1. Introduction
In vitro cell models are often employed as high throughput screening tools used to gather data sets that are difficult to obtain in vivo. For example, cell proliferation assays can be used to indicate cytotoxicity and inform animal and human studies targeted to assessing the biological function of new therapeutics or functional ingredients from fruits, vegetables, and other plant sources. Cell proliferation assays exploit various cell functions, such as mitochondrial enzyme activity, cell membrane permeability, ATP production, or measurements of cellular DNA content. The CellTiter 96
® AQueous One Solution Cell Proliferation Assay (Promega Corporation, Madison, WI, USA) is one such assay and contains a tetrazolium compound, MTS (3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium inner salt), and an electron coupling reagent PES (phenazine ethosulfate). The MTS reagent is reduced by cells into a coloured 1-(4,5-dimethylthiazol-2-yl)-3,5-diphenylformazan product that is soluble in cell culture medium and measured by a change in absorbance at 490 nm. This reduction occurs in the presence of NADPH or NADH produced by dehydrogenase enzymes in metabolically active cells [
1].
Studies have revealed that some compounds found in plant extracts interfere with the estimation of cell viability using related cell proliferation assays, like the MTT (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide) assay, potentially leading to an overestimation of viable cells [
2]. The MTT assay is based on the reduction of the tetrazolium dye MTT to insoluble formazan, while related tetrazolium dyes, such as MTS (3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfonyl)-2H-tetrazolium), are used in conjunction with the intermediate electron acceptor PMS (1-methoxy phenazine methosulfate). Some phenolics, like kaempferol and epigallocatechin-3-gallate found in green tea, have been reported to reduce MTT in cell-free media, and when in the presence of cells, interference of the phenolics with MTT can be reduced by washing the cells prior to adding the MTT reagent [
3]. Other biological molecules, like proteins and carbohydrates with reductive capacity, can also reduce MTT-based reagents.
Terminalia ferdinandiana is a native Australian fruit that has been used by Indigenous people for centuries on hunting trips for quick energy and refreshment. Recent research has shown that
T. ferdinandiana has the highest reported vitamin C content of all fruits, and is rich in other hydrophilic (phenolics, flavonols) and lipophilic (α-tocopherol, γ-tocopherol, δ-tocopherol, lutein, chlorophyll a and b) antioxidants, nutrients, and other phenolics such as ellagic acid [
4,
5]. Overall, ellagic acid and vitamin C are the most abundant, and possibly the most bioactive antioxidants present in
T. ferdinandiana fruits [
6]. To investigate the general safety and bioactivity of fruit extracts using various cell-based in vitro assays (like antioxidant or anti-inflammatory assays), initial assessments of cytotoxicity are needed to determine cell treatment ranges and ensure that bioactivity can be observed in viable cells. To identify the most suitable cell viability assay, different assays were used including the CellTiter 96
® AQ
ueous One Solution Cell Proliferation Assay, CellTiter-Blue
® Cell Viability Assay (Promega Corporation, Madison, WI, USA), and the CyQUANT
® NF Assay (Invitrogen™, Molecular Probes™, Thermo Fisher Scientific Corporation, Carlsbad, CA, USA). Based on the results observed in this study, the variable composition of different plant extracts necessitates careful selection of cell viability assays to remove, or at least reduce, direct interaction of extracts with cell viability reagents masking the actual impact of extracts on cell proliferation in vitro.
3. Results and Discussion
In this study, we initially employed the CellTiter 96
® AQ
ueous One Solution Cell Proliferation Assay to measure the effect of antioxidant-rich
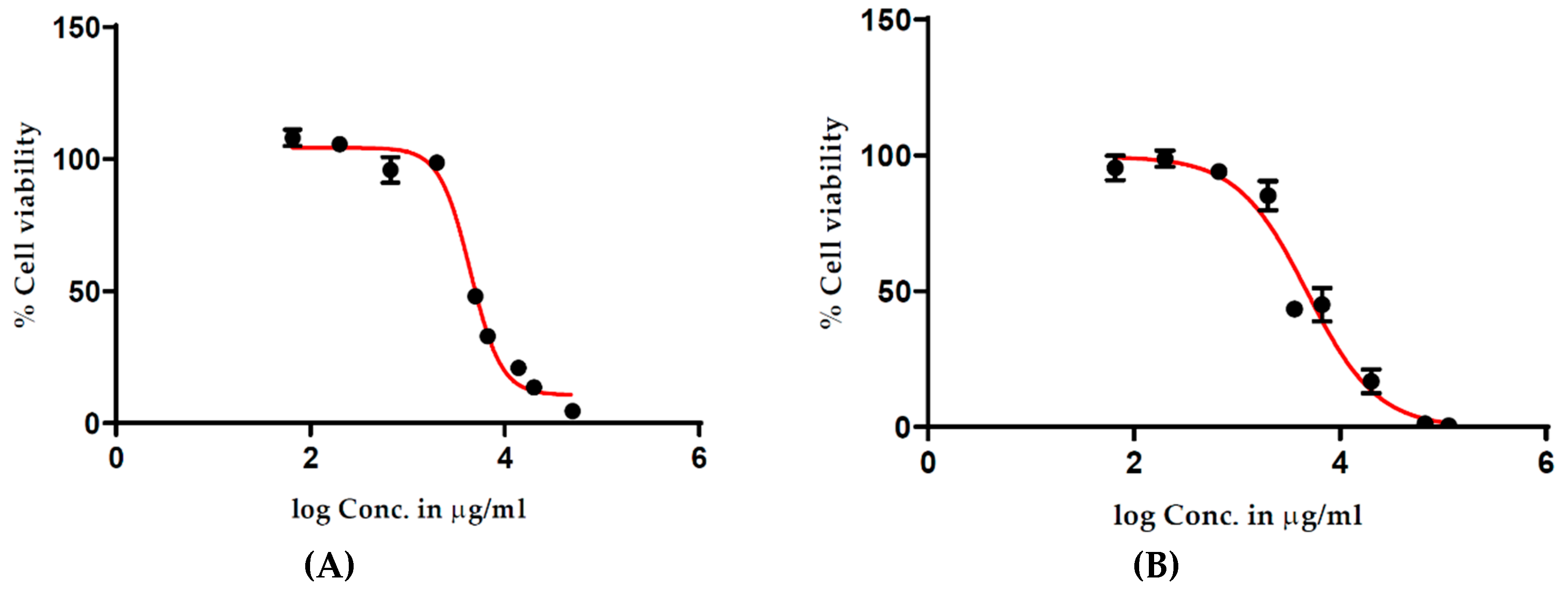
T. ferdinandiana extracts on the viability of human HT29-MTX-E12 cells. Cells were treated with 1000–10,000 µg/mL extracts for 2 h. Cell viability was expressed as a percentage compared to untreated cells with treatment–response curves drawn by plotting the cell viability percentage against the extract treatment concentration. An unexpected increase in cell viability was observed at higher treatment concentrations (
Figure 1), suggesting a 20% increase in cell viability following the 2 h treatment in response to 4000–8000 µg/mL of methanolic (
Figure 1A) and water extracts (
Figure 1B). Conversely, cell viability appeared to be lower with lower treatment concentrations of the extracts. These results suggested interactions between the cell viability assay reagents and the extracts.
A cell-free experiment was then performed to determine whether the fruit extract could directly bio-reduce the cell viability assay reagent, creating false cell viability results. All the absorbance readings from methanolic (
Figure 1C) and water (
Figure 1D) extracts increased in accordance with an increase in treatment concentrations, providing evidence that even in the absence of cells, the fruit extracts were reducing the assay reagent. In an attempt to identify a more suitable viability assay, we performed a CellTiter-Blue
® Cell Viability Assay, commonly known as the resazurin assay. This assay is based on the reduction of resazurin to its highly fluorescent product resorufin by metabolically active cells, with fluorescence proportional to the number of viable cells. Results of a cell-free experiments using the CellTiter-Blue
® Cell Viability Assay showed increased fluorescence with increasing concentrations of the methanolic (
Figure 2A) and water (
Figure 2B) extracts of
T. ferdinandiana fruits. No autofluorescence was observed in response to the extracts alone.
To identify a cell viability assay compatible with antioxidant-rich extracts that could provide a more accurate estimation of cell viability, the CyQUANT
® NF Cell Proliferation Assay was employed. The CyQUANT
® NF Cell Proliferation assay measures cellular DNA content via fluorescent dye binding, and with cellular DNA content highly regulated and closely proportional to the cell number, cell viability is measured by fluorescence [
8]. The CyQUANT
® NF assay results are presented in
Figure 3 and show a typical treatment–response curve, with reduced cell viability observed at higher concentrations of the methanolic (
Figure 3A) and water (
Figure 3B) extracts of fruits. Subsequently, by comparing the results from the different assays, it appears as though the antioxidants present in the extract promote the conversion of MTS to formazan, producing false positive viability results.
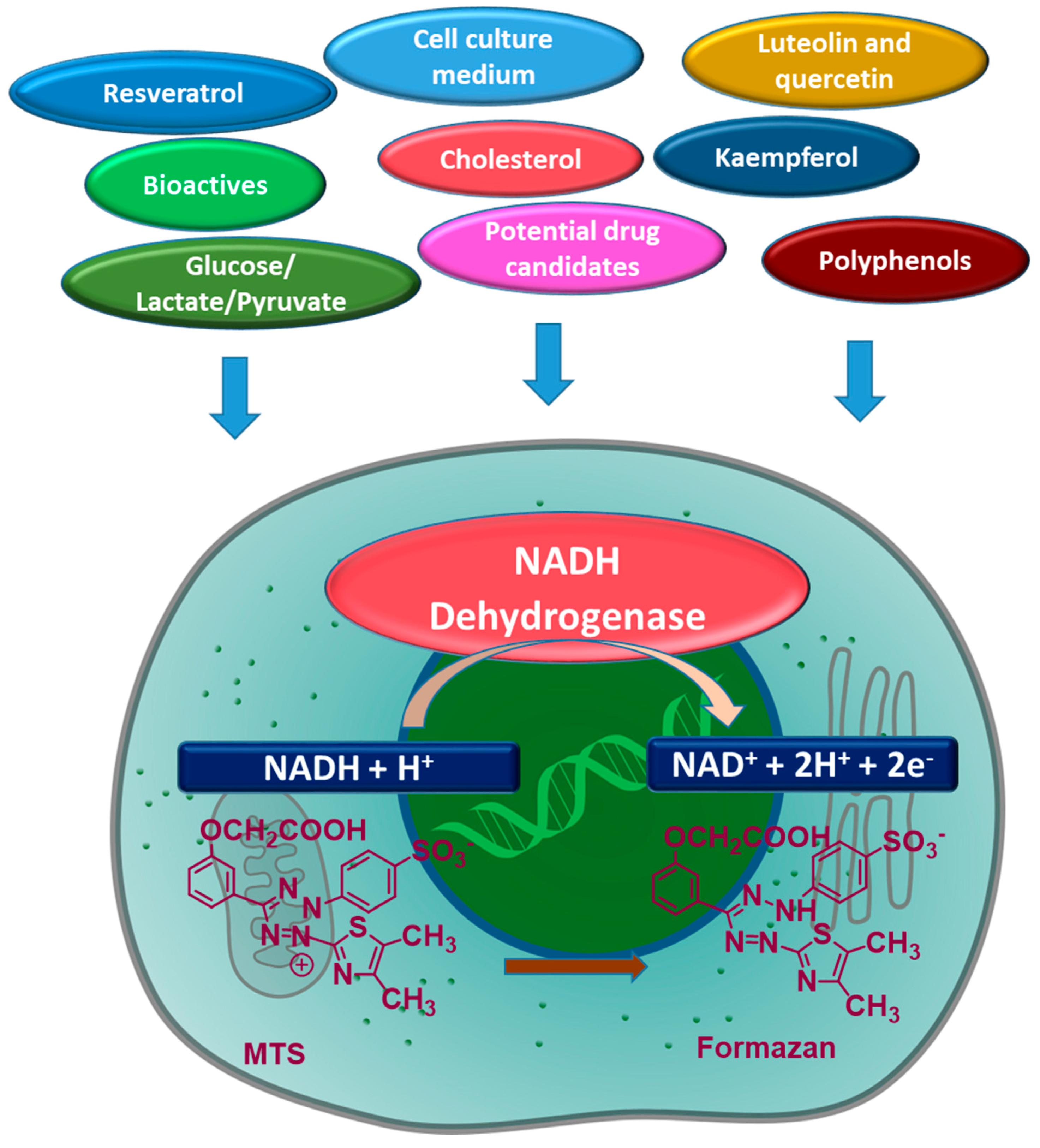
T. ferdinandiana fruits are naturally rich in antioxidants like ellagic acid and vitamin C, and based on the outcomes of this study, can directly bio-reduce cell viability assays dependent upon the activity of dehydrogenase enzymes. In support of the findings presented here, other studies report the direct reduction of MTS to formazan by phenolic antioxidants (
Figure 4), including kaempferol and ellagic acid, in the absence of cells [
2,
3]. The increased cell viability observed in the CellTiter 96
® AQ
ueous One Solution Cell Proliferation Assay after treating cells with
T. ferdinandiana fruit extract can be explained by the presence of redox active antioxidants, such as vitamin C [
9]. Other studies involving plant extracts containing vitamin E, luteolin, quercetin, flavones, flavonones, alkaloids, glycosides, and epigallocatechin-3-gallate (
Figure 4) have also reported direct interaction with MTT in cell-free systems [
2]. The MTT assay, and to a lesser degree MTS, and other related assays like XTT (2,3-bis-(2-methoxy-4-nitro-5-sulfophenyl)-2
H-tetrazolium-5-carboxanilide) or WST (water soluble tetrazolium salts), are a widely exploited approach for measuring cell viability in response to different targets such as drugs.
The reduction of MTT, however, can occur through many different mechanisms, ultimately impacting cell viability results. For example, intracellular MTT reduction can occur through NADH and NADPH by oxidoreductases or superoxides, whereas extracellular reduction can occur through NADH and NADPH by cell surface oxidoreductases. Mitochondrial and non-mitochondrial enzymes, endoplasmic reticulum, cytosol, and plasma membranes can also participate in reducing MTT [
10]. Factors that influence intracellular trafficking of MTT-formazan, such as disturbances in cellular metabolic and energy homeostasis, rate of endo- or exocytosis, oxidative stress, glucose uptake and glycolysis, lactate and pyruvate levels, and formazan, can also impact MTT reduction [
10]. Off-target effects of the investigated molecules, such as potential drug candidates and other bioactives, can also cause under or over-estimation of cell viability [
2]. Cell culture medium composition, cell growth state, concentration and consumption rate of energy supply metabolites, nanoparticles, polypeptides, and X-ray radiation may also impact MTT reduction and measured cell viability [
2].
All cell viability assays offer advantages and limitations; for example, the MTT/MTS assays are easy to perform but variable metabolic behaviour can be observed under different cell culture conditions and the solubility of the insoluble formazan products can sometimes be problematic, whereas WSTs are water soluble and easy one-step assays. However, variable metabolic behaviour under different cell culture conditions can also be observed with WSTs. DNA assays such as CyQUANT
® can be directly correlated with proliferation, however, black plates are needed and the assay requires RNase treatment for the detection of DNA and vice versa [
11].
In this study, methanolic and water extracts of
T. ferdinandiana fruits were incubated with MTS (
Figure 1A,B) in a cell-free setting, producing direct MTS reduction after a 2-h incubation period, whereas positive and negative controls did not produce formazan. Based on the findings presented here and by others, different variables, such as the pH and osmolarity of cell culture media, positive and negative control substances, interactions with MTS in a cell-free system, and washing steps, are important considerations when performing cell viability assays. The application of the CellTiter 96
® AQ
ueous One Solution Cell Proliferation Assay as a screening system for plant extracts requires pre-screening in cell-free systems to determine whether the extracts exhibit a reductive capacity that might prevent optimum performance of this assay. Cell washing steps can be added to reduce the potential interference by reductive extracts, however, washing cannot prevent the impact of redox active extracts on intracellular redox status that will also impact the cell viability measured through MTS or MTT.

 ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}



