Screening of In Vitro Health Benefits of Tangerine Tomatoes




Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Tomato Varieties and Extracts
2.3. Prostate Cancer Cell Lines and Culture Media
2.4. Cell Lines and Culture Media for the Anti-Inflammatory Assays
2.5. Antioxidant Assays
2.5.1. FRAP Assay
2.5.2. ABTS Assay
2.6. Prostate Cancer Proliferation Assay
2.7. Anti-Inflammatory Assays
2.8. Data Analyses
3. Results
3.1. Antioxidant Assays
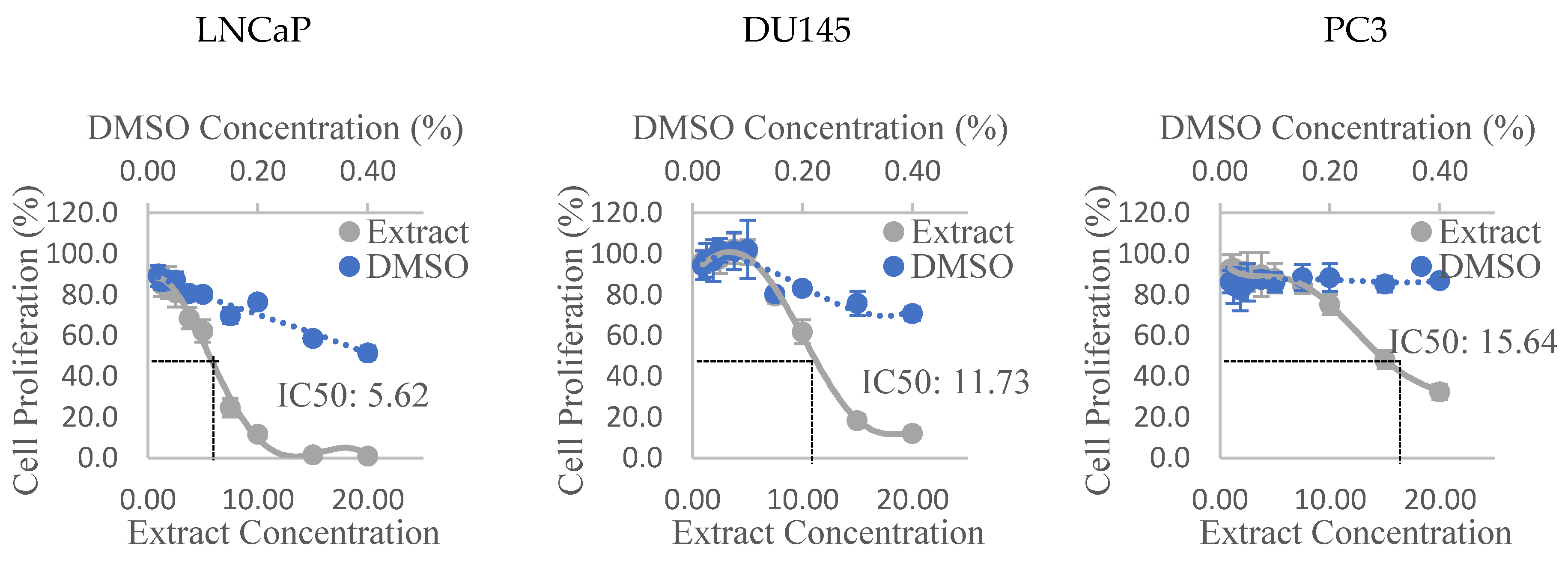
3.2. Anti-Proliferative Activity
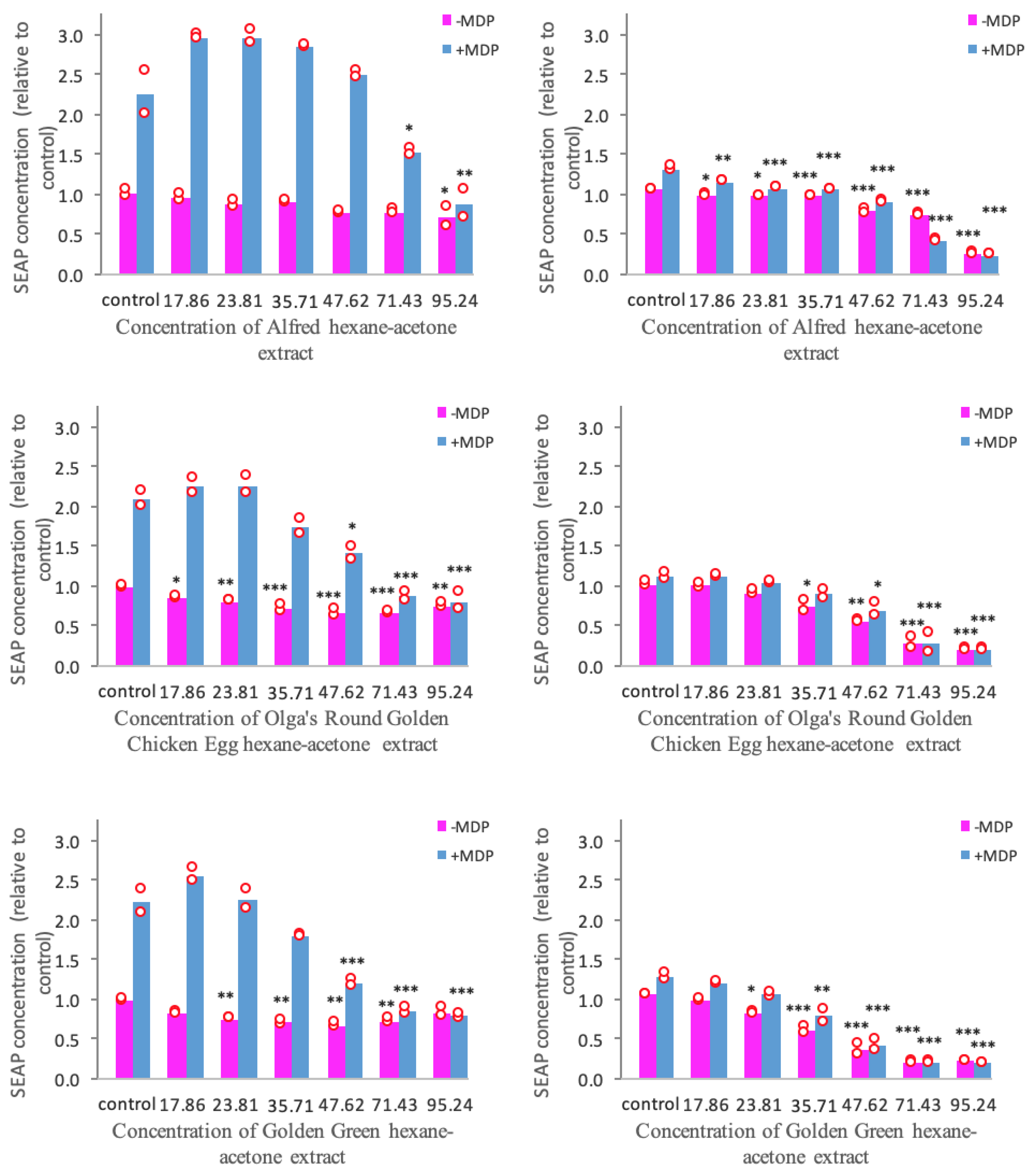
3.3. Anti-Inflammatory Activity Mediated through the TLR2 and TLR4 Pathways
3.4. NOD2-Mediated Anti-Inflammatory Pathway
4. Discussion
4.1. Antioxidant Activity
4.2. Anti-Proliferative Activity
4.3. Anti-Inflammatory Activity
4.3.1. TLR2 and TLR4 Pathways
4.3.2. NOD2 Pathway
4.4. Future Research
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cheng, H.M.; Koutsidis, G.; Lodge, J.K.; Ashor, A.; Siervo, M.; Lara, J. Tomato and lycopene supplementation and cardiovascular risk factors: A systematic review and meta-analysis. Atherosclerosis 2017, 257, 100–108. [Google Scholar] [CrossRef] [PubMed]
- Rowles, J.L.; Ranard, K.M.; Applegate, C.C.; Jeon, S.; An, R.; Erdman, J.W. Processed and raw tomato consumption and risk of prostate cancer: A systematic review and dose–response meta-analysis. Prostate Cancer Prostatic Dis. 2018, 21, 319–336. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Yang, X.; Wang, X.; Wang, Y.; Song, Z. The role of tomato products and lycopene in the prevention of gastric cancer: A meta-analysis of epidemiologic studies. Med. Hypotheses 2013, 80, 383–388. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Xu, J. Meta-analysis of the association between dietary lycopene intake and ovarian cancer risk in postmenopausal women. Sci. Rep. 2014, 4, 4885. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.M.; Koutsidis, G.; Lodge, J.K.; Ashor, A.W.; Siervo, M.; Lara, J. Lycopene and tomato and risk of cardiovascular diseases: A systematic review and meta-analysis of epidemiological evidence. Crit. Rev. Food Sci. Nutr. 2019, 59, 141–158. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Zhang, W.; Wang, X.; Zhao, K.; Negi, D.S.; Zhuo, L.; Qi, M.; Wang, X.; Zhang, X. Lycopene and risk of prostate cancer: A systematic review and meta-analysis. Medicine 2015, 94. [Google Scholar] [CrossRef] [PubMed]
- Cooperstone, J.L.; Ralston, R.A.; Riedl, K.M.; Haufe, T.C.; Schweiggert, R.M.; King, S.A.; Timmers, C.D.; Francis, D.M.; Lesinski, G.B.; Clinton, S.K. Enhanced bioavailability of lycopene when consumed as cis-isomers from tangerine compared to red tomato juice, a randomized, cross-over clinical trial. Mol. Nutr. Food Res. 2015, 59, 658–669. [Google Scholar] [CrossRef]
- Unlu, N.Z.; Bohn, T.; Francis, D.; Clinton, S.K.; Schwartz, S.J. Carotenoid absorption in humans consuming tomato sauces obtained from tangerine or high-β-carotene varieties of tomatoes. J. Agric. Food Chem. 2007, 55, 1597–1603. [Google Scholar] [CrossRef]
- Gärtner, C.; Stahl, W.; Sies, H. Lycopene is more bioavailable from tomato paste than from fresh tomatoes. Am. J. Clin. Nutr. 1997, 66, 116–122. [Google Scholar] [CrossRef]
- Boileau, T.W.M.; Boileau, A.C.; Erdman, J.W., Jr. Bioavailability of all-trans and cis–Isomers of Lycopene. Exp. Biol. Med. 2002, 227, 914–919. [Google Scholar] [CrossRef]
- Wang, X.D. Lycopene metabolism and its biological significance. Am. J. Clin. Nutr. 2012, 96, 1214S–1222S. [Google Scholar] [CrossRef] [PubMed]
- Isaacson, T.; Ronen, G.; Zamir, D.; Hirschberg, J. Cloning of tangerine from tomato reveals a carotenoid isomerase essential for the production of β-carotene and xanthophylls in plants. Plant Cell 2002, 14, 333–342. [Google Scholar] [CrossRef] [PubMed]
- Wertz, K.; Siler, U.; Goralczyk, R. Lycopene: Modes of action to promote prostate health. Arch. Biochem. Biophys. 2004, 430, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: Globocan estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed]
- Ministry of Health. New Cancer Registrations 2016. Available online: https://www.health.govt.nz/publication/new-cancer-registrations-2016 (accessed on 27 May 2019).
- Hwang, E.S.; Bowen, P.E. Effects of tomato paste extracts on cell proliferation, cell-cycle arrest and apoptosis in LNCaP human prostate cancer cells. Biofactors 2005, 23, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Rao, A.; Waseem, Z.; Agarwal, S. Lycopene content of tomatoes and tomato products and their contribution to dietary lycopene. Food Res. Int. 1998, 31, 737–741. [Google Scholar] [CrossRef]
- Clough, J.M.; Pattenden, G. Stereochemical assignment of prolycopene and other poly-Z-isomeric carotenoids in fruits of the tangerine tomato Lycopersicon esculentum var.‘Tangella’. J. Chem. Soc. Perkin Trans. 1 1983, 3011–3018. [Google Scholar] [CrossRef]
- Taungbodhitham, A.K.; Jones, G.P.; Wahlqvist, M.L.; Briggs, D.R. Evaluation of extraction method for the analysis of carotenoids in fruits and vegetables. Food Chem. 1998, 63, 577–584. [Google Scholar] [CrossRef]
- Masters, J.R.; Thomson, J.A.; Daly-Burns, B.; Reid, Y.A.; Dirks, W.G.; Packer, P.; Toji, L.H.; Ohno, T.; Tanabe, H.; Arlett, C.F. Short tandem repeat profiling provides an international reference standard for human cell lines. Proc. Natl. Acad. Sci. USA 2001, 98, 8012–8017. [Google Scholar] [CrossRef]
- Philpott, M.; Mackay, L.; Ferguson, L.R.; Forbes, D.; Skinner, M. Cell culture models in developing nutrigenomics foods for inflammatory bowel disease. Mutat. Res./Fundam. Mol. Mech. Mutagenes. 2007, 622, 94–102. [Google Scholar] [CrossRef]
- Folkard, D.L.; Marlow, G.; Mithen, R.F.; Ferguson, L.R. Effect of Sulforaphane on NOD2 via NF-κB: Implications for Crohn’s disease. J. Inflamm. 2015, 12, 6. [Google Scholar] [CrossRef]
- Benzie, I.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef]
- Miller, N.J.; Rice-Evans, C.; Davies, M.J.; Gopinathan, V.; Milner, A. A novel method for measuring antioxidant capacity and its application to monitoring the antioxidant status in premature neonates. Clin. Sci. 1993, 84, 407–412. [Google Scholar] [CrossRef]
- Ozgen, M.; Reese, R.N.; Tulio, A.Z.; Scheerens, J.C.; Miller, A.R. Modified 2, 2-azino-bis-3-ethylbenzothiazoline-6-sulfonic acid (ABTS) method to measure antioxidant capacity of selected small fruits and comparison to ferric reducing antioxidant power (FRAP) and 2, 2 ‘-diphenyl-1-picrylhydrazyl (DPPH) methods. J. Agric. Food Chem. 2006, 54, 1151–1157. [Google Scholar] [CrossRef]
- Müller, L.; Goupy, P.; Fröhlich, K.; Dangles, O.; Caris-Veyrat, C.; Böhm, V. Comparative study on antioxidant activity of lycopene (Z)-isomers in different assays. J. Agric. Food Chem. 2011, 59, 4504–4511. [Google Scholar] [CrossRef]
- Skehan, P.; Storeng, R.; Scudiero, D.; Monks, A.; McMahon, J.; Vistica, D.; Warren, J.T.; Bokesch, H.; Kenney, S.; Boyd, M.R. New colorimetric cytotoxicity assay for anticancer-drug screening. JNCI J. Natl. Cancer Inst. 1990, 82, 1107–1112. [Google Scholar] [CrossRef]
- Tomblin, V.; Ferguson, L.R.; Han, D.Y.; Murray, P.; Schlothauer, R. Potential pathway of anti-inflammatory effect by New Zealand honeys. Int. J. Gen. Med. 2014, 7, 149. [Google Scholar] [CrossRef]
- Peng, Y.; Bishop, K.; Ferguson, L.; Quek, S. Screening of Cytotoxicity and Anti-Inflammatory Properties of Feijoa Extracts Using Genetically Modified Cell Models Targeting TLR2, TLR4 and NOD2 Pathways, and the Implication for Inflammatory Bowel Disease. Nutrients 2018, 10, 1188. [Google Scholar] [CrossRef]
- Cohen, J. Statistical Power Analysis for the Behavioral Sciences, 2nd ed.; Routledge: New York, NY, USA, 1988; pp. 66–67. [Google Scholar]
- Apak, R.; Gorinstein, S.; Böhm, V.; Schaich, K.M.; Özyürek, M.; Güçlü, K. Methods of measurement and evaluation of natural antioxidant capacity/activity (IUPAC Technical Report). Pure Appl. Chem. 2013, 85, 957–998. [Google Scholar] [CrossRef]
- AOAC. AOAC SMPR 2011.011 Standard Method Performance Requirements for In Vitro Determination of Total Antioxidant Activity. Available online: http://www.aoac.org/aoac_prod_imis/AOAC_Docs/OMA/OMAChangesinMethods_050913.pdf (accessed on 27 May 2019).
- Schaich, K.M.; Tian, X.; Xie, J. Reprint of “Hurdles and pitfalls in measuring antioxidant efficacy: A critical evaluation of ABTS, DPPH, and ORAC assays”. J. Funct. Foods 2015, 18, 782–796. [Google Scholar] [CrossRef]
- Müller, L.; Fröhlich, K.; Böhm, V. Comparative antioxidant activities of carotenoids measured by ferric reducing antioxidant power (FRAP), ABTS bleaching assay (αTEAC), DPPH assay and peroxyl radical scavenging assay. Food Chem. 2011, 129, 139–148. [Google Scholar] [CrossRef]
- McGhie, T.K.; Cordiner, S. Carotenoid Composition of Tomatoes. Available online: https://www.heritagefoodcrops.org.nz/static/media/2015-carotenoid-composition-of-tomatoes.9d149f68.pdf (accessed on 27 May 2019).
- Kotake-Nara, E.; Kushiro, M.; Zhang, H.; Sugawara, T.; Miyashita, K.; Nagao, A. Carotenoids affect proliferation of human prostate cancer cells. J. Nutr. 2001, 131, 3303–3306. [Google Scholar] [CrossRef]
- Williams, A.W.; Boileau, T.W.-M.; Zhou, J.R.; Clinton, S.K.; Erdman, J.W., Jr. β-Carotene modulates human prostate cancer cell growth and may undergo intracellular metabolism to retinol. J. Nutr. 2000, 130, 728–732. [Google Scholar] [CrossRef]
- Dulińska, J.; Gil, D.; Zagajewski, J.; Hartwich, J.; Bodzioch, M.; Dembińska-Kieć, A.; Langmann, T.; Schmitz, G.; Laidler, P. Different effect of beta-carotene on proliferation of prostate cancer cells. Biochimica Biophysica Acta (BBA)-Mol. Basis Dis. 2005, 1740, 189–201. [Google Scholar] [CrossRef]
- Bank, S.; Andersen, P.S.; Burisch, J.; Pedersen, N.; Roug, S.; Galsgaard, J.; Turino, S.Y.; Brodersen, J.B.; Rashid, S.; Rasmussen, B.K. Polymorphisms in the inflammatory pathway genes TLR2, TLR4, TLR9, LY96, NFKBIA, NFKB1, TNFA, TNFRSF1A, IL6R, IL10, IL23R, PTPN22, and PPARG are associated with susceptibility of inflammatory bowel disease in a Danish cohort. PLoS ONE 2014, 9, e98815. [Google Scholar] [CrossRef]
- Jostins, L.; Ripke, S.; Weersma, R.K.; Duerr, R.H.; McGovern, D.P.; Hui, K.Y.; Lee, J.C.; Schumm, L.P.; Sharma, Y.; Anderson, C.A. Host–microbe interactions have shaped the genetic architecture of inflammatory bowel disease. Nature 2012, 491, 119. [Google Scholar] [CrossRef]
- Curkovic, I.; Egbring, M.; Kullak-Ublick, G.A. Risks of inflammatory bowel disease treatment with glucocorticosteroids and aminosalicylates. Dig. Dis. 2013, 31, 368–373. [Google Scholar] [CrossRef]
- Hou, J.K.; Lee, D.; Lewis, J. Diet and inflammatory bowel disease: Review of patient-targeted recommendations. Clin. Gastroenterol. Hepatol. 2014, 12, 1592–1600. [Google Scholar] [CrossRef]
- Hartman, C.; Eliakim, R.; Shamir, R. Nutritional status and nutritional therapy in inflammatory bowel diseases. World J. Gastroenterol. WJG 2009, 15, 2570. [Google Scholar] [CrossRef]
- Marcotorchino, J.; Romier, B.; Gouranton, E.; Riollet, C.; Gleize, B.; Malezet-Desmoulins, C.; Landrier, J.F. Lycopene attenuates LPS-induced TNF-α secretion in macrophages and inflammatory markers in adipocytes exposed to macrophage-conditioned media. Mol. Nutr. Food Res. 2012, 56, 725–732. [Google Scholar] [CrossRef]
- Riso, P.; Visioli, F.; Grande, S.; Guarnieri, S.; Gardana, C.; Simonetti, P.; Porrini, M. Effect of a tomato-based drink on markers of inflammation, immunomodulation, and oxidative stress. J. Agric. Food Chem. 2006, 54, 2563–2566. [Google Scholar] [CrossRef]
- Maekawa, S.; Ohto, U.; Shibata, T.; Miyake, K.; Shimizu, T. Crystal structure of NOD2 and its implications in human disease. Nat. Commun. 2016, 7, 11813. [Google Scholar] [CrossRef]
- Cooperstone, J.L.; Francis, D.M.; Schwartz, S.J. Thermal processing differentially affects lycopene and other carotenoids in cis-lycopene containing, tangerine tomatoes. Food Chem. 2016, 210, 466–472. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Solvent | Ratio | Source |
|---|---|---|
| Hexane | - | [17] |
| Hexane:Acetone:Ethanol | 2:1:1 | [18] |
| Hexane:Acetone | 1:1 | [19] |
| Hexane:Ethanol | 3:4 | [20] |
| Extraction Solvent | Tomato Variety | |||
|---|---|---|---|---|
| Alfred | Olga’s Round Golden Chicken Egg | Golden Green | Golden Eye | |
| FRAP Assay | ||||
| Hexane | 11.71 ± 0.62 d/B | 6.24 ± 0.30 c/A | 12.52 ± 0.54 c/B | 12.50 ± 0.35 d/B |
| HAE | 3.66 ± 0.12 b | 2.23 ± 0.21 a | 5.99 ± 0.31 a | 6.18 ± 0.34 b |
| HA | 7.10 ± 0.45 c | 4.72 ± 0.36 b | 10.54 ± 0.17 b | 8.60 ± 0.54 c |
| HE | 2.11 ± 0.22 a | 1.79 ± 0.22 a | 5.58 ± 0.25 a | 3.23 ± 0.23 a |
| ABTS Assay | ||||
| Hexane | 20.04 ± 1.40 d/B | 14.27 ± 1.03 d/A | 26.22 ± 0.32 c/C | 18.32 ± 1.16 d/B |
| HAE | 6.23 ± 0.50 b | 6.82 ± 0.64 b | 14.42 ± 1.06 a | 8.24 ± 0.70 b |
| HA | 13.84 ± 0.33 c | 11.46 ± 1.16 c | 19.76 ± 1.60 b | 11.87 ± 0.69 c |
| HE | 1.99 ± 0.20 a | 6.11 ± 0.44 a | 13.42 ± 1.04 a | 3.77 ± 0.26 a |
| Tomato Variety | Extraction Solvent | |||
|---|---|---|---|---|
| Hexane | HAE | HA | HE | |
| LNCaP | ||||
| Alfred | 14.46 | X | 11.76† | X |
| Olga’s | 5.62 † | 16.93 | 10.1 | X |
| Golden Green | 8.08 † | 15.45 | 10.19 | X |
| Golden Eye | + | + | + | + |
| DU145 | ||||
| Alfred | X | X | -† | X |
| Olga’s | 11.73 † | X | - | X |
| Golden Green | 17.84 † | X | 18.01 | X |
| Golden Eye | + | + | + | + |
| PC3 | ||||
| Alfred | X | X | -† | X |
| Olga’s | 15.64 † | X | - | X |
| Golden Green | -† | X | X | X |
| Golden Eye | + | + | + | + |
| - | Concentration (mg Dry Tomato Equivalent/mL) | Ibuprofen (mg/mL) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Alfred Hexane | Alfred HA | Olga’s Hexane | Olga’s HA | GG Hexane | GG HA | GE Hexane | GE HA | ||
| HEK-Blue™ hTLR2 | |||||||||
| IC30 | 60.24 | 59.35 | 53.69 * | 39.96 | 50.01 * | 32.83 | N/A | N/A | 0.27 |
| IC50 | 88.46 | 74.73 | 66.22 * | 55.62 | 61.16 * | 50.41 | N/A | N/A | 0.4 |
| IC70 | N/A | 90.1 | 78.75 * | 71.28 | 72.3 * | 67.99 | N/A | N/A | N/A |
| HEK-Blue™ hTLR4 | |||||||||
| IC30 | 84.56 * | 64.47 | 52.2 * | 51.51 | 56.69 * | 42.76 | N/A | N/A | 0.15 |
| IC50 | N/A | 76.01 | 61.44 * | 62.53 | 62.89 * | 53.08 | N/A | N/A | 0.27 |
| IC70 | N/A | 87.55 | 70.68 * | 73.55 | 69.09 * | 63.4 | N/A | N/A | 0.38 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tanambell, H.; Quek, S.Y.; Bishop, K.S. Screening of In Vitro Health Benefits of Tangerine Tomatoes. Antioxidants 2019, 8, 230. https://doi.org/10.3390/antiox8070230
Tanambell H, Quek SY, Bishop KS. Screening of In Vitro Health Benefits of Tangerine Tomatoes. Antioxidants. 2019; 8(7):230. https://doi.org/10.3390/antiox8070230
Chicago/Turabian StyleTanambell, Hartono, Siew Young Quek, and Karen Suzanne Bishop. 2019. "Screening of In Vitro Health Benefits of Tangerine Tomatoes" Antioxidants 8, no. 7: 230. https://doi.org/10.3390/antiox8070230
APA StyleTanambell, H., Quek, S. Y., & Bishop, K. S. (2019). Screening of In Vitro Health Benefits of Tangerine Tomatoes. Antioxidants, 8(7), 230. https://doi.org/10.3390/antiox8070230