Suppression of Light-Induced Oxidative Stress in the Retina by Mitochondria-Targeted Antioxidant


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Animals and Ethics Statement
2.3. Experimental Model
2.4. Measurements of Outer Nuclear Layer Thickness
2.5. Retinal Samples
2.6. Total Protein Concentration
2.7. Malondialdehyde Concentration
2.8. Hydrogen Peroxide Concentration
2.9. Total Antioxidant Activity
2.10. Antioxidant Enzymes Activity
2.11. Content of Disulfide Dimers of Arrestin
2.12. Statistical Analysis
3. Results
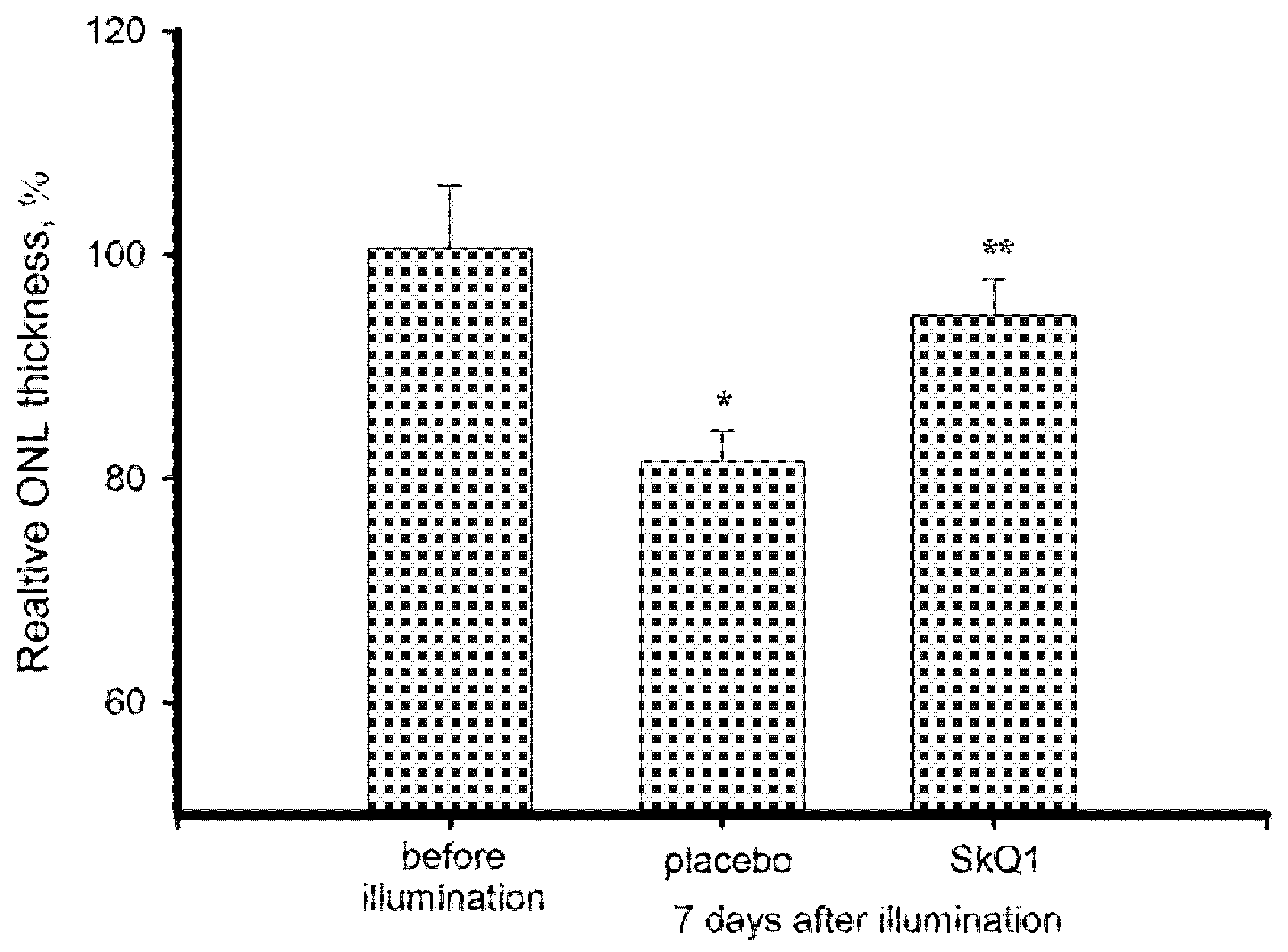
3.1. Morphological State of Bright Light-Exposed Retina: Thickness of Outer Nuclear Layer
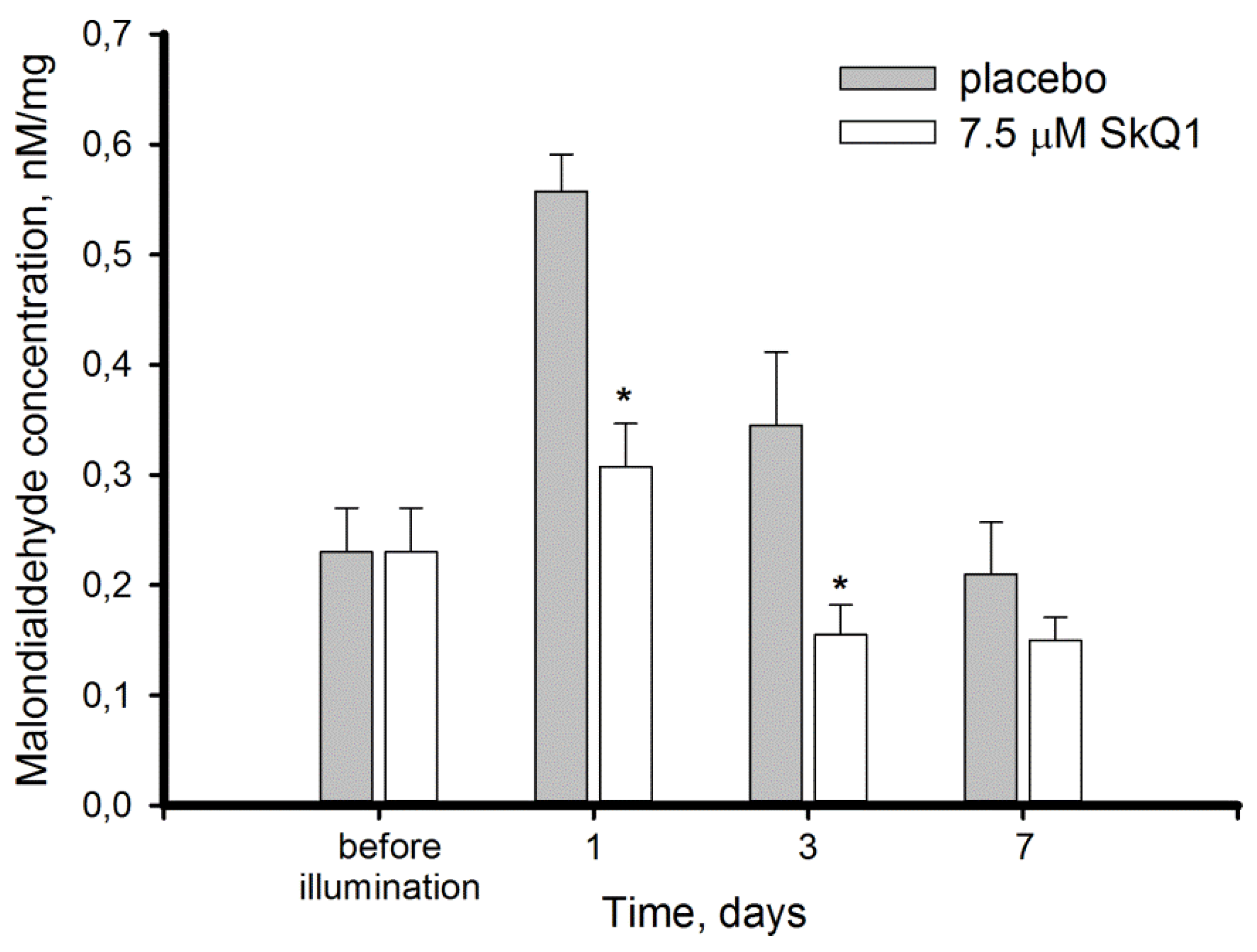
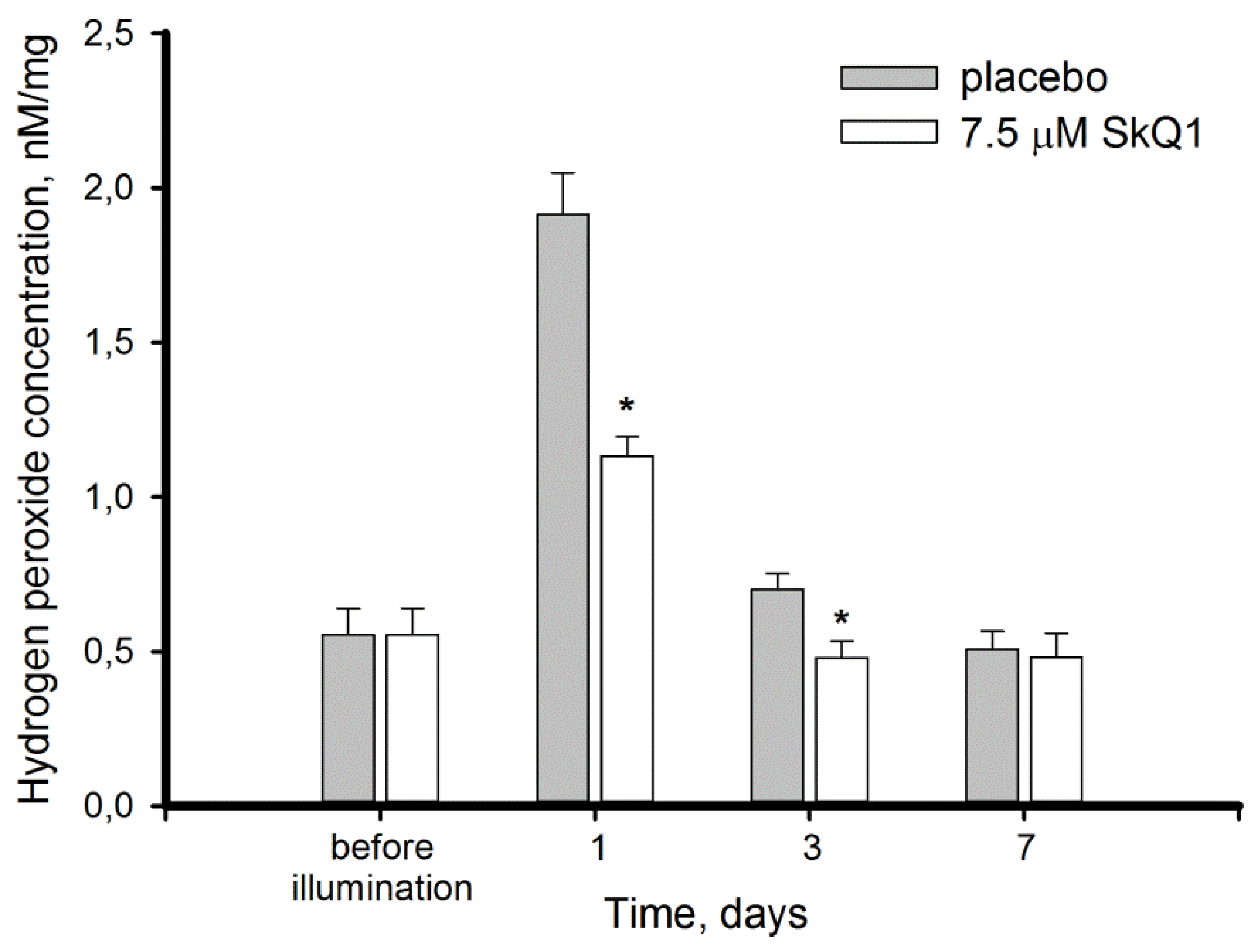
3.2. Light-Induced Oxidative Stress in the Retina: Lipid Peroxidation and Hydrogen Peroxide Accumulation
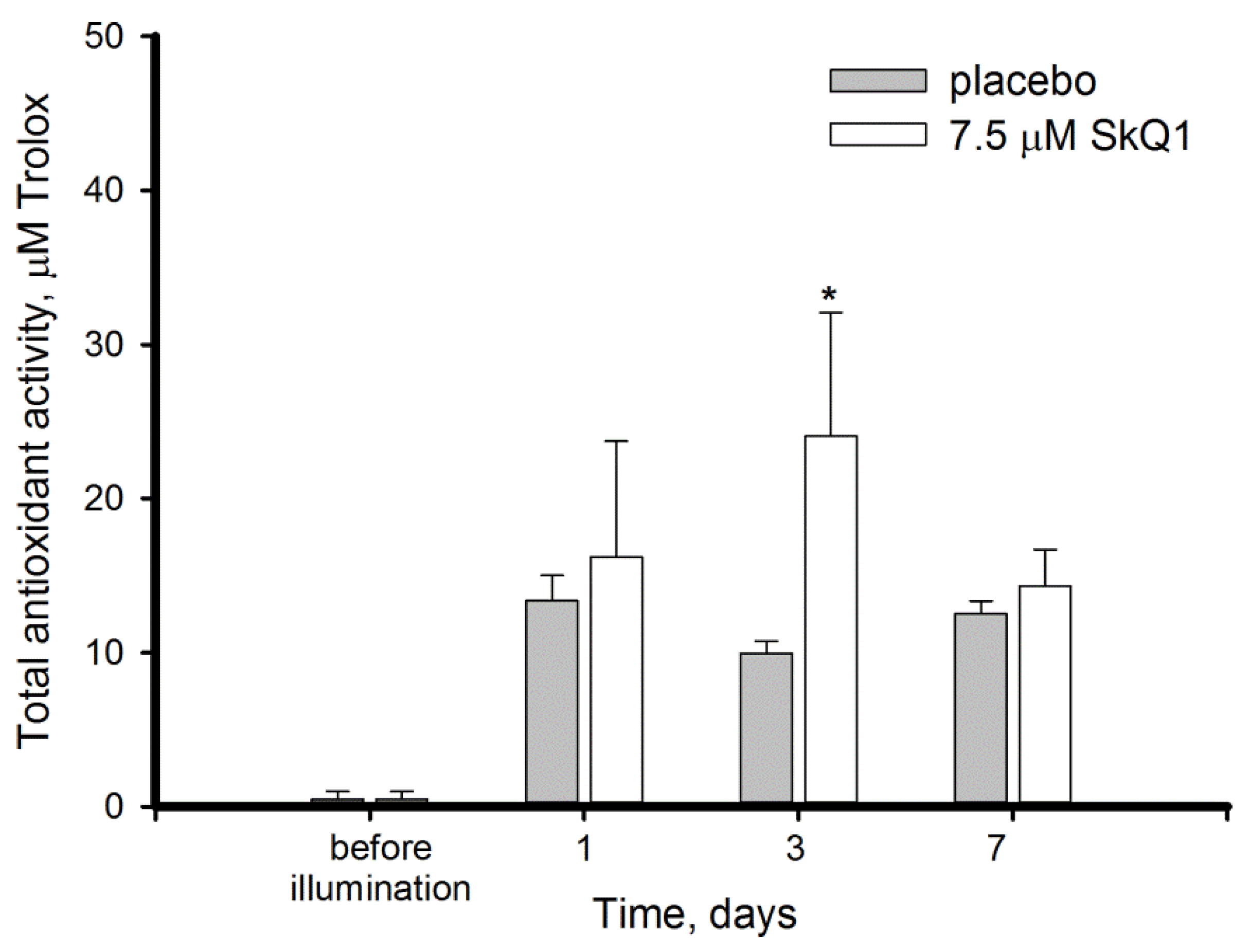
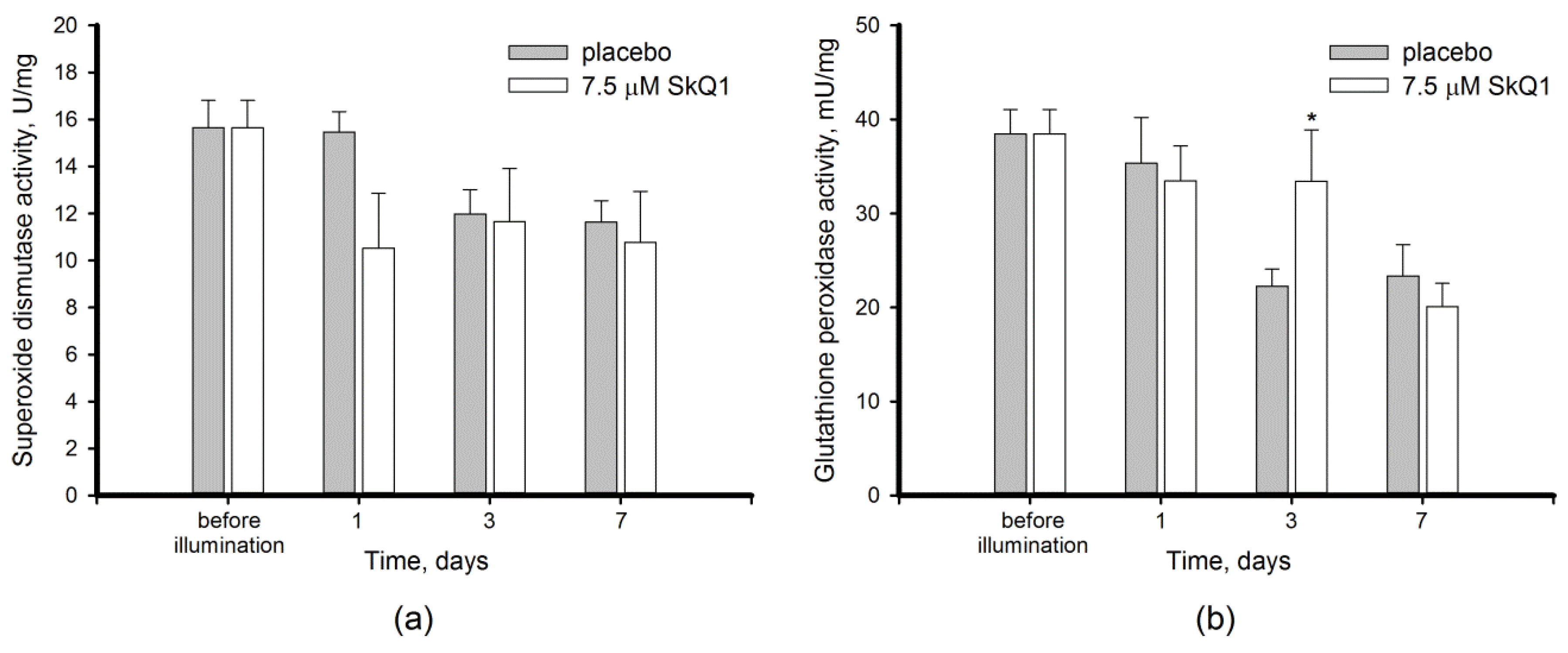
3.3. Antioxidant Activity in the Bright Light-Exposed Retina: Total Antioxidant Activity and Antioxidant Enzyme Functioning
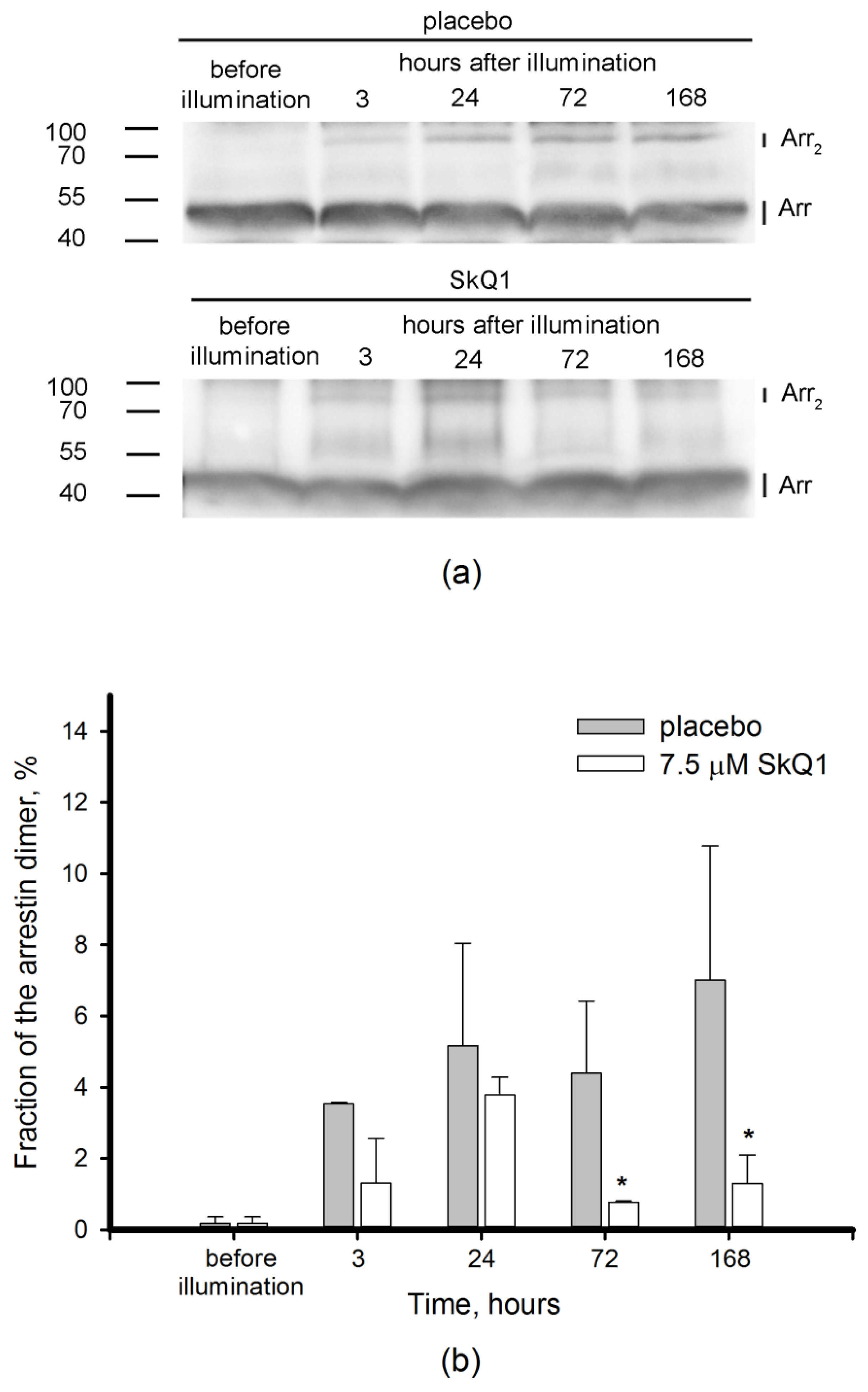
3.4. Redox-Sensitive Proteins in Light-Induced Oxidative Stress: Visual Arrestin
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AMD | Age-related macular degeneration |
| RPE | Retinal pigment epithelium |
| ROS | Reactive oxygen species |
| LIRD | Light-induced retinal damage |
| SkQ1 | 10-(6′-plastoquinonyl)-decyltriphenylphosphonium |
| AOA | Total antioxidant activity |
| SOD | Superoxide dismutase |
| GPx | Glutathione peroxidase |
| PBS | Phosphate buffer saline |
References
- Rajala, R.V.; Rajala, A.; Kooker, C.; Wang, Y.; Anderson, R.E. The Warburg effect mediator pyruvate kinase M2 expression and regulation in the retina. Sci. Rep. 2016, 6, 37727. [Google Scholar] [CrossRef] [PubMed]
- Beatty, S.; Koh, H.-H.; Phil, M.; Henson, D.; Boulton, M. The role of oxidative stress in the pathogenesis of age-related macular degeneration. Surv. Ophthalmol. 2000, 45, 115–134. [Google Scholar] [CrossRef]
- Miceli, M.V.; Liles, M.R.; Newsome, D.A. Evaluation of oxidative processes in human pigment epithelial cells associated with retinal outer segment phagocytosis. Exp. Cell Res. 1994, 214, 242–249. [Google Scholar] [CrossRef] [PubMed]
- Liles, M.R.; Newsome, D.A.; Oliver, P.D. Antioxidant enzymes in the aging human retinal pigment epithelium. Arch. Ophthalmol. 1991, 109, 1285–1288. [Google Scholar] [CrossRef] [PubMed]
- Tate, D.; Newsome, D.; Oliver, P. Metallothionein shows an age-related decrease in human macular retinal pigment epithelium. Investig. Ophthalmol. Vis. Sci. 1993, 34, 2348–2351. [Google Scholar]
- Zernii, E.Y.; Baksheeva, V.E.; Iomdina, E.N.; Averina, O.A.; Permyakov, S.E.; Philippov, P.; Zamyatnin, A.; Senin, I. Rabbit models of ocular diseases: New relevance for classical approaches. CNS Neurol. Disord. Drug Targets 2016, 15, 267–291. [Google Scholar] [CrossRef] [PubMed]
- Van Norren, D.; Vos, J.J. Light damage to the retina: An historical approach. Eye 2016, 30, 169–172. [Google Scholar] [CrossRef]
- Organisciak, D.T.; Vaughan, D.K. Retinal light damage: Mechanisms and protection. Prog. Retin. Eye Res. 2010, 29, 113–134. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez Martin-Moro, J.; Hernandez Verdejo, J.L.; Zarallo Gallardo, J. Photic maculopathy: A review of the literature (I). Arch. Soc. Espan. Oftalmol. 2018. [CrossRef]
- Gonzalez Martin-Moro, J.; Hernandez Verdejo, J.L.; Zarallo Gallardo, J. Photic maculopathy: A review of the literature (II). Arch. Soc. Espan. Oftalmol. 2018. [CrossRef]
- Marquioni-Ramella, M.D.; Suburo, A.M. Photo-damage, photo-protection and age-related macular degeneration. Photochem. Photobiol. Sci. 2015, 14, 1560–1577. [Google Scholar] [CrossRef] [PubMed]
- Boulton, M.; Rozanowska, M.; Rozanowski, B. Retinal photodamage. J. Photochem. Photobiol. B Biol. 2001, 64, 144–161. [Google Scholar] [CrossRef]
- Organisciak, D.T.; Darrow, R.M.; Barsalou, L.; Darrow, R.A.; Kutty, R.K.; Kutty, G.; Wiggert, B. Light history and age-related changes in retinal light damage. Investig. Ophthalmol. Vis. Sci. 1998, 39, 1107–1116. [Google Scholar]
- Marc, R.E.; Jones, B.; Watt, C.; Vazquez-Chona, F.; Vaughan, D.; Organisciak, D. Extreme retinal remodeling triggered by light damage: Implications for age related macular degeneration. Mol. Vis. 2008, 14, 782–806. [Google Scholar]
- Cousins, S.W.; Espinosa-Heidmann, D.G.; Alexandridou, A.; Sall, J.; Dubovy, S.; Csaky, K. The role of aging, high fat diet and blue light exposure in an experimental mouse model for basal laminar deposit formation. Exp. Eye Res. 2002, 75, 543–553. [Google Scholar] [CrossRef] [PubMed]
- Espinosa-Heidmann, D.G.; Sall, J.; Hernandez, E.P.; Cousins, S.W. Basal laminar deposit formation in APO B100 transgenic mice: Complex interactions between dietary fat, blue light, and vitamin E. Investig. Ophthalmol. Vis. Sci. 2004, 45, 260–266. [Google Scholar] [CrossRef]
- Wielgus, A.; Collier, R.; Martin, E.; Lih, F.; Tomer, K.; Chignell, C.; Roberts, J. Blue light induced A2E oxidation in rat eyes–experimental animal model of dry AMD. Photochem. Photobiol. Sci. 2010, 9, 1505–1512. [Google Scholar] [CrossRef]
- Albert, D.M.; Neekhra, A.; Wang, S.; Darjatmoko, S.R.; Sorenson, C.M.; Dubielzig, R.R.; Sheibani, N. Development of choroidal neovascularization in rats with advanced intense cyclic light-induced retinal degeneration. Arch. Ophthalmol. 2010, 128, 212–222. [Google Scholar]
- Novikova, Y.P.; Gancharova, O.S.; Eichler, O.; Philippov, P.; Grigoryan, E. Preventive and therapeutic effects of SkQ1-containing Visomitin eye drops against light-induced retinal degeneration. Biochemistry 2014, 79, 1101–1110. [Google Scholar] [CrossRef]
- Zernii, E.Y.; Nazipova, A.A.; Gancharova, O.S.; Kazakov, A.S.; Serebryakova, M.V.; Zinchenko, D.V.; Tikhomirova, N.K.; Senin, I.I.; Philippov, P.P.; Permyakov, E.A. Light-induced disulfide dimerization of recoverin under ex vivo and in vivo conditions. Free Radic. Biol. Med. 2015, 83, 283–295. [Google Scholar] [CrossRef]
- Wolffe, M. How safe is the light during ophthalmic diagnosis and surgery. Eye 2016, 30, 186–188. [Google Scholar] [CrossRef] [PubMed]
- Hunter, J.J.; Morgan, J.I.W.; Merigan, W.H.; Sliney, D.H.; Sparrow, J.R.; Williams, D.R. The susceptibility of the retina to photochemical damage from visible light. Prog. Retin. Eye Res. 2012, 31, 28–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, J.; Seregard, S.; Spångberg, B.; Oskarsson, M.; Chen, E. Blue light induced apoptosis in rat retina. Eye 1999, 13, 577–583. [Google Scholar] [CrossRef] [PubMed]
- Donovan, M.; Carmody, R.J.; Cotter, T.G. Light-induced photoreceptor apoptosis in vivo requires neuronal nitric-oxide synthase and guanylate cyclase activity and is caspase-3-independent. J. Biol. Chem. 2001, 276, 23000–23008. [Google Scholar] [CrossRef] [PubMed]
- Maeda, A.; Maeda, T.; Golczak, M.; Chou, S.; Desai, A.; Hoppel, C.L.; Matsuyama, S.; Palczewski, K. Involvement of all-trans-retinal in acute light-induced retinopathy of mice. J. Biol. Chem. 2009, 284, 15173–15183. [Google Scholar] [CrossRef] [PubMed]
- Liang, F.-Q.; Godley, B.F. Oxidative stress-induced mitochondrial DNA damage in human retinal pigment epithelial cells: A possible mechanism for RPE aging and age-related macular degeneration. Exp. Eye Res. 2003, 76, 397–403. [Google Scholar] [CrossRef]
- Bhagavan, H.N.; Chopra, R.K. Plasma coenzyme Q10 response to oral ingestion of coenzyme Q10 formulations. Mitochondrion 2007, 7, S78–S88. [Google Scholar] [CrossRef]
- Manach, C.; Williamson, G.; Morand, C.; Scalbert, A.; Remesy, C. Bioavailability and bioefficacy of polyphenols in humans. I. Review of 97 bioavailability studies. Am. J. Clin. Nutr. 2005, 81, 230S–242S. [Google Scholar] [CrossRef] [Green Version]
- Gueven, N.; Woolley, K.; Smith, J. Border between natural product and drug: Comparison of the related benzoquinones idebenone and coenzyme Q10. Redox Biol. 2015, 4, 289–295. [Google Scholar] [CrossRef]
- Fletcher, A.E. Free radicals, antioxidants and eye diseases: Evidence from epidemiological studies on cataract and age-related macular degeneration. Ophthalmic Res. 2010, 44, 191–198. [Google Scholar] [CrossRef]
- Antonenko, Y.N.; Avetisyan, A.; Bakeeva, L.; Chernyak, B.; Chertkov, V.; Domnina, L.; Ivanova, O.Y.; Izyumov, D.; Khailova, L.; Klishin, S. Mitochondria-targeted plastoquinone derivatives as tools to interrupt execution of the aging program. 1. Cationic plastoquinone derivatives: Synthesis and in vitro studies. Biochemistry 2008, 73, 1273–1287. [Google Scholar] [CrossRef] [PubMed]
- Neroev, V.; Archipova, M.; Bakeeva, L.; Fursova, A.Z.; Grigorian, E.; Grishanova, A.Y.; Iomdina, E.; Ivashchenko, Z.N.; Katargina, L.; Khoroshilova-Maslova, I. Mitochondria-targeted plastoquinone derivatives as tools to interrupt execution of the aging program. 4. Age-related eye disease. SkQ1 returns vision to blind animals. Biochemistry 2008, 73, 1317–1328. [Google Scholar] [CrossRef] [PubMed]
- Brzheskiy, V.V.; Efimova, E.L.; Vorontsova, T.N.; Alekseev, V.N.; Gusarevich, O.G.; Shaidurova, K.N.; Ryabtseva, A.A.; Andryukhina, O.M.; Kamenskikh, T.G.; Sumarokova, E.S.; et al. Results of a Multicenter, Randomized, Double-Masked, Placebo-Controlled Clinical Study of the Efficacy and Safety of Visomitin Eye Drops in Patients with Dry Eye Syndrome. Adv. Ther. 2015, 32, 1263–1279. [Google Scholar] [CrossRef] [PubMed]
- Zernii, E.Y.; Gancharova, O.S.; Baksheeva, V.E.; Golovastova, M.O.; Kabanova, E.I.; Savchenko, M.S.; Tiulina, V.V.; Sotnikova, L.F.; Zamyatnin, A.A.; Philippov, P.P. Mitochondria-Targeted Antioxidant SkQ1 Prevents Anesthesia-Induced Dry Eye Syndrome. Oxid. Med. Cell. Longev. 2017, 2017, 9281519. [Google Scholar] [CrossRef] [PubMed]
- Baldin, A.V.; Grishina, A.N.; Korolev, D.O.; Kuznetsova, E.B.; Golovastova, M.O.; Kalpinskiy, A.S.; Alekseev, B.Y.; Kaprin, A.D.; Zinchenko, D.V.; Savvateeva, L.V.; et al. Autoantibody against arrestin-1 as a potential biomarker of renal cell carcinoma. Biochimie 2018, 157, 26–37. [Google Scholar] [CrossRef] [PubMed]
- Erel, O. A new automated colorimetric method for measuring total oxidant status. Clin. Biochem. 2005, 38, 1103–1111. [Google Scholar] [CrossRef] [PubMed]
- Gulidova, O.; Liubitskiĭ, O.; Klebanov, G.; Chesnokova, N. Changes in the antioxidative activity of tears during experimental eye burns. Biull. Eksp. Biol. Med. 1999, 128, 571–574. [Google Scholar] [CrossRef]
- Zernii, E.Y.; Golovastova, M.O.; Baksheeva, V.E.; Kabanova, E.I.; Ishutina, I.E.; Gancharova, O.S.; Gusev, A.E.; Savchenko, M.S.; Loboda, A.P.; Sotnikova, L.F.; et al. Alterations in Tear Biochemistry Associated with Postanesthetic Chronic Dry Eye Syndrome. Biochemistry 2016, 81, 1549–1557. [Google Scholar] [CrossRef]
- Zernii, E.Y.; Baksheeva, V.E.; Kabanova, E.I.; Tiulina, V.V.; Golovastova, M.O.; Gancharova, O.S.; Savchenko, M.S.; Sotikova, L.F.; Zamyatnin, A.A., Jr.; Filippov, P.P.; et al. Effect of General Anesthesia Duration on Recovery of Secretion and Biochemical Properties of Tear Fluid in the Post-Anesthetic Period. Bull. Exp. Biol. Med. 2018, 165, 269–271. [Google Scholar] [CrossRef]
- GelAnalyzer 2010a. Available online: http://gelanalyzer.com/ (accessed on 21 October 2018).
- Lieven, C.J.; Ribich, J.D.; Crowe, M.E.; Levin, L.A. Redox Proteomic Identification of Visual Arrestin Dimerization in Photoreceptor Degeneration after Photic Injury. Investig. Ophthalmol. Vis. Sci. 2012, 53, 3990–3998. [Google Scholar] [CrossRef] [Green Version]
- Xie, Z.; Wu, X.; Gong, Y.; Song, Y.; Qiu, Q.; Li, C. Intraperitoneal injection of Ginkgo biloba extract enhances antioxidation ability of retina and protects photoreceptors after light-induced retinal damage in rats. Curr. Eye Res. 2007, 32, 471–479. [Google Scholar] [CrossRef] [PubMed]
- Saenz-de-Viteri, M.; Heras-Mulero, H.; Fernandez-Robredo, P.; Recalde, S.; Hernandez, M.; Reiter, N.; Moreno-Orduna, M.; Garcia-Layana, A. Oxidative stress and histological changes in a model of retinal phototoxicity in rabbits. Oxid. Med. Cell. Longev. 2014, 2014, 637137. [Google Scholar] [CrossRef]
- Rózanowska, M.; Jarvis-Evans, J.; Korytowski, W.; Boulton, M.E.; Burke, J.M.; Sarna, T. Blue light-induced reactivity of retinal age pigment in vitro generation of oxygen-reactive species. J. Biol. Chem. 1995, 270, 18825–18830. [Google Scholar] [CrossRef] [PubMed]
- Catalá, A. An overview of lipid peroxidation with emphasis in outer segments of photoreceptors and the chemiluminescence assay. Int. J. Biochem. Cell Biol. 2006, 38, 1482–1495. [Google Scholar] [CrossRef] [PubMed]
- Daemen, F.J. Vertebrate rod outer segment membranes. Biochim. Biophys. Acta 1973, 300, 255–288. [Google Scholar] [CrossRef]
- Fine, S.L.; Berger, J.W.; Maguire, M.G.; Ho, A.C. Age-related macular degeneration. N. Engl. J. Med. 2000, 342, 483–492. [Google Scholar] [CrossRef] [PubMed]
- Weismann, D.; Hartvigsen, K.; Lauer, N.; Bennett, K.L.; Scholl, H.P.; Charbel Issa, P.; Cano, M.; Brandstatter, H.; Tsimikas, S.; Skerka, C.; et al. Complement factor H binds malondialdehyde epitopes and protects from oxidative stress. Nature 2011, 478, 76–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crabb, J.W.; Miyagi, M.; Gu, X.; Shadrach, K.; West, K.A.; Sakaguchi, H.; Kamei, M.; Hasan, A.; Yan, L.; Rayborn, M.E. Drusen proteome analysis: An approach to the etiology of age-related macular degeneration. Proc. Natl. Acad. Sci. USA 2002, 99, 14682–14687. [Google Scholar] [CrossRef] [Green Version]
- Hollyfield, J.G.; Salomon, R.G.; Crabb, J.W.; Gu, X. Diagnostic Methods for Age Related Macular Degeneration. U.S. Patent 7172874B2, 6 February 2007. [Google Scholar]
- Yamashita, H.; Horie, K.; Yamamoto, T.; Nagano, T.; Hirano, T. Light-induced retinal damage in mice. Hydrogen peroxide production and superoxide dismutase activity in retina. Retina 1992, 12, 59–66. [Google Scholar] [CrossRef]
- Lam, S.; Tso, M.O.; Gurne, D.H. Amelioration of retinal photic injury in albino rats by dimethylthiourea. Arch. Ophthalmol. 1990, 108, 1751–1757. [Google Scholar] [CrossRef]
- Winkler, B.S.; Boulton, M.E.; Gottsch, J.D.; Sternberg, P. Oxidative damage and age-related macular degeneration. Mol. Vis. 1999, 5, 32. [Google Scholar] [PubMed]
- Jin, G.-F.; Hurst, J.S.; Godley, B.F. Rod outer segments mediate mitochondrial DNA damage and apoptosis in human retinal pigment epithelium. Curr. Eye Res. 2001, 23, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Jarrett, S.G.; Boulton, M.E. Consequences of oxidative stress in age-related macular degeneration. Mol. Aspects Med. 2012, 33, 399–417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tokarz, P.; Kaarniranta, K.; Blasiak, J. Role of antioxidant enzymes and small molecular weight antioxidants in the pathogenesis of age-related macular degeneration (AMD). Biogerontology 2013, 14, 461–482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trevithick, J.R.; Dzialoszynski, T. Endogenous superoxide-like species and antioxidant activity in ocular tissues detected by luminol luminescence. IUBMB Life 1997, 41, 695–705. [Google Scholar] [CrossRef]
- Penn, J.S.; Naash, M.I.; Anderson, R.E. Effect of light history on retinal antioxidants and light damage susceptibility in the rat. Exp. Eye Res. 1987, 44, 779–788. [Google Scholar] [CrossRef]
- Hiramitsu, T.; Ohshiro, M. Effects of superoxide dismutase and catalase on UV-light induced retina lipid peroxidation. J. Eye 1988, 5, 1024–1026. [Google Scholar]
- Ohira, A.; Tanito, M.; Kaidzu, S.; Kondo, T. Glutathione peroxidase induced in rat retinas to counteract photic injury. Investig. Ophthalmol. Vis. Sci. 2003, 44, 1230–1236. [Google Scholar] [CrossRef]
- Calvert, P.D.; Strissel, K.J.; Schiesser, W.E.; Pugh, E.N., Jr.; Arshavsky, V.Y. Light-driven translocation of signaling proteins in vertebrate photoreceptors. Trends Cell Biol. 2006, 16, 560–568. [Google Scholar] [CrossRef]
- Chen, Y.; Perusek, L.; Maeda, A. Autophagy in light-induced retinal damage. Exp. Eye Res. 2016, 144, 64–72. [Google Scholar] [CrossRef] [Green Version]
- Hall, M.O.; Hall, D.O. Superoxide-Dismutase of Bovine and Frog Rod Outer Segments. Biochem. Biophys. Res. Commun. 1975, 67, 1199–1204. [Google Scholar] [CrossRef]
- Trujillo, M.; Alvarez, B.; Radi, R. One- and two-electron oxidation of thiols: Mechanisms, kinetics and biological fates. Free Radic. Res. 2016, 50, 150–171. [Google Scholar] [CrossRef] [PubMed]
- Davies, M.J. Protein oxidation and peroxidation. Biochem. J. 2016, 473, 805–825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Escribano-Lopez, I.; Diaz-Morales, N.; Rovira-Llopis, S.; de Maranon, A.M.; Orden, S.; Alvarez, A.; Banuls, C.; Rocha, M.; Murphy, M.P.; Hernandez-Mijares, A.; et al. The mitochondria-targeted antioxidant MitoQ modulates oxidative stress, inflammation and leukocyte-endothelium interactions in leukocytes isolated from type 2 diabetic patients. Redox Biol. 2016, 10, 200–205. [Google Scholar] [CrossRef]
- Gurevich, V.V.; Hanson, S.M.; Song, X.; Vishnivetskiy, S.A.; Gurevich, E.V. The functional cycle of visual arrestins in photoreceptor cells. Prog. Retin. Eye Res. 2011, 30, 405–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koch, K.W.; Dell’Orco, D. Protein and Signaling Networks in Vertebrate Photoreceptor Cells. Front. Mol. Neurosci. 2015, 8, 67. [Google Scholar] [CrossRef] [PubMed]
- Kiser, P.D.; Golczak, M.; Maeda, A.; Palczewski, K. Key enzymes of the retinoid (visual) cycle in vertebrate retina. Biochim. Biophys. Acta 2012, 1821, 137–151. [Google Scholar] [CrossRef] [Green Version]
- Adler, L.T.; Boyer, N.P.; Chen, C.; Ablonczy, Z.; Crouch, R.K.; Koutalos, Y. The 11-cis Retinal Origins of Lipofuscin in the Retina. Prog. Mol. Biol. Transl. Sci. 2015, 134, e1–e12. [Google Scholar] [CrossRef]
- Maeda, T.; Golczak, M.; Maeda, A. Retinal photodamage mediated by all-trans-retinal. Photochem. Photobiol. 2012, 88, 1309–1319. [Google Scholar] [CrossRef]
- Chen, J.; Simon, M.I.; Matthes, M.T.; Yasumura, D.; LaVail, M.M. Increased susceptibility to light damage in an arrestin knockout mouse model of Oguchi disease (stationary night blindness). Invest. Ophthalmol. Vis. Sci. 1999, 40, 2978–2982. [Google Scholar]
- Wang, A.L.; Lukas, T.J.; Yuan, M.; Du, N.; Tso, M.O.; Neufeld, A.H. Autophagy and exosomes in the aged retinal pigment epithelium: Possible relevance to drusen formation and age-related macular degeneration. PLoS ONE 2009, 4, e4160. [Google Scholar] [CrossRef] [PubMed]






© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baksheeva, V.E.; Tiulina, V.V.; Tikhomirova, N.K.; Gancharova, O.S.; Komarov, S.V.; Philippov, P.P.; Zamyatnin, A.A., Jr.; Senin, I.I.; Zernii, E.Y. Suppression of Light-Induced Oxidative Stress in the Retina by Mitochondria-Targeted Antioxidant. Antioxidants 2019, 8, 3. https://doi.org/10.3390/antiox8010003
Baksheeva VE, Tiulina VV, Tikhomirova NK, Gancharova OS, Komarov SV, Philippov PP, Zamyatnin AA Jr., Senin II, Zernii EY. Suppression of Light-Induced Oxidative Stress in the Retina by Mitochondria-Targeted Antioxidant. Antioxidants. 2019; 8(1):3. https://doi.org/10.3390/antiox8010003
Chicago/Turabian StyleBaksheeva, Viktoriia E., Veronika V. Tiulina, Natalia K. Tikhomirova, Olga S. Gancharova, Sergey V. Komarov, Pavel P. Philippov, Andrey A. Zamyatnin, Jr., Ivan I. Senin, and Evgeni Yu. Zernii. 2019. "Suppression of Light-Induced Oxidative Stress in the Retina by Mitochondria-Targeted Antioxidant" Antioxidants 8, no. 1: 3. https://doi.org/10.3390/antiox8010003
APA StyleBaksheeva, V. E., Tiulina, V. V., Tikhomirova, N. K., Gancharova, O. S., Komarov, S. V., Philippov, P. P., Zamyatnin, A. A., Jr., Senin, I. I., & Zernii, E. Y. (2019). Suppression of Light-Induced Oxidative Stress in the Retina by Mitochondria-Targeted Antioxidant. Antioxidants, 8(1), 3. https://doi.org/10.3390/antiox8010003