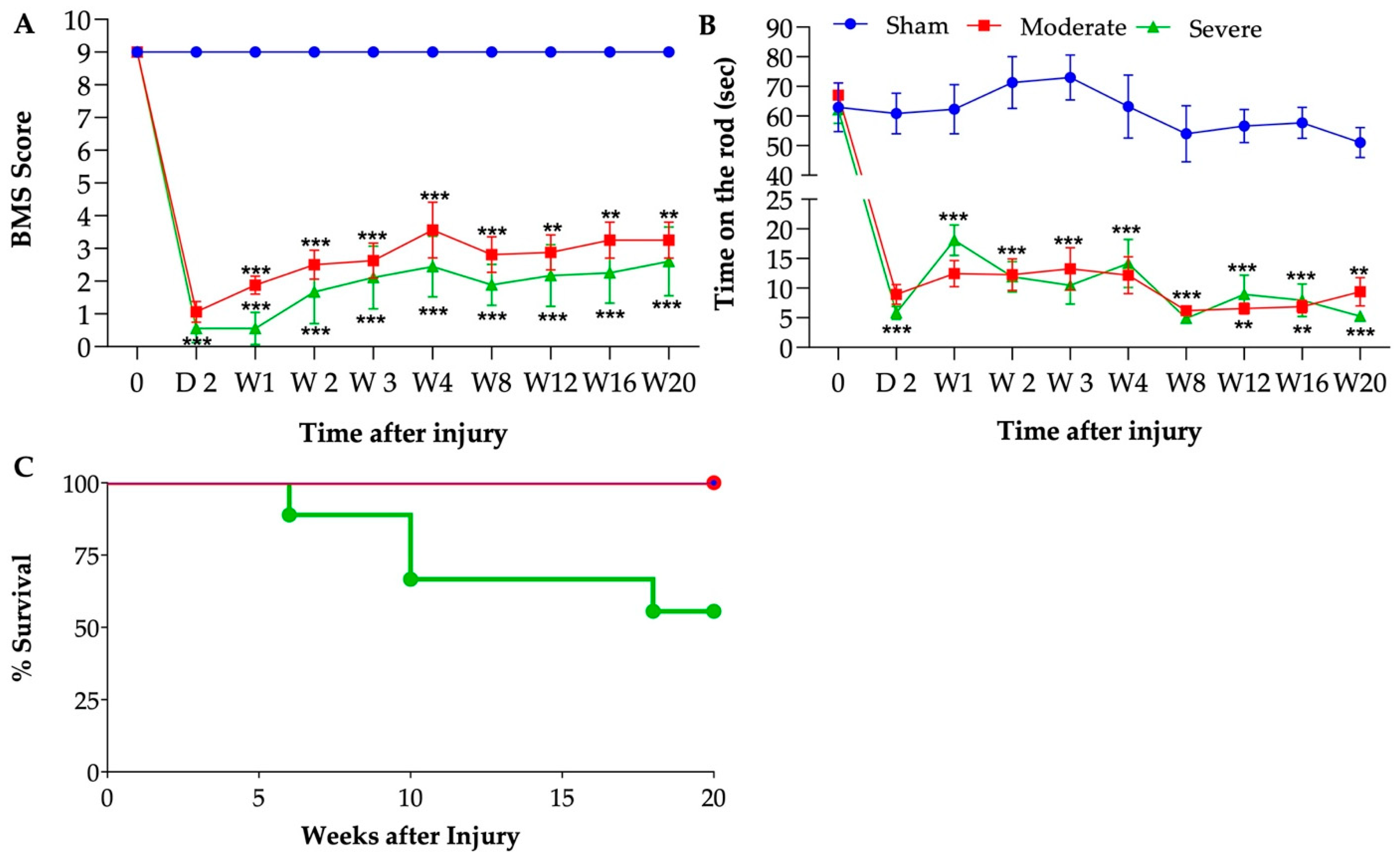
3.2. Sustained Reduction of Motor Function after Chronic SCI
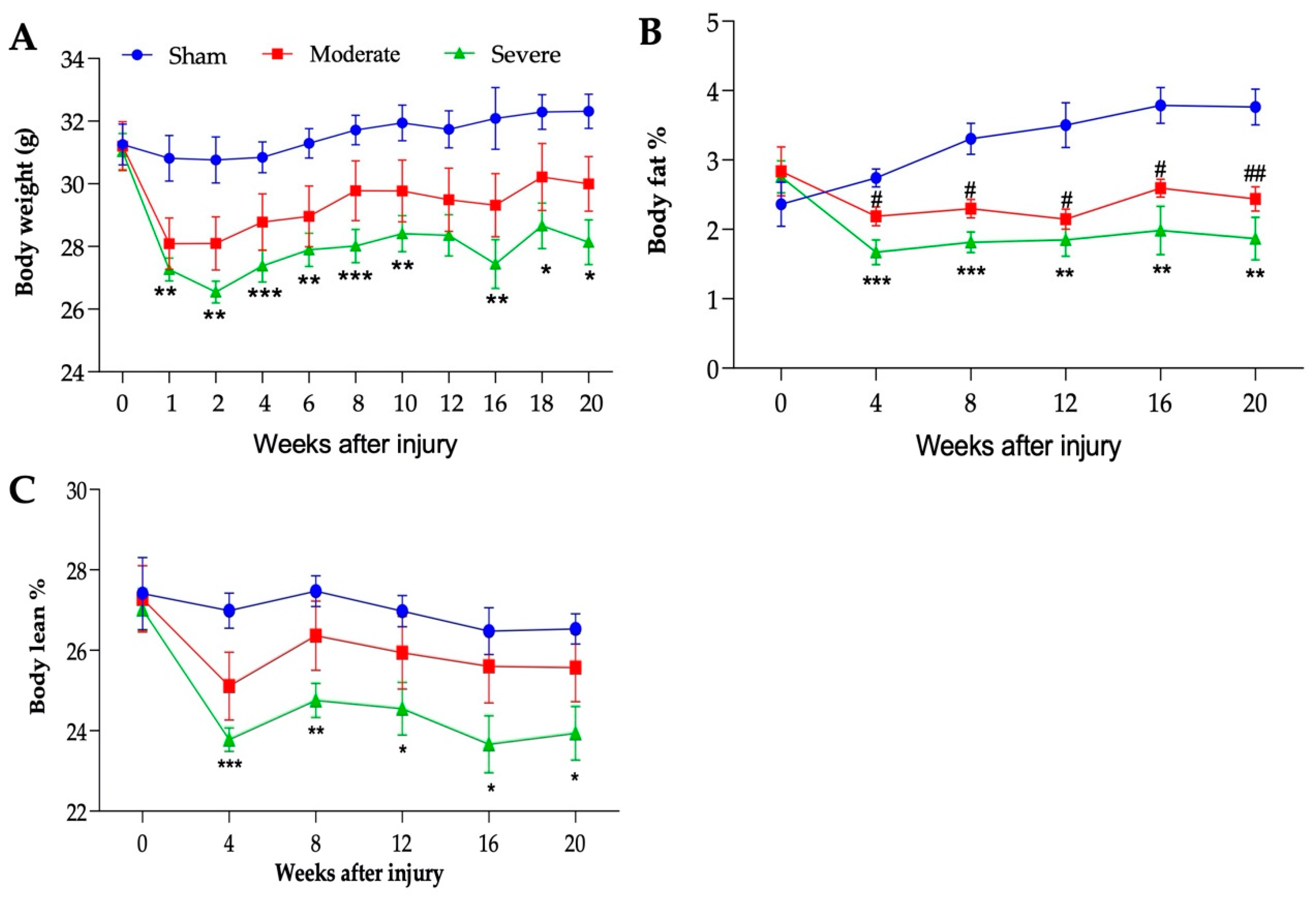
3.3. Severity-Dependent Reduction of Body Weight and Body Composition
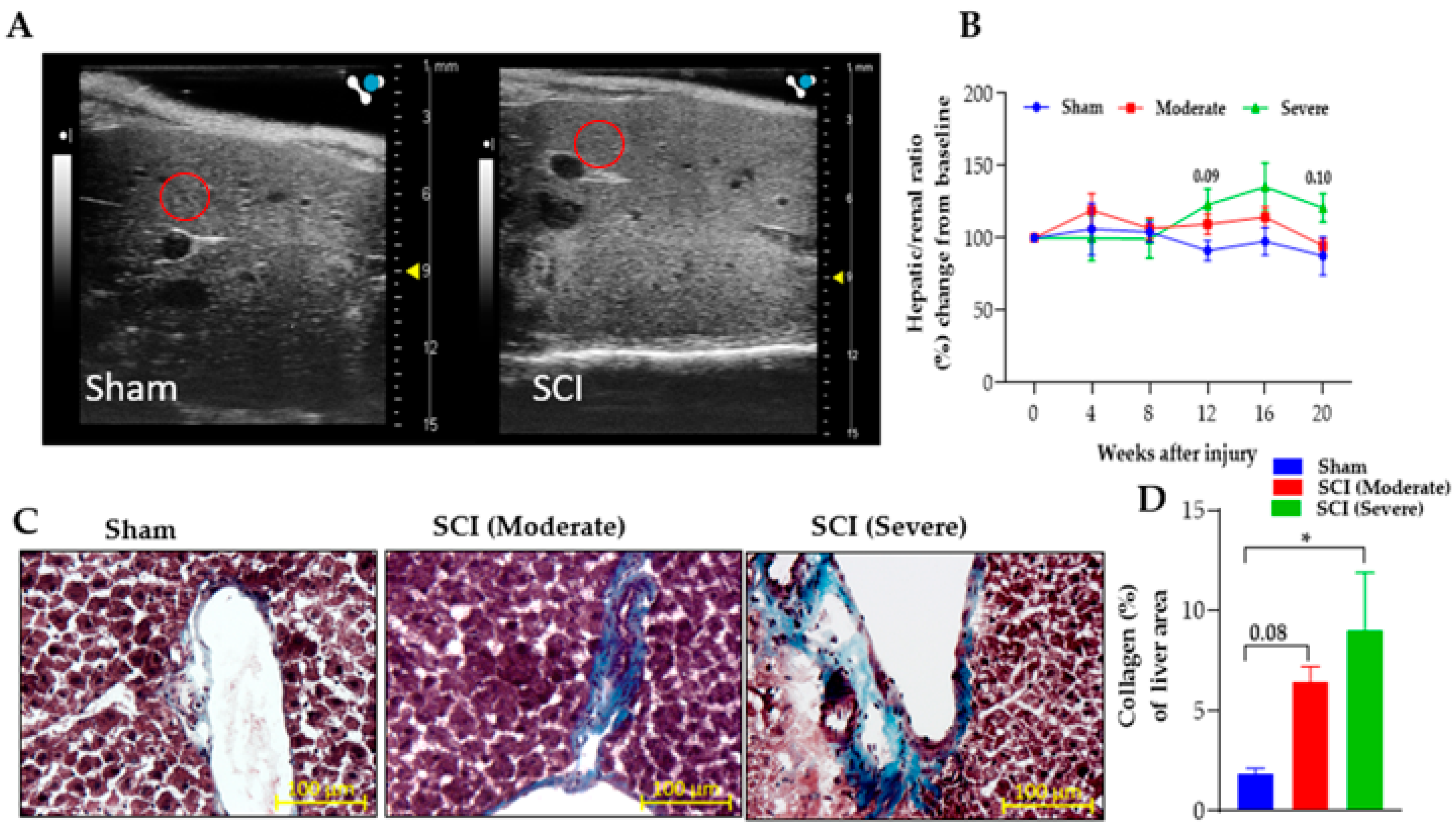
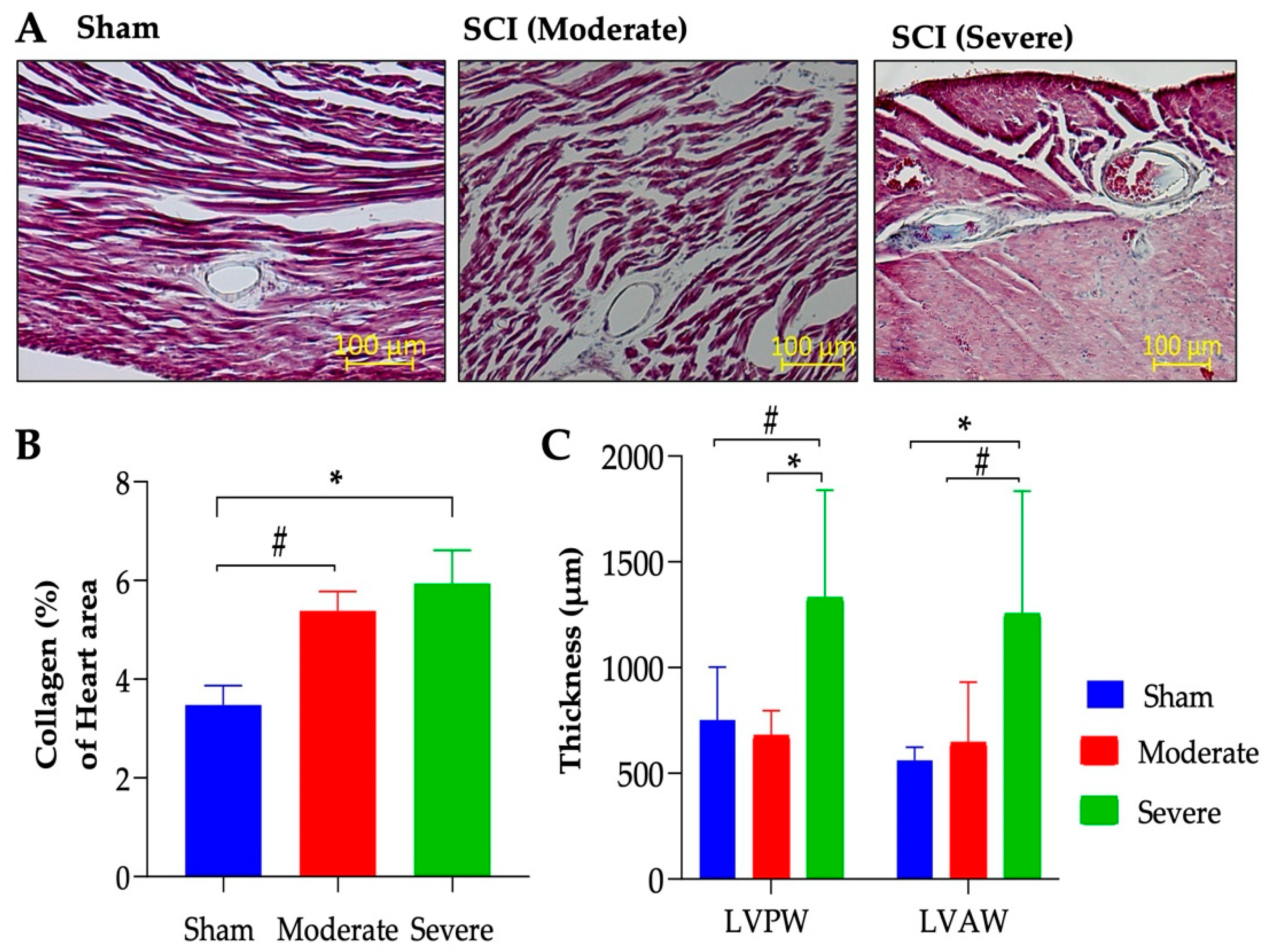
3.4. Severe SCI Induced Stronger Liver Pathology
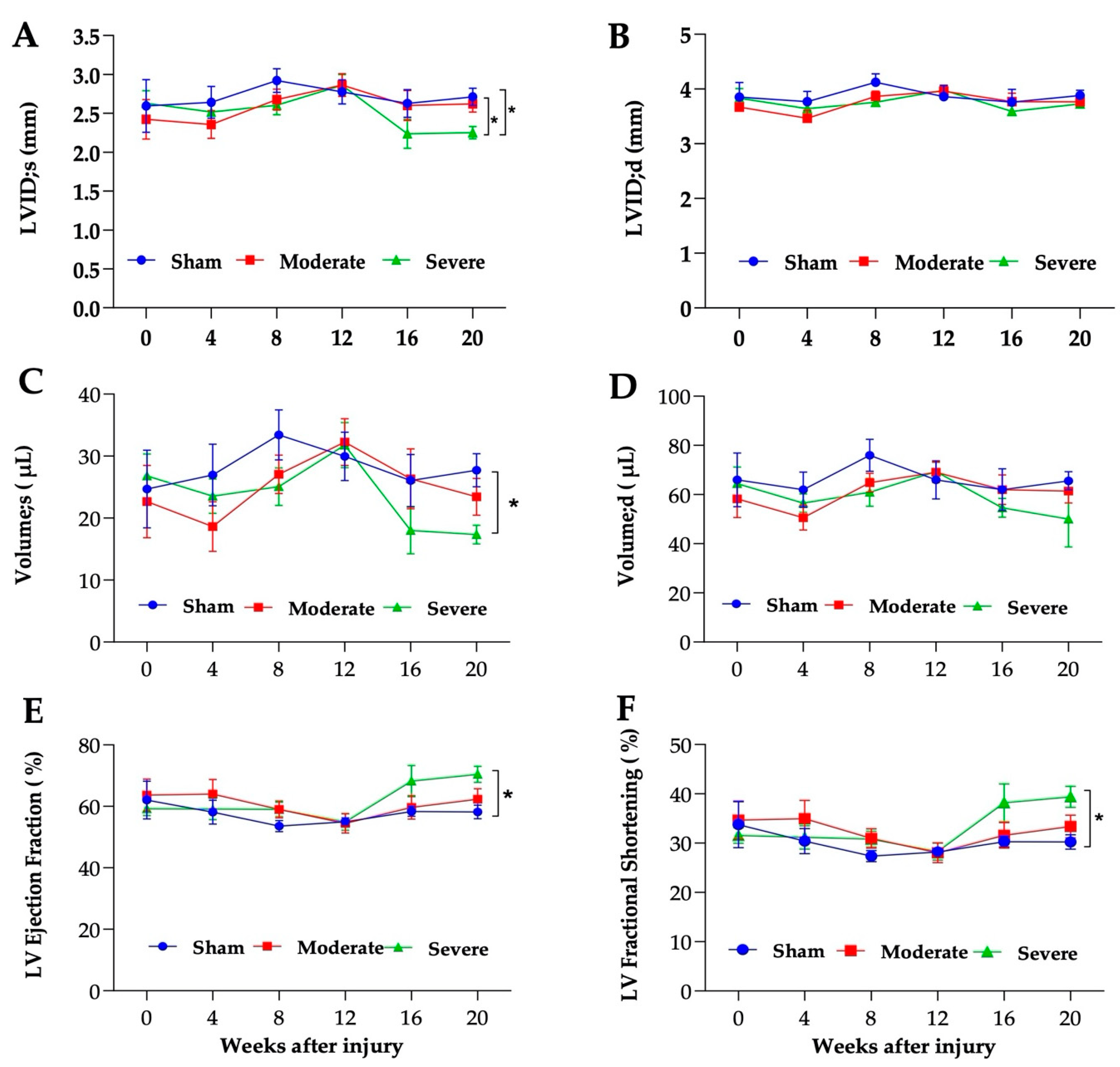
3.5. Severe SCI Induced Cardiac Dysfunction
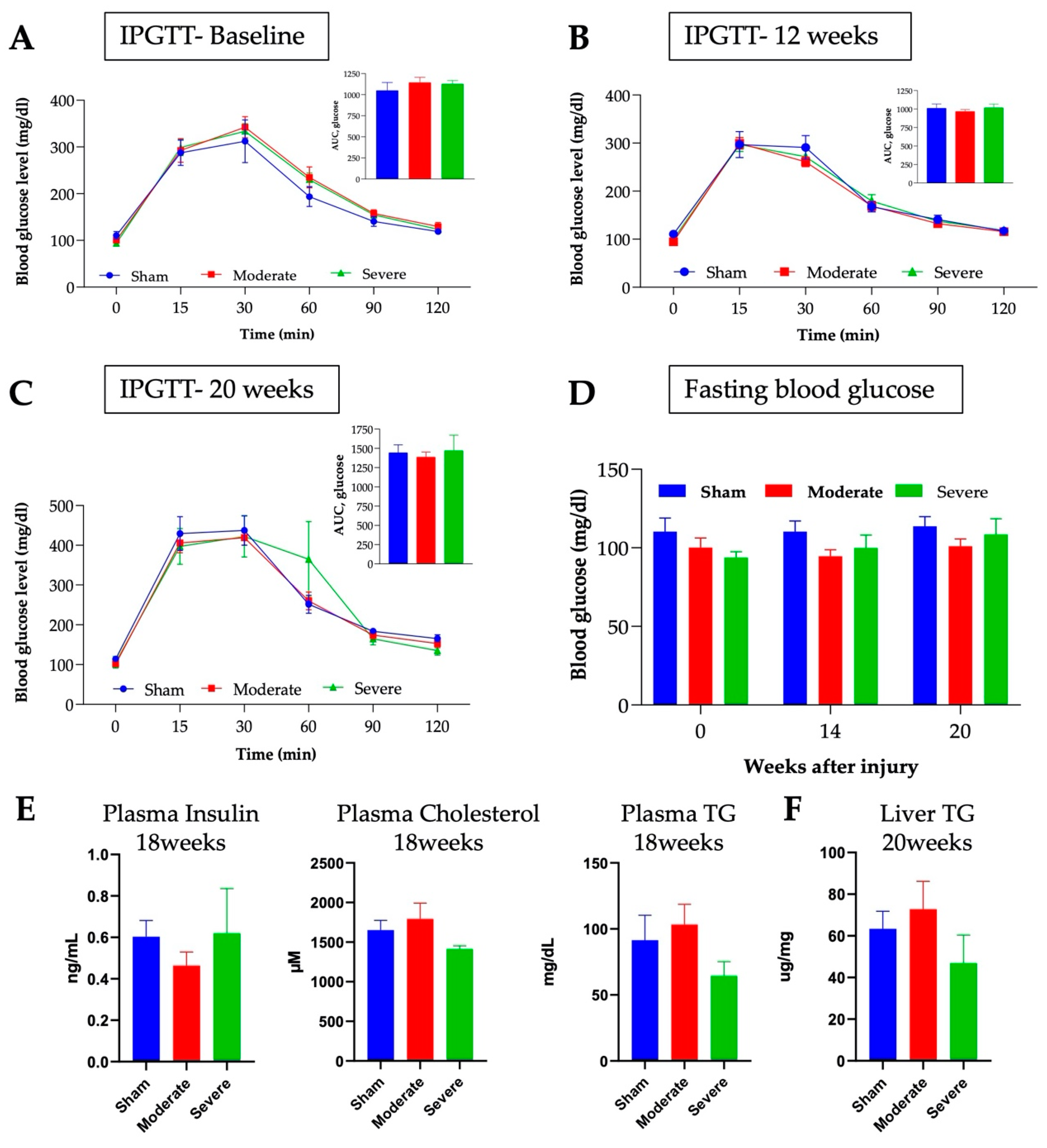
3.6. No Changes in Glucose or Lipids Metabolism after Chronic SCI in Mice
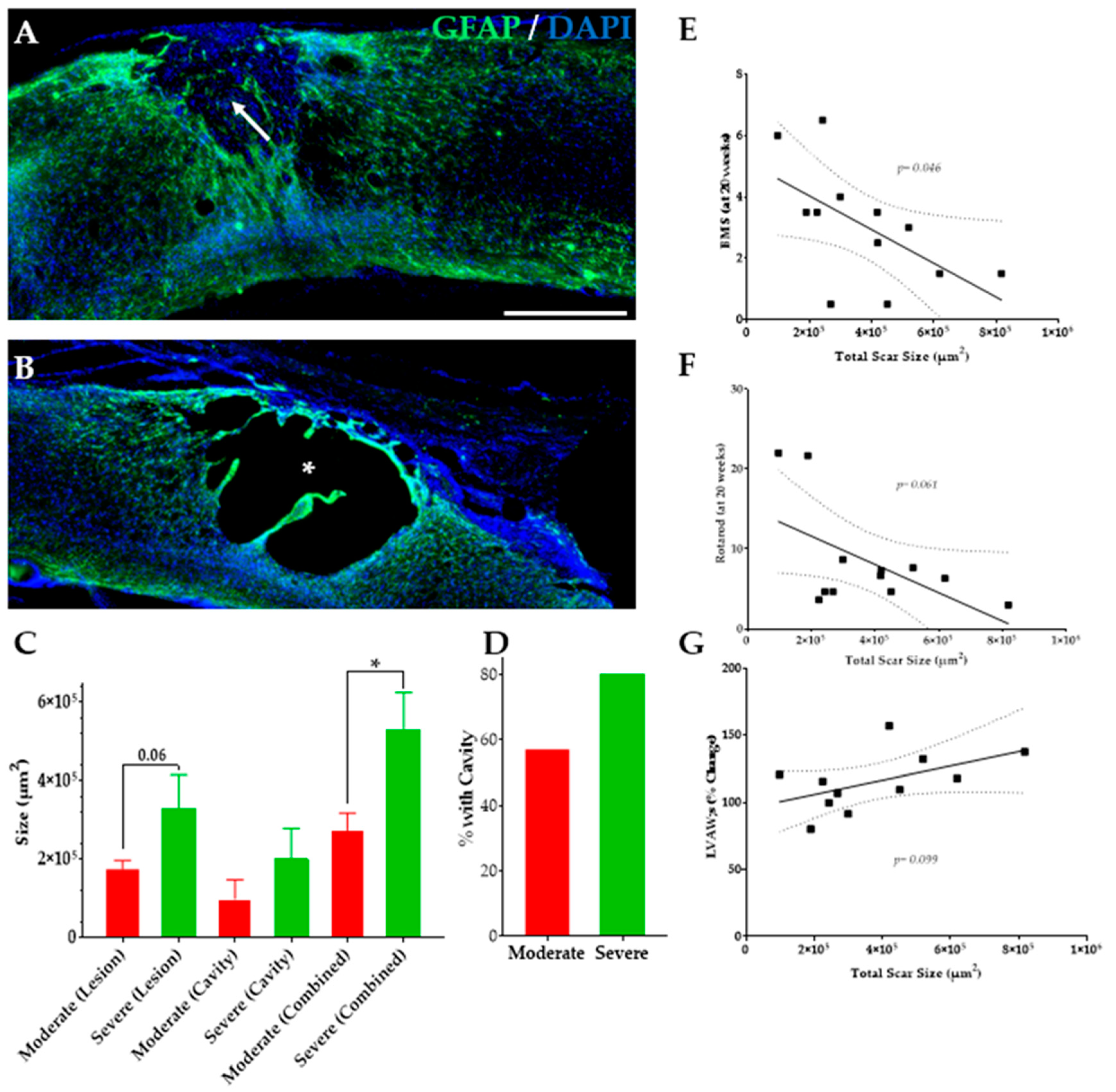
3.7. Severe SCI Induced Changes in SC Injury
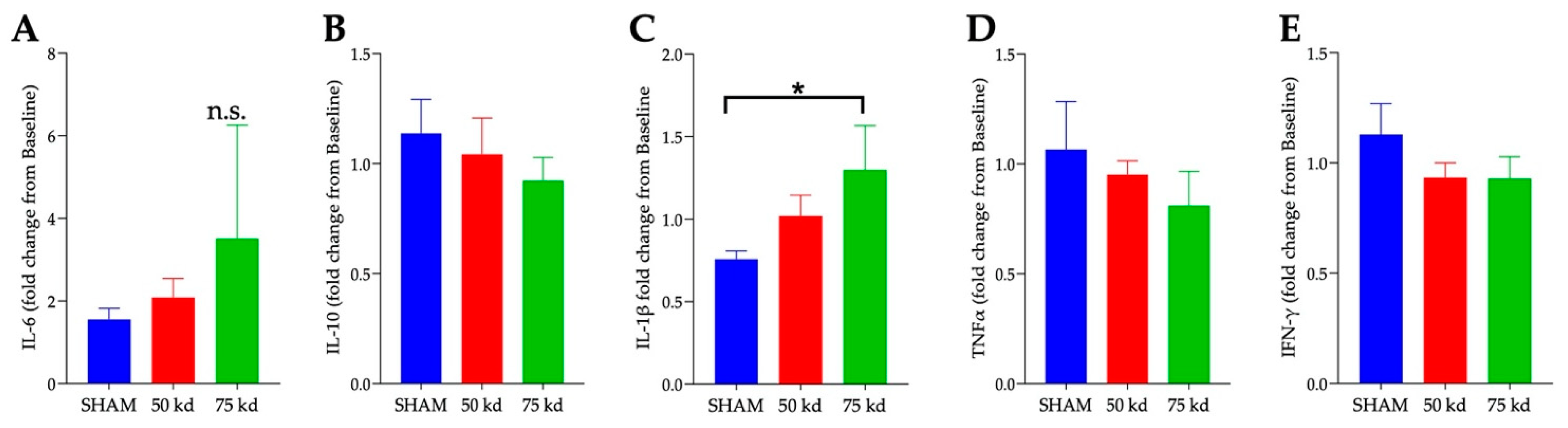
3.8. SCI and Plasma Cytokines
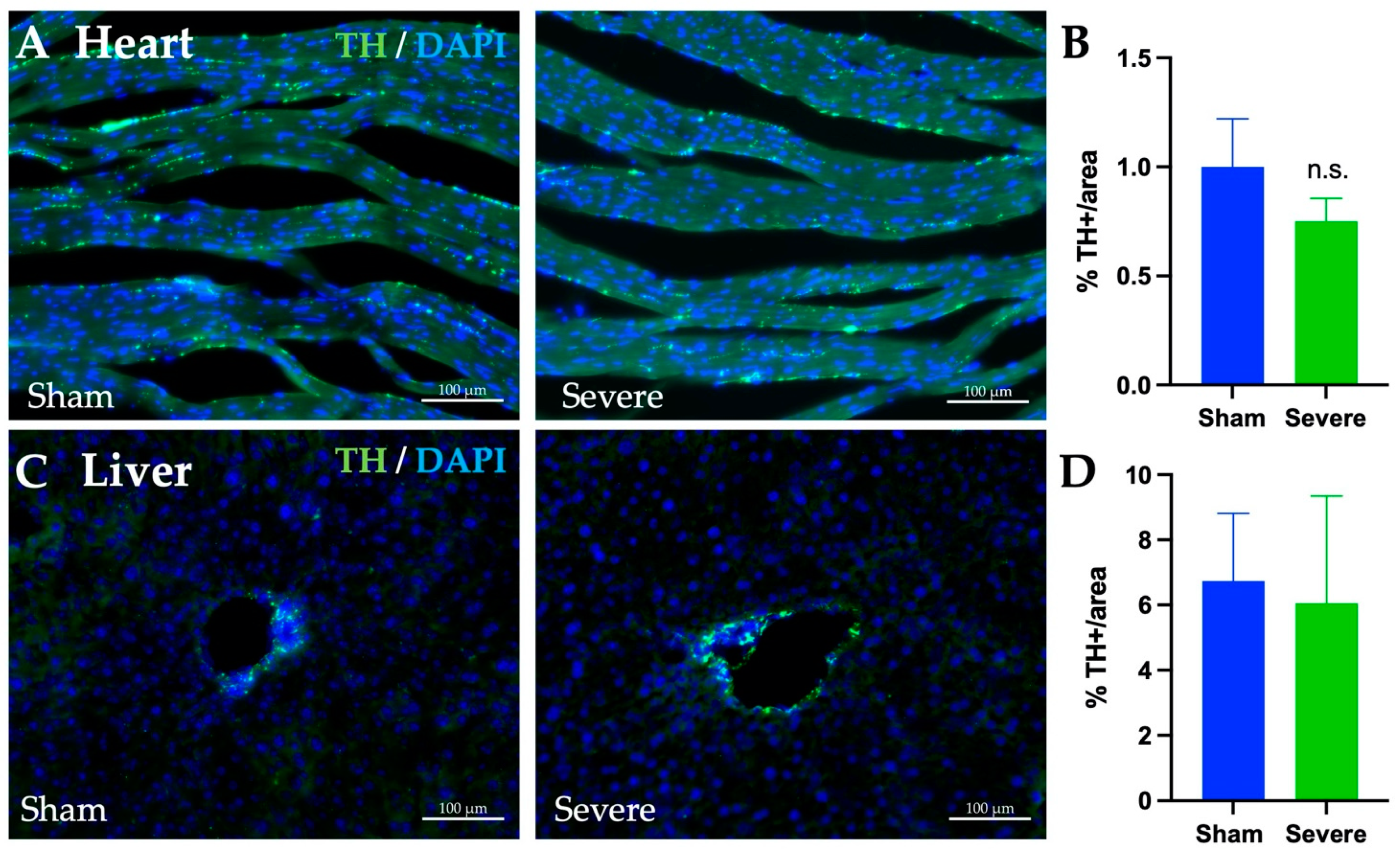
3.9. SCI, Heart, and Liver Innervation
4. Discussion
4.1. SCI Severity and Hepatic Dysfunctions
4.2. SCI Severity and Cardiac Functions
4.3. SCI Severity and Body Composition
4.4. SCI Severity, Motor Function, and Injury Size
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- SCI Facts and Figures. J. Spinal Cord Med. 2017, 40, 872–873. [CrossRef] [PubMed]
- Alizadeh, A.; Dyck, S.M.; Karimi-Abdolrezaee, S. Traumatic Spinal Cord Injury: An Overview of Pathophysiology, Models and Acute Injury Mechanisms. Front. Neurol. 2019, 10, 282. [Google Scholar] [CrossRef] [Green Version]
- Ahuja, C.S.; Wilson, J.R.; Nori, S.; Kotter, M.R.N.; Druschel, C.; Curt, A.; Fehlings, M.G. Traumatic spinal cord injury. Nat. Rev. Dis. Primers 2017, 3, 17018. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Jones, Z.B.; Chen, X.M.; Zhou, L.; So, K.F.; Ren, Y. Multiple organ dysfunction and systemic inflammation after spinal cord injury: A complex relationship. J. Neuroinflamm. 2016, 13, 260. [Google Scholar] [CrossRef] [Green Version]
- Smith, D.L., Jr.; Yarar-Fisher, C. Contributors to Metabolic Disease Risk Following Spinal Cord Injury. Curr. Phys. Med. Rehabil. Rep. 2016, 4, 190–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gill, S.; Sumrell, R.M.; Sima, A.; Cifu, D.X.; Gorgey, A.S. Waist circumference cutoff identifying risks of obesity, metabolic syndrome, and cardiovascular disease in men with spinal cord injury. PLoS ONE 2020, 15, e0236752. [Google Scholar] [CrossRef]
- Gorgey, A.S.; Gater, D.R., Jr. Prevalence of Obesity After Spinal Cord Injury. Top. Spinal Cord Inj. Rehabil. 2007, 12, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Nash, M.S.; Mendez, A.J. A guideline-driven assessment of need for cardiovascular disease risk intervention in persons with chronic paraplegia. Arch. Phys. Med. Rehabil. 2007, 88, 751–757. [Google Scholar] [CrossRef]
- Nash, M.S.; Gater, D.R., Jr. Cardiometabolic Disease and Dysfunction Following Spinal Cord Injury: Origins and Guideline-Based Countermeasures. Phys. Med. Rehabil. Clin. N. Am. 2020, 31, 415–436. [Google Scholar] [CrossRef]
- Cornier, M.A.; Dabelea, D.; Hernandez, T.L.; Lindstrom, R.C.; Steig, A.J.; Stob, N.R.; Van Pelt, R.E.; Wang, H.; Eckel, R.H. The metabolic syndrome. Endocr. Rev. 2008, 29, 777–822. [Google Scholar] [CrossRef]
- Eckel, R.H.; Grundy, S.M.; Zimmet, P.Z. The metabolic syndrome. Lancet 2005, 365, 1415–1428. [Google Scholar] [CrossRef]
- Grundy, S.M.; Hansen, B.; Smith, S.C., Jr.; Cleeman, J.I.; Kahn, R.A.; American Heart, A.; National Heart, L.; Blood, I.; American Diabetes, A. Clinical management of metabolic syndrome: Report of the American Heart Association/National Heart, Lung, and Blood Institute/American Diabetes Association conference on scientific issues related to management. Arterioscler. Thromb. Vasc. Biol. 2004, 24, e19–e24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buchholz, A.C.; Pencharz, P.B. Energy expenditure in chronic spinal cord injury. Curr. Opin. Clin. Nutr. Metab. Care 2004, 7, 635–639. [Google Scholar] [CrossRef]
- Gurka, M.J.; Filipp, S.L.; DeBoer, M.D. Geographical variation in the prevalence of obesity, metabolic syndrome, and diabetes among US adults. Nutr. Diabetes 2018, 8, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stein, C.J.; Colditz, G.A. The epidemic of obesity. J. Clin. Endocrinol. Metab. 2004, 89, 2522–2525. [Google Scholar] [CrossRef] [Green Version]
- Gater, D.R., Jr.; Farkas, G.J.; Berg, A.S.; Castillo, C. Prevalence of metabolic syndrome in veterans with spinal cord injury. J. Spinal Cord Med. 2019, 42, 86–93. [Google Scholar] [CrossRef]
- Shulman, G.I. Ectopic fat in insulin resistance, dyslipidemia, and cardiometabolic disease. N. Engl. J. Med. 2014, 371, 2237–2238. [Google Scholar] [CrossRef]
- Bao, F.; Omana, V.; Brown, A.; Weaver, L.C. The systemic inflammatory response after spinal cord injury in the rat is decreased by alpha4beta1 integrin blockade. J. Neurotrauma 2012, 29, 1626–1637. [Google Scholar] [CrossRef] [Green Version]
- Gorgey, A.S.; Wells, K.M.; Austin, T.L. Adiposity and spinal cord injury. World J. Orthop. 2015, 6, 567–576. [Google Scholar] [CrossRef]
- Bauman, W.A.; Spungen, A.M. Metabolic changes in persons after spinal cord injury. Phys. Med. Rehabil. Clin. N. Am. 2000, 11, 109–140. [Google Scholar] [CrossRef]
- Bauman, W.A.; Spungen, A.M. Carbohydrate and lipid metabolism in chronic spinal cord injury. J. Spinal Cord Med. 2001, 24, 266–277. [Google Scholar] [CrossRef] [PubMed]
- Spungen, A.M.; Adkins, R.H.; Stewart, C.A.; Wang, J.; Pierson, R.N., Jr.; Waters, R.L.; Bauman, W.A. Factors influencing body composition in persons with spinal cord injury: A cross-sectional study. J. Appl. Physiol. (1985) 2003, 95, 2398–2407. [Google Scholar] [CrossRef] [PubMed]
- Shojaei, M.H.; Alavinia, S.M.; Craven, B.C. Management of obesity after spinal cord injury: A systematic review. J. Spinal Cord Med. 2017, 40, 783–794. [Google Scholar] [CrossRef] [PubMed]
- Gater, D.R., Jr. Obesity after spinal cord injury. Phys. Med. Rehabil. Clin. N. Am. 2007, 18, 333–351. [Google Scholar] [CrossRef]
- Castro, M.J.; Apple, D.F., Jr.; Rogers, S.; Dudley, G.A. Influence of complete spinal cord injury on skeletal muscle mechanics within the first 6 months of injury. Eur. J. Appl. Physiol. 2000, 81, 128–131. [Google Scholar] [CrossRef]
- Shah, P.K.; Stevens, J.E.; Gregory, C.M.; Pathare, N.C.; Jayaraman, A.; Bickel, S.C.; Bowden, M.; Behrman, A.L.; Walter, G.A.; Dudley, G.A.; et al. Lower-extremity muscle cross-sectional area after incomplete spinal cord injury. Arch. Phys. Med. Rehabil. 2006, 87, 772–778. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.Y.; Myers, J.; Hayes, A.; Madan, S.; Froelicher, V.F.; Perkash, I.; Kiratli, B.J. C-reactive protein, metabolic syndrome, and insulin resistance in individuals with spinal cord injury. J. Spinal Cord Med. 2005, 28, 20–25. [Google Scholar] [CrossRef]
- Cragg, J.J.; Noonan, V.K.; Krassioukov, A.; Borisoff, J. Cardiovascular disease and spinal cord injury: Results from a national population health survey. Neurology 2013, 81, 723–728. [Google Scholar] [CrossRef] [Green Version]
- Van der Scheer, J.W.; Totosy de Zepetnek, J.O.; Blauwet, C.; Brooke-Wavell, K.; Graham-Paulson, T.; Leonard, A.N.; Webborn, N.; Goosey-Tolfrey, V.L. Assessment of body composition in spinal cord injury: A scoping review. PLoS ONE 2021, 16, e0251142. [Google Scholar] [CrossRef]
- Neto, F.R.; Lopes, G.H. Body composition modifications in people with chronic spinal cord injury after supervised physical activity. J. Spinal Cord Med. 2011, 34, 586–593. [Google Scholar] [CrossRef] [Green Version]
- Felleiter, P.; Krebs, J.; Haeberli, Y.; Schmid, W.; Tesini, S.; Perret, C. Post-traumatic changes in energy expenditure and body composition in patients with acute spinal cord injury. J. Rehabil. Med. 2017, 49, 579–584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cardus, D.; McTaggart, W.G. Body composition in spinal cord injury. Arch. Phys. Med. Rehabil. 1985, 66, 257–259. [Google Scholar] [CrossRef]
- Kocina, P. Body composition of spinal cord injured adults. Sports Med. 1997, 23, 48–60. [Google Scholar] [CrossRef] [PubMed]
- Bechmann, L.P.; Hannivoort, R.A.; Gerken, G.; Hotamisligil, G.S.; Trauner, M.; Canbay, A. The interaction of hepatic lipid and glucose metabolism in liver diseases. J. Hepatol. 2012, 56, 952–964. [Google Scholar] [CrossRef] [Green Version]
- Rui, L. Energy metabolism in the liver. Compr. Physiol. 2014, 4, 177–197. [Google Scholar] [CrossRef] [Green Version]
- Kitade, H.; Chen, G.; Ni, Y.; Ota, T. Nonalcoholic Fatty Liver Disease and Insulin Resistance: New Insights and Potential New Treatments. Nutrients 2017, 9, 387. [Google Scholar] [CrossRef] [Green Version]
- Sipski, M.L.; Estores, I.M.; Alexander, C.J.; Guo, X.; Chandralapaty, S.K. Lack of justification for routine abdominal ultrasonography in patients with chronic spinal cord injury. J. Rehabil. Res. Dev. 2004, 41, 101–108. [Google Scholar] [CrossRef]
- Rankin, K.C.; O’Brien, L.C.; Segal, L.; Khan, M.R.; Gorgey, A.S. Liver Adiposity and Metabolic Profile in Individuals with Chronic Spinal Cord Injury. Biomed. Res. Int. 2017, 2017, 1364818. [Google Scholar] [CrossRef]
- Sauerbeck, A.D.; Laws, J.L.; Bandaru, V.V.; Popovich, P.G.; Haughey, N.J.; McTigue, D.M. Spinal cord injury causes chronic liver pathology in rats. J. Neurotrauma 2015, 32, 159–169. [Google Scholar] [CrossRef] [Green Version]
- Campbell, S.J.; Zahid, I.; Losey, P.; Law, S.; Jiang, Y.; Bilgen, M.; van Rooijen, N.; Morsali, D.; Davis, A.E.; Anthony, D.C. Liver Kupffer cells control the magnitude of the inflammatory response in the injured brain and spinal cord. Neuropharmacology 2008, 55, 780–787. [Google Scholar] [CrossRef]
- Hundt, H.; Fleming, J.C.; Phillips, J.T.; Lawendy, A.; Gurr, K.R.; Bailey, S.I.; Sanders, D.; Bihari, R.; Gray, D.; Parry, N.; et al. Assessment of hepatic inflammation after spinal cord injury using intravital microscopy. Injury 2011, 42, 691–696. [Google Scholar] [CrossRef] [PubMed]
- Garshick, E.; Kelley, A.; Cohen, S.A.; Garrison, A.; Tun, C.G.; Gagnon, D.; Brown, R. A prospective assessment of mortality in chronic spinal cord injury. Spinal Cord 2005, 43, 408–416. [Google Scholar] [CrossRef] [Green Version]
- Claydon, V.E.; Krassioukov, A.V. Orthostatic hypotension and autonomic pathways after spinal cord injury. J. Neurotrauma 2006, 23, 1713–1725. [Google Scholar] [CrossRef] [PubMed]
- Ravensbergen, H.J.; de Groot, S.; Post, M.W.; Slootman, H.J.; van der Woude, L.H.; Claydon, V.E. Cardiovascular function after spinal cord injury: Prevalence and progression of dysfunction during inpatient rehabilitation and 5 years following discharge. Neurorehabil. Neural. Repair. 2014, 28, 219–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michael, F.M.; Patel, S.P.; Rabchevsky, A.G. Intraspinal Plasticity Associated With the Development of Autonomic Dysreflexia After Complete Spinal Cord Injury. Front. Cell. Neurosci. 2019, 13, 505. [Google Scholar] [CrossRef]
- Hou, S.; Rabchevsky, A.G. Autonomic consequences of spinal cord injury. Compr. Physiol. 2014, 4, 1419–1453. [Google Scholar] [CrossRef]
- West, C.R.; Mills, P.; Krassioukov, A.V. Influence of the neurological level of spinal cord injury on cardiovascular outcomes in humans: A meta-analysis. Spinal Cord 2012, 50, 484–492. [Google Scholar] [CrossRef] [Green Version]
- West, C.R.; Poormasjedi-Meibod, M.S.; Manouchehri, N.; Williams, A.M.; Erskine, E.L.; Webster, M.; Fisk, S.; Morrison, C.; Short, K.; So, K.; et al. A porcine model for studying the cardiovascular consequences of high-thoracic spinal cord injury. J. Physiol. 2020, 598, 929–942. [Google Scholar] [CrossRef]
- Lujan, H.L.; Janbaih, H.; DiCarlo, S.E. Dynamic interaction between the heart and its sympathetic innervation following T5 spinal cord transection. J. Appl. Physiol. (1985) 2012, 113, 1332–1341. [Google Scholar] [CrossRef] [Green Version]
- Lujan, H.L.; Janbaih, H.; DiCarlo, S.E. Structural remodeling of the heart and its premotor cardioinhibitory vagal neurons following T(5) spinal cord transection. J. Appl. Physiol. (1985) 2014, 116, 1148–1155. [Google Scholar] [CrossRef] [Green Version]
- Squair, J.W.; Liu, J.; Tetzlaff, W.; Krassioukov, A.V.; West, C.R. Spinal cord injury-induced cardiomyocyte atrophy and impaired cardiac function are severity dependent. Exp. Physiol. 2018, 103, 179–189. [Google Scholar] [CrossRef] [PubMed]
- Squair, J.W.; West, C.R.; Popok, D.; Assinck, P.; Liu, J.; Tetzlaff, W.; Krassioukov, A.V. High Thoracic Contusion Model for the Investigation of Cardiovascular Function after Spinal Cord Injury. J. Neurotrauma 2017, 34, 671–684. [Google Scholar] [CrossRef] [PubMed]
- Bhalala, O.G.; Pan, L.; North, H.; McGuire, T.; Kessler, J.A. Generation of Mouse Spinal Cord Injury. Bio Protoc. 2013, 3, e886. [Google Scholar] [CrossRef] [PubMed]
- Gruner, J.A. A monitored contusion model of spinal cord injury in the rat. J. Neurotrauma 1992, 9, 123–126, discussion 126–128. [Google Scholar] [CrossRef]
- Lee, J.K.; Geoffroy, C.G.; Chan, A.F.; Tolentino, K.E.; Crawford, M.J.; Leal, M.A.; Kang, B.; Zheng, B. Assessing spinal axon regeneration and sprouting in Nogo-, MAG-, and OMgp-deficient mice. Neuron 2010, 66, 663–670. [Google Scholar] [CrossRef] [Green Version]
- Basso, D.M.; Fisher, L.C.; Anderson, A.J.; Jakeman, L.B.; McTigue, D.M.; Popovich, P.G. Basso Mouse Scale for locomotion detects differences in recovery after spinal cord injury in five common mouse strains. J. Neurotrauma 2006, 23, 635–659. [Google Scholar] [CrossRef]
- Nixon, J.P.; Zhang, M.; Wang, C.; Kuskowski, M.A.; Novak, C.M.; Levine, J.A.; Billington, C.J.; Kotz, C.M. Evaluation of a quantitative magnetic resonance imaging system for whole body composition analysis in rodents. Obesity 2010, 18, 1652–1659. [Google Scholar] [CrossRef] [Green Version]
- Pandit, H.; Tinney, J.P.; Li, Y.; Cui, G.; Li, S.; Keller, B.B.; Martin, R.C.G., 2nd. Utilizing Contrast-Enhanced Ultrasound Imaging for Evaluating Fatty Liver Disease Progression in Pre-clinical Mouse Models. Ultrasound Med. Biol. 2019, 45, 549–557. [Google Scholar] [CrossRef]
- Ghnenis, A.B.; Burns, D.T.; Osimanjiang, W.; He, G.; Bushman, J.S. A Long-Term Pilot Study on Sex and Spinal Cord Injury Shows Sexual Dimorphism in Functional Recovery and Cardio-Metabolic Responses. Sci. Rep. 2020, 10, 2762. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Q.; Yan, T.; Li, L.; Chopp, M.; Venkat, P.; Qian, Y.; Li, R.; Wu, R.; Li, W.; Lu, M.; et al. Immune Response Mediates Cardiac Dysfunction after Traumatic Brain Injury. J. Neurotrauma 2019, 36, 619–629. [Google Scholar] [CrossRef]
- Wu, C.; Yan, F.; Li, M.; Tu, Y.; Guo, Z.; Chen, Y.; Wu, Y.; Li, Q.; Yu, C.; Fu, Y.; et al. Whole-Mount Kidney Clearing and Visualization Reveal Renal Sympathetic Hyperinnervation in Heart Failure Mice. Front. Physiol. 2021, 12, 696286. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Rigel, D.F. Echocardiographic examination in rats and mice. Methods Mol. Biol. 2009, 573, 139–155. [Google Scholar] [CrossRef] [PubMed]
- Tong, C.W.; Stelzer, J.E.; Greaser, M.L.; Powers, P.A.; Moss, R.L. Acceleration of crossbridge kinetics by protein kinase A phosphorylation of cardiac myosin binding protein C modulates cardiac function. Circ. Res. 2008, 103, 974–982. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, A.; Barajas, S.; Lammoglia, G.M.; Reyna, A.J.; Morley, T.S.; Johnson, J.A.; Scherer, P.E.; Rutkowski, J.M. Vascular Endothelial Growth Factor-D (VEGF-D) Overexpression and Lymphatic Expansion in Murine Adipose Tissue Improves Metabolism in Obesity. Am. J. Pathol. 2019, 189, 924–939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, D.; Patel, V.V.; Ricciotti, E.; Zhou, R.; Levin, M.D.; Gao, E.; Yu, Z.; Ferrari, V.A.; Lu, M.M.; Xu, J.; et al. Cardiomyocyte cyclooxygenase-2 influences cardiac rhythm and function. Proc. Natl. Acad. Sci. USA 2009, 106, 7548–7552. [Google Scholar] [CrossRef] [Green Version]
- Jackson, S.J.; Andrews, N.; Ball, D.; Bellantuono, I.; Gray, J.; Hachoumi, L.; Holmes, A.; Latcham, J.; Petrie, A.; Potter, P.; et al. Does age matter? The impact of rodent age on study outcomes. Lab. Anim. 2017, 51, 160–169. [Google Scholar] [CrossRef] [Green Version]
- Goodus, M.T.; Carson, K.E.; Sauerbeck, A.D.; Dey, P.; Alfredo, A.N.; Popovich, P.G.; Bruno, R.S.; McTigue, D.M. Liver inflammation at the time of spinal cord injury enhances intraspinal pathology, liver injury, metabolic syndrome and locomotor deficits. Exp. Neurol. 2021, 342, 113725. [Google Scholar] [CrossRef]
- Mathiesen, U.L.; Franzen, L.E.; Aselius, H.; Resjo, M.; Jacobsson, L.; Foberg, U.; Fryden, A.; Bodemar, G. Increased liver echogenicity at ultrasound examination reflects degree of steatosis but not of fibrosis in asymptomatic patients with mild/moderate abnormalities of liver transaminases. Dig. Liver. Dis. 2002, 34, 516–522. [Google Scholar] [CrossRef]
- Raymond, J.; Harmer, A.R.; Temesi, J.; van Kemenade, C. Glucose tolerance and physical activity level in people with spinal cord injury. Spinal Cord 2010, 48, 591–596. [Google Scholar] [CrossRef] [Green Version]
- Gorgey, A.S.; Dolbow, D.R.; Dolbow, J.D.; Khalil, R.K.; Castillo, C.; Gater, D.R. Effects of spinal cord injury on body composition and metabolic profile—Part I. J. Spinal Cord Med. 2014, 37, 693–702. [Google Scholar] [CrossRef] [Green Version]
- Graham, Z.A.; Liu, X.H.; Harlow, L.; Pan, J.; Azulai, D.; Tawfeek, H.A.; Wnek, R.D.; Mattingly, A.J.; Bauman, W.A.; Yarrow, J.F.; et al. Effects of a High-Fat Diet on Tissue Mass, Bone, and Glucose Tolerance after Chronic Complete Spinal Cord Transection in Male Mice. Neurotrauma Rep. 2020, 1, 17–31. [Google Scholar] [CrossRef] [PubMed]
- Primeaux, S.D.; Tong, M.; Holmes, G.M. Effects of chronic spinal cord injury on body weight and body composition in rats fed a standard chow diet. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2007, 293, R1102–R1109. [Google Scholar] [CrossRef] [Green Version]
- Squair, J.W.; West, C.R.; Krassioukov, A.V. Neuroprotection, Plasticity Manipulation, and Regenerative Strategies to Improve Cardiovascular Function following Spinal Cord Injury. J. Neurotrauma 2015, 32, 609–621. [Google Scholar] [CrossRef] [PubMed]
- Hagen, E.M.; Rekand, T.; Gronning, M.; Faerestrand, S. Cardiovascular complications of spinal cord injury. Tidsskr. Nor. Laegeforen 2012, 132, 1115–1120. [Google Scholar] [CrossRef] [PubMed]
- Williams, A.M.; Gee, C.M.; Voss, C.; West, C.R. Cardiac consequences of spinal cord injury: Systematic review and meta-analysis. Heart 2019, 105, 217–225. [Google Scholar] [CrossRef] [PubMed]
- West, C.R.; Squair, J.W.; McCracken, L.; Currie, K.D.; Somvanshi, R.; Yuen, V.; Phillips, A.A.; Kumar, U.; McNeill, J.H.; Krassioukov, A.V. Cardiac Consequences of Autonomic Dysreflexia in Spinal Cord Injury. Hypertension 2016, 68, 1281–1289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- West, C.R.; Crawford, M.A.; Poormasjedi-Meibod, M.S.; Currie, K.D.; Fallavollita, A.; Yuen, V.; McNeill, J.H.; Krassioukov, A.V. Passive hind-limb cycling improves cardiac function and reduces cardiovascular disease risk in experimental spinal cord injury. J. Physiol. 2014, 592, 1771–1783. [Google Scholar] [CrossRef]
- Lindsey, M.L.; Kassiri, Z.; Virag, J.A.I.; de Castro Bras, L.E.; Scherrer-Crosbie, M. Guidelines for measuring cardiac physiology in mice. Am. J. Physiol. Heart Circ. Physiol. 2018, 314, H733–H752. [Google Scholar] [CrossRef] [Green Version]
- Kai, H.; Kuwahara, F.; Tokuda, K.; Imaizumi, T. Diastolic dysfunction in hypertensive hearts: Roles of perivascular inflammation and reactive myocardial fibrosis. Hypertens Res. 2005, 28, 483–490. [Google Scholar] [CrossRef] [Green Version]
- Lijnen, P.; Petrov, V. Induction of cardiac fibrosis by aldosterone. J. Mol. Cell. Cardiol. 2000, 32, 865–879. [Google Scholar] [CrossRef]
- Essick, E.E.; Sam, F. Cardiac hypertrophy and fibrosis in the metabolic syndrome: A role for aldosterone and the mineralocorticoid receptor. Int. J. Hypertens 2011, 2011, 346985. [Google Scholar] [CrossRef] [Green Version]
- Kong, P.; Christia, P.; Frangogiannis, N.G. The pathogenesis of cardiac fibrosis. Cell. Mol. Life Sci. 2014, 71, 549–574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diez, J. Mechanisms of cardiac fibrosis in hypertension. J. Clin. Hypertens (Greenwich) 2007, 9, 546–550. [Google Scholar] [CrossRef] [PubMed]
- Ho, C.Y.; Lopez, B.; Coelho-Filho, O.R.; Lakdawala, N.K.; Cirino, A.L.; Jarolim, P.; Kwong, R.; Gonzalez, A.; Colan, S.D.; Seidman, J.G.; et al. Myocardial fibrosis as an early manifestation of hypertrophic cardiomyopathy. N. Engl. J. Med. 2010, 363, 552–563. [Google Scholar] [CrossRef] [Green Version]
- Naser, J.A.; Anupraiwan, O.; Adigun, R.O.; Maleszewski, J.J.; Pislaru, S.V.; Pellikka, P.A.; Pislaru, C. Myocardial Stiffness by Cardiac Elastography in Hypertrophic Cardiomyopathy: Relationship With Myocardial Fibrosis and Clinical Outcomes. JACC Cardiovasc. Imaging 2021, 14, 2051–2053. [Google Scholar] [CrossRef] [PubMed]
- Basso, C.; Thiene, G.; Corrado, D.; Buja, G.; Melacini, P.; Nava, A. Hypertrophic cardiomyopathy and sudden death in the young: Pathologic evidence of myocardial ischemia. Hum. Pathol. 2000, 31, 988–998. [Google Scholar] [CrossRef] [PubMed]
- Frieler, R.A.; Mortensen, R.M. Immune cell and other noncardiomyocyte regulation of cardiac hypertrophy and remodeling. Circulation 2015, 131, 1019–1030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mann, D.L. Innate immunity and the failing heart: The cytokine hypothesis revisited. Circ. Res. 2015, 116, 1254–1268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wenzl, F.A.; Ambrosini, S.; Mohammed, S.A.; Kraler, S.; Luscher, T.F.; Costantino, S.; Paneni, F. Inflammation in Metabolic Cardiomyopathy. Front. Cardiovasc. Med. 2021, 8, 742178. [Google Scholar] [CrossRef]
- Koopman, F.A.; van Maanen, M.A.; Vervoordeldonk, M.J.; Tak, P.P. Balancing the autonomic nervous system to reduce inflammation in rheumatoid arthritis. J. Intern. Med. 2017, 282, 64–75. [Google Scholar] [CrossRef] [Green Version]
- Crane, D.A.; Little, J.W.; Burns, S.P. Weight gain following spinal cord injury: A pilot study. J. Spinal Cord Med. 2011, 34, 227–232. [Google Scholar] [CrossRef] [Green Version]
- Castro, M.J.; Apple, D.F., Jr.; Hillegass, E.A.; Dudley, G.A. Influence of complete spinal cord injury on skeletal muscle cross-sectional area within the first 6 months of injury. Eur. J. Appl. Physiol. Occup. Physiol. 1999, 80, 373–378. [Google Scholar] [CrossRef] [PubMed]
- Harris, K.K.; Himel, A.R.; Duncan, B.C.; Grill, R.J.; Grayson, B.E. Energy balance following diets of varying fat content: Metabolic dysregulation in a rodent model of spinal cord contusion. Physiol. Rep. 2019, 7, e14207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goodus, M.T.; McTigue, D.M. Hepatic dysfunction after spinal cord injury: A vicious cycle of central and peripheral pathology? Exp. Neurol. 2020, 325, 113160. [Google Scholar] [CrossRef] [PubMed]
- Laclaustra, M.; Van Den Berg, E.L.; Hurtado-Roca, Y.; Castellote, J.M. Serum lipid profile in subjects with traumatic spinal cord injury. PLoS ONE 2015, 10, e0115522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raguindin, P.F.; Frankl, G.; Itodo, O.A.; Bertolo, A.; Zeh, R.M.; Capossela, S.; Minder, B.; Stoyanov, J.; Stucki, G.; Franco, O.H.; et al. The neurological level of spinal cord injury and cardiovascular risk factors: A systematic review and meta-analysis. Spinal Cord 2021, 59, 1135–1145. [Google Scholar] [CrossRef] [PubMed]
- McFarlane, K.; Otto, T.E.; Bailey, W.M.; Veldhorst, A.K.; Donahue, R.R.; Taylor, B.K.; Gensel, J.C. Effect of Sex on Motor Function, Lesion Size, and Neuropathic Pain after Contusion Spinal Cord Injury in Mice. J. Neurotrauma 2020, 37, 1983–1990. [Google Scholar] [CrossRef]
- Kakuta, Y.; Adachi, A.; Yokohama, M.; Horii, T.; Mieda, T.; Iizuka, Y.; Takagishi, K.; Chikuda, H.; Iizuka, H.; Nakamura, K. Spontaneous functional full recovery from motor and sensory deficits in adult mice after mild spinal cord injury. Heliyon 2019, 5, e01847. [Google Scholar] [CrossRef] [Green Version]
- Fouad, K.; Hurd, C.; Magnuson, D.S. Functional testing in animal models of spinal cord injury: Not as straight forward as one would think. Front. Integr. Neurosci. 2013, 7, 85. [Google Scholar] [CrossRef] [Green Version]
- Phillips, A.A.; Krassioukov, A.V. Contemporary Cardiovascular Concerns after Spinal Cord Injury: Mechanisms, Maladaptations, and Management. J. Neurotrauma 2015, 32, 1927–1942. [Google Scholar] [CrossRef]
- Rose-John, S. Interleukin-6 Family Cytokines. Cold Spring Harb. Perspect. Biol. 2018, 10, a028415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wieckowska, A.; Papouchado, B.G.; Li, Z.; Lopez, R.; Zein, N.N.; Feldstein, A.E. Increased hepatic and circulating interleukin-6 levels in human nonalcoholic steatohepatitis. Am. J. Gastroenterol. 2008, 103, 1372–1379. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number of Mice | Set of Experiments | |||||||
|---|---|---|---|---|---|---|---|---|
| Mice Group | Sham | 50 kd | 75 kd | BMS & Rotarod | Echocardiography & Liver Ultrasound | EchoMRI | Blood Samples | IPGTT |
| Pre SCI | 6 | 8 | 9 | + | + | + | + | + |
| W1 | 6 | 8 | 9 | + | - | - | - | - |
| W2 | 6 | 8 | 9 | + | - | - | + | - |
| W3 | 6 | 8 | 9 | + | - | - | - | - |
| W4 | 6 | 8 | 9 | + | + | + | + | + |
| W5 | 6 | 8 | 9 | - | - | - | - | - |
| W6 | 6 | 8 | 9 | - | - | - | + | - |
| W7 | 6 | 8 | 9 | - | - | - | - | - |
| W8 | 6 | 8 | 8 | + | + | + | + | + |
| W9 | 6 | 8 | 8 | - | - | - | - | - |
| W10 | 6 | 8 | 6 | - | - | - | + | - |
| W11 | 6 | 8 | 6 | - | - | - | - | - |
| W12 | 6 | 8 | 6 | + | + | + | + | + |
| W13 | 6 | 8 | 6 | - | - | - | - | - |
| W14 | 6 | 8 | 6 | - | - | - | + | - |
| W15 | 6 | 8 | 6 | - | - | - | - | - |
| W16 | 6 | 8 | 6 | + | + | + | + | + |
| W17 | 6 | 8 | 6 | - | - | - | - | - |
| W18 | 6 | 8 | 5 | - | - | - | + | - |
| W19 | 6 | 8 | 5 | - | - | - | - | - |
| W20 | 6 | 8 | 5 | + | + | + | + | + |
| Parameters | Surgery | HR (BPM) | SV (µL) | CO (mL/min) | LV Mass (mg) | LVAW;s (mm) | LVAW;d (mm) | LVPW;s (mm) | LVPW;d (mm) |
|---|---|---|---|---|---|---|---|---|---|
| Time | |||||||||
| Baseline | Sham | 494 ± 31.8 | 38.6 ± 3.1 | 18.8 ± 1.2 | 128 ± 13.2 | 1.5 ± 0.1 | 1.0 ± 0.1 | 1.4 ± 0.1 | 1.1 ± 0.1 |
| Moderate | 449 ± 14.7 | 35.5 ± 2.8 | 15.8 ± 1.1 | 134 ± 16.6 | 1.4 ± 0.1 | 0.9 ± 0.5 | 1.7 ± 0.1 | 1.3 ± 0.2 | |
| Severe | 447 ± 15.2 | 38.1 ± 3.2 | 17.1 ± 1.6 | 117 ± 4.8 | 1.3 ± 0.0 | 0.94 ± 0.0 | 1.4 ± 0.1 | 1.0 ± 0.1 | |
| 4weeks | Sham | 493 ± 21.8 | 35 ± 3.2 | 17.1 ± 1.3 | 119.4. ± 5.5 | 1.3 ± 0.1 | 0.94 ± 0.0 | 1.46 ± 0.1 | 1.1 ± 0.1 |
| Moderate | 482 ± 12.5 | 32 ± 3.3 | 15.5 ± 1.7 | 121 ± 10.2 | 1.4 ± 0.1 | 0.97 ± 0.0 | 1.6 ± 0.1 | 1.2 ± 0.1 | |
| Severe | 497 ± 12.4 | 32.9 ± 2.1 | 16.4 ± 1.3 | 134 ± 17.8 | 1.45 ± 0.1 | 1.1 ± 0.1 | 1.56 ± 0.1 | 1.1 ± 0.1 | |
| 8 weeks | Sham | 451 ± 21.1 | 40.2 ± 2.8 | 17.9 ± 0.7 | 103.3 ± 5.9 | 1.25 ± 0.1 | 0.9 ± 0.01 | 1.1 ± 0.0 | 0.8 ± 0.0 |
| Moderate | 478 ± 20.1 | 37.7 ± 1.1 | 17.9 ± 0.6 | 119.2 ± 12.1 | 1.5 ± 0.1 | 1.0 ± 0.1 | 1.3 ± 0.1 | 0.87 ± 0.1 | |
| Severe | 472 ± 16.2 | 35.8 ± 3.2 | 16.9 ± 1.7 | 113.3 ± 18.8 | 1.4 ± 0.1 | 0.9 ± 0.0 | 1.4 ± 0.1 | 0.98 ± 0.1 | |
| 12 weeks | Sham | 498 ± 16.3 | 36 ± 3.9 | 17.8 ± 1.8 | 143 ± 16.6 | 1.4 ± 0.1 | 1.0 ± 0.1 | 1.5 ± 0.1 | 1.16 ± 0.2 |
| Moderate | 492 ± 17.7 | 36.7 ± 1.5 | 18 ± 1.0 | 110.4 ± 7.5 | 1.3 ± 0.5 | 0.9 ± 0.0 | 1.2 ± 0.1 | 0.89 ± 0.1 | |
| Severe | 519 ± 15.7 | 37..5 ± 0.5 | 19.4 ± 0.5 | 110.6 ± 7.6 | 1.3 ± 0.1 | 0.9 ± 0.0 | 1.25 ± 0.1 | 0.89 ± 0.1 | |
| 16 weeks | Sham | 501 ± 24.7 | 35.9 ± 4.5 | 17.9 ± 2.3 | 118.7 ± 9.1 | 1.3 ± 0.1 | 0.9 ± 0.0 | 1.5 ± 0.1 | 1.1 ± 0.1 |
| Moderate | 476 ± 17.5 | 35.6 ± 2.0 | 16.9 ± 1.1 | 116.2 ± 7.8 | 1.4 ± 0.1 | 1.0 ± 0.1 | 1.3 ± 0.0 | 0.96 ± 0.0 | |
| Severe | 503 ± 3.5 | 36.5 ± 2.6 | 18.4 ± 1.3 | 131.6 ± 20.4 | 1.4 ± 0.1 | 0.96 ± 0.0 | 1.8 ± 0.2 | 1.2 ± 0.2 | |
| 20 weeks | Sham | 449 ± 10 | 37.8 ± 1.7 | 17 ± 0.8 | 142.4 ± 22.2 | 1.5 ± 0.0 | 1.1 ± 0.1 | 1.4 ± 0.1 | 1.1 ± 0.2 |
| Moderate | 480 ± 6.7 | 37.9 ± 3.3 | 18.1 ± 1.4 | 119.3 ± 6.9 | 1.5 ± 0.1 | 1.0 ± 0.0 | 1.4 ± 0.1 | 1.0 ± 0.1 | |
| Severe | 477 ± 17.5 | 42.6 ± 5.8 | 20 ± 2.4 | 119.6 ± 5.9 | 1.5 ± 0.1 | 1.0 ± 0.0 | 1.5 ± 0.1 | 1.0 ± 0.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ghnenis, A.B.; Jones, C.; Sefiani, A.; Douthitt, A.J.; Reyna, A.J.; Rutkowski, J.M.; Geoffroy, C.G. Evaluation of the Cardiometabolic Disorders after Spinal Cord Injury in Mice. Biology 2022, 11, 495. https://doi.org/10.3390/biology11040495
Ghnenis AB, Jones C, Sefiani A, Douthitt AJ, Reyna AJ, Rutkowski JM, Geoffroy CG. Evaluation of the Cardiometabolic Disorders after Spinal Cord Injury in Mice. Biology. 2022; 11(4):495. https://doi.org/10.3390/biology11040495
Chicago/Turabian StyleGhnenis, Adel B., Calvin Jones, Arthur Sefiani, Ashley J. Douthitt, Andrea J. Reyna, Joseph M. Rutkowski, and Cédric G. Geoffroy. 2022. "Evaluation of the Cardiometabolic Disorders after Spinal Cord Injury in Mice" Biology 11, no. 4: 495. https://doi.org/10.3390/biology11040495