
H2O2 and Ca2+ Signaling Crosstalk Counteracts ABA to Induce Seed Germination
by
 , and
, and
Mengjie Cheng
1,†, 
Yanliang Guo
1,†,
Qing Liu
1,
Sanwa Nan
1,
Yuxing Xue
1,
Chunhua Wei
1,
Yong Zhang
1,
Feishi Luan
2,
Xian Zhang
1 and
Hao Li
1,* 1
College of Horticulture, Northwest A&F University, Yangling 712100, China
2
College of Horticulture and Landscape Architecture, Northeast Agricultural University, Harbin 150000, China
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Antioxidants 2022, 11(8), 1594; https://doi.org/10.3390/antiox11081594
Submission received: 24 June 2022
/
Revised: 13 August 2022
/
Accepted: 16 August 2022
/
Published: 17 August 2022
(This article belongs to the Special Issue Hydrogen Peroxide Signaling in Physiology and Pathology)
Abstract
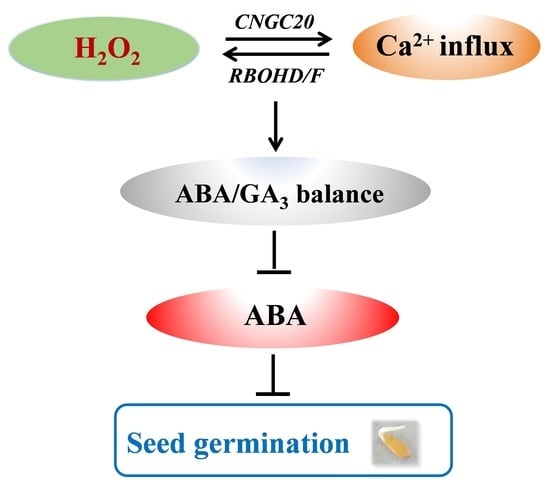
:Seed germination is a critical stage and the first step in the plant’s life cycle. H2O2 and Ca2+ act as important signal molecules in regulating plant growth and development and in providing defense against numerous stresses; however, their crosstalk in modulating seed germination remains largely unaddressed. In the current study, we report that H2O2 and Ca2+ counteracted abscisic acid (ABA) to induce seed germination in melon and Arabidopsis by modulating ABA and gibberellic acid (GA3) balance. H2O2 treatment induced a Ca2+ influx in melon seeds accompanied by the upregulation of cyclic nucleotide-gated ion channel (CNGC) 20, which encodes a plasma membrane Ca2+-permeable channel. However, the inhibition of cytoplasmic free Ca2+ elevation in the melon seeds and Arabidopsis mutant atcngc20 compromised H2O2-induced germination under ABA stress. CaCl2 induced H2O2 accumulation accompanied by the upregulation of respiratory burst oxidase homologue (RBOH) D and RBOHF in melon seeds with ABA pretreatment. However, inhibition of H2O2 accumulation in the melon seeds and Arabidopsis mutant atrbohd and atrbohf abolished CaCl2-induced germination under ABA stress. The current study reveals a novel mechanism in which H2O2 and Ca2+ signaling crosstalk offsets ABA to induce seed germination. H2O2 induces Ca2+ influx, which in turn increases H2O2 accumulation, thus forming a reciprocal positive-regulatory loop to maintain a balance between ABA and GA3 and promote seed germination under ABA stress.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
Seed germination is the first and vital step in a plant’s life cycle [1]. This physiological process starts with water absorption by dry seeds and ends with radicle emergence, during which the imbibed seeds shift from a quiescent state to active metabolism [2]. The transition from seed dormancy to germination involves simultaneous seed reserve mobilization and seed coat rupture, which cumulate in radicle emergence and subsequent seedling establishment [3,4]. Seed germination is a complex process that depends on multiple environmental factors, such as temperature, water availability, light, and oxygen, as well as certain intrinsic factors such as phytohormones [5]. Both poor seed quality and sowing conditions adversely affect seed germination and subsequent crop establishment, health, and yield [6].
Abscisic acid (ABA) and gibberellic acid (GA) are two central hormones that transduce environmental information and play antagonistic roles in regulating seed germination [7]. In fact, seed dormancy or germination largely depends on the dynamic balance of ABA and GA [8]. ABA promotes seed dormancy but inhibits seed germination. Knockout of ABA biosynthesis- or signaling-related genes promoted seed germination and reduced seed dormancy [9]. However, the mutation of ABA catabolic enzymes or overexpression of ABA biosynthetic enzymes delays seed germination but prolongs seed dormancy [2,10]. Conversely, GA breaks seed dormancy and promotes seed germination by activating the growth potential of embryos and weakening the embryo surrounding tissues [11].
In plants, reactive oxygen species (ROS), such as hydroxyl radicals (OH.), superoxide (O2−), and hydrogen peroxide (H2O2), play important roles in regulating seed dormancy or germination [12,13]. The controlled germination of ROS during seed imbibition can oxidize a subset of biomolecules such as mRNAs, nucleic acids, amino acids, and proteins, resulting in adequate cell functions that trigger seed germination [14]. In particular, H2O2 generated by respiratory burst oxidase homologues (RBOHs)-encoded NADPH oxidase is a critical secondary signaling molecule that regulates ABA and GA balance and affects their signaling pathways in regulating seed germination [15,16].
In addition to ROS, calcium (Ca2+) functions as a vital second messenger and plays an important role in regulating various physiological processes, including seed germination [17,18]. The Ca2+ signal functions by eliciting characteristic transient fluctuations of cytoplasmic free Ca2+ ([Ca2+]cyt) concentration through membrane transport proteins-mediated activities of Ca2+ influx and efflux [19]. Activated Ca2+ influx channels, such as two-pore channels (TPCs), glutamate receptor homolog (GLR) 3.5, and cyclic nucleotide-gated ion channels (CNGCs), induce a transient [Ca2+]cyt elevation and then trigger Ca2+ signal, which is decoded and relayed to downstream regulators of gene expression by a set of Ca2+ sensor proteins [19]. Abundant evidence suggests that the Ca2+ signal plays an important role in regulating seed germination by altering ABA and GA signaling [20,21].
Despite abundant studies on the individual role of H2O2 and Ca2+ in regulating seed germination, their potential interaction in modulating ABA/GA balance and subsequent seed germination remains largely unknown. The crosstalk of H2O2 (a major ROS) and Ca2+ regulating plant growth and development and its role as a defense against numerous stresses have been well documented [22]. For example, in plant response to cold exposure, Ca2+ signal is rapidly induced and then activates RBOH activity to trigger H2O2 production, which in turn induces Ca2+ transient influx in plant cells, forming a reciprocal positive-regulatory loop [23,24,25]. This raises the possibility that H2O2 and Ca2+ may also interact with each other in modulating seed germination. We demonstrate that the positive crosstalk of H2O2 and Ca2+ promotes seed germination under ABA stress. H2O2 promotes Ca2+ influx, which in turn increases H2O2 accumulation to regulate ABA and GA balance.
2. Materials and Methods
2.1. Plant Materials
In this study, melon (Cucumis melo L. cv. SSMA) seeds were provided by the Watermelon and Melon Research Group of Northwest A&F University, Yangling, China. Seeds of Arabidopsis mutant atrbohd (SALK_120299), atrbohf (SALK_034674), and atcngc20 (SALK_074919C) with a Columbia genetic background were obtained from the Arabidopsis Biological Resource Center (https://www.arabidopsis.org/, accessed on 1 June 2018).
2.2. Experimental Design
To investigate the role and mechanisms of H2O2 and Ca2+ in counteracting ABA to promote seed germination, the melon or Arabidopsis seeds were soaked in double distilled water (as control) or different test solutions for 7 h and then were rinsed three times with double-distilled water. The rinsed melon seeds were incubated at 30 °C in the dark for 7 days in 9 cm diameter Petri dishes containing double-layered rolled filter paper moistened with distilled water. The rinsed Arabidopsis seeds were incubated at 21 °C under continuous light at 100 μM m−2 s−1 for 7 days in 3 cm Petri dishes containing moistened filter papers. Each treatment comprised three replicates. Each replicate consisted of 30 seeds. Seeds were considered germinated when the radicle emerged (1–2 mm). Germination was scored daily as radicle emergence.
Test solutions used to treat melon seeds included ABA (1 mM), ABA (1 mM) + H2O2 (10 mM), ABA (1 mM) + CaCl2 (0.2, 0.5, 1, or 3 mM), ABA (1 mM) + diphenyleneiodonium (DPI, an inhibitor of NADPH oxidases, which produces ROS, 10 μM) [26,27,28], ABH (1 mM ABA + 10 mM H2O2) + ethylene glycol-bis (2-aminoethylether)-N,N,N′,N′-tetraacetic acid (EGTA, a Ca2+ chelator, 5 mM) [18,29], ABH + LaCl3 (a plasma membrane-located Ca2+ channel blocker, 5 mM) [18,29], ACa (1 mM ABA + 1 mM CaCl2) + DPI (10 μM). Test solutions used to treat Arabidopsis seeds included ABA (0.2 mM), ABA (0.2 mM) + H2O2 (5 mM), ABA (0.2 mM) + CaCl2 (0.2, 0.5, or 1 mM). The concentrations of the different chemicals are referred to Li et al. [30].
Notably, the chemical concentrations were higher in this study than those in previous studies. These high concentrations of chemicals were also used to assess their functions in various plant species [27,30]. The chemical concentrations were justified previously by using a range of concentrations, and the optimum concentration was used [30]. Generally, in previous studies, the chemicals such as ABA were added to the incubation medium during seed germination for several days. In the current study, the seeds were soaked with test solutions for 7 h and then rinsed three times with distilled water. Such treatments can increase endogenous chemical content and avoid exogenous chemicals’ constant and complex influence during seed incubation. After ABA (1 mM) pretreatment for 7 h, the endogenous ABA in melon seeds increased from 16.8 to 133.6 ng g−1 [30]. Therefore, a high concentration of ABA did not completely inhibit seed germination owing to the short treatment time (see Figure 1).
2.3. Net Ca2+ Flux Assay
The net Ca2+ flux of the seed embryo cells was measured with a Noninvasive Micro-test Technology (NMT) Physiolyzer (Xuyue Science and Technology Company Limited, Beijing, China) following the method of Li et al. [30]. The flux unit is pmol cm−2 s−1, and the negative and positive values represent Ca2+ influx and efflux, respectively.
2.4. Analysis of H2O2
The H2O2 content in seeds was measured according to the method described by Willekens et al. [31]. In brief, 0.2 g samples were ground in 3 mL of ice-cold HClO4 (1 M). After centrifugation at 6000× g for 5 min at 4 °C, the supernatant pH was adjusted to 6.0–7.0 with KOH (4 M). The supernatants were further centrifuged at 12,000× g for 5 min at 4 °C, passed through an AG1-X8 prepacked column (Bio-Rad, Hercules, CA, USA), and then were eluted with 4 mL ddH2O. The sample extract (800 μL) was combined with 400 μL of 100 mM potassium acetate (pH 4.4) containing horseradish peroxidase (0.25 U), 2,2′-azino-di (3-ethylbenzthiazoline-6-sulfonic acid) (4 mM), and 400 μL deionized water. The H2O2 content was measured at OD412.
2.5. Analysis of ABA and GA3
The plant hormones were extracted as described by Yang et al. [32] with minor modifications. In brief, 0.3 g of frozen seed samples was homogenized in 4 mL of methanol (80%, v/v) containing 1 mM 2,6-di-t-butyl-p-cresol as an antioxidant. The homogenates were incubated at 4 °C for 4 h and then were centrifuged at 1000× g for 20 min at 4 °C. The obtained supernatants (crude extracts) were filtered through C18 Sep-Park Cartridge (Millipore, Millford, MA, USA), dried under N2 (gas) flow, and then were dissolved in 5 mL of 50 mM Tris (pH 8.1) containing 10% (v/v) methanol, 1 mM MgCl2, and 150 mM NaCl. ABA and GA3 contents were analyzed with an immunoassay kit (China Agricultural University, Beijing, China) following the manufacturer’s instructions. Colorimetric readings were conducted using a Multimode Plate Reader M200 pro (Tecan, Männedorf, Switzerland).
2.6. qRT-PCR Analysis
The total RNA were isolated from the seeds using an RNA extraction kit (Axgen, Union City, CA, USA). After extraction, a DNase Mini Kit (Qiagen, Hilden, Germany) was used to remove residual DNA. Then, total RNA (1 µg per sample) was reverse-transcribed to cDNA using a FastKing RT kit (TIANGEN, Beijing, China). qRT-PCR was conducted on an iCycler Iq TM Multicolor PCR Detection System (Bio-Rad, Hercules, CA, USA) using SYBR® Premix ExTaqTM II (2×) kit (Takara, Tokyo, Japan). The gene-specific primers were designed according to the EST sequences (http://cucurbitgenomics.org/, accessed on 1 March 2018): 5′-TTGGTGCTGGCGAATTGGTTGA-3′ and 5′-ATGATCTGAGGCAGCGGCAAA-3′ for CmCNGC20 (MELO3C001941); 5′-AGTGAGTGACAGCCGAGTTCTAAGT-3′ and 5′-CTGCTCTGTGACGGTATTGGATGAA-3′ for CmRBOHD (MELO3C026754); 5′-GCACGAGTTGAAGGCTGAGTTGA-3′ and 5′-GGAATCCATCCTTGGCGAGCTTATC’ for CmRBOHF (MELO3C005718); and 5′-ATTCTTGCATCTCTAAGTACCTTCC-3′ and 5′-CCAACTAAAGGGAAATAACTCACC-3′ for CmActin (MELO3C008032). CmActin was used as the internal control genes [30]. The relative expression of mRNA was calculated as described previously [33].
2.7. Statistical Analysis
The experiment was a completely randomized design with three independent replications. Each replicate included 30 seeds. Analysis of variance (ANOVA) was used to test for significance, and significant differences among treatments were determined using Tukey’s test at the p < 0.05 level.
3. Results
3.1. H2O2 and Ca2+ Counteract ABA to Induce Seed Germination
Exogenous ABA pretreatment remarkably delayed germinating time and severely reduced germination rates of both melon and Arabidopsis seeds (Figure 1). However, exogenous H2O2 or CaCl2 pretreatment attenuated the ABA-induced inhibition of seed germination in melon and Arabidopsis seeds. The most effective concentration of CaCl2 was 1 mM and 0.5 mM for melon and Arabidopsis seeds, respectively. The promoting effect of CaCl2 on seed germination was attenuated with CaCl2 concentrations both lower and higher than the optimum concentration. For example, the germination rates of melon seeds pretreated with ABA (1 mM) + H2O2 (10 mM) and ABA (1 mM) + CaCl2 (1 mM) were 53.3% and 74.4%, respectively, far higher than that of seeds with ABA pretreatment.
Figure 1.
H2O2 and Ca2+ counteract abscisic acid (ABA) to promote seed germination in melon and Arabidopsis. (A,B) The effects of ABA and the combination of ABA and H2O2 (ABH) on the germination of melon and Arabidopsis seeds. (C,D) The effects of ABA and the combination of ABA and CaCl2 (ACa) on the germination of melon and Arabidopsis seeds. The melon seeds were presoaked in double-distilled water or test solutions for 7 h and then were incubated at 30 °C for 7 days. In (C,D), the germination rates were recorded on the seventh day after seed incubation. Data are means ± SD (3 replicates × 30 seeds). The different letters denote a significant difference (p < 0.05) according to Tukey’s test.
Figure 1.
H2O2 and Ca2+ counteract abscisic acid (ABA) to promote seed germination in melon and Arabidopsis. (A,B) The effects of ABA and the combination of ABA and H2O2 (ABH) on the germination of melon and Arabidopsis seeds. (C,D) The effects of ABA and the combination of ABA and CaCl2 (ACa) on the germination of melon and Arabidopsis seeds. The melon seeds were presoaked in double-distilled water or test solutions for 7 h and then were incubated at 30 °C for 7 days. In (C,D), the germination rates were recorded on the seventh day after seed incubation. Data are means ± SD (3 replicates × 30 seeds). The different letters denote a significant difference (p < 0.05) according to Tukey’s test.

3.2. Ca2+ Signal Is Involved in H2O2-Induced Seed Germination
To investigate whether the Ca2+ signal participates in H2O2-induced seed germination under ABA stress, we firstly analyzed the effects of H2O2 and its deficiency on Ca2+ flux. As shown in Figure 2, the ABA pretreatment had very little effect on the net flux of Ca2+ in both melon and Arabidopsis seeds. Compared to ABA, ABH (ABA+H2O2) pretreatment induced Ca2+ influx, accompanied by an increase in the transcript level of CmCNGC20, a gene encoding a plasma membrane Ca2+-permeable channel (Figure 2A,B). However, ABA+DPI (an inhibitor of NADPH oxidase-mediated ROS generation, 10 μM) decreased Ca2+ influx and downregulated the expression of CmCNGC20. Consistently, atrbohd and atrbohf mutation in Arabidopsis seeds inhibited Ca2+ influx under both control conditions and ABA stress (Figure 2C). These results suggest that RBOH-dependent ROS mediates Ca2+ influx during seed germination under ABA stress.
To further investigate the role of Ca2+ signals in H2O2-induced seed germination and ABA/GA3 balance under ABA stress, EGTA (a Ca2+ chelator) and LaCl3 (a plasma membrane-located Ca2+ channel blocker) were used to chelate Ca2+ and block Ca2+ influx, respectively. As shown in Figure 3A, both EGTA and LaCl3 inhibited H2O2-induced seed germination under ABA stress. The germination rates of seeds pretreated with ABA, ABA+H2O2 (ABH), ABH+EGTA, and ABH+LaCl3 were 36.7%, 90.0%, 58.9%, and 57.8%, respectively. EGTA but not LaCl3 promoted ABH-induced decrease in ABA level (Figure 3B). However, both EGTA and LaCl3 completely compromised the ABH-induced increase of GA level. As a result, LaCl3 but not EGTA abolished ABH-decreased ABA and GA3 ratio. The ABA/GA3 ratio in seeds with ABH+LaCl3 pretreatment was 29.0, significantly higher than that (14.7) in seeds with ABH pretreatment. When compared to wild-type Arabidopsis seeds, seeds of atcngc20 mutant showed lower germination rates under ABA stress (Figure 3D). However, AtCNGC20 deletion attenuated H2O2-induced seed germination under ABA stress.
3.3. H2O2 Mediates Ca2+-Induced Seed Germination
To investigate whether H2O2 was involved in Ca2+-induced seed germination under ABA stress, we analyzed the effects of CaCl2 on H2O2 accumulation under ABA stress. During incubation at 30 °C, the H2O2 level in melon seeds increased by ABA pretreatment from the first day, reached their peaks on the third day, and then declined gradually (Figure 4). CaCl2 promoted ABA-induced H2O2 accumulation. On the 1st and third day of incubation at 30 °C, H2O2 content in melon seeds pretreated with ABA+CaCl2 (ACa) were 115.2% and 25.0% higher, respectively, than that in the seeds pretreated with ABA alone. The expression of CmRBOHD was increased by ABA pretreatment, and this increase was promoted by CaCl2 on the first and seventh days during seed incubation. The expression of CmRBOHF was increased by ABA pretreatment from the third day of incubation. CaCl2 upregulated CmRBOHF expression in early response to ABA (within one day after seed incubation).
As well as H2O2, CaCl2 induced seed germination, ABA decrease, GA increase, and decrease of ABA/GA ratio under ABA stress (Figure 5). To further investigate the role of H2O2 in Ca2+-induced seed germination and ABA/GA3 balance under ABA stress, DPI, an inhibitor of NADPH oxidases, was used to inhibit H2O2 production. DPI attenuated or completely abolished Ca2+-induced seed germination, ABA decrease, GA increase, and decrease of ABA/GA ratio under ABA stress.
When compared to wild-type Arabidopsis seeds, seeds of the atrbohd and atrbohf mutants showed higher germination rates under ABA stress (Figure 6), suggesting that H2O2 is involved in ABA-induced inhibition of seed germination. However, the deletion of AtRBOHD and AtRBOHF completely abolished CaCl2-induced seed germination under ABA stress. Taken together, these results indicated that the Ca2+ and H2O2 signals interacted with each other, forming a reciprocal positive-regulatory loop, which antagonize ABA to promote seed germination.
4. Discussion
ABA and GA are two critical plant hormones and function antagonistically in regulating seed germination [7]. In the seeds exposed to salt and cold temperature, ABA accumulation was increased but GA3 accumulation was not decreased, indicating that ABA increase is the principal factor that disturbs the balance between ABA and GA3 and consequently inhibits seed germination unfavorable conditions [30].
As an important signal molecule, ROS generated by RBOHs, which encode NADPH oxidase, plays a dual role in regulating seed germination [30,34]. The H2O2 levels transiently increase during seed germination and inhibition of H2O2 production by NADPH inhibitor DPI or AtRBOHD deletion suppresses seed germination [35], suggesting that transient H2O2 generation is required for seed germination [14]. However, atrbohd and atrbohf double mutations impair ABA-induced promotion of ROS production and inhibition of seed germination, indicating that H2O2 was involved in ABA-induced seed dormancy [36].
In the present study, we found that exogenous H2O2 alleviated ABA-induced inhibition of seed germination, providing evidence on H2O2-ABA antagonism in regulating seed germination (Figure 1 and Figure 3). However, ABA pretreatment increased H2O2 accumulation and the transcript levels of RBOHD and RBOHF in melon seeds and Arabidopsis seeds with AtRBOHD or AtRBOHF deletion showed less sensitivity to ABA, indicating that H2O2 is required for ABA-induced seed dormancy (Figure 4 and Figure 6) [36]. In addition, the uncontrolled accumulation of H2O2 under stress conditions or during seed aging also inhibits seed germination [35]. Therefore, the occurrence of seed germination progresses is restricted to a critical range of H2O2 levels [34]. H2O2 at low or high levels would not permit progress toward germination. The previous study by Liu et al. showed that exogenous H2O2 promote ABA catabolism and GA biosynthesis during Arabidopsis seed imbibition [37]. Consistently, we found that exogenous H2O2 decreased the ABA level and increased the GA3 level during seed germination under ABA stress, suggesting that H2O2-induced seed germination was closely associated with its regulatory role in balancing ABA/GA3 (Figure 3).
Similar to H2O2, Ca2+ has a critical secondary signaling molecule and functions synergistically or antagonistically with ABA signaling during seed germination. For example, the repression of Arabidopsis GLR3.5 (AtGLR3.5), which encodes a plasma membrane Ca2+-permeable channel, impairs [Ca2+]cyt elevation and enhances seed sensitivity to ABA, whereas AtGLR3.5 overexpression promotes seed germination but reduce seed sensitivity to ABA by suppressing the expression of ABSCISIC ACID INSENSITIVE4, suggestive of a negative role of Ca2+ in ABA signaling [38]. However, the deletion of TPC1 (a vacuolar Ca2+ channel gene) and Ca2+ signaling-related genes (e.g., calcium-dependent protein kinase (CPK) 4, CPK11, and the calmodulin-like protein 39) results in insensitivity to ABA during seed germination, suggestive of a positive role of the Ca2+ signal in ABA signaling [39,40,41].
In the current study, we found that CaCl2 pretreatment decreased the ABA/GA3 ratio likely by promoting ABA catabolism and GA3 biosynthesis and thus promoted seed germination under ABA stress, suggesting that cytoplasmic Ca2+ signaling acts as a positive regulator in ABA-regulated seed germination. Therefore, the Ca2+ signal plays a dual role in seed germination response to ABA as well as H2O2. Extracellular Ca2+ enters the cytosol through plasma membrane Ca2+-permeable channels to positively regulate seed germination, whereas the Ca2+ release from the vacuole through vacuolar channels is required for ABA signaling that inhibits seed germination.
The crosstalk between the H2O2 derived from RBOHs and the Ca2+ signal is well documented in various physiological actions [22]. Evidently, the H2O2-triggered influx of Ca2+ has long been thought to be involved in H2O2 sensing and signalling [42]. Ca2+ acts as a key downstream component of AtRBOHD and AtRBOHF, transducing ROS signals in plant growth and in responses to various stresses [36,43,44,45]. For example, in ABA signalling in guard cells, H2O2 activates Ca2+-permeable channels and induces influx of Ca2+ and increases in [Ca2+]cyt in guard cells, which mediate stomatal closure induced by ABA [46,47]. However, the crosstalk between H2O2 and Ca2+ in regulating seed germination remains largely unknown. In the current study, H2O2 promoted Ca2+ influx in melon seeds under ABA stress, while the H2O2 deficiency in both melon and Arabidopsis seeds prevented Ca2+ influx under ABA stress (Figure 2). Similarly, the atrbohd and atrbohf mutations impair the hypoxia-elicited Ca2+ enhancement in Arabidopsis root cells [48]. CNGC20 is an important Ca2+ transport system that conducts external Ca2+ in the cytoplasm [49]. H2O2 induced the upregulation of CmCNGC20, suggesting that CmCNGC20 may be involved in H2O2-induced Ca2+ influx (Figure 2B). Moreover, blocking of Ca2+ influx by LaCl3 or chelation of Ca2+ by EGTA inhibited H2O2-induced ABA/GA3 balance and germination in melon seeds pretreated with ABA (Figure 3A–C). AtCNGC20 deletion in Arabidopsis seeds completely abolished H2O2-induced seed germination under ABA stress (Figure 3D). These results indicate that a Ca2+ signal is involved in H2O2-induced ABA/GA3 balance and subsequent seed germination under ABA stress.
Calcineurin-B-like (CBL) proteins and their interacting protein kinases (CIPKs) have been shown to function in many Ca2+-signaling processes, including seed germination. The mutations of CBL9 and CIPK3 exhibited hypersensitivity to ABA during seed germination, suggesting a negative role of Ca2+ signals in ABA-inhibited seed germination [50,51]. Interestingly, a calcium signal-activated CBL1/9-CIPK26 module can enhance ROS production via phosphorylation of RBOHF [52]. CPK5 phosphorylates RBOHD and produces ROS to induce systemic defense responses [53]. Here, CaCl2 increased H2O2 accumulation under ABA stress, accompanied by the upregulation of CmRBOHD and CmRBOHF (Figure 4).
Additional experiments showed that DPI prevented the CaCl2-induced ABA/GA3 balance and subsequent germination of melon seeds under ABA stress (Figure 5). Moreover, AtRBOHD and AtRBOHF deletion in Arabidopsis seeds completely abolished CaCl2-induced seed germination under ABA stress (Figure 6). Therefore, it is apparent that H2O2 participates in Ca2+-induced ABA/GA3 balance and subsequent seed germination under ABA stress. Taken together, the H2O2 and Ca2+ signals function together in a self-amplifying feedback loop, in which H2O2 induces Ca2+ influx, and Ca2+ subsequently increases H2O2 accumulation during seed response to ABA. Such an H2O2/Ca2+ activation circuit has also been reported to be required for rapid defense signal propagation in plants [53,54].
5. Conclusions
To date, the crosstalk underlying H2O2 and Ca2+ signals antagonize ABA to promote seed germination are unclear. In this study, we show that H2O2 and Ca2+ signals interact with each other to regulate ABA/GA3 balance and seed germination during ABA response. H2O2 promotes Ca2+ influx, which in turn increases H2O2 accumulation, forming a reciprocal positive-regulatory loop, to sustain Ca2+ influx-elicited signature and regulate ABA and GA3 balance. To our knowledge, this is the first study of its kind to provide evidence for the role of ROS and Ca2+ signaling in antagonizing ABA to regulate seed germination.
Author Contributions
Conceptualization, H.L.; data curation, M.C., Y.G., Q.L., S.N. and Y.X.; Formal analysis, Y.G. and H.L.; funding acquisition, X.Z. and H.L.; investigation, M.C., Y.G., Q.L., S.N., Y.X. and C.W.; project administration, H.L.; resources, Y.Z., F.L. and X.Z.; supervision, H.L.; writing—original draft, H.L.; writing—review and editing, H.L. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the China Agriculture Research System (CARS-25), the National Natural Science Foundation of China (31972479), the Young Science and Technology Star of Shaanxi (2022KJXX-80), and the Science and Technology Innovation Team of Shaanxi (2021TD-32).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All datasets generated for this study are included in the article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Donohue, K.; Rafael, R.D.C.; Burghardt, L.; Kovach, K.; Willis, C.G. Germination, post-germination adaptation, and species ecological ranges. Annu. Rev. Ecol. Evol. Syst. 2010, 41, 293–319. [Google Scholar] [CrossRef]
- Holdsworth, M.J.; Bentsink, L.; Soppe, W.J.J. Molecular networks regulating Arabidopsis seed maturation, after ripening, dormancy and germination. New Phytol. 2008, 179, 33–54. [Google Scholar] [CrossRef] [PubMed]
- Finkelstein, R.; Reeves, W.; Ariizumi, T.; Steber, C. Molecular aspects of seed dormancy. Annu. Rev. Plant Biol. 2008, 59, 387–415. [Google Scholar] [CrossRef]
- Rajjou, L.; Duval, M.; Gallardo, K.; Catusse, J.; Bally, J.; Job, C.; Job, D. Seed germination and vigor. Annu. Rev. Plant Biol. 2012, 63, 507–533. [Google Scholar] [CrossRef] [PubMed]
- Barba-Espin, G.; Diaz-Vivancos, P.; Clemente-Moreno, M.J.; Albacete, A.; Faize, L.; Faize, M.; Pérez-Alfocea, F.; Hernández, J.A. Interaction between hydrogen peroxide and plant hormones during germination and the early growth of pea seedlings. Plant Cell Environ. 2010, 33, 981–994. [Google Scholar] [CrossRef]
- Lamichhane, J.R.; Debaeke, P.; Steinberg, C.; You, M.P.; Barbetti, M.J.; Aubertot, J.N. Abiotic and biotic factors affecting crop seed germination and seedling emergence: A conceptual framework. Plant Soil 2018, 432, 1–28. [Google Scholar] [CrossRef]
- Footitt, S.; Douterelo-Soler, I.; Clay, H.; Finch-Savage, W.E. Dormancy cycling in Arabidopsis seeds is controlled by seasonally distinct hormone-signaling pathways. Proc. Natl. Acad. Sci. USA 2011, 108, 20236–20241. [Google Scholar] [CrossRef]
- Weitbrecht, K.; Müller, K.; Leubner-Metzger, G. First off the mark: Early seed germination. J. Exp. Bot. 2011, 62, 3289–3309. [Google Scholar] [CrossRef] [PubMed]
- Finkelstein, R.R.; Gampala, S.S.L.; Rock, C.D. Abscisic acid signaling in seeds and seedlings. Plant Cell 2002, 14, S15–S45. [Google Scholar] [CrossRef]
- Okamoto, M.; Kuwahara, A.; Seo, M.; Kushiro, T.; Asami, T.; Hirai, N.; Kamiya, H.Y.; Koshiba, T.; Nambara, E. CYP707A1 and CYP707A2, which encode abscisic acid 8′-hydroxylases, are indispensable for proper control of seed dormancy and germination in Arabidopsis. Plant Physiol. 2006, 141, 97–107. [Google Scholar] [CrossRef]
- Ogawa, M.; Hanada, A.; Yamauchi, Y.; Kuwahara, A.; Kamiya, Y.; Yamaguchi, S. Gibberellin biosynthesis and response during Arabidopsis seed germination. Plant Cell 2003, 15, 1591–1604. [Google Scholar] [CrossRef] [PubMed]
- Bailly, C. Active oxygen species and antioxidants in seed biology. Seed Sci. Res. 2004, 14, 93–107. [Google Scholar] [CrossRef]
- Gomes, M.P.; Garcia, Q.S. Reactive oxygen species and seed germination. Biologia 2013, 68, 351–357. [Google Scholar] [CrossRef]
- El-Maarouf-Bouteau, H.; Meimoun, P.; Job, C.; Bailly, C. Role of protein and mRNA oxidation in seed dormancy and germination. Front. Plant Sci. 2013, 4, 77. [Google Scholar] [CrossRef] [PubMed]
- Bahin, E.; Bailly, C.; Sotta, B.; Kranne, I.; Corbineau, F.; Leymarie, J. Crosstalk between reactive oxygen species and hormonal signalling pathways regulates grain dormancy in barley. Plant Cell Environ. 2011, 34, 980–993. [Google Scholar] [CrossRef] [PubMed]
- Bailly, C. The signalling role of ROS in the regulation of seed germination and dormancy. Biochem. J. 2019, 476, 3019–3032. [Google Scholar] [CrossRef]
- Berridge, M.J.; Lipp, P.; Bootman, M.D. The versatility and universality of calcium signalling. Nat. Rev. Mol. Cell Biol. 2000, 1, 11–21. [Google Scholar] [CrossRef]
- Zhang, Z.; Wu, P.; Zhang, W.; Yang, Z.; Liu, H.; Ahammed, G.J.; Cui, J.X. Calcium is involved in exogenous NO-induced enhancement of photosynthesis in cucumber (Cucumis sativus L.) seedlings under low temperature. Sci. Hortic. 2020, 261, 108953. [Google Scholar] [CrossRef]
- Dodd, A.N.; Kudla, J.; Sanders, D. The language of calcium signaling. Annu. Rev. Plant Biol. 2010, 61, 593–620. [Google Scholar] [CrossRef]
- Salahshoor, F.; Kazemi, F. Effect of calcium on reducing salt stress in seed germination and early growth stage of Festuca ovina L. Plant Soil Environ. 2016, 62, 460–466. [Google Scholar] [CrossRef]
- Singh, K.L.; Chaudhuri, A.; Kar, R.K. Role of peroxidase activity and Ca2+ in axis growth during seed germination. Planta 2015, 242, 997–1007. [Google Scholar] [CrossRef]
- Niu, L.J.; Liao, W.B. Hydrogen peroxide signaling in plant development and abiotic responses: Crosstalk with nitric oxide and calcium. Front. Plant Sci. 2016, 7, 230. [Google Scholar] [CrossRef] [PubMed]
- Kawarazaki, T.; Kimura, S.; Iizuka, A.; Hanamata, S.; Nibori, H.; Michikawa, M.; Imai, A.; Abe, M.; Kaya, H.; Kuchitsu, K. A low temperature-inducible protein AtSRC2 enhances the ROS-producing activity of NADPH oxidase AtRbohF. BBA-Mol. Cell Res. 2013, 1833, 2775–2780. [Google Scholar] [CrossRef] [PubMed]
- Baxter, A.; Mittler, R.; Suzuki, N. ROS as key players in plant stress signalling. J. Exp. Bot. 2013, 65, 1229–1240. [Google Scholar] [CrossRef]
- Yuan, P.; Yang, T.; Poovaiah, B.W. Calcium signaling-mediated plant response to cold stress. Int. J. Mol. Sci. 2018, 19, 3896. [Google Scholar] [CrossRef] [PubMed]
- Miller, G.; Schlauch, K.; Tam, R.; Cortes, D.; Torres, M.A.; Shulaev, V.; Dangl, J.L.; Mittler, R. The plant NADPH oxidase RBOHD mediates rapid systemic signaling in response to diverse stimuli. Sci. Signal. 2009, 2, 45. [Google Scholar] [CrossRef]
- Xia, X.J.; Wang, Y.J.; Zhou, Y.H.; Tao, Y.; Mao, W.H.; Shi, K.; Asami, T.; Chen, Z.X.; Yu, J.Q. Reactive oxygen species are involved in brassinosteroid-induced stress tolerance in cucumber. Plant Physiol. 2009, 150, 801–814. [Google Scholar] [CrossRef]
- Xia, X.J.; Zhou, Y.H.; Ding, J.; Shi, K.; Asami, T.; Chen, Z.X.; Yu, J.Q. Induction of systemic stress tolerance by brassinosteroid in Cucumis sativus. New Phytol. 2011, 191, 706–720. [Google Scholar] [CrossRef]
- Khangembam, L.S.; Anindita, M.; Ru, K.K. Early axis growth during seed germination is gravitropic and mediated by ROS and calcium. J. Plant Physiol. 2017, 216, 181–187. [Google Scholar] [CrossRef]
- Li, H.; Guo, Y.L.; Lan, Z.X.; Zhang, Z.X.; Ahammed, G.J.; Chang, J.J.; Zhang, Y.; Wei, C.H.; Zhang, X. Melatonin antagonizes ABA action to promote seed germination by regulating Ca2+ efflux and H2O2 accumulation. Plant Sci. 2021, 303, 110761. [Google Scholar] [CrossRef]
- Willekens, H.; Chamnongpol, S.; Davey, M.; Schraudner, M.; Langebartels, C.; Van, M.M.; Inzé, D.; Van, C.W. Catalase is a sink for H2O2 and is indispensable for stress defence in C3 plants. EMBO J. 1997, 16, 4806–4816. [Google Scholar] [CrossRef]
- Yang, J.; Zhang, J.; Wang, Z.; Zhu, Q.; Wang, W. Hormonal changes in the grains of rice subjected to water stress during grain filling. Plant Physiol. 2001, 127, 315–323. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Bailly, C.; El-Maarouf-Bouteau, H.; Corbineau, F. From intracellular signaling networks to cell death: The dual role of reactive oxygen species in seed physiology. CR Biol. 2008, 331, 806–814. [Google Scholar] [CrossRef] [PubMed]
- Leymarie, J.; Vitkauskaité, G.; Hoang, H.H.; Gendreau, E.; Chazoule, V.; Meimoun, P.; El-Maarouf-Bouteau, H.; Bailly, C. Role of reactive oxygen species in the regulation of Arabidopsis seed dormancy. Plant Cell Physiol. 2012, 53, 96–106. [Google Scholar] [CrossRef]
- Kwak, J.M.; Mori, I.C.; Pei, Z.M.; Leonhardt, N.; Torres, M.A.; Dangl, J.L.; Bloom, R.E.; Bodde, S.; Jones, J.D.; Schroeder, J.I. NADPH oxidase AtrbohD and AtrbohF genes function in ROS-dependent ABA signaling in Arabidopsis. EMBO J. 2003, 22, 2623–2633. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Ye, N.; Liu, R.; Chen, M.; Zhang, J. H2O2 mediates the regulation of ABA catabolism and GA biosynthesis in Arabidopsis seed dormancy and germination. J. Exp. Bot. 2010, 61, 2979–2990. [Google Scholar] [CrossRef]
- Kon, D.D.; Ju, C.L.; Parihar, A.; Kim, S.; Cho, D.; Kwak, J.M. Arabidopsis glutamate receptor homolog3.5 modulates cytosolic Ca2+ level to counteract effect of abscisic acid in seed germination. Plant Physiol. 2015, 167, 1630–1642. [Google Scholar] [CrossRef]
- Peiter, E.; Maathuis, F.J.M.; Mills, L.N.; Knight, H.; Pelloux, J.; Hetherington, A.; Sanders, D. The vacuolar Ca2+-activated channel TPC1 regulates germination and stomatal movement. Nature 2005, 434, 404–408. [Google Scholar] [CrossRef]
- Zhu, S.Y.; Yu, X.C.; Wang, X.J.; Zhao, R.; Zhang, D.P. Two Calcium-Dependent Protein Kinases, CPK4 and CPK11, Regulate abscisic acid signal transduction in Arabidopsis. Plant Cell 2007, 19, 3019–3036. [Google Scholar] [CrossRef] [PubMed]
- Ubaid, M.; Ting, M.K.Y.; Teresinski, H.J.; Snedden, W.A. The calmodulin-like protein, CML39, is involved in regulating seed development, germination, and fruit development in Arabidopsis. Plant Mol. Biol. 2018, 96, 375–392. [Google Scholar] [CrossRef]
- Wu, F.; Chi, Y.; Jiang, Z.; Xu, Y.Y.; Xie, L.; Huang, F.F.; Wan, D.; Ni, J.; Yuan, F.; Wu, X.M.; et al. Hydrogen peroxide sensor HPCA1 is an LRR receptor kinase in Arabidopsis. Nature 2020, 578, 577–581. [Google Scholar] [CrossRef] [PubMed]
- Mori, I.C.; Schroeder, J.I. Reactive oxygen species activation of plant Ca2+ channels. A signaling mechanism in polar growth, hormone transduction, stress signaling, and hypothetically mechanotransduction. Plant Physiol. 2004, 135, 702–708. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.Y.; Zhang, H.; Sun, L.R.; Jiao, Y.H.; Zhang, G.Z.; Miao, C.; Hao, F.S. NADPH oxidase AtrbohD and AtrbohF function in ROS dependent regulation of Na+/K+ homeostasis in Arabidopsis under salt stress. J. Exp. Bot. 2012, 63, 305–317. [Google Scholar] [CrossRef]
- Jiao, Y.H.; Sun, L.R.; Song, Y.L.; Wang, L.M.; Liu, L.P.; Zhang, L.Y.; Liu, B.; Li, N.; Miao, C.; Hao, F.S. AtrbohD and AtrbohF positively regulate abscisic acid inhibited primary root growth by affecting Ca2+ signaling and auxin response of roots in Arabidopsis. J. Exp. Bot. 2013, 64, 4183–4192. [Google Scholar] [CrossRef] [PubMed]
- Pei, Z.M.; Murata, Y.; Benning, G.; Thomine, S.; Klüsener, B.; Allen, G.J.; Grill, E.; Schroeder, J.I. Calcium channels activated by hydrogen peroxide mediate abscisic acid signalling in guard cells. Nature 2000, 406, 731–734. [Google Scholar] [CrossRef]
- Brandt, B.; Munemasa, S.; Wang, C.; Nguyen, D.; Yong, T.M.; Yang, P.G.; Poretsky, E.; Belknap, T.F.; Waadt, R.; Alemán, F.; et al. Correction: Calcium specificity signaling mechanisms in abscisic acid signal transduction in Arabidopsis guard cells. eLife 2015, 4, e03599. [Google Scholar] [CrossRef]
- Liu, B.; Sun, L.; Ma, L.; Hao, F.S. Both AtrbohD and AtrbohF are essential for mediating responses to oxygen deficiency in Arabidopsis. Plant Cell Rep. 2017, 36, 947–957. [Google Scholar] [CrossRef]
- Spalding, E.P.; Harper, J.F. The ins and outs of cellular Ca2+ transport. Curr. Opin. Plant Bio. 2011, 14, 715–720. [Google Scholar] [CrossRef]
- Pandey, G.K.; Cheong, Y.H.; Kyung-Nam, K.; Grant, J.J.; Li, L.G.; Wendy, H.; Cecilia, D.; Stefan, W.; Jörg, K.; Sheng, L. The calcium sensor calcineurin B-like 9 modulates abscisic acid sensitivity and biosynthesis in Arabidopsis. Plant Cell 2004, 16, 1912–1924. [Google Scholar] [CrossRef]
- Pandey, G.K.; Grant, J.J.; Cheong, Y.H.; Beom-Gi, K.; Li, L.G.; Sheng, L. Calcineurin-B-like protein CBL9 interacts with target kinase CIPK3 in the regulation of ABA response in seed germination. Mol. Plant 2008, 1, 238–248. [Google Scholar] [CrossRef] [PubMed]
- Drerup, M.M.; Schlücking, K.; Hashimoto, K.; Manishankar, P.; Steinhorst, L.; Kuchitsu, K.; Kudla, J. The Calcineurin B-like calcium sensors CBL1 and CBL9 together with their interacting protein kinase CIPK26 regulate the Arabidopsis NADPH oxidase RBOHF. Mol. Plant 2013, 6, 559–569. [Google Scholar] [CrossRef] [PubMed]
- Dubiella, U.; Seybold, H.; Durian, G.; Komander, E.; Lassig, R.; Witte, C.P.; Schulze, W.X.; Romeis, T. Calcium-dependent protein kinase/NADPH oxidase activation circuit is required for rapid defense signal propagation. Proc. Natl. Acad. Sci. USA 2013, 110, 8744–8749. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.K. Abiotic stress signaling and responses in plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef]
Figure 2.
The effects of H2O2 on Ca2+ flux in seeds under ABA stress. (A,B) Ca2+ flux and CmCNGC20 expression in melon seeds pre-treated with ABA, a combination of ABA and H2O2 (ABH), and a combination of ABA and diphenyleneiodonium (DPI, an inhibitor of NADPH oxidases, which produces H2O2). (C) Ca2+ flux in seeds of wild-type (WT) Arabidopsis and atrbohd and atrbohf mutants under normal conditions and ABA stress. In (A,C), dry melon seeds or Arabidopsis were soaked in double distilled water or test solutions for 7 h and then were used to analyze the changes in Ca2+ flux. In (B), melon seeds were treated as that in Figure 1. Data are means ± SD (3 replicates × 30 seeds). The different letters denote a significant difference (p < 0.05) according to Tukey’s test.
Figure 2.
The effects of H2O2 on Ca2+ flux in seeds under ABA stress. (A,B) Ca2+ flux and CmCNGC20 expression in melon seeds pre-treated with ABA, a combination of ABA and H2O2 (ABH), and a combination of ABA and diphenyleneiodonium (DPI, an inhibitor of NADPH oxidases, which produces H2O2). (C) Ca2+ flux in seeds of wild-type (WT) Arabidopsis and atrbohd and atrbohf mutants under normal conditions and ABA stress. In (A,C), dry melon seeds or Arabidopsis were soaked in double distilled water or test solutions for 7 h and then were used to analyze the changes in Ca2+ flux. In (B), melon seeds were treated as that in Figure 1. Data are means ± SD (3 replicates × 30 seeds). The different letters denote a significant difference (p < 0.05) according to Tukey’s test.

Figure 3.
The role of Ca2+ in H2O2 counteracting ABA to promote seed germination. (A) The germination rates of melon seeds treated with ABA, a combination of ABA and H2O2 (ABH), a combination of ABH and EGTA (a Ca2+ chelator), and a combination of ABH and LaCl3 (a plasma membrane-located Ca2+ channel blocker). (B,C) ABA and GA3 content and ABA/GA3 ratio in melon seeds treated with ABA, ABH, ABH+EGTA, and ABH+LaCl3. ABA and GA3 contents were measured on the third day after seed incubation. (D) The effects of ABA and ABH on the germination rates of seeds in wild-type (WT) Arabidopsis and atcngc20 mutant. In both (A,D), germination rates were recorded on the seventh day after seed incubation. Data are means ± SD (3 replicates × 30 seeds). The different letters denote significant difference (p < 0.05) according to Tukey’s test.
Figure 3.
The role of Ca2+ in H2O2 counteracting ABA to promote seed germination. (A) The germination rates of melon seeds treated with ABA, a combination of ABA and H2O2 (ABH), a combination of ABH and EGTA (a Ca2+ chelator), and a combination of ABH and LaCl3 (a plasma membrane-located Ca2+ channel blocker). (B,C) ABA and GA3 content and ABA/GA3 ratio in melon seeds treated with ABA, ABH, ABH+EGTA, and ABH+LaCl3. ABA and GA3 contents were measured on the third day after seed incubation. (D) The effects of ABA and ABH on the germination rates of seeds in wild-type (WT) Arabidopsis and atcngc20 mutant. In both (A,D), germination rates were recorded on the seventh day after seed incubation. Data are means ± SD (3 replicates × 30 seeds). The different letters denote significant difference (p < 0.05) according to Tukey’s test.

Figure 4.
The effects of ABA and combination of ABA and CaCl2 (ACa) on the H2O2 accumulation and the relative expression of CmRBOHD and CmRBOHF during melon seed incubation. Data are means ± SD (3 replicates × 30 seeds).
Figure 4.
The effects of ABA and combination of ABA and CaCl2 (ACa) on the H2O2 accumulation and the relative expression of CmRBOHD and CmRBOHF during melon seed incubation. Data are means ± SD (3 replicates × 30 seeds).

Figure 5.
Involvement of H2O2 in Ca2+ counteracting ABA to promote seed germination. (A) The germination rate of melon seeds treated with ABA, a combination of ABA and CaCl2 (ACa), and a combination of ACa and diphenyleneiodonium (DPI, an inhibitor of NADPH oxidases, which produces ROS). Germination rates were recorded on the seventh day after seed incubation. (B,C) ABA and GA3 content and ABA/GA3 ratio in melon seeds treated with ABA, a combination of ABA and CaCl2 (ACa), and a combination of ACa and DPI. ABA and GA3 contents were measured on the third day after seed incubation. Data are means ± SD (3 replicates × 30 seeds). The different letters denote a significant difference (p < 0.05) according to Tukey’s test.
Figure 5.
Involvement of H2O2 in Ca2+ counteracting ABA to promote seed germination. (A) The germination rate of melon seeds treated with ABA, a combination of ABA and CaCl2 (ACa), and a combination of ACa and diphenyleneiodonium (DPI, an inhibitor of NADPH oxidases, which produces ROS). Germination rates were recorded on the seventh day after seed incubation. (B,C) ABA and GA3 content and ABA/GA3 ratio in melon seeds treated with ABA, a combination of ABA and CaCl2 (ACa), and a combination of ACa and DPI. ABA and GA3 contents were measured on the third day after seed incubation. Data are means ± SD (3 replicates × 30 seeds). The different letters denote a significant difference (p < 0.05) according to Tukey’s test.

Figure 6.
The effects of ABA and combination of ABA and CaCl2 (ACa) on the germination rates of seeds in wild-type (WT) Arabidopsis and atrbohd and atrbohf mutants. Germination rates were recorded on the seventh day after seed incubation. Data are means ± SD (3 replicates × 30 seeds). The different letters denote a significant difference (p < 0.05) according to Tukey’s test.
Figure 6.
The effects of ABA and combination of ABA and CaCl2 (ACa) on the germination rates of seeds in wild-type (WT) Arabidopsis and atrbohd and atrbohf mutants. Germination rates were recorded on the seventh day after seed incubation. Data are means ± SD (3 replicates × 30 seeds). The different letters denote a significant difference (p < 0.05) according to Tukey’s test.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Cheng, M.; Guo, Y.; Liu, Q.; Nan, S.; Xue, Y.; Wei, C.; Zhang, Y.; Luan, F.; Zhang, X.; Li, H. H2O2 and Ca2+ Signaling Crosstalk Counteracts ABA to Induce Seed Germination. Antioxidants 2022, 11, 1594. https://doi.org/10.3390/antiox11081594
AMA Style
Cheng M, Guo Y, Liu Q, Nan S, Xue Y, Wei C, Zhang Y, Luan F, Zhang X, Li H. H2O2 and Ca2+ Signaling Crosstalk Counteracts ABA to Induce Seed Germination. Antioxidants. 2022; 11(8):1594. https://doi.org/10.3390/antiox11081594
Chicago/Turabian StyleCheng, Mengjie, Yanliang Guo, Qing Liu, Sanwa Nan, Yuxing Xue, Chunhua Wei, Yong Zhang, Feishi Luan, Xian Zhang, and Hao Li. 2022. "H2O2 and Ca2+ Signaling Crosstalk Counteracts ABA to Induce Seed Germination" Antioxidants 11, no. 8: 1594. https://doi.org/10.3390/antiox11081594
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.