
Mulberry Leaf Polysaccharides Attenuate Oxidative Stress Injury in Peripheral Blood Leukocytes by Regulating Endoplasmic Reticulum Stress



Abstract
1. Introduction
2. Materials and Methods
2.1. MLPs
2.1.1. Determination of Molecular Weights
2.1.2. Monosaccharide Composition Analysis
2.2. PBL Isolation
2.3. Experimental Treatment and Sampling
2.4. Chemical Analyses
2.4.1. Determination of PBLs’ Antioxidant Capacity
2.4.2. Real-Time PCR Analysis (qRT-PCR)
2.5. Statistical Analyses
3. Results
3.1. Monosaccharide Composition and Molecular Weights of MLPs
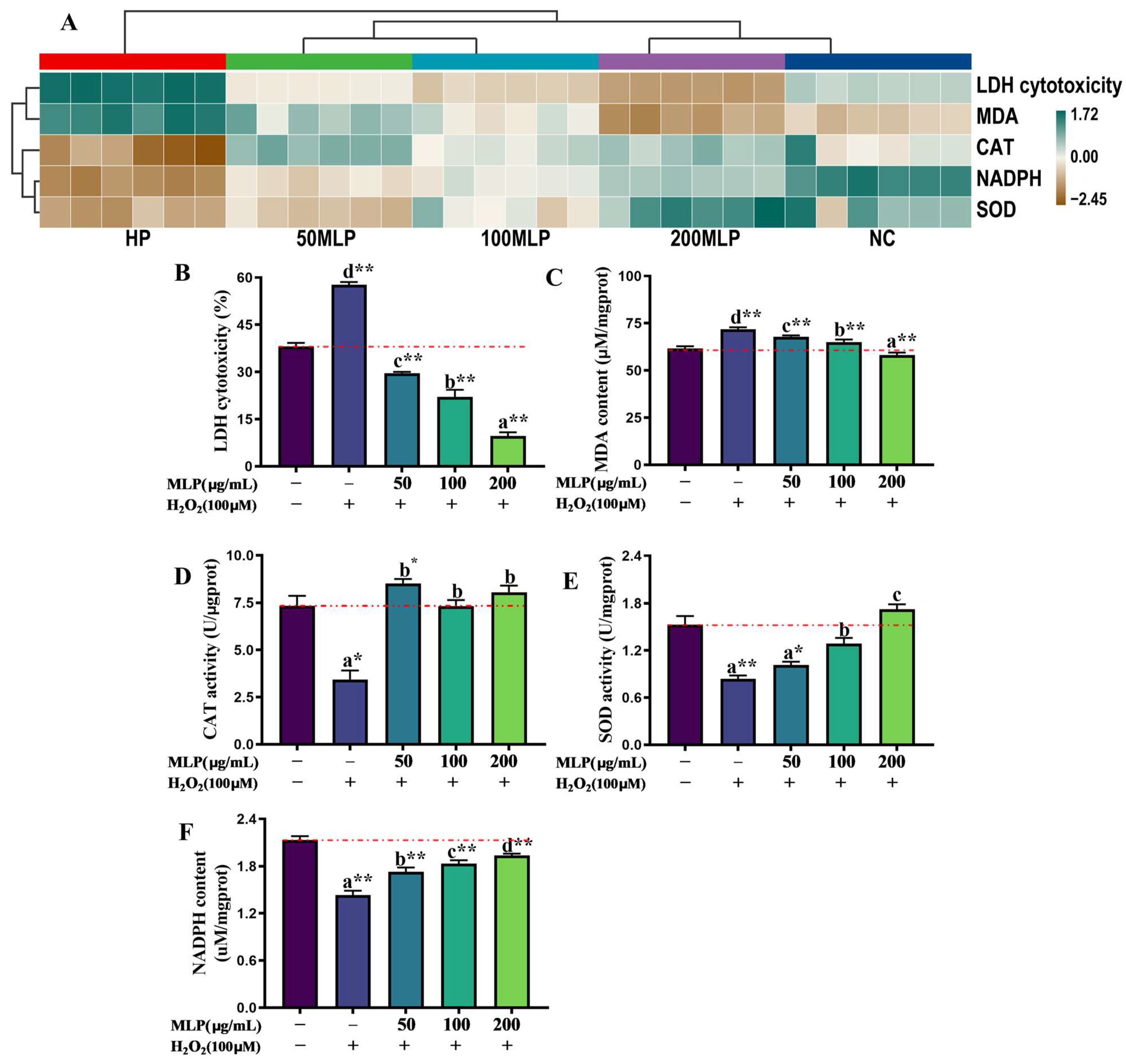
3.2. Effect of MLPs on the Antioxidant Capacity of PBLs Exposed to H2O2
3.3. Assessment of ROS Generation and the MMP Status
3.4. Assessment of Gene Expression
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Huang, K.; Jiang, L.; Huang, W.; Li, X.; Yuan, L.; Jiang, J.; Zhou, S.; Wang, Y.; Xie, J. Investigating the interplay between Cryptocaryon irritans ectoparasite infection and the immune responses of the head kidney in silver pomfret (Pampus argenteus). Aquaculture 2023, 575, 739780. [Google Scholar] [CrossRef]
- Giri, S.S.; Kim, S.G.; Woo, K.J.; Jung, W.J.; Lee, S.B.; Lee, Y.M.; Jo, S.J.; Kim, J.H.; Park, S.C. Impact of dandelion polysaccharides on growth and immunity response in common carp Cyprinus carpio. Fish Shellfish Immunol. 2022, 128, 371–379. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wu, Z.; Chen, H.; Liu, R.; Zhang, W.; Chen, X. Astragalus polysaccharides protect against inactivated Vibrio alginolyticus-induced inflammatory injury in macrophages of large yellow croaker. Fish Shellfish Immunol. 2022, 131, 95–104. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Geng, C.; Qu, Y.K.; Cheng, B.X.; Zhang, Y.; Wang, A.M.; Zhang, J.H.; Liu, B.; Tian, H.Y.; Yang, W.P. The feeding of dietary Codonopsis pilosula polysaccharide enhances the immune responses, the expression of immune-related genes and the growth performance of red swamp crayfish (Procambarus clarkii). Fish Shellfish Immunol. 2020, 103, 321–331. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Fang, J.; Ruan, Y.; Wang, X.; Sun, Y.; Wu, N.; Zhao, Z.; Chang, Y.; Ning, N.; Guo, H. Structures, bioactivities and future prospective of polysaccharides from Morus alba (white mulberry): A review. Food Chem. 2018, 245, 899–910. [Google Scholar] [CrossRef]
- Jian, F.X. The Regulation of Mouse Immune Functions by olysaccharides from Mulberry Bark. Sci. Seric. 2009, 35, 907–912. [Google Scholar]
- Guo, C.; Li, R.; Zheng, N.; Xu, L.; Liang, T.; He, Q. Anti-diabetic effect of ramulus mori polysaccharides, isolated from Morus alba L., on STZ-diabetic mice through blocking inflammatory response and attenuating oxidative stress. Int. Immunopharmacol. 2013, 16, 93–99. [Google Scholar] [CrossRef]
- Liu, C.J.; Lin, J.Y. Anti-inflammatory and anti-apoptotic effects of strawberry and mulberry fruit polysaccharides on lipopolysaccharide-stimulated macrophages through modulating pro-/anti-inflammatory cytokines secretion and Bcl-2/Bak protein ratio. Food Chem. Toxicol. 2012, 50, 3032–3039. [Google Scholar] [CrossRef]
- Zhang, Y.; Ren, C.; Lu, G.; Mu, Z.; Cui, W.; Gao, H.; Wang, Y. Anti-diabetic effect of mulberry leaf polysaccharide by inhibiting pancreatic islet cell apoptosis and ameliorating insulin secretory capacity in diabetic rats. Int. Immunopharmacol. 2014, 22, 248–257. [Google Scholar] [CrossRef]
- Banerjee, P.; Mukherjee, S.; Bera, K.; Ghosh, K.; Ali, I.; Khawas, S.; Ray, S. Polysaccharides from Thymus vulgaris leaf: Structural features, antioxidant activity and interaction with bovine serum albumin. Int. J. Biol. Macromol. 2019, 125, 580–587. [Google Scholar] [CrossRef]
- Tahmouzi, S.; Nejat, M.R.S. New infertility therapy effects of polysaccharides from Althaea officinalis leaf with emphasis on characterization, antioxidant and anti-pathogenic activity. Int. J. Biol. Macromol. 2020, 145, 777–787. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Zhou, X.; Deng, Q.; Yang, M.; Li, S.; Zhang, Q.; Chen, H. Extraction, structural characterization and biological activities of polysaccharides from mulberry leaves: A review. Int. J. Biol. Macromol. 2023, 257, 128669. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Cai, B.; Wang, J.; Sheng, Z.; Yang, H.; Wang, D.; Chen, J.; Ning, Q. Mulberry leaf-derived polysaccharide modulates the immune response and gut microbiota composition in immunosuppressed mice. J. Funct. Foods 2021, 83, 104545. [Google Scholar] [CrossRef]
- Liu, C.; Ma, Y.; Zhang, X. Effects of mulberry leaf polysaccharide on oxidative stress in pancreatic β-cells of type 2 diabetic rats. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 2482–2488. [Google Scholar] [PubMed]
- Ren, C.; Zhang, Y.; Cui, W.; Lu, G.; Wang, Y.; Gao, H.; Huang, L.; Mu, Z. A polysaccharide extract of mulberry leaf ameliorates hepatic glucose metabolism and insulin signaling in rats with type 2 diabetes induced by high fat-diet and streptozotocin. Int. J. Biol. Macromol. 2015, 72, 951–959. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Li, L.; Li, Z.K.; Wu, F.; Hang, B.Y.; Cai, B.Y.; Du, Y.G. Effect and mechanism of mulberry leaf polysaccharide on type 1 diabetic nephropathy in rats. Zhonghua Yi Xue Za Zhi 2018, 98, 1792–1796. [Google Scholar] [PubMed]
- Voorhees, R. Membrane Protein Biosynthesis at the Endoplasmic Reticulum. Microsc. Microanal. 2020, 26, 116. [Google Scholar] [CrossRef]
- Zhang, H.; Hu, W.; Zhong, Y.; Guo, Z. Meta-analysis of the effects of smooth endoplasmic reticulum aggregation on birth outcome. BMC Pregnancy Childbirth 2021, 21, 374. [Google Scholar] [CrossRef]
- Raote, I.; Malhotra, V. Tunnels for protein export from the endoplasmic reticulum. Annu. Rev. Biochem. 2021, 90, 605–630. [Google Scholar] [CrossRef]
- Wang, Y.; Hu, H.; Li, H.; Ma, H.; Xu, F.; Qu, B. Effects of lead exposure on placental cellular apoptosis and endoplasmic reticulum stress in rats. Chin. Med. J. 2014, 127, 1744–1748. [Google Scholar] [CrossRef]
- Wu, K.; Luo, Z.; Hogstrand, C.; Chen, G.H.; Wei, C.C.; Li, D.D. Zn stimulates the phospholipids biosynthesis via the pathways of oxidative and endoplasmic reticulum stress in the intestine of freshwater teleost yellow catfish. Environ. Sci. Technol. 2018, 52, 9206–9214. [Google Scholar] [CrossRef] [PubMed]
- Bravo, R.; Parra, V.; Gatica, D.; Rodríguez, A.E.; Torrealba, N.; Paredes, F.; Wang, Z.V.; Zorzano, A.; Hill, J.A.; Jaimovich, E.; et al. Endoplasmic reticulum and the unfolded protein response: Dynamics and metabolic integration. Int. Rev. Cell Mol. Biol. Rep. 2013, 301, 215–290. [Google Scholar]
- Hu, R.; Wang, S.; Feng, L.; Jiang, W.; Wu, P.; Liu, Y.; Jin, X.; Kuang, S.; Tang, L.; Zhang, L. Alleviation of hypoxia stress induced oxidative damage, endoplasmic reticulum stress (ERS) and autophagy in grass carp (Ctenopharyngodon idellu) by TTO (Melaleuca alternifolia essential oil). Aquaculture 2023, 564, 739073. [Google Scholar] [CrossRef]
- Ou, H.; Liang, J.; Liu, J. Effects of acute ammonia exposure on oxidative stress, endoplasmic reticulum stress and apoptosis in the kuruma shrimp (Marsupenaeus japonicus). Aquac. Rep. 2022, 27, 101383. [Google Scholar] [CrossRef]
- Fu, C.; Qiu, Z.; Huang, Y.; Lin, Q.; Jin, L.; Tu, H.; Ye, J.; Zheng, C.; Zhong, W.; Ma, D. Achyranthes bidentata polysaccharides alleviate endoplasmic reticulum stress in osteoarthritis via lncRNA NEAT1/miR-377-3p pathway. Biomed. Pharmacother. 2022, 154, 113551. [Google Scholar] [CrossRef]
- Sun, S.; Yang, S.; An, N.; Wang, G.; Xu, Q.; Liu, J.; Mao, Y. Astragalus polysaccharides inhibits cardiomyocyte apoptosis during diabetic cardiomyopathy via the endoplasmic reticulum stress pathway. J. Ethnopharmacol. 2019, 238, 111857. [Google Scholar] [CrossRef]
- Yang, F.; Wei, Y.; Liao, B.; Wei, G.; Qin, H.; Pang, X.; Wang, J. Effects of Lycium barbarum Polysaccharide on Endoplasmic Reticulum Stress and Oxidative Stress in Obese Mice. Front. Pharmacol. 2020, 11, 742. [Google Scholar] [CrossRef]
- Shelley, L.K.; Ross, P.S.; Kennedy, C.J. The effects of an in vitro exposure to 17β-estradiol and nonylphenol on rainbow trout (Oncorhynchus mykiss) peripheral blood leukocytes. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2012, 155, 440–446. [Google Scholar] [CrossRef]
- Liu, B.; Yang, Z.; Bo, L.; Zhao, Z.; Sun, C. Cytotoxic effects, inflammatory response and apoptosis induction of cyclophosphamide in the peripheral blood leukocyte of blunt snout bream (Megalobrama amblycephala). Fish Shellfish Immunol. 2019, 93, 174–182. [Google Scholar] [CrossRef]
- Huang, X.; Miao, L.H.; Lin, Y.; Pan, W.J.; Ren, M.C.; Ge, X.P.; Liu, B.; Zhou, Q.L. High glucose affected respiratory burst activity of peripheral leukocyte via G6PD and NOX inhibition in Megalobrama amblycephala. Fish Shellfish Immunol. 2018, 83, 243–248. [Google Scholar] [CrossRef]
- Betoulle, S.; Duchiron, C.; Deschaux, P. Lindane differently modulates intracellular calcium levels in two populations of rainbow trout (Oncorhynchus mykiss) immune cells: Head kidney phagocytes and peripheral blood leucocytes. Toxicology 2000, 145, 203–215. [Google Scholar] [CrossRef] [PubMed]
- Hao, H.; Cao, L.; Jiang, C.; Che, Y.; Zhang, S.; Takahashi, S. Farnesoid X Receptor Regulation of the NLRP3 Inflammasome Underlies Cholestasis-Associated Sepsis. Cell Metab. 2017, 25, 856–867.e855. [Google Scholar] [CrossRef] [PubMed]
- Du, L.; Lin, L.; Li, Q.; Liu, K.; Huang, Y.; Wang, X. IGF-2 Preprograms Maturing Macrophages to Acquire Oxidative Phosphorylation-Dependent Anti-inflammatory Properties. Cell Metab. 2019, 29, 1363–1375.e1368. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Sun, L.; Hao, M.; Liu, X. Circ-SWT1 Ameliorates H2O2-Induced Apoptosis, Oxidative Stress and Endoplasmic Reticulum Stress in Cardiomyocytes via miR-192-5p/SOD2 Axis. Cardiovasc. Toxicol. 2022, 22, 378–389. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Liao, X.H.; Huang, L.L.; Sun, H.; Liu, Q.; Zhang, L. Overexpression of augmenter of liver regeneration (ALR) mitigates the effect of H2O2-induced endoplasmic reticulum stress in renal tubule epithelial cells. Apoptosis 2019, 24, 278–289. [Google Scholar] [CrossRef] [PubMed]
- Jia, R.; Du, J.; Cao, L.; Feng, W.; He, Q.; Xu, P. Chronic exposure of hydrogen peroxide alters redox state, apoptosis and endoplasmic reticulum stress in common carp (Cyprinus carpio). Aquat. Toxicol. 2020, 229, 105657. [Google Scholar] [CrossRef] [PubMed]
- Pobre, K.F.R.; Poet, G.J.; Hendershot, L.M. The endoplasmic reticulum (ER) chaperone BiP is a master regulator of ER functions: Getting by with a little help from ERdj friends. J. Biol. Chem. 2019, 294, 2098–2108. [Google Scholar] [CrossRef]
- Rao, J.; Yue, S.; Fu, Y.; Zhu, J.; Wang, X.; Busuttil, R.W. ATF6 Mediates a Pro-Inflammatory Synergy Between ER Stress and TLR Activation in the Pathogenesis of Liver Ischemia-Reperfusion Injury. Am. J. Transplant. 2014, 14, 1552–1561. [Google Scholar] [CrossRef]
- Zhou, X.; Lu, B.; Fu, D.; Gui, M.; Yao, L.; Li, J. Huoxue Qianyang decoction ameliorates cardiac remodeling in obese spontaneously hypertensive rats in association with ATF6-CHOP endoplasmic reticulum stress signaling pathway regulation. Biomed. Pharmacother. 2020, 121, 109518. [Google Scholar] [CrossRef]
- Wu, L.; Li, H.; Xu, W.; Dong, B.; Geng, H.; Jin, J.; Han, D.; Liu, H.; Zhu, X.; Yang, Y. Emodin alleviates acute hypoxia-induced apoptosis in gibel carp (Carassius gibelio) by upregulating autophagy through modulation of the AMPK/mTOR pathway. Aquaculture 2022, 548, 737689. [Google Scholar] [CrossRef]
- Huang, C.; Yao, R.; Zhu, Z.; Pang, D.; Cao, X.; Feng, B.; Paulsen, B.S.; Li, L.; Yin, Z.; Chen, X. A pectic polysaccharide from water decoction of Xinjiang Lycium barbarum fruit protects against intestinal endoplasmic reticulum stress. Int. J. Biol. Macromol. 2019, 130, 508–514. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Crisóstomo, N.P.; Rodríguez Martínez, E.; Selva, R.A. Oxidative stress activates the transcription factors FoxO 1a and FoxO 3a in the hippocampus of rats exposed to low doses of ozone. Oxid. Med. Cell Longev. 2014, 2014, 805764. [Google Scholar] [CrossRef] [PubMed]
- Shen, M.; Lin, F.; Zhang, J.; Tang, Y.; Chen, W.K.; Liu, H. Involvement of the up-regulated FoxO1 expression in follicular granulosa cell apoptosis induced by oxidative stress. J. Biol. Chem. 2012, 287, 25727–25740. [Google Scholar] [CrossRef] [PubMed]
- Ding, H.R.; Tang, Z.T.; Tang, N.; Zhu, Z.Y.; Liu, H.Y.; Pan, C.Y.; Hu, A.Y.; Lin, Y.Z.; Gou, P.; Yuan, X.W. Protective properties of FOXO1 inhibition in a murine model of non-alcoholic fatty liver disease are associated with attenuation of ER stress and necroptosis. Front. Physiol. 2020, 11, 177. [Google Scholar] [CrossRef]
- Safra, M.; Fickentscher, R.; Levi-Ferber, M.; Danino, Y.M.; Haviv-Chesner, A.; Hansen, M.; Juven-Gershon, T.; Weiss, M.; Henis-Korenblit, S. The FOXO transcription factor DAF-16 bypasses ire-1 requirement to promote endoplasmic reticulum homeostasis. Cell Metab. 2014, 20, 870–881. [Google Scholar] [CrossRef] [PubMed]
- Farag, M.R.; Alagawany, M.; Khalil, S.R.; Moustafa, A.A.; Mahmoud, H.K.; Abdel-Latif, H.M. Astragalus membranaceus polysaccharides modulate growth, hemato-biochemical indices, hepatic antioxidants, and expression of HSP70 and apoptosis-related genes in Oreochromis niloticus exposed to sub-lethal thallium toxicity. Fish Shellfish Immunol. 2021, 118, 251–260. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Fang, Y.; Zou, C. Pomelo polysaccharide extract inhibits oxidative stress, inflammation, and mitochondrial apoptosis of Epinephelus coioides. Aquaculture 2021, 544, 737040. [Google Scholar] [CrossRef]
- Zuo, Z.; Wang, S.; Wang, Q.; Wang, D.; Wu, Q.; Xie, S.; Zou, J. Effects of partial replacement of dietary flour meal with seaweed polysaccharides on the resistance to ammonia stress in the intestine of hybrid snakehead (Channa maculatus ♀ × Channa argus ♂). Fish Shellfish Immunol. 2022, 127, 271–279. [Google Scholar] [CrossRef]
- Mohammadi, G.; Karimi, A.A.; Hafezieh, M.; Dawood, M.A.; Abo-Al-Ela, H.G. Pistachio hull polysaccharide protects Nile tilapia against LPS-induced excessive inflammatory responses and oxidative stress, possibly via TLR2 and Nrf2 signaling pathways. Fish Shellfish Immunol. 2022, 121, 276–284. [Google Scholar] [CrossRef]
- Fan, H.; Sun, M.; Li, J.; Zhang, S.; Tu, G.; Liu, K.; Xia, Q.; Jiang, Y.; Liu, B. Structure characterization and immunomodulatory activity of a polysaccharide from Saposhnikoviae Radix. Int. J. Biol. Macromol. 2023, 233, 123502. [Google Scholar] [CrossRef]
- Xiao, Z.; Deng, Q.; Zhou, W.; Zhang, Y. Immune activities of polysaccharides isolated from Lycium barbarum L. What do we know so far? Pharm. Ther. 2022, 229, 107921. [Google Scholar] [CrossRef]
- Cao, W.; Wu, J.; Zhao, X.; Li, Z.; Yu, J.; Shao, T.; Han, J. Structural elucidation of an active polysaccharide from Radix Puerariae lobatae and its protection against acute alcoholic liver disease. Carbohydr. Polym. 2024, 325, 121565. [Google Scholar] [CrossRef] [PubMed]
- Qin, X.; Nong, K.; Liu, Z.; Fang, X.; Zhang, B.; Chen, W.; Wang, Z.; Wu, Y.; Shi, H.; Wang, X.; et al. Regulation of the intestinal flora using polysaccharides from Callicarpa nudiflora Hook to alleviate ulcerative colitis and the molecular mechanisms involved. Int. J. Biol. Macromol. 2024, 258, 128887. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Liu, Y.; Sun, Y.; Mou, Q.; Wang, B.; Zhang, Y.; Huang, L. Structural characterization of LbGp1 from the fruits of Lycium barbarum L. Food Chem. 2014, 159, 137–142. [Google Scholar] [CrossRef] [PubMed]
- Gong, G.; Dang, T.; Deng, Y.; Han, J.; Zou, Z.; Jing, S.; Zhang, Y.; Liu, Q.; Huang, L.; Wang, Z. Physicochemical properties and biological activities of polysaccharides from Lycium barbarum prepared by fractional precipitation. Int. J. Biol. Macromol. 2018, 109, 611–618. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.; Li, K.; Xiao, L.; Lei, Z.; Zhang, Z. Characterization of polysaccharide from Helicteres angustifolia L. and its immunomodulatory activities on macrophages RAW264. 7. Biomed. Pharmacother. 2019, 109, 262–270. [Google Scholar] [CrossRef] [PubMed]
- Csordás, G.R.; Renken, C.; Várnai, P.; Walter, L.; Weaver, D.; Buttle, K.F.; Balla, T.; Mannella, C.A.; Hajnóczky, G.R. Structural and functional features and significance of the physical linkage between ER and mitochondria. J. Cell Biol. 2006, 174, 915–921. [Google Scholar] [CrossRef] [PubMed]
- Peng, J.; Peng, C.; Wang, L.; Cao, H.; Xing, C.; Li, G.; Hu, G.; Yang, F. Endoplasmic reticulum-mitochondria coupling attenuates vanadium-induced apoptosis via IP3R in duck renal tubular epithelial cells. J. Inorg. Chem. 2022, 232, 111809. [Google Scholar] [CrossRef]
- De Brito, O.M.; Scorrano, L. Mitofusin 2 tethers endoplasmic reticulum to mitochondria. Nature. 2008, 456, 605–610. [Google Scholar] [CrossRef]
- Erustes, A.G.; D’Eletto, M.; Guarache, G.C.; Ureshino, R.P.; Bincoletto, C.; Silva Pereira, G.J.; Piacentini, M.; Smaili, S.S. Overexpression of α-synuclein inhibits mitochondrial Ca2+ trafficking between the endoplasmic reticulum and mitochondria through MAMs by altering the GRP75–IP3R interaction. J. Neurosci. Res. 2021, 99, 2932–2947. [Google Scholar] [CrossRef]
- Wang, W.; Xie, Q.; Zhou, X.; Yao, J.; Zhu, X.; Huang, P.; Zhang, L.; Wei, J.; Xie, H.; Zhou, L. Mitofusin-2 triggers mitochondria Ca2+ influx from the endoplasmic reticulum to induce apoptosis in hepatocellular carcinoma cells. Cancer Lett. 2015, 358, 47–58. [Google Scholar] [CrossRef] [PubMed]
- Misrani, A.; Tabassum, S.; Yang, L. Mitochondrial Dysfunction and Oxidative Stress in Alzheimer’s Disease. Front. Aging Neurosci. 2021, 13, 57. [Google Scholar] [CrossRef] [PubMed]
- Sharma, C.; Kim, S.; Nam, Y.; Jung, U.J.; Kim, S.R. Mitochondrial Dysfunction as a Driver of Cognitive Impairment in Alzheimer’s Disease. Int. J. Mol. Sci. 2021, 22, 4850. [Google Scholar] [CrossRef]
- Xiang, Q.D.; Yu, Q.; Wang, H.; Zhao, M.M.; Liu, S.Y.; Nie, S.P.; Xie, M.Y. Immunomodulatory Activity of Ganoderma atrum Polysaccharide on Purified T Lymphocytes through Ca2+/CaN and Mitogen-Activated Protein Kinase Pathway Based on RNA Sequencing. J. Agric. Food Chem. 2017, 65, 5306–5315. [Google Scholar] [CrossRef] [PubMed]
- Miao, X.Y.; Zhu, X.X.; Gu, Z.Y.; Fu, B.; Cui, S.Y.; Zu, Y.; Rong, L.J.; Hu, F.; Chen, X.M.; Gong, Y.P.; et al. Astragalus polysaccharides reduce high-glucose-induced rat aortic endothelial cell senescence and inflammasome activation by modulating the mitochondrial Na+/Ca2+ exchanger. Cell Biochem. Biophys. 2022, 80, 1–13. [Google Scholar] [CrossRef]
- Sun, S.L.; Guo, L.; Ren, Y.C.; Wang, B.; Li, R.H.; Qi, Y.S.; Yu, H.; Chang, N.D.; Li, M.H.; Peng, H.S. Anti-apoptosis effect of polysaccharide isolated from the seeds of Cuscuta chinensis Lam on cardiomyocytes in aging rats. Mol. Biol. Rep. 2014, 41, 6117–6124. [Google Scholar] [CrossRef]
- Dai, H.; Jia, G.; Liu, X.; Liu, Z.; Wang, H. Astragalus polysaccharide inhibits isoprenaline-induced cardiac hypertrophy via suppressing Ca2+-mediated calcineurin/NFATc3 and CaMKII signaling cascades. Environ. Toxicol. Pharmacol. Ther. 2014, 38, 263–271. [Google Scholar] [CrossRef]
- Hu, S.; Wang, D.; Zhang, J.; Du, M.; Cheng, Y.; Liu, Y.; Zhang, N.; Wang, D.; Wu, Y. Mitochondria related pathway is essential for polysaccharides purified from sparassis crispa mediated neuro-protection against glutamate-induced toxicity in differentiated PC12 cells. Int. J. Mol. Sci. 2016, 17, 133. [Google Scholar] [CrossRef]
- Russo, A.; Cardile, V.; Graziano, A.C.; Avola, R.; Bruno, M.; Rigano, D. Involvement of Bax and Bcl-2 in induction of apoptosis by essential oils of three Lebanese Salvia species in human prostate cancer cells. Int. J. Mol. Sci. 2018, 19, 292. [Google Scholar] [CrossRef]
- Zhou, H.; Wang, X.; Zhang, B. Depression of lncRNA NEAT1 antagonizes LPS-evoked acute injury and inflammatory response in alveolar epithelial cells via HMGB1-RAGE signaling. Mediat. Inflamm. 2020, 2020, 1–11. [Google Scholar] [CrossRef]
- Suski, J.M.; Lebiedzinska, M.; Bonora, M.; Pinton, P.; Duszynski, J.; Wieckowski, M.R. Relation between mitochondrial membrane potential and ROS formation. In Mitochondrial Bioenergetics: Methods Protocols; Springer: Berlin/Heidelberg, Germany, 2018; pp. 357–381. [Google Scholar]
- Duan, Y.; Gross, R.A.; Sheu, S.S. Ca2+-dependent generation of mitochondrial reactive oxygen species serves as a signal for poly (ADP-ribose) polymerase-1 activation during glutamate excitotoxicity. J. Physiol. 2007, 585, 741–758. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B.; Gutteridge, J.M. Free Radicals in Biology and Medicine; Oxford University Press: New York, NY, USA, 2015. [Google Scholar]
- Cui, Y.T.; Liu, B.; Xie, J.; Xu, P.; Habte-Tsion, H.; Zhang, Y.Y. The effect of emodin on cytotoxicity, apoptosis and antioxidant capacity in the hepatic cells of grass carp (Ctenopharyngodon idellus). Fish Shellfish Immunol. 2014, 38, 74–79. [Google Scholar] [CrossRef] [PubMed]
- Yi, J.; Li, X.; Wang, S.; Wu, T.; Liu, P. Steam explosion pretreatment of Achyranthis bidentatae radix: Modified polysaccharide and its antioxidant activities. Food Chem. 2022, 375, 131746. [Google Scholar] [CrossRef]
- Zhu, X.; Liu, X.; Xia, C.; Li, M.; Niu, X.; Wang, G.; Zhang, D. Effects of dietary Astragalus Propinquus Schischkin polysaccharides on growth performance, immunological parameters, antioxidants responses and inflammation-related gene expression in Channa argus. Comparative biochemistry and physiology. Toxicol. Pharmacol. 2021, 249, 109121. [Google Scholar] [CrossRef]
- Shi, F.L.; Zhi, J.Y.; Min, X.L.; Feng, Z.; Fan, B.Z.; Li, J.L.; Yan, A.L.; Qing, Q.L.; Jiang, T.L.; Jun, L.; et al. Astragalus polysaccharides mediate the immune response and intestinal microbiota in grass carp (Ctenopharyngodon idellus). Aquaculture 2021, 534, 736205. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Primer Sequence (5′-3′) | Product Length (bps) | Accession No. | Amplification Efficiency | |
|---|---|---|---|---|---|
| foxO1α | Forward | GGATGATGCCTGAGATGG | 148 | XM_048206068.1 | 109.8% |
| Reverse | GACCTGGAGGCTCGTGT | ||||
| bip | Forward | CCTGGGACAGAAGGTCACAC | 133 | XM_048186701.1 | 100.4% |
| Reverse | AGCAGTTGGCTCGTTGATGA | ||||
| atf6 | Forward | CGATCAGGATGGAGAGTGGGATA | 157 | XM_048207041.1 | 108.3% |
| Reverse | AGGGCTACTCCACAATGGGT | ||||
| chop | Forward | ATGTGGTGCAGAGTTGGAGG | 124 | XM_048198700.1 | 108.2% |
| Reverse | CACATCCAGAAACTCGGGCT | ||||
| mcu | Forward | AAGGGAGCGAAGAGAACACG | 128 | XM_048205815.1 | 106.6% |
| Reverse | ATTTGCTGGACAGGGAGGTG | ||||
| vdac1 | Forward | GATAACCTGGAGACGGCAGTT | 146 | XM_048166591.1 | 104.7% |
| Reverse | GTATATCCCAGGCCGACGAG | ||||
| mfn2 | Forward | TGCTCACTCTGGACTGCAAG | 192 | XM_048210720.1 | 109.0% |
| Reverse | GACCGTCCTCTATGTGCCTG | ||||
| grp75 | Forward | CGGCGTTATGATGACCCAGA | 184 | XM_048175758.1 | 106.7% |
| Reverse | TCACAGCATGGCCCAAGTAG | ||||
| il-6 | Forward | AAGACAACCGCACACTCGAT | 122 | XM_048203704.1 | 97.2% |
| Reverse | CTGGGTCTCTTCACGCCTTT | ||||
| bax | Forward | AGTGTTTGCAGCAGATCGGA | 162 | XM_048196672.1 | 104.3% |
| Reverse | AGAAAAGAGCCACCACCCTG | ||||
| β-actin | Forward | TCGTCCACCGCAAATGCTTCTA | 190 | AY170122.2 | 99.9% |
| Reverse | CCGTCACCTTCACCGTTCCAGT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, W.; Lin, Y.; Qian, L.; Lu, S.; Shen, H.; Ge, X.; Miao, L. Mulberry Leaf Polysaccharides Attenuate Oxidative Stress Injury in Peripheral Blood Leukocytes by Regulating Endoplasmic Reticulum Stress. Antioxidants 2024, 13, 136. https://doi.org/10.3390/antiox13020136
Jiang W, Lin Y, Qian L, Lu S, Shen H, Ge X, Miao L. Mulberry Leaf Polysaccharides Attenuate Oxidative Stress Injury in Peripheral Blood Leukocytes by Regulating Endoplasmic Reticulum Stress. Antioxidants. 2024; 13(2):136. https://doi.org/10.3390/antiox13020136
Chicago/Turabian StyleJiang, Wenqiang, Yan Lin, Linjie Qian, Siyue Lu, Huaishun Shen, Xianping Ge, and Linghong Miao. 2024. "Mulberry Leaf Polysaccharides Attenuate Oxidative Stress Injury in Peripheral Blood Leukocytes by Regulating Endoplasmic Reticulum Stress" Antioxidants 13, no. 2: 136. https://doi.org/10.3390/antiox13020136
APA StyleJiang, W., Lin, Y., Qian, L., Lu, S., Shen, H., Ge, X., & Miao, L. (2024). Mulberry Leaf Polysaccharides Attenuate Oxidative Stress Injury in Peripheral Blood Leukocytes by Regulating Endoplasmic Reticulum Stress. Antioxidants, 13(2), 136. https://doi.org/10.3390/antiox13020136