
Cherries with Different Geographical Origins Regulate Neuroprotection in a Photoperiod-Dependent Manner in F344 Rats
by
 , , and
, , and
Francesca Manocchio
1,2,3,
Francisca Isabel Bravo
1,2,3,* ,
,
Gisela Helfer
4,* and
Begoña Muguerza
1,2,3 1
Nutrigenomics Research Group, Department of Biochemistry and Biotechnology, Universitat Rovira i Virgili, C Marcel·lí Domingo s/n, 43007 Tarragona, Spain
2
Nutrigenomics Research Group, Institut d’Investigació Sanitària Pere Virgili, C/Marcel·lí Domingo s/n, 43007 Tarragona, Spain
3
Center of Environmental, Food and Toxicological Technology (TecnATox), Universitat Rovira i Virgili, 43007 Tarragona, Spain
4
School of Chemistry and Biosciences, University of Bradford, Richmond Road, Bradford BD7 1DP, UK
*
Authors to whom correspondence should be addressed.
Antioxidants 2024, 13(1), 72; https://doi.org/10.3390/antiox13010072
Submission received: 29 November 2023
/
Revised: 24 December 2023
/
Accepted: 27 December 2023
/
Published: 3 January 2024
(This article belongs to the Special Issue Natural Products: Biological-, Antioxidant Properties and Health Effects - 2nd Edition)
Abstract
:The photoperiod is the main environmental cue that drives seasonal adaptive responses in reproduction, behavior, and metabolism in seasonal animals. Increasing evidence suggests that (poly)phenols contained in fruits can also modulate seasonal rhythms. (Poly)phenol-rich diets are associated with an improvement in cognitive function and neuroprotection due to their anti-inflammatory and antioxidative properties. However, it is unknown whether cherries affect neuroprotection in a photoperiod-dependent manner. To test this, F344 rats were exposed to L6 (6 h light/day), L12 (12 h light/day) and L18 (18 h light/day) photoperiods and fed a standard chow diet supplemented with either a control, lyophilized cherry 1 or cherry 2 with distinctive phenolic hallmarks. Physiological parameters (body weight, eating pattern index (EPI), testosterone, T4/T3) and hypothalamic key genes (Dio2, Dio3, Raldh1 and Ghrh) were strongly regulated by the photoperiod and/or fruit consumption. Importantly, we show for the first time that neurotrophs (Bdnf, Sod1 and Gpx1) in the hippocampus are also regulated by the photoperiod. Furthermore, the consumption of cherry 2, which was richer in total flavonols, but not cherry 1, which was richer in total anthocyanins and flavanols, enhanced neuroprotection in the hippocampus. Our results show that the seasonal consumption of cherry with a specific phenolic composition plays an important role in the hippocampal activation of neuroprotection in a photoperiod-dependent manner.
Keywords:
phenolic compounds; antioxidant-rich fruit; hippocampus; neurotroph; BDNF; seasonal; photoperiod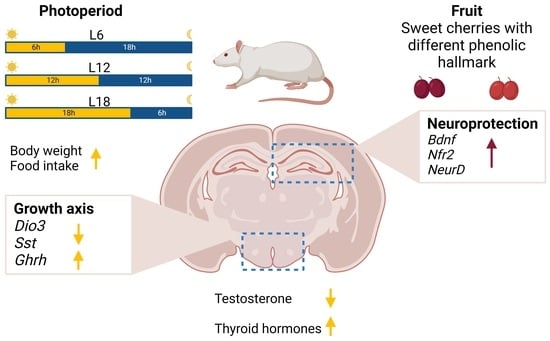
1. Introduction
Many organisms adapt to seasonal fluctuations in the climate with profound changes in their physiology, behavior and reproduction [1]. These seasonal changes are driven by changes in day length (photoperiod) and mediated via neuroendocrine processes in the hypothalamus. Briefly, photoperiod information is biochemically translated through the release of melatonin from the pineal gland during dark hours. Melatonin then acts on the pars tuberalis of the pituitary gland to regulate thyroid-stimulating hormone release, which, in turn, regulates the thyroid hormone availability of the deiodinase enzymes DIO2 and DIO3 in the hypothalamic tanycytes lining the third ventricle of the hypothalamus. Indeed, the hypothalamic tanycytes express thyroid-stimulating hormone receptors. Thus, they sense thyroid-stimulating hormone subunit β signals from the pituitary, expression of which is upregulated by the photoperiod and results in long-photoperiod-induced Dio2 mRNA expression (12, 14 or 16 h of light) and short-photoperiod-induced Dio3 mRNA expression (6 and 8 h of light). DIO2 converts the inactive form of the thyroid hormone, thyroxine (T4), into the active form, triiodothyronine (T3), increasing the hypothalamic concentration of T3 [1]. Thus, the hypothalamic–pituitary–thyroid and hypothalamic–pituitary–gonadal axes are sensitive to changes in photoperiod [2]. In addition, hypothalamic tanycytes express a variety of nutrient sensors [3]. Nutrients (glucose, amino acids and lipids), as well as hormones (leptin, ghrelin and insulin) produced in response to nutrient availability, can modulate gene expression in tanycytes, which ultimately regulates energy metabolism [4]. Moreover, the hypothalamus is sensitive to phenolic compounds, which are among the main bioactive components of fruits and vegetables [5]. In support, the administration of grapes and cherries, which are rich in phenolic compounds, to photoperiod-sensitive Fischer 344 (F344) rats increases hypothalamic leptin sensitivity when they are consumed during a short photoperiod [5,6]. Phenolic compounds can exert a neuroprotective effect. A recent study showed that the consumption of tart cherries, a fruit rich in phenolic compounds, significantly affects the plasma metabolome, associated with an improvement in cognitive function in humans consuming this fruit [7]. For example, tart cherry exerts anti-inflammatory properties, suppresses neuronal apoptosis and stimulates pro-survival signaling cascades in vitro [8] and in aged rats [7,8]. Furthermore, aged rats consuming blueberries, a fruit also rich in phenolic compounds, show an improvement in working memory and an upregulation of hippocampal brain-derived neurotrophic factor (Bdnf) gene expression compared with control rats [9]. The BDNF protein plays a key role in neuronal plasticity, and, together with nuclear factor erythroid-derived 2-like (NRF2), promotes the gene expression of antioxidant enzymes in the hippocampus [10].
Recently, our group showed that Royal Dawn sweet cherries increased serum and hepatic antioxidant markers in F344 rats depending on the cherry composition profile and the photoperiod of consumption [11]. Moreover, consumption of the same sweet cherries modulated the activity of transaminase enzymes and reduced lipid peroxidation markers in Fisher 344 rats, also depending on their phenolic profile and the photoperiod in which they were consumed [11]. Cherries are considered a good source of dietary phenolic compounds [12,13]. The predominant phenolic compounds are cyanidin derivatives (anthocyanins), with cyanidin-3-O-rutinoside being the major compound [14,15]. Moreover, cherries also contain high levels of hydroxycinnamic acids, caffeoylquinic acid derivatives and rutin, although their profiles can differ depending on the method used to extract the cherry’s phenolic fraction [15]. Interestingly, as (poly)phenols are plant stress metabolites, the cherry’s phenolic profile is distinctive and depends on the location, time of harvest and storage conditions, among other factors [16,17]. Although cherries are seasonal fruits (available from May to August), out of season they are harvested and imported from different geographical locations to allow for continuous sales and consumption [18]. However, it is yet unknown whether the consumption of cherries with different phenolic compositions exerts a neuroprotective effect in a photoperiod-dependent manner, reducing oxidative stress. The aim of this study was to investigate if the consumption of sweet cherries plays a role in neuroprotection in a photoperiod- and composition-profile-dependent manner.
2. Materials and Methods
2.1. Fruit Preparation and Composition
Royal Dawn sweet cherries (Prunus avium L.) were harvested in two different geographical locations in order to obtain cherries with different compositions. Cherries from the Tarragona region in Spain (Ch1) were kindly donated by a local farmer. Cherries from Chachapoal in Chile (Ch2) were purchased in a local market. Sweet cherry pulps were frozen in liquid nitrogen, ground and lyophilized. After lyophilization, the obtained cherry powders were kept at −20 °C until administration to animals. Basic composition was analyzed according to the AOAC method [19], and the phenolic content of these two cherries has been published in Cruz-Carrión et al. 2020 [11]. Phenolic compounds in the cherries were extracted for analysis using 65% methanol (1% formic acid) at a ratio of 80 mL/g of cherry, 72 °C, and 100 min of agitation at 500 rpm [11,15]. Phenolic composition was analyzed using spectrophotometric methods, total polyphenols by the Folin–Ciocalteu method, total anthocyanins by the pH differential method, total flavanols by the DMACA method, and total flavonols by mixing the sample with 0.1% HCl and measuring the absorbance at 360 nm after 15 min of reaction [15,20,21]. Ch1 showed higher contents of total ash, protein, lipids, fiber, polyphenols, anthocyanins and flavanols than Ch2; however, Ch2 showed higher levels of total carbohydrates, sugars and total flavonols than Ch1 [11]. Additional HPLC-ESI-MS/MS studies showed that anthocyanins were the major phenolic family in both cherries (75 and 52% of the total quantified phenolic compounds for Ch1 and Ch2, respectively), followed by flavonols, flavanols and phenolic acids. The major compounds were cyanidin-3-O-rutinoside (Ch1 > Ch2, 3.42 times), quercetin-3-O-rutinoside (Ch2 > Ch1, 1.18 times), catechin (Ch1 > Ch2, 3.20 times), 4-hydroxybenzoic acid (Ch2 > Ch1, 1.04 times) and 3-O-caffeoylquinic acid (Ch1 > Ch2, 1.33 times) (data not published). Cherries were characterized during the same period in which they were administered to the animals (Section 2.2).
2.2. Experimental Design
Animal experiments were conducted in accordance with the European Communities Council Directive (86/609/EEC) and approved by the Animal Ethics Review Committee for Animal Experimentation of the Universitat Rovira i Virgili and Generalitat de Catalunya (permission number 9495, FUE-2017-00499873). A total of 72 male Fischer 344/IcoCrl rats (7–8 weeks old, body weight 216–244 g) were purchased from Charles River Laboratories (Barcelona, Spain) and housed in pairs at 22 °C and 50% humidity under three photoperiods (light intensity 700 lx): L6 (short photoperiod; 6 h light/day), L12 (12 h light/day) and L18 (long photoperiod; 18 h light/day). Throughout the experiment, the rats were fed a standard chow diet (AO4, Panlab, Barcelona, Spain) and tap water ad libitum. After 4 weeks of photoperiod acclimatization, F344 rats in each photoperiod group were randomly subdivided into three groups. One group of F344 rats received chow diet, which was supplemented with 100 mg Ch1 dry weight (dw)/kg body weight (bw) (Ch1 group), other group received 100 mg Ch2 dw/kg (Ch2 group) and the control group (C group) received 21.2 mg glucose /kg bw and 21.2 mg fructose /kg bw to match the sugar contents of the diets between the three groups (n = 8 per group). The group size [22,23,24] and cherry dose [24] were selected based on the results of previous studies. During the four weeks of photoperiod acclimatization, rats were trained to lick a syringe with water, which allowed us to avoid oral gavage, thus reducing stress and improving animal welfare during the seven weeks of fruit supplementation. Fruit supplements were dissolved in tap water and administered by voluntary licking with a syringe. The experimenter visually confirmed that the full dose was administered. Rats were weighed every week, and food intake was measured in the last week. The weight (in grams) of the pellets of the standard diet was recorded when the food was provided and subtracted after 24 h to calculate how many grams of rat pellets were consumed. The eating pattern index (EPI) was calculated according to the calculations used by Ruiz de Azua et al. 2023 [16] and considers the kcal consumed as well as the fact that animals eat mainly during the hours of darkness: EPI (kcal/h): (kcal/day)/hours of darkness/day. Health checks were performed daily, and no welfare-related issues were observed.
At the end of the experiment, rats were deprived of food after supplementation with Ch1, Ch2 or C and were killed by decapitation 1 h later between ZT1 and ZT2 (i.e., 1 and 2 h after the beginning of the light phase, respectively). Paired testes weights were recorded, and brains were dissected and immediately frozen in liquid nitrogen and stored at −80 °C until use. The hypothalamus and hippocampus were dissected at −20 °C on the day of RNA extraction to maintain RNA integrity. Figure 1 shows a graphical representation of the experimental design used in this study.
2.3. High-Performance Liquid Chromatography Coupled to Triple Quadrupole (LC-QqQ)
Serum testosterone T3 and T4 concentrations were measured using high-performance liquid chromatography coupled with triple quadrupole (LC-QqQ) following the method and using the equipment described by Domenech-Coca et al. 2019 [25]. Samples were prepared by mixing 50 μL serum with 250 μL methanol containing the internal standard (2 ng/mL). The mixture was then vortexed and centrifuged for 5 min at 4 °C at 20,000× g. The supernatant was mixed with 700 μL of 0.1% formic acid in water and subjected to solid-phase extraction using an Oasis HLB 96-well plate (Waters, Milford, MA, USA) previously conditioned with methanol and 0.1% formic acid in Milli-Q water. The cartridge was washed with 0.1% formic acid in Milli-Q water and dried under a high vacuum. The compounds were eluted with 500 μL methanol. Samples were evaporated in a SpeedVac at 45 °C, reconstituted with 50 μL Milli-Q water: methanol (60:40, v/v) and transferred to a glass vial for analysis. Then, 5 µL of solution was injected into a spectrometry UHPLC 1290 Infinity II Series (Agilent Technologies, Santa Clara, CA, USA) coupled to a QqQ/MS 6490 Series (Agilent Technologies) using a C18 analytical column (Zorbax Eclipse, 150 × 2.1 mm, Agilent Technologies). The chromatographic separation was performed with a gradient (40% of B for 0.5 min, 40–80% of B for 10.5 min, 80–100% for 0.1 min, 100% for 1.1 min, 100–40% for 0.1, and 40% for 3.5 min); mobile phase A was water with 0.5% acetic acid and 10 mM ammonium acetate, while B was 100% methanol. The column temperature was set at 45 °C and the flow rate was 3 mL/min. Standards and internal standards (3,3′,5’-triiodo-L-thyronine-13C6 solution, L-thyroxine-d4 and testosterone d3 for T3 and T4 and testosterone, respectively) were dissolved in methanol. Calibration curves were constructed in the ranges 0.05–10 ng/mL for testosterone, 0.05–5 ng/mL for T3 and 1–100 ng/mL for T4.
2.4. Gene Expression Analysis in the Hypothalamus and Hippocampus
Approximately 70–80 mg of hypothalamus and hippocampus tissues were dissected from each rat brain, transferred to cell disruption buffer and homogenized with 1.5 mm homogenization beads (Triple-Pure™, Molecular Biology Grade Zirconium Beads) using the BeadBug microtube homogenizer. After homogenization, RNA extraction was performed according to the manufacturer’s protocol using the PARIS™ kit (Invitrogen, Thermo Fisher Scientific, Inc., Waltham, MA, USA). The RNA was quantified using a Nanodrop™ spectrophotometer (Nanodrop Lite; Thermo Fisher Scientific). RNA (500 ng) was used to synthesize cDNA using a High-Capacity cDNA Reverse Transcription Kit, and qPCR was performed using the Bio-Rad real-time system CFX96 to amplify gene expression levels with the following parameters: 40 cycles of 2 min at 50 °C, 5 min at 95 °C, 10 s at 95 °C and 30 s at 60 °C. mRNA levels were normalized to D-Box, and fold changes were calculated using the 2−ΔΔCt method. The primers used to amplify these genes are listed in Supplementary Table S1.
2.5. Statistical Analysis
Data are expressed as mean ± standard deviation (SD). One-way ANOVA (Tukey’s post-hoc test) was used to determine the differences between the groups (p < 0.05). Two-way repeated-measures ANOVA (Tukey’s post-hoc test) analysis of Photoperiod x Fruit was used to assess the differences between groups (F being fruit and P being photoperiod effect, with PxF being the effect of the interaction). The interaction was considered statistically significant with a p value of p < 0.05. Statistical tests were performed using the SPSS Statistics 22 software (SPSS Inc., Chicago, IL, USA). Outliers were identified using SPSS using Tukey’s method and were removed before statistical analysis.
3. Results
3.1. Effects of Photoperiod on Body Weight, Eating Pattern Index and Serum Testosterone Levels in F344 Rats
The photoperiod responsiveness of F344 rats was assessed using body weight, EPI, testes weight, serum testosterone and the T4/T3 ratio (Table 1). After 11 weeks of photoperiod exposure, two-way ANOVA revealed a significant difference in body weight between the groups (P, p < 0.05), with a 6% lower body weight in L6-C than in L12-C (Tukey’s post hoc test, p = 0.06). Within the photoperiod groups, no significant difference was found in cherry consumption; however, L6-Ch1 showed a significant decrease in body weight compared with L12-Ch1 (p < 0.05). To assess eating behavior, EPI was calculated for all groups and represented as kilocalories consumed by the rats and the hours of darkness when the animals were active and fed (Table 1). EPI was significantly affected by the photoperiod as follows: L6-C < L12-C < L18-C (P, two-way ANOVA, p < 0.05). Similar to body weight, no differences were found in EPI between Ch1 or Ch2.
No significant difference was found in paired testes weights (Table 1); however, two-way ANOVA revealed a significant interaction between photoperiod and cherry consumption for serum testosterone levels (P, PxF, two-way ANOVA, p < 0.05). Specifically, testosterone concentrations were significantly lower in L18-C than in L6-C and L12-C (Table 1). Ch1 increased serum testosterone levels in the L6-Ch1 and L12-Ch1 groups compared with their control groups (the L6-C and L12-C groups, respectively); however, no effects of cherry consumption were found in the L18 photoperiod.
Circulating levels of thyroid hormones in serum (T4/T3 ratio) were affected by photoperiod and fruit (PxF, two-way ANOVA, p < 0.05), with the lowest T4/T3 ratio in L6-C compared with the other photoperiod control groups (Table 1). Moreover, Ch1 significantly increased the T4:T3 ratio in F344 rats exposed to the L6 photoperiod compared with L6-C animals (Table 1).
3.2. Hypothalamic Gene Expression Responds to Photoperiod
Previous studies have shown a range of photoperiodically regulated genes in the hypothalamus in F344 rats, including genes involved in thyroid hormone and retinoic acid signaling [26,27,28]. Here, we tested the key genes involved in photoperiodic time measurements and assessed whether they were also regulated by cherry consumption. Hypothalamic Dio2 expression did not show photoperiod responsiveness in the control groups after 11 weeks of photoperiod exposure; however, Ch1 consumption significantly upregulated Dio2 expression in F344 rats exposed to the L6 photoperiod compared with all other groups, except L12-C (Figure 2A; two-way ANOVA P, PxF p < 0.05). Dio3 was downregulated in L18-C compared with the L6-C and L12-C groups and in L12-Ch2 compared with L12-C (Figure 2B) (P, F two-way ANOVA, p < 0.05).
An interaction between photoperiod and fruit was found in the expression levels of Retinaldehyde dehydrogenase 1 (Raldh1) (PxF, two-way ANOVA, p < 0.05); however, we did not find differences in photoperiod (Figure 2C). After Ch1 consumption, hypothalamic Raldh1 expression levels were downregulated in the L12-Ch1 and L18-Ch1 groups compared with the L6-Ch1 group.
Furthermore, mRNA expression levels of hypothalamic Growth-hormone-releasing hormone (Ghrh) and Somatostatin (Sst) were analyzed. Ghrh mRNA levels were strongly affected by photoperiod, with lower levels in L6-C than in L12-C and L18-C; however, this effect disappeared after the consumption of both cherries (L6-Ch1 and L6-Ch2 groups) (Figure 2D). Interestingly, Ghrh mRNA expression was downregulated by the consumption of Ch2 in L18, reaching expression levels similar to those in the L6-C group (Figure 2D). Two-way ANOVA revealed an effect of photoperiod and an interaction between photoperiod and fruit (two-way ANOVA; PxF, P, p < 0.05). Sst levels were downregulated in L18-C compared with in those L12-C and L6-C groups. Moreover, the Ch2 group showed downregulation of Sst mRNA compared with all other groups (two-way ANOVA, F, p < 0.05). Hence, we found that both genes not only respond to photoperiod but also to sweet cherry consumption within a specific photoperiod, as Ch2 exacerbated the downregulation of Ghrh and Sst in the L18 group.
3.3. Sweet Cherry Consumption Exerts a Hippocampal Neuroprotective Effect, Which Was Modulated by Specific Phenolic Hallmarks and Photoperiod
Next, we investigated the possible neuroprotective effects of seasonal consumption of sweet cherries with different phenolic hallmarks on gene expression markers in the hippocampus. Growing evidence suggests that the consumption of fruit is associated with neuroprotective effects [29,30], but reports on the effect of photoperiod on neuroprotection are less common. Bdnf mRNA levels were modulated by P, F and PxF (two-way ANOVA, p < 0.05), with Ch2 consumption strongly upregulating Bdnf expression compared with all other groups (Figure 3A). Nrf2 expression levels were affected by fruit consumption and photoperiod (PxF, p < 0.05). The L18-Ch2 group showed higher expression levels compared with the L18-Ch1 group (Figure 3B). Neuronal differentiation 1 (NeuroD) was regulated by photoperiod, with lower NeuroD expression in the L18-C group than in the L6-C group; however, Ch2 consumption significantly upregulated NeuroD expression compared with all other groups (Figure 3C). In addition, superoxide dismutase type 1 (Sod1) was significantly affected by photoperiod and was upregulated in the L6-C group compared with the L12-C and L18-C groups. Consumption of Ch1 significantly downregulated Sod1 levels, whereas Ch2 consumption maintained the expression levels of Sod1 (Figure 3D). Glutathione peroxidase 1 (Gpx1) showed a similar expression pattern to Sod1; however, Ch2 consumption significantly upregulated Sod1 expression compared with L18-C and Ch1 (Figure 3E).
4. Discussion
In this study, we investigated the effects of the consumption of seasonal cherry, which is rich in anthocyanins, on the activation of neuroprotective pathways in the hippocampus of photoperiod-sensitive F344 rats. Two cherries obtained from different geographical regions were selected for the study in order to obtain cherries with different chemical compositions. We showed that the markers of neuroprotection are strongly regulated by photoperiod and/or seasonal fruit consumption.
The consumption of fruit is associated with several beneficial health effects, including the reduction of blood pressure and cell damage through its antioxidant effects [29,30]. Phenolic compounds are one of the main compounds responsible for the health benefits of fruit, which can have antioxidant and anti-inflammatory effects [31]. Specifically, it has been reported that anthocyanins, found in cherry and blueberry for example, have excellent antioxidant properties, improving oxidative stress and neurodegeneration, among others [32]. Importantly, phenolic compounds can cross the blood–brain barrier [33,34]. The phenolic compounds quercetin and naringin have been shown to promote neuroprotection in the dentate gyrus of the hippocampus [35], which is the main brain region that hosts neurogenic niches [36]. In support, the consumption of blueberries, a fruit rich in phenolic compounds, exerts positive effects on the brain by enhancing memory and learning in animals and humans [37].
Firstly, to evaluate the photoperiod responsiveness of the F344 rats in this study, we analyzed physiological parameters (body weight, eating pattern), photoperiod-responsive hormones and photoperiod-regulated genes in the hypothalamus. As expected, we found that body weight and feeding behavior responded to the photoperiod. These results are consistent with previous studies in F344 rats showing that body weight and food intake are strongly regulated by photoperiod, with lower body weight and food intake in short photoperiods (L6) compared with long photoperiods (L18) [2,16,22]. However, compared with these studies, the effect of photoperiod on body weight in our study was more subtle. Differences between studies could be attributed to photoperiod refractoriness, as this study was carried out during eleven weeks of photoperiods. Refractoriness is a physiological response of seasonal species such as hamsters and sheep to a change in photoperiod that spontaneously reverts to the initial state, even if the photoperiod remains unchanged [38]. Another reason could be that different sub-strains of F344 respond differently to photoperiod; for example, F344/NHsd rats are more sensitive to photoperiod than F344⁄NCrHsd [39]. Interestingly, Ch1 exacerbated the photoperiod effect by decreasing body weight under a short photoperiod.
A shared mechanism of photoperiod responsiveness among different seasonal species is the change in hypothalamic thyroid hormone availability, depending on the photoperiod of exposure [1]. Here, we show that hypothalamic thyroid signaling was not only affected by photoperiod but also varied depending on cherry consumption. It has been widely described that the gene expression of Dio2 and Dio3 enzymes in tanycytes regulates hypothalamic thyroid hormone availability by converting between biologically active T3 and inactive T4 [40]. In our study, only hypothalamic Dio3 expression was regulated by photoperiod, with a downregulation in the long photoperiod as expected [28,41,42]. Species-specific differences in Dio2 and Dio3 expression have been discussed previously [40], although previous studies in F344 rats indicated a photoperiod response with upregulation of Dio2 in long photoperiods [2,27]. However, in these studies, the maximum photoperiod exposure was four weeks, whereas, in our study, rats were exposed to a photoperiod for eleven weeks, and photoperiod refractoriness could not be ruled out.
Intra-hypothalamic thyroid hormone metabolism plays an important role in seasonal response [43], but the seasonal pattern of serum thyroid hormone levels remains controversial. According to a clinical study in 700,000 people, only thyroid-stimulating hormone levels showed a seasonal rhythm, whereas T3 and T4 levels were not affected [44]. Nevertheless, another study with over 7000 volunteers showed monthly changes in serum T3 and T4 levels [45]. It has been suggested that diets rich in fruits (i.e., rich in (poly)phenols) have beneficial effects on thyroid hormones in population studies [46]. Here, we show that the T4/T3 ratio increased with the consumption of Ch1 under short photoperiods, suggesting that the effect of cherry consumption on the thyroid hormones depends on photoperiod in F344 rats and on chemical composition. Different studies have shown that phenolic compounds can act as activators or inhibitors of the thyroperoxidase enzyme, which is involved in thyroid hormone synthesis [47,48]. Thus, the effect of Ch1 could be the result of the sum of all the inhibitory and activating actions produced by several phenolic compounds.
RALDH1 is involved in retinoic acid synthesis, and its absence is associated with decreased food intake and body weight in F344 rats [26]. Our results contrast with those of other studies carried out in F344 rats exposed to photoperiods for three and fourteen days, where Raldh1 showed a rapid and strong photoperiod effect and was downregulated under short photoperiods [26]. The differences between the two studies could be attributed to the duration of photoperiod exposure, as photoperiod refractoriness could be involved in this pathway. The growth hormone axis is seasonally regulated, and its increase is associated with an increase in day length in seasonal species [49]. Growth hormone is involved in the promotion of anabolic functions when nutrients are available. In contrast, SST inhibits its activity as a feedback regulatory mechanism [50]. Different nutrients, such as amino acids, vitamins and minerals, play a role in the regulation of the growth axis [50], and it has been suggested that a Mediterranean diet, rich in fruits and vegetables, is correlated with a peak in growth hormone in humans [51]. In this study, we demonstrated that Ghrh and Sst expressions were regulated by photoperiod in F344 rats and that the consumption of Ch2 was associated with decreased expression. These effects could be attributed to the fact that Ch2 contained more flavonols and hydrobenzoic acid derivatives (unpublished data).
In mammals, reproduction is strongly modulated by photoperiod [52]. For example, the testes weight of F344 rats decreases during the first weeks of a short photoperiod, although these changes disappear after 10 weeks of photoperiod exposure [53]. These findings agree with the results observed in the present study, since no differences were observed in testes mass after 11 weeks of photoperiod exposure. We observed that testosterone levels are modulated by Ch1 but not Ch2 in the L6 and the L12 photoperiods, which could be linked to the different compositions of the two sweet cherries. These differential effects may be due to Ch1′s higher concentration of cyanidin derivatives and catechin than Ch2 (unpublished data), which may be the result of the harvesting conditions in different geographical areas. Moreover, sex hormones have been correlated with neuroprotection in neuronal cell models; however, in the presence of oxidative stress, they increase cell loss in both neuronal and glial cells [54]. This evidence allowed us to evaluate photoperiod responsiveness in this study to later focus on the neuroprotective role of cherries with different origins and compositions.
Among the factors that regulate hippocampal homeostasis, diets rich in fruits and vegetables have been associated with improvements in cognition and working memory [55]. For instance, the consumption of tart cherry in aged F344 rats decreases the expression of neuroinflammatory markers such as NADPH oxidase and cyclooxygenase 2 [56]. Other studies have observed that phenolic compounds, such as quercetin and kaempferol, activate key proteins involved in neuroprotection in the hippocampus, including NRF2 and BDNF. NRF2 acts in the nucleus by inducing the transcription of genes containing a specific region called the antioxidant response element (ARE) [57]. This signaling promotes the expression of antioxidant enzymes such as GPX1 and SOD by reducing cell damage and the production of reactive oxygen species [35]. Neuroprotection in the brain is correlated with the antioxidant enzymatic activity of a subset of proteins, which can be stimulated by external factors such as the intake of fruits rich in bioactive compounds [58]. Additionally, thyroid hormones have been observed to play an important role in adult neurogenesis in the hippocampus, promoting cell survival and upregulating the expression of genes involved in cell differentiation, such as NeuroD [59]. Considering that thyroid hormones play a pivotal role in generating seasonal rhythms, it is feasible to hypothesize that neuronal function in the hippocampus is affected by photoperiod. In support, we recently showed that a short photoperiod impaired memory in F344 rats [22]. In the current study, hippocampal Bdnf, Nrf2, NeuroD and Gpx1 expression levels were upregulated when rats consumed Ch2 in the L18 photoperiod, promoting hippocampal neuroprotection and stimulating the expression of genes involved in antioxidant signaling, suggesting that cherry effects are regulated by photoperiod. Given that Ch1 did not exert any effects, the compositions of the sweet cherries must be pivotal in their different effects. The phenolic compositions of the two cherries were different: Ch1 had a higher content of total polyphenol, anthocyanin and flavanol than Ch2, while Ch2 showed a higher flavonol content than Ch1 [11]. In addition, Ch2 had a higher content of quercetin, quercetin derivatives and kaempferol derivatives than Ch1 (data not published). As mentioned above, quercetin and kaempferol can activate proteins involved in hippocampal neuroprotection [57]. Thus, these flavonols may be involved in the neuroprotective effects of Ch2. Previously, we showed that lipid peroxidation markers due to free radicals are reduced by the consumption of both cherries in L18, with one of the cherries being more effective in lowering these markers, highlighting that the composition of the cherry influences the differential response [11]. Moreover, the effects could be different from one organ to another because, in the case of the brain, the blood–brain barrier is highly selective. It has been observed that metabolites derived from the digestion of (poly)phenols cross the blood–brain barrier depending on their molecular structure, and their metabolism in the endothelial layer produces a variety of metabolites with additional bioactive properties [33]. While a previous study did not show a photoperiod response in Bdnf mRNA expression in the hippocampus of hamsters exposed to short or long photoperiods [60], clinical studies have shown that circulating markers of neuroprotection, such as BDNF, show a seasonal pattern [61]. Moreover, we showed that testosterone levels were strongly increased by cherry consumption in short and intermediate photoperiods, and it has been previously suggested that the consumption of anthocyanins is associated with a reduction in oxidative-stress-induced testosterone deficiency [62]. To the best of our knowledge, this is the first study showing photoperiod-dependent regulation of neuroprotective markers in the hippocampus with a specific type of sweet cherry.
5. Conclusions
In conclusion, seasonal consumption of sweet cherry with a specific composition could play an important role in the hippocampal activation of neuroprotection in a photoperiod-dependent manner by increasing the expression markers of neuroprotection in a long photoperiod. However, further research is needed to elucidate how the time of year could affect the benefits of fruit consumption in humans by improving neuroprotective markers, including clinical trials and bioavailability studies to elucidate the possible phenolic metabolites responsible for the observed effects.
Supplementary Materials
The following supporting information can be downloaded at https://www.mdpi.com/article/10.3390/antiox13010072/s1. Table S1: Primer nucleotide sequences used for real-time quantitative PCR.
Author Contributions
Conceptualization, G.H., F.I.B. and B.M.; formal analysis, F.M.; funding acquisition, F.I.B., B.M. and F.M.; investigation, F.M.; methodology, F.M.; supervision, G.H., F.I.B. and B.M.; writing—original draft, F.M.; writing—review and editing, F.M., F.I.B., B.M. and G.H. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by grant number PID2020-113739RB-I00 funded by MCIN/AEI/10.13039/501100011033 and by Pect-Nutrisalt funded by the European Regional Development Fund of the European Commission through the Operative Program Erdf of Catalonia 2014–2020. The authors thank the British Society for Neuroendocrinology (BSN) for providing a research visit Grant to F.M (Grant number: BSN-2022-1452). F.M. is the recipient of a predoctoral fellowship from Universitat Rovira i Virgili—Martí i Franquès (Grant number: 2019PMF-PIPF-19).
Institutional Review Board Statement
The animal experiment was conducted in accordance with the European Communities Council Directive (86/609/EEC) and approved by the Animal Ethics Review Committee for Animal Experimentation of the Universitat Rovira i Virgili and by Generalitat de Catalunya (permission number 9495, FUE-2017-00499873).
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are contained within the article and the Supplementary Materials.
Acknowledgments
The authors thank Niurka Dariela Llópiz and Rosa Maria Pastor, from the Universitat Rovira i Virgili, for their valuable support in the laboratory. F.I.B is a Serra Húnter Fellow.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Helfer, G.; Barrett, P.; Morgan, P.J. A unifying hypothesis for control of body weight and reproduction in seasonally breeding mammals. J. Neuroendocrinol. 2019, 31, e12680. [Google Scholar] [CrossRef] [PubMed]
- Tavolaro, F.M.; Thomson, L.M.; Ross, A.W.; Morgan, P.J.; Helfer, G. Photoperiodic effects on seasonal physiology, reproductive status and hypothalamic gene expression in young male F344 rats. J. Neuroendocrinol. 2015, 27, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Elizondo-Vega, R.J.; Recabal, A.; Oyarce, K. Nutrient sensing by hypothalamic tanycytes. Front. Endocrinol. 2019, 10, 244. [Google Scholar] [CrossRef] [PubMed]
- Langlet, F. Tanycyte gene expression dynamics in the regulation of energy homeostasis. Front. Endocrinol. 2019, 10, 286. [Google Scholar] [CrossRef] [PubMed]
- Ibars, M.; Aragonès, G.; Ardid-Ruiz, A.; Gibert-Ramos, A.; Arola-Arnal, A.; Suárez, M.; Bladé, C. Seasonal consumption of polyphenol-rich fruits affects the hypothalamic leptin signaling system in a photoperiod-dependent mode. Sci. Rep. 2018, 8, 13572. [Google Scholar] [CrossRef]
- Arola-Arnal, A.; Cruz-Carrión, Á.; Torres-Fuentes, C.; Ávila-Román, J.; Aragonès, G.; Mulero, M.; Bravo, F.I.; Muguerza, B.; Arola, L.; Suárez, M. Chrononutrition and polyphenols: Roles and diseases. Nutrients 2019, 11, 2602. [Google Scholar] [CrossRef]
- Kimble, R.; Keane, K.M.; Lodge, J.K.; Cheung, W.; Haskell-Ramsay, C.F.; Howatson, G. Polyphenol-rich tart cherries (Prunus Cerasus, cv Montmorency) improve sustained attention, feelings of alertness and mental fatigue and influence the plasma metabolome in middle-aged adults: A randomised, placebo-controlled trial. Br. J. Nutr. 2022, 128, 2409–2420. [Google Scholar] [CrossRef]
- Shukitt-Hale, B.; Kelly, M.; Bielinski, D.; Fisher, D. Tart cherry extracts reduce inflammatory and oxidative stress signaling in microglial cells. Antioxidants 2016, 5, 33. [Google Scholar] [CrossRef]
- Rendeiro, C.; Vauzour, D.; Kean, R.J.; Butler, L.T.; Rattray, M.; Spencer, J.P.E.; Williams, C.M. Blueberry supplementation induces spatial memory improvements and region-specific regulation of hippocampal BDNF mRNA expression in young rats. Psychopharmacology 2012, 223, 319–330. [Google Scholar] [CrossRef]
- Bruna, B.; Lobos, P.; Herrera-Molina, R.; Hidalgo, C.; Paula-Lima, A.; Adasme, T. The signaling pathways underlying BDNF-induced Nrf2 hippocampal nuclear translocation involve ROS, RyR-Mediated Ca2+ signals, ERK and PI3K. Biochem. Biophys. Res. Commun. 2018, 505, 201–207. [Google Scholar] [CrossRef]
- Cruz-Carrión, Á.; Ruiz de Azua, M.J.; Mulero, M.; Arola-Arnal, A.; Suárez, M. Oxidative stress in rats is modulated by seasonal consumption of sweet cherries from different geographical origins: Local vs. non-local. Nutrients 2020, 12, 2854. [Google Scholar] [CrossRef] [PubMed]
- Ferretti, G.; Bacchetti, T.; Belleggia, A.; Neri, D. Cherry antioxidants: From farm to table. Molecules 2010, 15, 6993–7005. [Google Scholar] [CrossRef] [PubMed]
- Fonseca, L.R.S.; Silva, G.R.; Luís, Â.; Cardoso, H.J.; Correia, S.; Vaz, C.V.; Duarte, A.P.; Socorro, S. Sweet cherries as anti-cancer agents: From bioactive compounds to function. Molecules 2021, 26, 2941. [Google Scholar] [CrossRef] [PubMed]
- Martini, S.; Conte, A.; Tagliazucchi, D. Phenolic compounds profile and antioxidant properties of six sweet cherry (Prunus avium) cultivars. Food Res. Int. 2017, 97, 15–26. [Google Scholar] [CrossRef] [PubMed]
- Iglesias-Carres, L.; Mas-Capdevila, A.; Bravo, F.I.; Mulero, M.; Muguerza, B.; Arola-Arnal, A. Optimization and characterization of Royal Dawn cherry (Prunus avium) phenolics extraction. Sci. Rep. 2019, 9, 17626. [Google Scholar] [CrossRef] [PubMed]
- Ruiz de Azua, M.J.; Manocchio, F.; Cruz-Carrión, Á.; Arola-Arnal, A.; Torres-Fuentes, C.; Bernal, C.A.; Saín, J.; Suarez, M. Fatty acid metabolism in liver and muscle is strongly modulated by photoperiod in Fischer 344 rats. J. Photochem. Photobiol. B Biol. 2023, 238, 112621. [Google Scholar] [CrossRef] [PubMed]
- Chockchaisawasdee, S.; Golding, J.B.; Vuong, Q.V.; Papoutsis, K.; Stathopoulos, C.E. Sweet cherry: Composition, postharvest preservation, processing and trends for its future use. Trends Food Sci. Technol. 2016, 55, 72–83. [Google Scholar] [CrossRef]
- Blando, F.; Oomah, B.D. Sweet and sour cherries: Origin, distribution, nutritional composition and health benefits. Trends Food Sci. Technol. 2019, 86, 517–529. [Google Scholar] [CrossRef]
- AOAC International. Official Methods of Analysis, 15th ed.; Helrich, K., Ed.; Association of Official Analytical Chemists: Arlington, VA, USA, 1990; Volume 1. [Google Scholar]
- Arnous, A.; Makris, D.P.; Kefalas, P. Correlation of pigment and flavanol content with antioxidant properties in selected aged regional wines from Greece. J. Food Compos. Anal. 2002, 15, 655–665. [Google Scholar] [CrossRef]
- Cacace, J.E.; Mazza, G. Optimization of extraction of anthocyanins from black currants with aqueous ethanol. J. Food Sci. 2003, 68, 240–248. [Google Scholar] [CrossRef]
- McLean, S.L.; Yun, H.; Tedder, A.; Helfer, G. The effect of photoperiod and high fat diet on the cognitive response in photoperiod-sensitive F344 rats. Physiol. Behav. 2021, 239, 113496. [Google Scholar] [CrossRef] [PubMed]
- Navarro-Masip, È.; Manocchio, F.; Rodríguez, R.M.; Bravo, F.I.; Torres-Fuentes, C.; Muguerza, B.; Aragonès, G. Photoperiod-dependent effects of grape-seed proanthocyanidins on adipose tissue metabolic markers in healthy rats. Mol. Nutr. Food Res. 2023, 67, 2300035. [Google Scholar] [CrossRef] [PubMed]
- Mariné-Casadó, R.; Domenech-Coca, C.; del Bas, J.M.; Bladé, C.; Caimari, A.; Arola, L. Cherry consumption out of season alters lipid and glucose homeostasis in normoweight and cafeteria-fed obese Fischer 344 rats. J. Nutr. Biochem. 2019, 63, 72–86. [Google Scholar] [CrossRef] [PubMed]
- Domenech-Coca, C.; Mariné-Casadó, R.; Caimari, A.; Arola, L.; del Bas, J.M.; Bladé, C.; Rodriguez-Naranjo, M.I. Dual liquid-liquid extraction followed by LC-MS/MS method for the simultaneous quantification of melatonin, cortisol, triiodothyronine, thyroxine and testosterone levels in serum: Applications to a photoperiod study in rats. J. Chromatogr. B 2019, 1108, 11–16. [Google Scholar] [CrossRef] [PubMed]
- Helfer, G.; Ross, A.W.; Russell, L.; Thomson, L.M.; Shearer, K.D.; Goodman, T.H.; McCaffery, P.J.; Morgan, P.J. Photoperiod regulates vitamin A and Wnt/β-catenin signaling in F344 Rats. Endocrinology 2012, 153, 815–824. [Google Scholar] [CrossRef] [PubMed]
- Ross, A.W.; Helfer, G.; Russell, L.; Darras, V.M.; Morgan, P.J. Thyroid hormone signalling genes are regulated by photoperiod in the hypothalamus of F344 rats. PLoS ONE 2011, 6, e21351. [Google Scholar] [CrossRef] [PubMed]
- Helfer, G.; Ross, A.W.; Morgan, P.J. Neuromedin U partly mimics thyroid-stimulating hormone and triggers Wnt/β-Catenin signalling in the photoperiodic response of F344 rats. J. Neuroendocrinol. 2013, 25, 1264–1272. [Google Scholar] [CrossRef]
- Habauzit, V.; Morand, C. Evidence for a protective effect of polyphenols-containing foods on cardiovascular health: An update for clinicians. Ther. Adv. Chronic Dis. 2012, 3, 87–106. [Google Scholar] [CrossRef]
- Lapuente, M.; Estruch, R.; Shahbaz, M.; Casas, R. Relation of fruits and vegetables with major cardiometabolic risk factors, Markers of oxidation, and inflammation. Nutrients 2019, 11, 2381. [Google Scholar] [CrossRef]
- Cosme, F.; Pinto, T.; Aires, A.; Morais, M.C.; Bacelar, E.; Anjos, R.; Ferreira-Cardoso, J.; Oliveira, I.; Vilela, A.; Gonçalves, B. Red fruits composition and their health benefits—A Review. Foods 2022, 11, 644. [Google Scholar] [CrossRef]
- Haminiuk, C.W.I.; Maciel, G.M.; Plata-Oviedo, M.S.V.; Peralta, R.M. Phenolic compounds in fruits—An overview. Int. J. Food Sci. Technol. 2012, 47, 2023–2044. [Google Scholar] [CrossRef]
- Figueira, I.; Menezes, R.; Macedo, D.; Costa, I.; dos Santos, C.N. Polyphenols beyond barriers: A glimpse into the brain. Curr. Neuropharmacol. 2017, 15, 562–594. [Google Scholar] [CrossRef] [PubMed]
- Ardid-Ruiz, A.; Harazin, A.; Barna, L.; Walter, F.R.; Bladé, C.; Suárez, M.; Deli, M.A.; Aragonès, G. The effects of Vitis vinifera L. phenolic compounds on a blood-brain barrier culture model: Expression of leptin receptors and protection against cytokine-induced damage. J. Ethnopharmacol. 2020, 247, 112253. [Google Scholar] [CrossRef] [PubMed]
- Moosavi, F.; Hosseini, R.; Saso, L.; Firuzi, O. Modulation of neurotrophic signaling pathways by polyphenols. Drug Des. Dev. Ther. 2015, 10, 23–42. [Google Scholar] [CrossRef]
- Jurkowski, M.P.; Bettio, L.; Woo, E.K.; Patten, A.; Yau, S.-Y.; Gil-Mohapel, J. Beyond the hippocampus and the SVZ: Adult neurogenesis throughout the brain. Front. Cell. Neurosci. 2020, 14, 576444. [Google Scholar] [CrossRef] [PubMed]
- Vauzour, D. Dietary polyphenols as modulators of brain functions: Biological actions and molecular mechanisms underpinning their beneficial effects. Oxid. Med. Cell. Longev. 2012, 2012, 914273. [Google Scholar] [CrossRef] [PubMed]
- Dardente, H.; Migaud, M. Thyroid hormone and hypothalamic stem cells in seasonal functions. Vitam. Horm. 2021, 116, 91–131. [Google Scholar] [CrossRef]
- Ross, A.W.; Johnson, C.E.; Bell, L.M.; Reilly, L.; Duncan, J.S.; Barrett, P.; Heideman, P.D.; Morgan, P.J. Divergent regulation of hypothalamic neuropeptide Y and Agouti-related protein by photoperiod in F344 rats with differential food intake and growth. J. Neuroendocrinol. 2009, 21, 610–619. [Google Scholar] [CrossRef]
- Helfer, G.; Dumbell, R. Endocrine drivers of photoperiod response. Curr. Opin. Endocr. Metab. Res. 2020, 11, 49–54. [Google Scholar] [CrossRef]
- Nakao, N.; Ono, H.; Yamamura, T.; Anraku, T.; Takagi, T.; Higashi, K.; Yasuo, S.; Katou, Y.; Kageyama, S.; Uno, Y.; et al. Thyrotrophin in the pars tuberalis triggers photoperiodic response. Nature 2008, 452, 317–322. [Google Scholar] [CrossRef]
- Hanon, E.A.; Lincoln, G.A.; Fustin, J.-M.; Dardente, H.; Masson-Pévet, M.; Morgan, P.J.; Hazlerigg, D.G. Ancestral TSH mechanism signals summer in a photoperiodic mammal. Curr. Biol. 2008, 18, 1147–1152. [Google Scholar] [CrossRef]
- Yoshimura, T. Thyroid hormone and seasonal regulation of reproduction. Front. Neuroendocrinol. 2013, 34, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Santi, D.; Spaggiari, G.; Brigante, G.; Setti, M.; Tagliavini, S.; Trenti, T.; Simoni, M. Semi-annual seasonal pattern of serum thyrotropin in adults. Sci. Rep. 2019, 9, 10786. [Google Scholar] [CrossRef] [PubMed]
- Yamada, S.; Horiguchi, K.; Akuzawa, M.; Sakamaki, K.; Shimomura, Y.; Kobayashi, I.; Andou, Y.; Yamada, M. Seasonal variation in thyroid function in over 7000 healthy subjects in an iodine-sufficient area and literature review. J. Endocr. Soc. 2022, 6, bvac054. [Google Scholar] [CrossRef]
- Bellastella, G.; Scappaticcio, L.; Caiazzo, F.; Tomasuolo, M.; Carotenuto, R.; Caputo, M.; Arena, S.; Caruso, P.; Maiorino, M.I.; Esposito, K. Mediterranean diet and thyroid: An interesting alliance. Nutrients 2022, 14, 4130. [Google Scholar] [CrossRef]
- de Souza dos Santos, M.C.; Gonçalves, C.F.L.; Vaisman, M.; Ferreira, A.C.F.; de Carvalho, D.P. Impact of flavonoids on thyroid function. Food Chem. Toxicol. 2011, 49, 2495–2502. [Google Scholar] [CrossRef] [PubMed]
- Habza-Kowalska, E.; Kaczor, A.A.; Bartuzi, D.; Piłat, J.; Gawlik-Dziki, U. Some dietary phenolic compounds can activate thyroid peroxidase and inhibit lipoxygenase-preliminary study in the model systems. Int. J. Mol. Sci. 2021, 22, 5108. [Google Scholar] [CrossRef]
- Dumbell, R. An appetite for growth: The role of the hypothalamic—Pituitary—Growth hormone axis in energy balance. J. Neuroendocrinol. 2022, 34, e13133. [Google Scholar] [CrossRef]
- Caputo, M.; Pigni, S.; Agosti, E.; Daffara, T.; Ferrero, A.; Filigheddu, N.; Prodam, F. Regulation of GH and GH Signaling by Nutrients. Cells 2021, 10, 1376. [Google Scholar] [CrossRef]
- Muscogiuri, G.; Barrea, L.; Laudisio, D.; Di Somma, C.; Pugliese, G.; Salzano, C.; Colao, A.; Savastano, S. Somatotropic axis and obesity: Is there any role for the Mediterranean diet? Nutrients 2019, 11, 2228. [Google Scholar] [CrossRef]
- Dardente, H.; Wood, S.; Ebling, F.; Sáenz de Miera, C. An integrative view of mammalian seasonal neuroendocrinology. J. Neuroendocrinol. 2019, 31, e12729. [Google Scholar] [CrossRef] [PubMed]
- Shoemaker, M.B.; Heideman, P.D. Reduced body mass, food intake, and testis size in response to short photoperiod in adult F344 rats. BMC Physiol. 2002, 2, 11. [Google Scholar] [CrossRef] [PubMed]
- Duong, P.; Tenkorang, M.A.A.; Trieu, J.; McCuiston, C.; Rybalchenko, N.; Cunningham, R.L. Neuroprotective and neurotoxic outcomes of androgens and estrogens in an oxidative stress environment. Biol. Sex Differ. 2020, 11, 12. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Huang, J.; Song, D.; Deng, R.; Wei, J.; Zhang, Z. Increased consumption of fruit and vegetables is related to a reduced risk of cognitive impairment and dementia: Meta-analysis. Front. Aging Neurosci. 2017, 9, 18. [Google Scholar] [CrossRef] [PubMed]
- Thangthaeng, N.; Poulose, S.M.; Gomes, S.M.; Miller, M.G.; Bielinski, D.F.; Shukitt-Hale, B. Tart cherry supplementation improves working memory, hippocampal inflammation, and autophagy in aged rats. Age 2016, 38, 393–404. [Google Scholar] [CrossRef] [PubMed]
- Moratilla-Rivera, I.; Sánchez, M.; Valdés-González, J.A.; Gómez-Serranillos, M.P. Natural products as modulators of Nrf2 signaling pathway in neuroprotection. Int. J. Mol. Sci. 2023, 24, 3748. [Google Scholar] [CrossRef] [PubMed]
- Babazadeh, A.; Vahed, F.M.; Liu, Q.; Siddiqui, S.A.; Kharazmi, M.S.; Jafari, S.M. Natural bioactive molecules as neuromedicines for the treatment/prevention of neurodegenerative diseases. ACS Omega 2023, 8, 3667–3683. [Google Scholar] [CrossRef]
- Kapoor, R.; Fanibunda, S.E.; Desouza, L.A.; Guha, S.K.; Vaidya, V.A. Perspectives on thyroid hormone action in adult neurogenesis. J. Neurochem. 2015, 133, 599–616. [Google Scholar] [CrossRef]
- Ikeno, T.; Weil, Z.M.; Nelson, R.J. Photoperiod affects the diurnal rhythm of hippocampal neuronal morphology of siberian hamsters. Chronobiol. Int. 2013, 30, 1089–1100. [Google Scholar] [CrossRef]
- Molendijk, M.L.; Haffmans, J.P.M.; Bus, B.A.A.; Spinhoven, P.; Penninx, B.W.J.H.; Prickaerts, J.; Voshaar, R.C.O.; Elzinga, B.M. Serum BDNF concentrations show strong seasonal variation and correlations with the amount of ambient sunlight. PLoS ONE 2012, 7, e48046. [Google Scholar] [CrossRef]
- Hu, J.; Li, X.; Wu, N.; Zhu, C.; Jiang, X.; Yuan, K.; Li, Y.; Sun, J.; Bai, W. Anthocyanins prevent AAPH-induced steroidogenesis disorder in leydig cells by counteracting oxidative stress and StAR abnormal expression in a structure-dependent manner. Antioxidants 2023, 12, 508. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Experimental design and timeline. F344 rats were acclimatized to L6 (6 h light/day), L12 (12 h light/day) or L18 (18 h light/day) on a chow diet for four weeks. Rats in each photoperiod group (n = 24 per photoperiod) were randomly assigned to three groups. One group in each photoperiod was provided with chow and either Cherry 1 from Spain, Cherry 2 from Chile or a sugar-content-matched control. The experiment was completed after seven weeks of photoperiod and diet intervention.
Figure 1.
Experimental design and timeline. F344 rats were acclimatized to L6 (6 h light/day), L12 (12 h light/day) or L18 (18 h light/day) on a chow diet for four weeks. Rats in each photoperiod group (n = 24 per photoperiod) were randomly assigned to three groups. One group in each photoperiod was provided with chow and either Cherry 1 from Spain, Cherry 2 from Chile or a sugar-content-matched control. The experiment was completed after seven weeks of photoperiod and diet intervention.

Figure 2.
Hypothalamic (A) deiodinase 2 (Dio2), (B) deiodinase 3 (Dio3), (C) retinaldehyde dehydrogenase 1 (Raldh1), (D) growth hormone-releasing hormone (Ghrh) and (E) somatostatin (Sst) mRNA expression in Fisher 344 rats exposed to L6 (6 h light/18 h dark), L12 (12 h light/12 h dark) or L18 (18 h light/6 h dark) photoperiods for eleven weeks and administered vehicle (C), cherry 1 (Ch1, 100 mg/kg body weight) or cherry 2 (Ch2, 100 mg/kg body weight) during the last seven weeks. Values are expressed as mean ± standard deviation and normalized relative to the L12-C group (n = 8 per group). P, F and PxF indicate significant differences produced by photoperiod, fruit and the interaction between photoperiod and fruit, respectively (two-way ANOVA, p < 0.05). Different letters above the bars indicate significant differences by post-hoc Tukey’s test (one-way ANOVA, p < 0.05). (F) Schematic representation of the brain. The region included in the red square shows the analyzed brain region.
Figure 2.
Hypothalamic (A) deiodinase 2 (Dio2), (B) deiodinase 3 (Dio3), (C) retinaldehyde dehydrogenase 1 (Raldh1), (D) growth hormone-releasing hormone (Ghrh) and (E) somatostatin (Sst) mRNA expression in Fisher 344 rats exposed to L6 (6 h light/18 h dark), L12 (12 h light/12 h dark) or L18 (18 h light/6 h dark) photoperiods for eleven weeks and administered vehicle (C), cherry 1 (Ch1, 100 mg/kg body weight) or cherry 2 (Ch2, 100 mg/kg body weight) during the last seven weeks. Values are expressed as mean ± standard deviation and normalized relative to the L12-C group (n = 8 per group). P, F and PxF indicate significant differences produced by photoperiod, fruit and the interaction between photoperiod and fruit, respectively (two-way ANOVA, p < 0.05). Different letters above the bars indicate significant differences by post-hoc Tukey’s test (one-way ANOVA, p < 0.05). (F) Schematic representation of the brain. The region included in the red square shows the analyzed brain region.

Figure 3.
Hippocampal (A) brain-derived neurotrophic enzyme (Bdnf), (B) nuclear factor erythroid-derived 2-like 2 (Nrf2), (C) neuronal differentiation 1 (NeuroD), (D) superoxide dismutase type 1 (Sod1) and (E) glutathione peroxidase 1 (Gpx1) mRNA expression in Fisher 344 rats exposed to L6 (6 h light/18 h dark), L12 (12 h light/12 h dark), or L18 (18 h light/6 h dark) photoperiods for eleven weeks and administered vehicle (C), cherry 1 (Ch1, 100 mg/kg body weight) or cherry 2 (Ch2, 100 mg/kg body weight) during the last seven weeks. Values are expressed as mean ± standard deviation and normalized relative to the L12-C group (n = 8 per group). P, F and PxF indicate significant differences produced by photoperiod, fruit and the interaction between photoperiod and fruit, respectively (two-way ANOVA, p < 0.05). Different letters above the bars indicate significant differences by post-hoc Tukey’s test, (one-way ANOVA, p < 0.05).
Figure 3.
Hippocampal (A) brain-derived neurotrophic enzyme (Bdnf), (B) nuclear factor erythroid-derived 2-like 2 (Nrf2), (C) neuronal differentiation 1 (NeuroD), (D) superoxide dismutase type 1 (Sod1) and (E) glutathione peroxidase 1 (Gpx1) mRNA expression in Fisher 344 rats exposed to L6 (6 h light/18 h dark), L12 (12 h light/12 h dark), or L18 (18 h light/6 h dark) photoperiods for eleven weeks and administered vehicle (C), cherry 1 (Ch1, 100 mg/kg body weight) or cherry 2 (Ch2, 100 mg/kg body weight) during the last seven weeks. Values are expressed as mean ± standard deviation and normalized relative to the L12-C group (n = 8 per group). P, F and PxF indicate significant differences produced by photoperiod, fruit and the interaction between photoperiod and fruit, respectively (two-way ANOVA, p < 0.05). Different letters above the bars indicate significant differences by post-hoc Tukey’s test, (one-way ANOVA, p < 0.05).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Biometric parameters and blood hormone levels of Fisher 344 rats exposed to different photoperiods and administered vehicle or cherries for seven weeks.
Table 1.
Biometric parameters and blood hormone levels of Fisher 344 rats exposed to different photoperiods and administered vehicle or cherries for seven weeks.
| Parameters | L6 Photoperiod | L12 Photoperiod | L18 Photoperiod | 2wA | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| C | Ch1 | Ch2 | C | Ch1 | Ch2 | C | Ch1 | Ch2 | ||
| Final BW (g) | 367.7 ± 13.7 ab | 369.5 ± 20.0 b | 370.8 ± 21.3 ab | 389.8 ± 25.4 a | 393.7 ± 14.9 a | 381.8 ± 17.1 ab | 383.7 ± 27.8 a | 387.8 ± 20.7 ab | 376.1 ± 24.0 ab | P |
| EPI (kcal/h of dark) | 3.00 ± 0.04 a | 3.16 ± 0.19 a | 3.06 ± 0.25 a | 4.62 ± 0.27 b | 4.68 ± 0.27 b | 4.48 ± 0.26 b | 9.15 ± 0.22 c | 9.02 ± 0.22 c | 9.19 ± 0.47 c | P |
| Testosterone (ng/mL) | 2.98 ± 0.72 a | 4.16 ± 1.02 b | 3.45 ± 1.43 ab | 2.95 ± 0.98 a | 4.20 ± 1.09 b | 3.68 ± 1.83 ab | 1.63 ± 1.0 c | 1.86 ± 0.98 c | 2.43 ± 1.39 ac | P, PxF |
| Testes (g) | 3.08 ± 0.16 | 3.13 ± 0.14 | 3.15 ± 0.12 | 3.15 ± 0.13 | 3.20 ± 0.1 | 3.26 ± 0.14 | 3.05 ± 0.17 | 3.2 ± 0.15 | 3.2 ± 0.17 | |
| T4/T3 | 42.46 ± 2.64 a | 50.68 ± 4.89 b | 47.76 ± 4.85 ab | 48.31 ± 4.41 b | 47.45 ± 5.90 ab | 47.89 ± 5.40 b | 51.25 ± 6.14 b | 51.97 ± 6.47 b | 47.6 ± 5.22 ab | PxF |
Fisher 344 rats were exposed to L6 (6 h light/day), L12 (12 h light/day) or L18 (18 h light/day) photoperiods for eleven weeks and administered vehicle (C), cherry 1 (Ch1, 100 mg/kg body weight) or cherry 2 (Ch2, 100 mg/kg body weight) during the last 7 weeks of the experiment. Values are expressed as mean ± standard deviation (n = 8/group). P and PxF indicate significant differences in photoperiod and interactions between photoperiod and fruit, respectively (two-way ANOVA, 2wA, p < 0.05). Different letters indicate significant differences among groups for each parameter (one-way ANOVA, post-hoc Tukey’s test, p < 0.05; one-way ANOVA, post-hoc Tukey’s test, p < 0.05-p < 0.01). Abbreviations: body weight (BW), eating pattern index (EPI), triiodothyronine (T3), thyroxine (T4).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Manocchio, F.; Bravo, F.I.; Helfer, G.; Muguerza, B. Cherries with Different Geographical Origins Regulate Neuroprotection in a Photoperiod-Dependent Manner in F344 Rats. Antioxidants 2024, 13, 72. https://doi.org/10.3390/antiox13010072
AMA Style
Manocchio F, Bravo FI, Helfer G, Muguerza B. Cherries with Different Geographical Origins Regulate Neuroprotection in a Photoperiod-Dependent Manner in F344 Rats. Antioxidants. 2024; 13(1):72. https://doi.org/10.3390/antiox13010072
Chicago/Turabian StyleManocchio, Francesca, Francisca Isabel Bravo, Gisela Helfer, and Begoña Muguerza. 2024. "Cherries with Different Geographical Origins Regulate Neuroprotection in a Photoperiod-Dependent Manner in F344 Rats" Antioxidants 13, no. 1: 72. https://doi.org/10.3390/antiox13010072
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.