Vitamin D Attenuates Oxidative Damage and Inflammation in Retinal Pigment Epithelial Cells


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Cell Viability Assay
2.3. Caspase-3/7 Activity
2.4. Measurement of Reactive Oxygen Species (ROS)
2.5. Enzyme-Linked Immunosorbent Assay (ELISA)
2.6. Biochemical Assay
2.7. Mouse Samples
2.8. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
2.9. Immunostaining
2.10. Western Blot
2.11. Statistical Data Analysis
3. Results
3.1. VITD Treatment Improved Cell Viability and Reduced ROS Production and Apoptosis
3.2. VITD Treatment Enhanced VDR) Expression in Stressed ARPE-19 Cells
3.3. VITD Treatment Upregulated the Expression of Antioxidant Genes
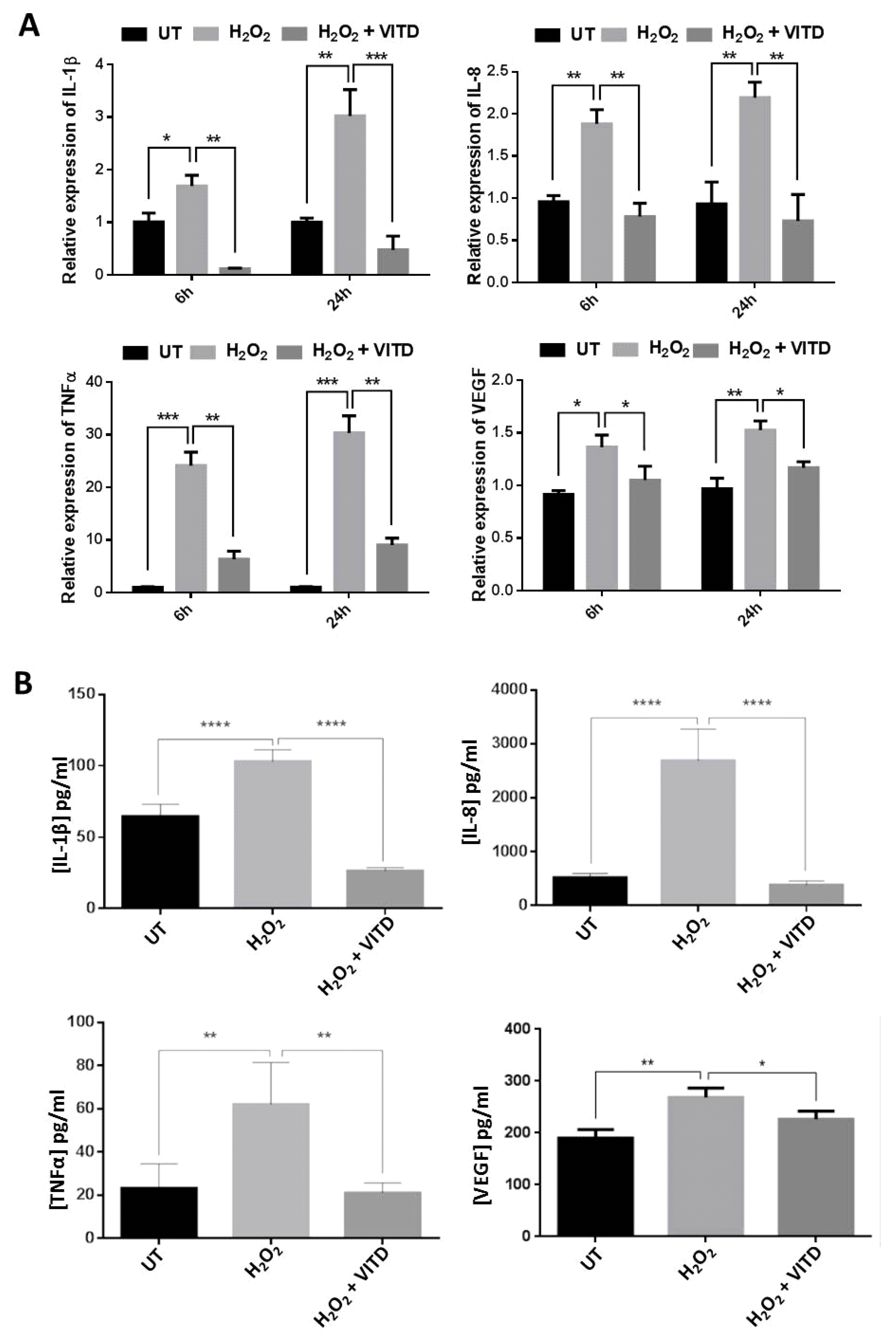
3.4. VITD Treatment Modulated the Expression of the Inflammatory Mediators
3.5. VITD Reversed the H2O2-Induced Change in NRF2 Expression
3.6. VITD Regulated IL-33 Expression
3.7. IL-33 Treatment Upregulated Antioxidant Gene Expression
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Cascella, R.; Ragazzo, M.; Strafella, C.; Missiroli, F.; Borgiani, P.; Angelucci, F.; Marsella, L.; Cusumano, A.; Novelli, G.; Ricci, F.; et al. Age-Related Macular Degeneration: Insights into Inflammatory Genes. J. Ophthalmol. 2014, 2014, 582842. [Google Scholar] [CrossRef] [PubMed]
- Priya, R.R.; Chew, E.Y.; Swaroop, A. Genetic studies of age-related macular degeneration: Lessons, challenges, and opportunities for disease management. Ophthalmology 2012, 119, 2526–2536. [Google Scholar] [CrossRef] [PubMed]
- Wong, W.L.; Su, X.; Li, X.; Cheung, C.M.; Klein, R.; Cheng, C.Y.; Wong, T.Y. Global prevalence of age-related macular degeneration and disease burden projection for 2020 and 2040: A systematic review and meta-analysis. Lancet Glob. Health 2014, 2, e106–e116. [Google Scholar] [CrossRef]
- Theodoropoulou, S.; Copland, D.A.; Liu, J.; Wu, J.; Gardner, P.J.; Ozaki, E.; Doyle, S.L.; Campbell, M.; Dick, A.D. Interleukin-33 regulates tissue. J. Pathol. 2017, 241, 45–56. [Google Scholar] [CrossRef] [PubMed]
- Holz, F.G.; Schmitz-Valckenberg, S.; Fleckenstein, M. Recent developments in the treatment of age-related macular degeneration. J. Clin. Investig. 2014, 124, 1430–1438. [Google Scholar] [CrossRef] [PubMed]
- Toomey, C.B.; Kelly, U.; Saban, D.R.; Bowes Rickman, C. Regulation of age-related macular degeneration-like pathology by complement factor H. Proc. Natl. Acad. Sci. USA 2015, 112, E3040–E3049. [Google Scholar] [CrossRef] [Green Version]
- Bikle, D.D. Vitamin D metabolism, mechanism of action, and clinical applications. Chem. Biol. 2014, 21, 319–329. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, A.V.; Feldman, D. Mechanisms of the anti-cancer and anti-inflammatory actions of vitamin D. Annu. Rev. Pharmacol. Toxicol. 2011, 51, 311–336. [Google Scholar] [CrossRef]
- Fernandes de Abreu, D.A.; Eyles, D.; Féron, F. Vitamin D, a neuro-immunomodulator: Implications for neurodegenerative and autoimmune diseases. Psychoneuroendocrinology 2009, 34, S265–S277. [Google Scholar] [CrossRef]
- Graffe, A.; Beauchet, O.; Fantino, B.; Milea, D.; Annweiler, C. Vitamin and macular thickness in the elderly: An optical coherence tomography study. Investig. Ophthalmol. Vis. Sci. 2014, 15, 5298–5303. [Google Scholar] [CrossRef]
- Itty, S.; Day, S.; Lyles, K.W.; Stinnett, S.S.; Vajzovic, L.M.; Mruthyunjaya, P. Vitamin deficiency in neovascular versus nonneovascular age-related macular degeneration. Retina 2014, 34, 1779–1786. [Google Scholar] [CrossRef]
- Millen, A.E.; Voland, R.; Sondel, S.A.; Parekh, N.; Horst, R.L.; Wallace, R.B.; Hageman, G.S.; Chappell, R.; Blodi, B.A.; Klein, M.L.; et al. Vitamin D status and early age-related macular degeneration in postmenopausal women. Arch. Ophthalmol. 2011, 129, 481–489. [Google Scholar] [CrossRef]
- Singh, A.; Falk, M.K.; Subhi, Y.; Sørensen, T.L. The association between plasma 25-hydroxyvitamin D and subgroups in age-related macular degeneration: A cross-sectional study. PLoS ONE 2013, 29, e70948. [Google Scholar] [CrossRef]
- Gadani, S.P.; Walsh, J.T.; Smirnov, I.; Zheng, J.; Kipnis, J. The glia-derived alarmin IL-33 orchestrates the immune response and promotes recovery following CNS injury. Neuron 2015, 85, 703–709. [Google Scholar] [CrossRef]
- Liew, F.; Pitman, N.; McInnes, I. Disease-associated functions of IL-33: The new kid in the IL-1 family. Nat. Rev. Immunol. 2010, 10, 103–110. [Google Scholar] [CrossRef]
- Yu, J.T.; Song, J.H.; Wang, N.D.; Wu, Z.C.; Zhang, Q.; Zhang, N.; Zhang, W.; Xuan, S.Y.; Tan, L. Implication of gene polymorphism in Chinese patients with Alzheimer’s disease. Neurobiol. Aging 2012, 33, 1014.e11–1014.e14. [Google Scholar] [CrossRef]
- Tohari, A.M.; Zhou, X.; Shu, X. Protection against oxidative stress by vitamin D in cone cells. Cell Biochem. Funct. 2016, 34, 82–94. [Google Scholar] [CrossRef]
- Chen, Y.; Zhang, J.; Ge, X.; Du, J.; Deb, D.; Li, Y. Vitamin D receptor inhibits nuclear factor κB activation by interacting with I κB kinase β protein. J. Biol. Chem. 2013, 288, 19450–19458. [Google Scholar] [CrossRef]
- Huang, Y.; Ho, Y.; Lai, C.; Chiu, C.; Wang, J. 1,25-Dihydroxyvitamin D3 attenuates endotoxin-induced production of inflammatory mediators by inhibiting MAPK activation in primary cortical neuron-glia cultures. J. Neuroinflamm. 2015, 12, 147. [Google Scholar] [CrossRef]
- Bao, B.Y.; Ting, H.J.; Hsu, J.W.; Lee, Y.F. Protective role of 1 alpha, 25-dihydroxyvitamin D3 against oxidative stress in human prostate epithelial cells. Int. J. Cancer 2008, 122, 2699–2706. [Google Scholar] [CrossRef]
- Hamden, K.; Carreau, S.; Jamoussi, K.; Miladi, S.; Lajmi, S.; Aloulou, D.; Ayadi, F.; Elfeki, A. 1 Alpha dihydroxyvitamin D3: Therapeutic and preventive effects against oxidative stress, hepatic, pancreatic and renal injury in alloxan-induced diabetes in rats. J. Nutr. Sci. Vitamin. 2009, 55, 215–222. [Google Scholar] [CrossRef]
- Peng, X.; Vaishnav, A.; Murillo, G.; Alimirah, F.; Torres, K.E.; Mehta, R.G. Protection against cellular stress by 25-hydroxyvitamin D3 in breast epithelial cells. J. Cell Biochem. 2010, 110, 1324–1333. [Google Scholar] [CrossRef]
- Polidoro, L.; Properzi, G.; Marampon, F.; Gravina, G.L.; Festuccia, C.; Di Cesare, E.; Scarsella, L.; Ciccarelli, C.; Zani, B.M.; Ferri, C. Vitamin D protects human endothelial cells from H₂O₂ oxidant injury through the Mek/Erk-Sirt1 axis activation. J. Cardiovasc. Transl. Res. 2013, 6, 221–231. [Google Scholar] [CrossRef]
- Redza-Dutordoir, M.; Averill-Bates, D.A. Activation of apoptosis pathways by reactive oxygen species. Biochim. Biophys. Acta 2016, 1863, 2977–2992. [Google Scholar] [CrossRef]
- Alsalem, J.; Patel, D.; Susarla, R.; Coca-Prados, M.; Bland, R.; Walker, E.; Rauz, S.; Wallace, G. Characterization of vitamin D production by human ocular barrier cells. Investig. Ophthalmol. Vis. Sci. 2014, 55, 2140–2141. [Google Scholar] [CrossRef]
- Dwyer, M.; Kazmin, D.; Hu, P.; McDonnell, D.; Malek, G. Research resource: Nuclear receptor atlas of human retinal pigment epithelial cells: Potential relevance to age-related macular degeneration. Mol. Endocrinol. 2011, 25, 360–372. [Google Scholar] [CrossRef]
- Ren, Z.; Li, W.; Zhao, Q.; Ma, L.; Zhu, J. The impact of 1,25-dihydroxy vitamin D3 on the expressions of vascular endothelial growth factor and transforming growth factor-β1 in the retinas of rats with diabetes. Diabetes Res. Clin. Pract. 2012, 98, 474–480. [Google Scholar] [CrossRef]
- Bhat, A.H.; Dar, K.B.; Anees, S.; Zargar, M.A.; Masood, A.; Sofi, M.A.; Ganie, S.A. Oxidative stress, mitochondrial dysfunction and neurodegenerative diseases: A mechanistic insight. Biomed. Pharmacother. 2015, 74, 101–110. [Google Scholar] [CrossRef]
- Nakai, K.; Fujii, H.; Kono, K.; Goto, S.; Kitazawa, R.; Kitazawa, S.; Hirata, M.; Shinohara, M.; Fukagawa, M.; Nishi, S. Vitamin D activates the Nrf2-Keap1 antioxidant pathway and ameliorates nephropathy in diabetic rats. Am. J. Hypertens. 2013, 27, 586–595. [Google Scholar] [CrossRef]
- Middleton, R.P.; Nelson, R.; Li, Q.; Blanton, A.; Labuda, J.A.; Vitt, J.; Inpanbutr, N. 1.25-Dihydroxyvitamin D3 and its analogues increase catalase at the mRNA, protein and activity level in a canine transitional carcinoma cell line. Vet. Comp. Oncol. 2015, 13, 452–463. [Google Scholar] [CrossRef]
- Eelen, G.; Verlinden, L.; Meyer, M.; Gijsbers, R.; Pike, J.; Bouillon, R.; Verstuyf, A. 1.25- Dihydroxyvitamin D3 and the aging-related forkhead box O and sestrin proteins in osteoblasts. J. Steroid Biochem. Mol. Biol. 2103, 136, 112–119. [Google Scholar] [CrossRef]
- Berridge, M.J. Vitamin D: A custodian of cell signalling stability in health and disease. Biochem. Soc. Trans. 2015, 43, 349–358. [Google Scholar] [CrossRef]
- Seki, K.; Sanada, S.; Kudinova, A.; Steinhauser, M.; Handa, V.; Gannon, J.; Lee, R. Interleukin-33 prevents apoptosis and improves survival after experimental myocardial infarction through ST2 signaling. Circ. Heart Fail. 2009, 2, 684–691. [Google Scholar] [CrossRef]
- Zhang, H.F.; Xie, S.L.; Chen, Y.X.; Mai, J.T.; Wang, J.F.; Zhu, W.L.; Zhu, L.G. Altered serum levels of IL-33 in patients with advanced systolic chronic heart failure: Correlation with oxidative stress. J. Transl. Med. 2012, 8, 120. [Google Scholar] [CrossRef]
- Barbour, M.; Allan, D.; Xu, H.; Pei, C.; Chen, M.; Niedbala, W.; Fukada, S.Y.; Besnard, A.G.; Alves-Filho, J.C.; Tong, X.; et al. IL-33 attenuates the development of experimental autoimmune uveitis. Eur. J. Immunol. 2014, 44, 3320–3329. [Google Scholar] [CrossRef] [Green Version]
- Xi, H.; Katschke, K.J.; Li, Y.; Truong, T.; Lee, W.P.; Diehl, L.; Rangell, L.; Tao, J.; Arceo, R.; Eastham-Anderson, J.; et al. IL-33 amplifies an innate immune response in the degenerating retina. J. Exp. Med. 2016, 213, 189–207. [Google Scholar] [CrossRef]
- Rui, T.; Zhang, J.; Xu, X.; Yao, Y.; Kao, R.; Martin, C. Reduction in IL-33 expression exaggerates ischaemia/reperfusion-induced myocardial injury in mice with diabetes mellitus. Cardiovasc. Res. 2012, 94, 370–378. [Google Scholar] [CrossRef]
- Layana, A.; Minnella, A.; Garhöfer, G.; Aslam, T.; Holz, F.; Leys, A.; Silva, R.; Delcourt, C.; Souied, E.; Seddon, J. Vitamin D and Age-Related Macular Degeneration. Nutrients 2017, 9, 1120. [Google Scholar] [CrossRef]
- Morrison, M.A.; Silveira, A.C.; Huynh, N.; Jun, G.; Smith, S.E.; Zacharaki, F.; Sato, H.; Loomis, S.; Andreoli, M.T.; Adams, S.M.; et al. Systems biology-based analysis implicates a novel role for vitamin D metabolism in the pathogenesis of age-related macular degeneration. Hum. Genom. 2011, 5, 538–568. [Google Scholar] [CrossRef] [Green Version]
- Parekh, N.; Chappell, R.J.; Millen, A.E.; Albert, D.M.; Mares, J.A. Association between vitamin D and age-related macular degeneration in the Third National Health and Nutrition Examination Survey, 1988 through 1994. Arch. Ophthalmol. 2007, 125, 661–669. [Google Scholar] [CrossRef]
- Lee, V.; Rekhi, E.; Hoh Kam, J.; Jeffery, G. Vitamin D rejuvenates aging eyes by reducing inflammation, clearing beta and improving visual function. Neurobiol. Aging 2012, 33, 2382–2389. [Google Scholar] [CrossRef]
- Christakos, S.; Dhawan, P.; Peng, X.; Obukhov, A.; Nowycky, M.; Benn, B.; Zhong, Y.; Liu, Y.; Shen, Q. New insights into the function and regulation of vitamin D target proteins. J. Steroid Biochem. Mol. Biol. 2007, 103, 405–410. [Google Scholar] [CrossRef] [Green Version]
- Zella, L.; Kim, S.; Shevde, N.; Pike, J. Enhancers located within two introns of the vitamin D receptor gene mediate transcriptional autoregulation by 1,25-dihydroxyvitamin D3. Mol. Endocrinol. 2006, 20, 1231–1247. [Google Scholar] [CrossRef]
- Eshaq, R.; Wright, W.; Harris, N. Oxygen delivery, consumption, and conversion to reactive oxygen species in experimental models of diabetic retinopathy. Redox Biol. 2014, 2, 661–666. [Google Scholar] [CrossRef] [Green Version]
- Samuel, S.; Sitrin, M.D. Vitamin D’s role in cell proliferation and differentiation. Nutr. Rev. 2008, 66, S116–S124. [Google Scholar] [CrossRef]
- Du, L.X.; Wang, Y.Q.; Hua, G.Q.; Mi, W.L. IL-33/ST2 pathway as a rational therapeutic target for CNS diseases. Neuroscience 2018, 369, 222–230. [Google Scholar] [CrossRef]
- Liu, X.C.; Liu, X.F.; Jian, C.X.; Li, C.J.; He, S.Z. IL-33 is induced by amyloid-β stimulation and regulates inflammatory cytokine production in retinal pigment epithelium cells. Inflammation 2012, 35, 776–784. [Google Scholar] [CrossRef]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tohari, A.M.; Alhasani, R.H.; Biswas, L.; Patnaik, S.R.; Reilly, J.; Zeng, Z.; Shu, X. Vitamin D Attenuates Oxidative Damage and Inflammation in Retinal Pigment Epithelial Cells. Antioxidants 2019, 8, 341. https://doi.org/10.3390/antiox8090341
Tohari AM, Alhasani RH, Biswas L, Patnaik SR, Reilly J, Zeng Z, Shu X. Vitamin D Attenuates Oxidative Damage and Inflammation in Retinal Pigment Epithelial Cells. Antioxidants. 2019; 8(9):341. https://doi.org/10.3390/antiox8090341
Chicago/Turabian StyleTohari, Ali Mohammad, Reem Hasaballah Alhasani, Lincoln Biswas, Sarita Rani Patnaik, James Reilly, Zhihong Zeng, and Xinhua Shu. 2019. "Vitamin D Attenuates Oxidative Damage and Inflammation in Retinal Pigment Epithelial Cells" Antioxidants 8, no. 9: 341. https://doi.org/10.3390/antiox8090341