
Novel Probiotic Lactic Acid Bacteria Were Identified from Healthy Infant Feces and Exhibited Anti-Inflammatory Capacities


1
State Key Laboratory of Food Science and Technology, Jiangnan University, Wuxi 214122, China
2
School of Food Science and Technology, Jiangnan University, Wuxi 214122, China
3
School of Medicine, Jiangnan University, Wuxi 214122, China
*
Authors to whom correspondence should be addressed.
Antioxidants 2022, 11(7), 1246; https://doi.org/10.3390/antiox11071246
Submission received: 11 May 2022
/
Revised: 16 June 2022
/
Accepted: 17 June 2022
/
Published: 24 June 2022
(This article belongs to the Special Issue The Role of Redox Modulation by Natural Agents under Steady State or Inflammatory Conditions)
Abstract
:The current study aims to evaluate the probiotic potential of lactic acid bacteria isolated from infant feces, and select candidates to be used as potential antioxidants for the treatment of oxidative stress-related diseases; To meet the criteria for probiotic attributes, the isolates were subjected to various in vitro tests and 16S rRNA genotypic characterization. Besides, anti-inflammatory and anti-oxidative effects of selected isolates were separately assessed by real-time quantitative PCR and Western blot; The selected strains belonged to Lactobacillus gasseri, Lactiplantibacillus plantarum and Lacticaseibacillus rhamnosus. Notably, three selected strains (L. gasseri FWJL-4, L. plantarum Fjias-5 and L. rhamnosus FSJ-13) particularly L. gasseri FWJL-4 significantly down-regulated mRNA expression levels of tumor necrosis factor α (TNFα), Interleukin-6 (IL-6) and IL-1β. Most importantly, three strains-treated RAW 264.7 murine macrophages displayed enhanced activities of antioxidant enzymes and reduced H2O2 production, which were associated with the enhanced expression levels of nuclear factor-erythroid 2 related factor 2 and heme oxygenase-1; Three selected strains, particularly L. gasseri FWJL-4, are good candidates that merit additional in vivo investigation for the validation and application of their health-promoting effects.
1. Introduction
Probiotics are live microorganisms that when consumed in adequate amounts, can exert beneficial effects on the host [1]. Well-known probiotic microorganisms include genera of lactic acid bacteria (LAB) (Lactococcus, Lactobacillus, Streptococcus, Pediococcus, Leuconostoc and Enterococcus), Bifidobacterium, Bacillus, as well as yeasts [1,2,3,4]. Among them, the genera Lactobacillus, Limosilactobacillus, Lacticaseibacillus, Lactiplantibacillus, etc. are the earliest found and most studied probiotics, which are Gram-positive and catalase-negative probiotics and have been shown to provide health benefits to the human host. As reported, LAB are natural inhabitants of the human gastrointestinal tract and play a critical role in restoring the human gut microbiota homeostasis [5]. In addition, LAB exhibit positive effects on various aspects of human health including nutrition, metabolism, immunity, and defense against pathogens [6]. Lacticaseibacillus casei, Lacticaseibacillus paracasei, Lactobacillus acidophilus, Limosilactobacillus fermentum, Lactiplantibacillus plantarum, Limosilactobacillus reuteri, Lacticaseibacillus rhamnosus, Lactobacillus gasseri and Lactobacillus crispatus are common probiotics existing in animal and human gastrointestinal and digestive systems [7]. These strains have been isolated from different environmental sources such as fermented foods [8] and human samples [9].
As reported, LAB can produce a high amount of lactic acid and other metabolites such as bacteriocins [10], short-chain fatty acids [11], soluble mediators [12], and exopolysaccharides [13]. Consequently, they possess the ability to withstand harsh conditions in the human body (such as salivary enzymes, low pH, and intestinal juice), colonize gut epithelial cells, and inhibit pathogens and reactive oxygen species (ROS) related to gut diseases, thus helping to maintain gut microbiota balance and immune homeostasis and exert physiological roles in human health [14,15].
Oxidative stress is an excessive production of ROS, which can provoke the onset and progression of various diseases, including diabetes [16], cancer [17] and cardiovascular diseases [18], etc. Accumulating studies have suggested that intestinal flora metabolites may participate in the modulation of inflammation-related diseases through regulating oxidative stress [19,20,21]. Despite the evident potential, knowledge is scarce about the antioxidant effects of probiotics, as an alternative for various disease protection against oxidative damage. The detailed mechanisms of probiotics in mediating inflammation and oxidative stress-related diseases remain to be fully explored. Currently, despite great efforts in the identification of probiotic strains, their functional substances (either metabolites or components) haven’t been specified.
The current study aims to isolate and identify novel bacterial strains belonging to the LAB from healthy infants, which can be used as potential probiotics against inflammation and oxidative stress-related human diseases. The candidate probiotic strains or their metabolites can be potentially used as novel antioxidants.
2. Materials and Methods
2.1. Sample Collection and Strain Isolation
A total of 100 stool samples of healthy infants (63 boys and 37 girls; without gastrointestinal tract diseases, allergic diseases and congenital diseases) aged under 3 weeks, were collected for the study. The present study was approved by the ethics committee of the Affiliated Wuxi Children’s Hospital of Nanjing Medical University [WXCH2018-08-007]. Written informed consent was obtained from the parents after a careful explanation of the research. After collection, samples were transferred to de Man-Rogosa-Sharpe (MRS) broth (purchased from Hope Bio-Technology Co., Qingdao, China) and cultured at 37 °C for 48–72 h. The MRS broth was composed of 2 g/L of dipotassium hydrogen phosphate, 20 g/L glucose, 0.2 g/L magnesium sulfate heptahydrate, 0.05 g/L manganous sulfate tetrahydrate, 8 g/L beef extract, 10 g/L peptone, 5 g/L sodium acetate trihydrate, 2 g/L triammonium citrate and 4 g/L yeast extract. After that, the samples were diluted with sterile saline and spread onto MRS agar medium, and incubated for 48–72 h at 37 °C under anaerobic conditions in an anaerobic workstation (Electrotek, West Yorkshire, UK). LAB colonies were purified twice using MRS agar medium. Then, the obtained bacterial isolates were used for morphological and biochemical characterization. The reference strain L. rhamnosus GG (LGG, ATCC 53103), gut-related pathogens, including Staphylococcus aureus ATCC 25923, Salmonella enterica ATCC 14028, Escherichia coli ATCC 25922, Listeria monocytogenes ATCC 13932 were obtained from the American Type Culture Collection (ATCC). Enterococcus faecalis E27 was obtained from the Culture Collection Center of Jiangnan University (Wuxi, China). Selected strains were kept in-house culture collection of the School of Food Science and Technology, Jiangnan University at −80 °C in 30% glycerol (v/v).
2.2. Morphological and Biochemical Characterization
The Gram staining test was used to characterize bacterial morphological properties. Biochemical properties were evaluated using the catalase test and analysis of carbohydrate fermentation. The biochemical identification tube system was purchased from Hope Bio-Technology Co., to analyze carbohydrate fermentation profiles of bacterial isolates. Fifteen kinds of carbohydrates were selected including glucose, galactose, maltose, mannose, lactose, sucrose, inositol, sorbitol, L-xylose, fructose, L-arabinose, rhamnose, cellobiose, esculin and mannitol. Each tube contained a specific carbohydrate. The signal bacterial colony was incubated in the relevant tube and cultured at 37 °C for 48–72 h under anaerobic conditions. The positive results were recorded according to the manufacturer’s instructions. The growth abilities of strains in the presence of 3% and 4.5% NaCl, and at 15 °C and 45 °C were also assessed [22,23]. Finally, catalase-negative and Gram-positive strains were selected as potential probiotic strains. The selected bacterial isolates were stored in 30% glycerol (v/v) at −80 °C for further use.
2.3. Preliminary Selection of LAB Using Specific Primers
The Gram-positive and catalase-negative strains were selected and used for further analysis. Bacterial genomic DNA of all strains was extracted using a TIANamp Bacteria DNA Kit (Tiangen Biotech Co., Beijing, China) according to the manufacturer’s instructions. Specific primers of LAB were used (forward: 5′-GCYGGTGCWAACCCNGTTGG -3′; reverse: 5′-AANGTNCCVCGVATCTTGTT-3′) [24]. The PCR mix contained 25 µL of 12.5 µL of 2 × Taq Plus Master Mix (CWBIO Biotech Co., Beijing, China), 1 µL of each primer (10 μmol/L), 1 µL of DNA templates, and 9.5 µL of ddH2O. The amplification program was performed over 30 cycles of 94 °C for 30 s, 58 °C for 30 s and 72 °C for 2 min, with a final extension step of 10 min at 72 °C. A 1% agarose gel electrophoresis was performed to determine the sizes and amounts of the amplicons. The positive strains were then selected for further analysis.
2.4. Antimicrobial Activity Assessment
To detect the inhibitory effects of selected LAB on several major gut-related pathogens, including Staphylococcus aureus ATCC 25923 (LB medium), Salmonella enterica ATCC 14028 (BHI medium), Escherichia coli ATCC 25922 (LB medium), Listeria monocytogenes ATCC 13932 (BHI medium), Enterococcus faecalis E27 (MRS medium), the Oxford cup method was used. In detail, the bacterial isolates cultured in MRS broth at 37 °C for 24–48 h were centrifuged at 12,000 rpm for 10 min. Then the supernatants were collected. The pathogens were incubated at 37 °C for 16–24 h (approximately 1 × 106 CFUs/mL) and spread onto a separated MRS medium. The sterilized Oxford cup was carefully placed and pressed onto the medium, and 200 μL of bacterial supernatants were distributed into the cups. After incubation at 37 °C for 24 h, the antimicrobial activity of the strains was recorded as a growth-free inhibition zone around the Oxford cups. The diameter of the inhibition zones was scored as follows: less and equal to 7 mm (negative, −); 7–15 mm (weak), >15 mm (strong). The antibiotic penicillin (30 mg/mL) was used as a positive control. A corresponding liquid medium was used as the negative control. Each test was conducted in triplicate.
2.5. Tolerance to Simulated Digestive Tract Conditions
The simulated digestive juice was prepared according to previous report with minor modifications [25]. Artificial saliva (pH 6.9) consisted of 3 g/L α-amylase (Sigma-Aldrich Co., St. Louis, MO, USA) in a sterile solution, which contained 6 g/L NaCl, 0.2 g/L CaCl2, 2 g/L KCl and 1 g/L NaHCO3. Simulated gastric fluid (pH 2) was prepared by dissolving 3.0 g/L pepsin from porcine gastric mucosa (Sigma-Aldrich) in a sterile solution, which contained 1.1 g/L KCl, 3 g/L NaCl, 0.6 g/L NaHCO3 and 0.15 g/L CaCl2. Artificial small intestinal juice (pH 7.4) was prepared by dissolving 3 g/L bile salt (Sigma-Aldrich), 0.1 g/L lipase (Sigma-Aldrich) and 1 g/L pancreatin (Sigma-Aldrich) in sterile solution, which consisted of 5 g/L NaCl, 0.6 g/L NaHCO3, 0.3 g/L CaCl2 and 0.6 g/L KCl. All the solution was filtered through a 0.22-µm filter before use. Selected bacterial cells (1 mL, 1 × 108–109 CFUs/mL) were firstly suspended in 1 mL artificial saliva for 5 min. The cells were then centrifuged (4 °C, 12,000 rpm, 2 min) and resuspended in 2 mL gastric fluid and incubated at 37 °C for 2 h. Subsequently, bacterial cells were recentrifuged (4 °C, 12,000 rpm, 2 min) and resuspended in 2 mL simulated small intestinal fluid and incubated at 37 °C for 2 h. Finally, the bacterial suspensions were diluted and plated on MRS agar and cultured at 37 °C for 36–48 h under anaerobic conditions. The total number of colonies was counted and the survival rates were recorded according to the formula as follows:
N1 represents the total count of strains after simulated digestive juice treatment. N0 represents the total count of strains before simulated digestive juice treatment. Each test was conducted in triplicate.
2.6. Adherence to Intestinal Epithelial Cells
The adhesion ability of the bacterial isolates to human intestinal epithelial cells was measured according to the procedures described by Balthazar et al. [26]. The human colon adenocarcinoma (Caco-2) cell line (obtained from China Cell Bank) was used as the target cell to examine cell adhesion of selected strains. Caco-2 cells were cultured in Dulbecco’s Modified Eagle’s Medium (DMEM) (Hyclone, Logan, UT, USA) supplemented with 1% penicillin-streptomycin (Ameresco, United States) and 10% fetal bovine serum (FBS) (Wisent, ST-BRUNO, QC, Canada) at 37 °C, 5% CO2. Firstly, Caco-2 cells were seeded in 24-well culture plates (2 × 105 cells/well) and incubated until the confluency. Before adhesion, the medium in each well was washed with sterile PBS and replaced with a prewarmed fresh medium without antibiotics. Overnight cultured fresh bacterial cells (108 CFUs/mL) were washed twice with sterile PBS and added to each well. Following co-incubation for 3 h at 37 °C, cells were washed twice. Then, the cells were treated with trypsin/EDTA (Sigma-Aldrich) at 37 °C for 3 min. The suspension from each well was transferred to serial saline for 10-fold dilution and plated onto MRS agar plates. After 36–48 h of incubation at 37 °C, the total number of colonies was counted under anaerobic conditions. The adhesion rate was calculated by measuring the total bacterial counts before and after bacteria adhered to the Caco-2 cells.
C1 represents the total bacterial counts adhered to Caco-2 cells. C0 represents the total bacterial counts of strains before treatment. Each test was conducted in triplicate.
2.7. Antibiotic Resistance Assay
Eight antibiotics including four cell membrane/wall inhibitors (penicillin, ampicillin, polymyxin and vancomycin) and four protein synthesis inhibitors (streptomycin, erythromycin, chloramphenicol and kanamycin), are common antibiotics used for clinical infection [27]. The concerns regarding specific strains are long-term use may develop antibacterial resistance [28]. All antibiotics were purchased from Sangon Biotech Co., Shanghai, China. Each antibiotic was dissolved with a proper solution and filtered before use.
Bacterial strains were incubated in MRS broth supplemented with different final concentrations (2, 4, 8, 16, 32, 64, 128, 256, 512 and 1024 μg/mL) of antibiotics for 24 h incubation at 37 °C, and assayed in triplicate in a microplate reader (OD at 610 nm) [29]. MICs (minimum inhibitory concentrations) were considered the lowest concentrations of antibiotics that could inhibit strain growth, which was used to evaluate the antibiotic resistance of selected strains.
2.8. Strain Identification Using 16S rRNA Sequence Analyses
Primers used for amplifying the complete sequence of 16S rRNA were 27F (5′-AGAGTTTGATCCTGGCCTCA-3′) and 1492R (5′-GGTTACCTTGTTACGACTT-3′) [30]. The PCR mix contains 25 µL of 12.5 µL of 2 × Taq Plus Master Mix, 1 µL of each primer (10 μ mol/L), 1 µL of DNA templates, and 9.5 µL of ddH2O. The amplification program was performed over 30 cycles of 94 °C for 30 s, 55 °C for 30 s and 72 °C for 2 min, with a final extension step of 10 min at 72 °C. A 2% agarose gel electrophoresis was performed to determine the sizes and amounts of the amplicons. The nucleotide sequences were used for sequence identity analysis (http://www.ncbi.nlm.nih.gov/blast, 16 S rRNA database, accessed on 27 May 2022).
2.9. Cytokine Measurement
Murine macrophages (RAW 264.7, obtained from China Cell Bank) were cultured in DMEM medium at 37 °C in a 5% CO2 humidified incubator [9]. A density of 1 × 105 cells/mL was seeded in each well of a 6-well culture plate for 24 h. The culture supernatants of three different strains (OD600 of 1.0 units) (L. gasseri FWJL-4, L. plantarum Fjias-5 and L. rhamnosus FSJ-13) and positive control (LGG, OD600 of 1.0 units) were administrated to each well for 1 h, and then lipopolysaccharides (LPS, 1 μg/mL) were added for 20 h. The cell supernatants were obtained by centrifugation and filtration using a 0.22-μm membrane. According to the manufacturer’s protocol, Trizol reagent (Invitrogen Corp., Carlsbad, CA, USA) was used to exact total RNA from RAW 264.7 cells. The mRNA expression levels of interleukin-10 (IL-10), IL-6, IL-1β and tumor necrosis factor (TNFα) were determined using the CFX Connect Real-Time System (Bio-Rad, Hercules, CA, USA). The relative mRNA expression levels were normalized using the mRNA levels of β-actin. Primers used for real-time quantitative PCR (RT-qPCR) were indicated in Table 1.
2.10. Determination of Antioxidant Enzymes and H2O2 Production
RAW 264.7 cells (1 × 105 cells/mL) were seeded in each well of a 24-well culture plate for 24 h. The culture supernatants of three different strains (OD600 of 1.0 units) (L. gasseri FWJL-4, L. plantarum Fjias-5 and L. rhamnosus FSJ-13) and positive control (LGG, OD600 of 1.0 units) were added to each well for 1 h, and then 1 μg/mL of LPS were added for 20 h. After that, the cells were collected for the determination of superoxide dismutase (SOD), glutathione peroxidase (GPx), glutathione (GSH) and oxidized glutathione (GSSG) using assay kits (Beyotime Biotechnology, Shanghai, China) according to the manufacturer’s instructions. For H2O2 measurement, the cells were firstly treated with culture supernatants of three different strains for 1 h, and then treated with 1 μg/mL of LPS for 2 h, 4 h, 6 h, 8 h, 10 h and 12 h, respectively. After LPS treatment, cells were washed and incubated with Amplex Red (50 μM, Beyotime) in the dark at 37 °C for 30 min [31]. Subsequently, cells were washed and the fluorescence density was measured at OD571 nm by a Varioskan LUX Multimode Reader (Thermo Fisher Scientific, Waltham, MA, USA).
2.11. Western Blotting
RAW 264.7 cells (1 × 105 cells/mL) were seeded in each well of a 6-well culture plate for 24 h. The culture supernatants of three different strains (OD600 of 1.0 units) (L. gasseri FWJL-4, L. plantarum Fjias-5 and L. rhamnosus FSJ-13) were added to each well for 1 h, and then 1 μg/mL of LPS were added for 6 h. A Nuclear and Cytoplasmic Protein Extraction Kit (Beyotime) was used to extract the cell proteins. Equal amounts of protein were separated by electrophoresis (Mini-PROTEAN® Tetra cell system, Bio-Rad) in 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis and then transferred to a nitrocellulose membrane (Millipore, Bedford, MA, USA). The membranes were firstly blocked with 5% nonfat milk and then incubated with respective primary antibodies containing 5% BSA overnight at 4 °C. The membranes were then incubated with horseradish peroxidase-conjugated anti-rabbit secondary antibodies (1:5000, Thermo Fisher Scientific) for 2 h. The blots were visualized by ChemiDoc Imager (Bio-Rad) after enhancing chemiluminescence reaction. Densitometric analyses of bands were quantified using Image Lab software (Version 3.0, Bio-Rad) with β-actin or histone H3 as the internal control. The following primary antibodies were used: polyclonal rabbit anti-Nrf-2 (Cat#16396-1-AP, 1:1000, Proteintech Group, Rosemont, IL, USA), polyclonal rabbit anti-HO-1 (Cat#10701-1-AP, 1:1000, Proteintech Group), monoclonal rabbit anti-histone 3 (Cat#9717, 1:1000, Cell Signaling Biotechnology, Danvers, MA, USA) and monoclonal rabbit anti-β-actin (Cat#AC026, 1:5000, Abclonal Biotechnology, Wuhan, China).
2.12. Statistical Analysis
Results were reported as mean ± SD of three triplicates. Differences among three or more groups were determined using one-way analysis of variance (ANOVA) followed by the Tukey post hoc test. Statistical significance was defined as * p < 0.05; ** p < 0.01; *** p < 0.001. Graphical presentations were generated by Graphpad Prism version 8.3.
2.13. Data Availability
Nucleotide sequence data reported are available in the Genbank database under the accession numbers: MZ220367 (FRY-2), MZ220368 (FXHB-6), MZ220369 (Fjias-5), MZ220370 (FRY-4X), MZ220371 (FWJL-4), MZ220372 (FZL-2), MZ220373 (FRY-6), MZ220374 (FSJ-13), MZ220375 (FZSJ-7), MZ220376 (FXHB-2), MZ220377 (FWJL-5), MZ220378 (FSJ-4X).
3. Results
3.1. Morphological and Biochemical Test Results
250 microbial strains were isolated from healthy infant feces. Among these strains, 27 isolates were selected based on morphological and biochemical tests. All strains were rod, Gram-positive and catalase-negative. They were able to grow in 3% and 4.5% NaCl (w/v), as well as at 45 °C. However, only eight strains could grow at the temperature of 15 °C. Carbohydrate fermentation profiles of each strain were recorded. The results showed that all strains were able to ferment glucose, galactose, maltose, mannose, lactose, sucrose, inositol, inositol, L-xylose, fructose, L-arabinose, rhamnose, cellobiose and esculin, and not able to ferment sorbitol and mannitol, preliminarily confirming that they belong to LAB.
3.2. Specific Primers Amplification for LAB
3.3. Antimicrobial Test Results
Twenty-seven bacterial isolates were used to evaluate antimicrobial activities against five pathogens using their cell-free culture supernatants. As shown in Table 2, seven isolates (FMM-1, FLY-17, FMM-7, FHHY-7, FSJ-6, FLT6-11 and FQM-3X) showed either poor or no activity against five pathogens, while other bacterial isolates exhibited better antimicrobial activities. Compared to penicillin, Fjias-5, FWJL-4, FSJ-11, FSJ-13, FXHB-2, FSJ-4X, FXHB-6, FXHB-2X and FHHY-1 showed strong antimicrobial activities against more than three pathogens. Notably, 23 out of 27 isolates had a strong inhibitory effect on Enterococcus faecalis E27.
3.4. Tolerance to Simulated Digestive Tract Condition
The bacterial isolates with significant antimicrobial activities (20 isolates) were selected to assess their survival rates in simulated digestive juice. As shown in Figure 2A,B, the artificial saliva and gastric fluid treatment (Figure 2A) had a less significant effect on the survival of bacteria than that of the simulated digestive juices (Figure 2B). However, the survival rates of most strains in the artificial saliva and gastric fluid were almost the same as those in the simulated digestive juice, suggesting that the pH of digestive juice played a crucial role. Compared with the reference strain LGG, several tested strains showed a higher level of tolerance (p < 0.05). In particular, five strains, namely FRY-3, Fjias-5, FWJL-4, FRY-6 and FXHB-2, exhibited >90% of survival rates in simulated digestive juice (Figure 2B). In contrast, most strains (FZL-21, FHHY-1, FSJ-2, FZYY-3, FSJ-13, FXHB-2X, FRY-4X) showed a lower level of tolerance than the reference strain LGG (p < 0.05) (Figure 2B).
3.5. Adherence to Caco-2 Cells
Based on the tolerance test results, three strains (FHHY-1, FSJ-2 and FZYY-3) with lower survival rates were excluded and the remaining strains were further tested for their adhesion ability to Caco-2 cells in vitro. As shown in Figure 3, ten bacterial isolates (FWJL-4, FZL-2, FRY-2, FRY-6, Fjias-5, FXHB-6, FRY-4X, FZSJ-7, FXHB-2 and FSJ-13) showed a higher adhesion ability than the reference strain LGG (p < 0.05). Among them, FRY-2, FRY-4X and Fjias-5 exhibited excellent adhesion rates (>80%). In contrast, other isolates, including FWJL-5, FRY-3, FSJ-1, FSJ-11, FSJ-4X, FZL-21 and FXHB-2X, showed a significantly lower adhesion ability (mostly < 30%) than LGG (p < 0.05).
3.6. Antibiotic Resistance
The MICs of tested strains in the presence of different concentrations of antibiotics were shown in Table 3. Most selected strains exhibited resistance to three antibiotics (kanamycin, vancomycin and polymyxin), especially to polymyxin, but were sensitive to other antibiotics. Nine strains including FRY-2, FXHB-6, FRY-3, FZL-21, Fjias-5, FRY-4X, FWJL-4, FZL-2 and FRY-6 showed antibiotic sensitivity to cell wall inhibitors such as penicillin G, ampicillin, and vancomycin. Furthermore, almost all strains were sensitive to protein synthesis inhibitors such as streptomycin, erythromycin and chloramphenicol.
3.7. 16S rRNA Sequencing and Phylogenetic Tree Results
According to the results obtained above, 11 probiotic candidates (FRY-2, FXHB-6, Fjias-5, FRY-4X, FWJL-4, FZL-2, FRY-6, FSJ-13, FZSJ-7, FXHB-2 and FSJ-4X) were further characterized using the Sanger sequencing method. The sequencing data analysis and the resulting phylogenetic tree demonstrated that these isolates belonged to L. gasseri, L. plantarum and L. rhamnosus strains (Table 4, Figure S1). All isolates’ names, sequence similarity and accession numbers were shown in Table 4. As shown in Table 4, three species of probiotic LAB including L. gasseri, L. plantarum and L. rhamnosus were finally selected as potential probiotics in this study. In addition, the phylogenetic tree was constructed by MEGA7 software (version 7.0) using the bootstrap method (1000 bootstraps have been performed) (Figure S1).
3.8. LAB Treatments Inhibit Inflammation in LPS-Treated RAW 264.7 Murine Macrophages
Three strains from different species (L. gasseri FWJL-4, L. plantarum Fjias-5 and L. rhamnosus FSJ-13) were selected to evaluate their health-promoting effects. Not surprisingly, LPS treatment significantly enhanced the mRNA expression levels of pro-inflammatory cytokines including TNFα, IL-6 and IL-1β (Figure 4A–C), whereas decreasing IL-10 (Figure 4D), a common anti-inflammatory cytokine. However, cells treated with four strains of probiotic LAB exhibited reduced expression levels of TNFα, IL-6 and IL-1β, and increased expression of IL-10, with a pronounced effect observed for L. gasseri FWJL-4 (Figure 4A–D), implying a potential immunomodulatory property of L. gasseri FWJL-4.
3.9. LAB Treatments Reduce ROS Production in LPS-Treated RAW 264.7 Murine Macrophages
Since inflammatory signaling or inflammasome maybe positively regulated by ROS [32,33]. Next, we test whether selected strains can reduce oxidative stress-related features, such as the cellular activity of several antioxidant enzymes and H2O2 generation. Compare to cells without any treatment, LPS supplementation reduced the activities of antioxidant enzymes while largely increasing the production of H2O2 (Figure 5A–C). Intriguingly, most LAB treatments significantly enhanced the enzymatic activities of SOD and GPx and the ratio of GSH/GSSG relative to LPS-treated cells (Figure 5A–C). Besides, the generation of cellular H2O2 was also inhibited by LAB (Figure 5D). In addition, LPS stimulation for 2 h or 4 h significantly increased intracellular H2O2 generation, which was markedly attenuated by LAB pretreatment (Figure 5D). Besides, only L. gasseri FWJL-4 exhibited the highest activity of SOD, GPx and GSH/GSSG ratio relative to reference strain LGG (Figure 5A–C), suggesting the metabolites of L. gasseri FWJL-4 might be good antioxidants for reducing oxidative stress.
3.10. Selected Probiotic LAB Induce Nuclear Factor-Erythroid 2 Related Factor 2 (Nrf2)-Mediated Signaling Pathways
Since we find the extraordinary anti-inflammatory capacity of selected LAB, we next examine the protein levels of Nrf2, and its downstream antioxidant enzyme heme oxygenase-1 (HO-1). The two proteins are multifunctional modulators of inflammation and oxidative stress-related disorders [34]. In the present study, the expression levels of Nrf2 and HO-1 were increased in LPS-treated RAW 264.7 cells (Figure 6). Consistently, probiotic LAB treatment markedly enhanced the expression levels of Nrf2 and HO-1, with a pronounced effect observed for L. gasseri FWJL-4. However, the cells treated with the culture supernatants of L. plantarum Fjias-5 exhibited a poorer antioxidant capacity relative to the other strains. Together, the results demonstrate a potential role of metabolites of L. gasseri FWJL-4 in reducing cellular oxidative stress.
4. Discussion
Over the years, due to multiple health benefits and market demands, probiotics have gained considerable attention. Accumulating studies have been conducted to isolate and characterize potential probiotics from various origins [9,35]. LAB are the most common probiotics found in human GI-tract. This indigenous microbiota plays a major role in maintaining the microbial ecosystem of gut and modulating gut immune homeostasis [36].
In this study, we isolated 250 microbial strains from healthy infant feces. The biochemical and morphological characteristics of these strains were then identified using the Gram-staining, the catalase test and analysis of carbohydrate fermentation profiles. 27 Gram-positive and catalase-negative strains were selected. Besides, they could ferment various carbohydrates except for mannitol and sorbitol. As reported, lactose intolerance could cause undesirable gastrointestinal symptoms [37], and the ability of lactose utilization could alleviate lactose intolerance in specific individuals. Furthermore, all strains exhibited abilities to utilize other carbohydrates except mannitol and sorbitol. Thus, they may be related to metabolizing human milk oligosaccharides in infants [38].
The safety of bacterial strains is a crucial element in selecting potential probiotics. Firstly, we found that the culture suspensions of selected strains could inhibit the growth of five common pathogens, including both Gram-positive pathogens including Staphylococcus aureus, Listeria monocytogenes and Enterococcus faecalis, and Gram-negative pathogens (Salmonella enterica and Escherichia coli). These findings indicated that the selected strains with broad-spectrum antimicrobial activities might effective in hospital-acquired infections. Additionally, our strains showed better inhibitory effects than those reported in earlier studies [39,40]. The broad antimicrobial effects of LAB species are attributable to the production of metabolites such as organic acid, antimicrobial peptides, etc. Notably, all selected strains exhibited strong inhibitory effects against E. faecalis E27. As reported, due to the exceptional multidrug resistance, most isolates of E. faecalis have caused clinical infections that are often hard to treat [41]. However, the antibiotic resistance of E. faecalis selected in our study hasn’t been assayed. The inhibitory mechanisms of selected LAB species against pathogens may be associated with inhibition of biofilm formation and exopolysaccharide synthesis [42].
Probiotics must be able to survive under harsh conditions including low pH, bile salts and digestive enzymes. In this study, we used artificial saliva and gastric fluid and simulated the digestive tract to evaluate the survival rates of selected strains. The major factor that influences the growth of strains is the acidic circumstance (pH 2). After 2 h of incubation, strains were considered to pass through the stomach. Isolates including FRY-3, Fjias-5, FWJL-4, FRY-6 and FXHB-2 showed a higher survival ability than the reference strain LGG under unfavorable conditions. The results obtained in this study agreed with those from previous studies [43,44,45]. Furthermore, the resistance of probiotics to low pH and bile salts varies greatly among species and strains, suggesting that this survival ability may be strain-specific [23,38]. Resistance to low pH is important for the development of fermented foods such as yogurt and cheese. These foods are often acidic and can affect strain viability. It is reported that the resistance to high bile salts is associated with physiological changes in probiotics such as carbohydrate fermentation and exopolysaccharide production [46]. The composition of membrane proteins and fatty acids and the inhibition of pathogen adhesion to human mucus are also related to the adaption of probiotics to bile salts [47,48]. Therefore, resistance to bile salts is an important trait for strains to compete with pathogens when used in functional foods.
The ability to colonize the GIT epithelial cells is another important feature for potential probiotic candidates. Here, the strains showing stronger adhesion rates than the reference strain LGG were selected for further tests. Microbial adhesion to epithelial cells is a complex process, which is closely correlated with the physicochemical composition of the probiotic strain cell [1]. The adhesion ability of a probiotic is a crucial trait, as it helps the probiotic compete with pathogenic bacteria to prevent their colonization of the gastrointestinal tract.
The presence of antibiotic resistance genes in probiotics is considered a safety issue as the resistance genes can transfer among the microorganism community of the gut. Our selected strains were all resistant to kanamycin and polymyxin. Consistent with the literature, the resistant ability to kanamycin has been confirmed for most LAB species [23,49]. Besides, strains show resistance to polymyxin, which is a Gram-negative bacteria inhibitor and has no effect on Gram-positive bacteria. Notably, nearly all isolates were resistant to vancomycin, which was in accordance with previous studies [29,50]. However, it is reported that when the antibiotic resistance is intrinsic (chromosomally encoded, non-transferable and non-inducible), the probiotic strains do not constitute a safety concern itself. Thus, the resistant probiotic strains can be used concomitantly or after antibiotic treatment to restore the gut microbiota [51]. In fact, due to the lack of cytochrome-mediated electron transport in Lactobacillus genera, and the presence of D-Ala-D-lactate in their peptidoglycan, resistance to streptomycin, kanamycin, and vancomycin is considered to be intrinsic [52].
Several antioxidant enzymes including SOD, GPx, GSH, GSSG and HO-1 are secreted to protect cells against oxidative damage. In accordance with other studies, these enzymes were collectively enhanced and ROS production was reduced by our selected LAB [53,54,55]. Nrf2 has long been considered a cytoprotective factor regulating anti-inflammatory and anti-oxidative proteins. In Nrf2-dependent cell antioxidant responses, HO-1 is one of the main effectors, exerting beneficial effects through the protection against cellular oxidative injury, modulation of inflammation and regulation of apoptosis [56,57]. Macrophage polarization is a hallmark of inflammation [58]. As reported, macrophages can be differentiated into two subsets: pro-inflammatory M1 macrophages and anti-inflammatory M2 macrophages. Pro-inflammatory cytokine (IL-1β, TNFα and IL-6) production further causes aggravated inflammation. Besides, excessive ROS production is involved in various chronic inflammation-mediated diseases. Several inflammatory signaling and inflammasome are positively regulated by ROS derived from uncharacterized organelles [59,60]. In oxidative stress and inflammation condition, enhancement of Nrf2/HO-1 expression exerts a crucial role in cell protection [56]. Various oxidative-inducing agents, including LPS, can induce the expression levels of Nrf2 and HO-1 [57]. Our data also showed that LPS increased the levels of Nrf2 and HO-1, however, the selected LAB, especially L. gasseri FWJL-4, could further enhance the levels of Nrf2 and HO-1. The high expression levels of Nrf2 and HO-1 can inhibit LPS-activated ROS production, thus decreasing the levels of pro-inflammatory cytokines in RAW 264.7 murine macrophages. These results are consistent with previous reports that probiotics participate in the modulation of inflammation-related diseases by regulating oxidative stress [20,21]. Compare to previous studies, the probiotics screened in the present study exhibited both strong anti-inflammatory and antioxidative properties [61,62,63,64], which were largely attributed to their secondary metabolites. Therefore, herein, we provide a possibility that the metabolites of probiotics maybe functional substances in alleviating inflammation and oxidative stress, which needs to be further explored, however.
5. Conclusions
Our study suggests that three selected strains (L. gasseri FWJL-4, L. plantarum Fjias-5 and L. rhamnosus FSJ-13) activate the Nrf2/HO-1 signaling pathway to reduce oxidative stress, thus inhibiting inflammation in macrophages (Figure 7). These strains maybe good candidates in the applications of human trials and may provide a promising therapeutic approach to preventing oxidative stress and inflammation-associated disorders. Our study also provides a theoretical basis for the exploration of probiotic functional metabolites.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/antiox11071246/s1, Figure S1: The phylogenetic tree shows the relationships between different LAB strains.
Author Contributions
Conceptualization, J.S. and L.-L.P.; methodology, B.L.; software, B.L.; validation, L.-L.P. and J.S.; formal analysis, B.L.; investigation, L.-L.P.; writing—original draft preparation, B.L.; writing—review and editing, L.-L.P.; visualization, J.S.; supervision, J.S.; project administration, J.S. and L.-L.P.; funding acquisition, J.S. and L.-L.P. All authors have read and agreed to the published version of the manuscript.
Funding
The work was funded by the National Natural Science Foundation of China, grant number 81870439, Natural Science Foundation for Distinguished Young Scholars of Jiangsu Province, grant number BK20200026, the Fundamental Research Funds for the Central Universities, grant number JUSRP22007, Jiangsu Province “Six Summit Talents” Program, grant number YY-038, Collaborative Innovation Center of Food Safety and Quality Control in Jiangsu Province, and Wuxi Taihu Talent Program.
Institutional Review Board Statement
The present study was approved by the ethics committee of the Affiliated Wuxi Children’s Hospital of Nanjing Medical University [WXCH2018-08-007].
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
The data is contained within this article.
Acknowledgments
We thank Le Zhang and Lin Ding for their assistance in sample collection.
Conflicts of Interest
The authors declare no conflict of interest.
References
- de Melo Pereira, G.V.; de Oliveira Coelho, B.; Magalhães Júnior, A.I.; Thomaz-Soccol, V.; Soccol, C.R. How to select a probiotic? A review and update of methods and criteria. Biotechnol. Adv. 2018, 36, 2060–2076. [Google Scholar] [CrossRef]
- Doron, S.; Snydman, D.R. Risk and safety of probiotics. Clin. Infect. Dis. 2015, 60 (Suppl. S2), S129–S134. [Google Scholar] [CrossRef] [Green Version]
- Prado, F.C.; Lindner, J.D.D.; Inaba, J.; Thomaz-Soccol, V.; Brar, S.K.; Soccol, C.R. Development and evaluation of a fermented coconut water beverage with potential health benefits. J. Funct. Foods 2015, 12, 489–497. [Google Scholar] [CrossRef]
- Maria Rosa Machado, C.R.S. Current Developments in Probiotics. J. Microb. Biochem. Technol. 2015, 7, 11–020. [Google Scholar] [CrossRef]
- KKumar, R.; Sood, U.; Gupta, V.; Singh, M.; Scaria, J.; Lal, R. Recent Advancements in the Development of Modern Probiotics for Restoring Human Gut Microbiome Dysbiosis. Indian J. Microbiol. 2019, 60, 12–25. [Google Scholar] [CrossRef]
- Kerry, R.G.; Patra, J.K.; Gouda, S.; Park, Y.; Shin, H.-S.; Das, G. Benefaction of probiotics for human health: A review. J. Food Drug Anal. 2018, 26, 927–939. [Google Scholar] [CrossRef] [Green Version]
- Dixit, Y.; Wagle, A.; Vakil, B. Patents in the Field of Probiotics, Prebiotics, Synbiotics: A Review. J. Food Microbiol. Saf. Hyg. 2016, 1, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.; Li, N.; Wang, Q.; Liu, Z.; Lee, Y.-K.; Liu, X.; Zhao, J.; Zhang, H.; Chen, W. Microbial diversity and volatile profile of traditional fermented yak milk. J. Dairy Sci. 2020, 103, 87–97. [Google Scholar] [CrossRef]
- Zhai, Q.; Shen, X.; Cen, S.; Zhang, C.; Tian, F.; Zhao, J.; Zhang, H.; Xue, Y.; Chen, W. Screening of Lactobacillus salivarius strains from the feces of Chinese populations and the evaluation of their effects against intestinal inflammation in mice. Food Funct. 2020, 11, 221–235. [Google Scholar] [CrossRef]
- Ooi, M.F.; Mazlan, N.; Foo, H.L.; Loh, T.C.; Rosfarizan, M.; Rahim, R.A.; Ariff, A. Effects of carbon and nitrogen sources on bacteriocin-inhibitory activity of postbiotic metabolites produced by Lactobacillus plantarum I-UL4. Malays. J. Microbiol. 2015, 176–184. [Google Scholar]
- Wang, G.; Yang, S.; Sun, S.; Si, Q.; Wang, L.; Zhang, Q.; Wu, G.; Zhao, J.; Zhang, H.; Chen, W. Lactobacillus rhamnosus Strains Relieve Loperamide-Induced Constipation via Different Pathways Independent of Short-Chain Fatty Acids. Front. Cell. Infect. Microbiol. 2020, 10, 423. [Google Scholar] [CrossRef]
- Tsilingiri, K.; Rescigno, M. Postbiotics: What else? Benef. Microbes 2013, 4, 101–107. [Google Scholar] [CrossRef]
- Liu, T.; Zhou, K.; Yin, S.; Liu, S.; Zhu, Y.; Yang, Y.; Wang, C. Purification and characterization of an exopolysaccharide produced by Lactobacillus plantarum HY isolated from home-made Sichuan Pickle. Int. J. Biol. Macromol. 2019, 134, 516–526. [Google Scholar] [CrossRef]
- Valladares, R.S.D.; Li, N.; Williams, E.; Lai, K.K.; Abdelgeliel, A.S.; Gonzalez, C.F. Lactobacillus johnsonii N6.2 Mitigates the Developmentof Type 1 Diabetes in BB-DP Rats. PLoS ONE 2010, 5, e10507. [Google Scholar]
- Alberda, C.G.L.; Meddings, J.; Field, C.; McCargar, L.; Kutsogiannis, D.; Fedorak, R. Effects of probiotic therapy in critically ill patients a randomized, double-blind, placebo-controlled trial. Am. J. Clin. Nutr. 2007, 85, 816–823. [Google Scholar] [CrossRef] [Green Version]
- García-Díez, E.; López-Oliva, M.E.; Caro-Vadillo, A.; Pérez-Vizcaíno, F.; Pérez-Jiménez, J.; Ramos, S.; Martín, M. Supplementation with a Cocoa–Carob Blend, Alone or in Combination with Metformin, Attenuates Diabetic Cardiomyopathy, Cardiac Oxidative Stress and Inflammation in Zucker Diabetic Rats. Antioxidants 2022, 11, 432. [Google Scholar] [CrossRef]
- Hayes, J.D.; Dinkova-Kostova, A.T.; Tew, K.D. Oxidative Stress in Cancer. Cancer Cell 2020, 38, 167–197. [Google Scholar] [CrossRef]
- Zhao, S.; Cheng, C.K.; Zhang, C.-L.; Huang, Y. Interplay Between Oxidative Stress, Cyclooxygenases, and Prostanoids in Cardiovascular Diseases. Antioxid. Redox Signal. 2021, 34, 784–799. [Google Scholar] [CrossRef]
- Carroll, I.M.; Andrus, J.M.; Bruno-Barcena, J.M.; Klaenhammer, T.R.; Hassan, H.M.; Threadgill, D.S. Anti-inflammatory properties of Lactobacillus gasseri expressing manganese superoxide dismutase using the interleukin 10-deficient mouse model of colitis. Am. J. Physiol. Gastrointest Liver Physiol. 2007, 293, G729–G738. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Yang, J.; Ye, Z.; Zhang, G.; Nie, W.; Cheng, H.; Peng, M.; Zhang, K.; Liu, J.; Zhang, Z.; et al. Physiologically Inspired Mucin Coated Escherichia coli Nissle 1917 Enhances Biotherapy by Regulating the Pathological Microenvironment to Improve Intestinal Colonization. ACS Nano 2022, 16, 4041–4058. [Google Scholar] [CrossRef]
- Finamore, A.; Ambra, R.; Nobili, F.; Garaguso, I.; Raguzzini, A.; Serafini, M. Redox Role of Lactobacillus casei Shirota Against the Cellular Damage Induced by 2,2’-Azobis (2-Amidinopropane) Dihydrochloride-Induced Oxidative and Inflammatory Stress in Enterocytes-Like Epithelial Cells. Front. Immunol. 2018, 9, 1131. [Google Scholar] [CrossRef] [Green Version]
- Jomehzadeh, N.; Javaherizadeh, H.; Amin, M.; Saki, M.; Al-Ouqaili, M.T.; Hamidi, H.; Seyedmahmoudi, M.; Gorjian, Z. Isolation and identification of potential probiotic Lactobacillus species from feces of infants in southwest Iran. Int. J. Infect. Dis. 2020, 96, 524–530. [Google Scholar] [CrossRef]
- BBazireh, H.; Shariati, P.; Jamalkandi, S.A.; Ahmadi, A.; Boroumand, M.A. Isolation of Novel Probiotic Lactobacillus and Enterococcus Strains From Human Salivary and Fecal Sources. Front. Microbiol. 2020, 11, 597946. [Google Scholar] [CrossRef]
- Lu, W.; Chen, W.; Zhai, Q.; Pan, M.; Xie, M.; Cui, S.; Zhao, J.; Zhang, H. A Method for Screening and/or Identifying Lactobacillus and Application. CN109971871A, 5 July 2019. [Google Scholar]
- Shang, J.; Wan, F.; Zhao, L.; Meng, X.; Li, B. Potential Immunomodulatory Activity of a Selected Strain Bifidobacterium bifidum H3-R2 as Evidenced in vitro and in Immunosuppressed Mice. Front. Microbiol. 2020, 11, 2089. [Google Scholar] [CrossRef]
- Balthazar, C.F.; Silva, H.L.; Esmerino, E.A.; Rocha, R.S.; Moraes, J.; Carmo, M.A.; Azevedo, L.; Camps, I.; Abud, Y.K.; Sant’Anna, C.; et al. The addition of inulin and Lactobacillus casei 01 in sheep milk ice cream. Food Chem. 2018, 246, 464–472. [Google Scholar] [CrossRef]
- Hernando-Amado, S.; Coque, T.M.; Baquero, F.; Martínez, J.L. Defining and combating antibiotic resistance from One Health and Global Health perspectives. Nat. Microbiol. 2019, 4, 1432–1442. [Google Scholar] [CrossRef]
- Young, S.; Rohr, J.R.; Harwood, V.J. Vancomycin resistance plasmids affect persistence of Enterococcus faecium in water. Water Res. 2019, 166, 115069. [Google Scholar] [CrossRef]
- Argyri, A.A.; Zoumpopoulou, G.; Karatzas, K.A.G.; Tsakalidou, E.; Nychas, G.-J.E.; Panagou, E.Z.; Tassou, C.C. Selection of potential probiotic lactic acid bacteria from fermented olives by in vitro tests. Food Microbiol. 2013, 33, 282–291. [Google Scholar] [CrossRef]
- Fabrícioz, L.T.; Winkelströter, L.K.; De Martini, E.C.P. Identification and evaluation of the probiotic potential of Lactobacillus paraplantarum ft259, a bacteriocinogenic strain isolated from brazilian semi-hard artisanal cheese. Anaerobe 2013, 22, 57–63. [Google Scholar]
- Yan, S.; Zhang, X.; Zheng, H.; Hu, D.; Zhang, Y.; Guan, Q.; Liu, L.; Ding, Q.; Li, Y. Clematichinenoside inhibits VCAM-1 and ICAM-1 expression in TNF-alpha-treated endothelial cells via NADPH oxidase-dependent IkappaB kinase/NF-kappaB pathway. Free Radic. Biol. Med. 2015, 78, 190–201. [Google Scholar] [CrossRef]
- Dong, X.; Wu, D.; Zhang, Y.; Jia, L.; Pan, X.; Sun, J.; Pan, L.-L. Cathelicidin Modulates Vascular Smooth Muscle Cell Phenotypic Switching through ROS/IL-6 Pathway. Antioxidants 2020, 9, 491. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Wang, Y.; Li, L.; Liu, S.; Wang, C.; Yuan, Y.; Yang, G.; Chen, Y.; Cheng, J.; Lu, Y.; et al. Mitochondrial ROS promote mitochondrial dysfunction and inflammation in ischemic acute kidney injury by disrupting TFAM-mediated mtDNA maintenance. Theranostics 2021, 11, 1845–1863. [Google Scholar] [CrossRef] [PubMed]
- Kang, R.; Li, R.; Dai, P.; Li, Z.; Li, Y.; Li, C. Deoxynivalenol induced apoptosis and inflammation of IPEC-J2 cells by promoting ROS production. Environ. Pollut. 2019, 251, 689–698. [Google Scholar] [CrossRef]
- Zhang, Q.; Liu, J.; Duan, H.; Li, R.; Peng, W.; Wu, C. Activation of Nrf2/HO-1 signaling: An important molecular mechanism of herbal medicine in the treatment of atherosclerosis via the protection of vascular endothelial cells from oxidative stress. J. Adv. Res. 2021, 34, 43–63. [Google Scholar] [CrossRef]
- Xing, J.; Wang, F.; Xu, Q.; Yin, B.; Fang, D.; Zhao, J.; Zhang, H.; Chen, Y.Q.; Wang, G.; Chen, W. Screening of potential probiotic lactic acid bacteria based on gastrointestinal properties and perfluorooctanoate toxicity. Appl. Microbiol. Biotechnol. 2016, 100, 6755–6766. [Google Scholar] [CrossRef]
- Rooks, M.G.; Garrett, W.S. Gut microbiota, metabolites and host immunity. Nat. Rev. Immunol. 2016, 16, 341–352. [Google Scholar] [CrossRef]
- Shaukat, A.; Levitt, M.D.; Taylor, B.C.; MacDonald, R.; Shamliyan, T.A.; Kane, R.L.; Wilt, T.J. Systematic review: Effective management strategies for lactose intolerance. Ann. Intern. Med. 2010, 152, 797–803. [Google Scholar] [CrossRef]
- Munoz-Quezada, S.; Chenoll, E.; Vieites, J.M.; Genoves, S.; Maldonado, J.; Bermudez-Brito, M.; Gomez-Llorente, C.; Matencio, E.; José Bernal, M.; Romero, F.; et al. Isolation, identification and characterisation of three novel probiotic strains (Lactobacillus paracasei CNCM I-4034, Bifidobacterium breve CNCM I-4035 and Lactobacillus rhamnosus CNCM I-4036) from the faeces of exclusively breast-fed infants. Br. J. Nutr. 2013, 109 (Suppl. S2), S51–S62. [Google Scholar] [CrossRef] [Green Version]
- Gheziel, C.; Russo, P.; Arena, M.P.; Spano, G.; Ouzari, H.-I.; Kheroua, O.; Saidi, D.; Fiocco, D.; Kaddouri, H.; Capozzi, V. Evaluating the Probiotic Potential of Lactobacillus plantarum Strains from Algerian Infant Feces: Towards the Design of Probiotic Starter Cultures Tailored for Developing Countries. Probiotics Antimicrob. Proteins 2018, 11, 113–123. [Google Scholar] [CrossRef]
- El-Deeb, W.M.; Fayez, M.; Elsohaby, I.; Ghoneim, I.; Al-Marri, T.; Kandeel, M.; Gioushy, M. Isolation and characterization of vaginal Lactobacillus spp. in dromedary camels (Camelus dromedarius): In vitro evaluation of probiotic potential of selected isolates. PeerJ 2020, 8, e8500. [Google Scholar] [CrossRef] [Green Version]
- Colomer-Winter, C.; Flores-Mireles, A.L.; Kundra, S.; Hultgren, S.J.J.A.L. (p)ppGpp and CodY Promote Enterococcus faecalis Virulence in a Murine Model of Catheter-Associated Urinary Tract Infection. mSphere 2019, 24, e00392-19. [Google Scholar] [CrossRef] [Green Version]
- Liu, F.; Sun, Z.; Wang, F.; Liu, Y.; Zhu, Y.; Du, L.; Wang, D.; Xu, W. Inhibition of biofilm formation and exopolysaccharide synthesis of Enterococcus faecalis by phenyllactic acid. Food Microbiol. 2019, 86, 103344. [Google Scholar] [CrossRef]
- Kaur, M.; Singh, H.; Jangra, M.; Kaur, L.; Jaswal, P.; Dureja, C.; Nandanwar, H.; Chaudhuri, S.R.; Raje, M.; Mishra, S.; et al. Lactic acid bacteria isolated from yak milk show probiotic potential. Appl. Microbiol. Biotechnol. 2017, 101, 7635–7652. [Google Scholar] [CrossRef]
- Liu, W.; Chen, M.; Duo, L.; Wang, J.; Guo, S.; Sun, H.; Menghe, B.; Zhang, H. Characterization of potentially probiotic lactic acid bacteria and bifidobacteria isolated from human colostrum. J. Dairy Sci. 2020, 103, 4013–4025. [Google Scholar] [CrossRef]
- Talib, N.; Mohamad, N.E.; Yeap, S.K.; Hussin, Y.; Aziz, M.N.M.; Masarudin, M.J.; Sharifuddin, S.A.; Hui, Y.W.; Ho, C.L.; Alitheen, N.B. Isolation and Characterization of Lactobacillus spp. from Kefir Samples in Malaysia. Molecules 2019, 24, 2606. [Google Scholar] [CrossRef] [Green Version]
- Ruas-Madiedo, P.; Gueimonde, M.; Arigoni, F.; Reyes-Gavilán, C.G.D.L.; Margolles, A. Bile Affects the Synthesis of Exopolysaccharides by Bifidobacterium animalis. Appl. Environ. Microbiol. 2009, 75, 1204–1207. [Google Scholar] [CrossRef] [Green Version]
- Ruiz, L.; Sanchez, B.; Ruas-Madiedo, P.; de Los Reyes-Gavilan, C.G.; Margolles, A. Cell envelope changes in Bifidobacterium animalis ssp. lactis as a response to bile. FEMS Microbiol. Lett. 2007, 274, 316–322. [Google Scholar] [CrossRef] [Green Version]
- Gueimonde, M.; Margolles, A.; de los Reyes-Gavilán, C.G.; Salminen, S. Competitive exclusion of enteropathogens from human intestinal mucus by Bifidobacterium strains with acquired resistance to bile—A preliminary study. Int. J. Food Microbiol. 2007, 113, 228–232. [Google Scholar] [CrossRef]
- Temmerman, R.; Pot, B.; Huys, G.; Swings, J. Identification and antibiotic susceptibility of bacterial isolates from probiotic products. Int. J. Food Microbiol. 2003, 81, 1–10. [Google Scholar] [CrossRef]
- Sharma, P.; Tomar, S.K.; Goswami, P.; Sangwan, V.; Singh, R. Antibiotic resistance among commercially available probiotics. Food Res. Int. 2014, 57, 176–195. [Google Scholar] [CrossRef]
- Gueimonde, M.; Sánchez, B.; de Los Reyes-Gavilán, C.G.; Margolles, A. Antibiotic resistance in probiotic bacteria. Front. Microbiol. 2013, 4, 202. [Google Scholar] [CrossRef] [Green Version]
- Monteagudo-Mera, A.; Rodríguez-Aparicio, L.; Rúa, J.; Martínez-Blanco, H.; Navasa, N.; García-Armesto, M.R.; Ferrero, M.Á. In vitro evaluation of physiological probiotic properties of different lactic acid bacteria strains of dairy and human origin. J. Funct. Foods 2012, 4, 531–541. [Google Scholar] [CrossRef]
- Izuddin, W.I.; Humam, A.M.; Loh, T.C.; Foo, H.L.; Samsudin, A.A. Dietary Postbiotic Lactobacillus plantarum Improves Serum and Ruminal Antioxidant Activity and Upregulates Hepatic Antioxidant Enzymes and Ruminal Barrier Function in Post-Weaning Lambs. Antioxidants 2020, 9, 250. [Google Scholar] [CrossRef] [Green Version]
- Kumar, A.; Joishy, T.; Das, S.; Kalita, M.C.; Mukherjee, A.K.; Khan, M.R. A Potential Probiotic Lactobacillus plantarum JBC5 Improves Longevity and Healthy Aging by Modulating Antioxidative, Innate Immunity and Serotonin-Signaling Pathways in Caenorhabditis elegans. Antioxidants 2022, 11, 268. [Google Scholar] [CrossRef]
- Wu, Y.; Wang, B.; Tang, L.; Zhou, Y.; Wang, Q.; Gong, L.; Ni, J.; Li, W. Probiotic Bacillus Alleviates Oxidative Stress-Induced Liver Injury by Modulating Gut-Liver Axis in a Rat Model. Antioxidants 2022, 11, 291. [Google Scholar] [CrossRef]
- Loboda, A.; Damulewicz, M.; Pyza, E.; Jozkowicz, A.; Dulak, J. Role of Nrf2/HO-1 system in development, oxidative stress response and diseases: An evolutionarily conserved mechanism. Cell. Mol. Life Sci. 2016, 73, 3221–3247. [Google Scholar] [CrossRef] [Green Version]
- Luo, J.-F.; Shen, X.-Y.; Lio, C.-K.; Dai, Y.; Cheng, C.-S.; Liu, J.-X.; Yao, Y.-D.; Yu, Y.; Xie, Y.; Luo, P.; et al. Activation of Nrf2/HO-1 Pathway by Nardochinoid C Inhibits Inflammation and Oxidative Stress in Lipopolysaccharide-Stimulated Macrophages. Front. Pharmacol. 2018, 9, 911. [Google Scholar] [CrossRef]
- Murray, P.J.; Wynn, T.A. Protective and pathogenic functions of macrophage subsets. Nat. Rev. Immunol. 2011, 11, 723–737. [Google Scholar] [CrossRef]
- Yuan, T.; Yang, T.; Chen, H.; Fu, D.; Hu, Y.; Wang, J.; Yuan, Q.; Yu, H.; Xu, W.; Xie, X. New insights into oxidative stress and inflammation during diabetes mellitus-accelerated atherosclerosis. Redox Biol. 2019, 20, 247–260. [Google Scholar] [CrossRef]
- Zhou, R.; Yazdi, A.S.; Menu, P.; Tschopp, J. A role for mitochondria in NLRP3 inflammasome activation. Nature 2011, 469, 221–225. [Google Scholar] [CrossRef]
- Wang, L.; Zhao, Z.; Zhao, L.; Zhao, Y.; Yang, G.; Wang, C.; Gao, L.; Niu, C.; Li, S. Lactobacillus plantarum DP189 Reduces alpha-SYN Aggravation in MPTP-Induced Parkinson’s Disease Mice via Regulating Oxidative Damage, Inflammation, and Gut Microbiota Disorder. J. Agric. Food Chem. 2022, 70, 1163–1173. [Google Scholar] [CrossRef]
- Gao, D.; Gao, Z.; Zhu, G. Antioxidant effects of Lactobacillus plantarum via activation of transcription factor Nrf2. Food Funct. 2013, 4, 982–989. [Google Scholar] [CrossRef]
- Li, J.; Li, Q.; Gao, N.; Wang, Z.; Li, F.; Li, J.; Shan, A. Exopolysaccharides produced by Lactobacillus rhamnosus GG alleviate hydrogen peroxide-induced intestinal oxidative damage and apoptosis through the Keap1/Nrf2 and Bax/Bcl-2 pathways in vitro. Food Funct. 2021, 12, 9632–9641. [Google Scholar] [CrossRef]
Figure 1.
The specific-PCR detection of 27 bacterial isolates. “P” represents positive LAB (LGG). “M” represents the DNA marker. Numbers 1–27 are strains selected in the present study.
Figure 1.
The specific-PCR detection of 27 bacterial isolates. “P” represents positive LAB (LGG). “M” represents the DNA marker. Numbers 1–27 are strains selected in the present study.
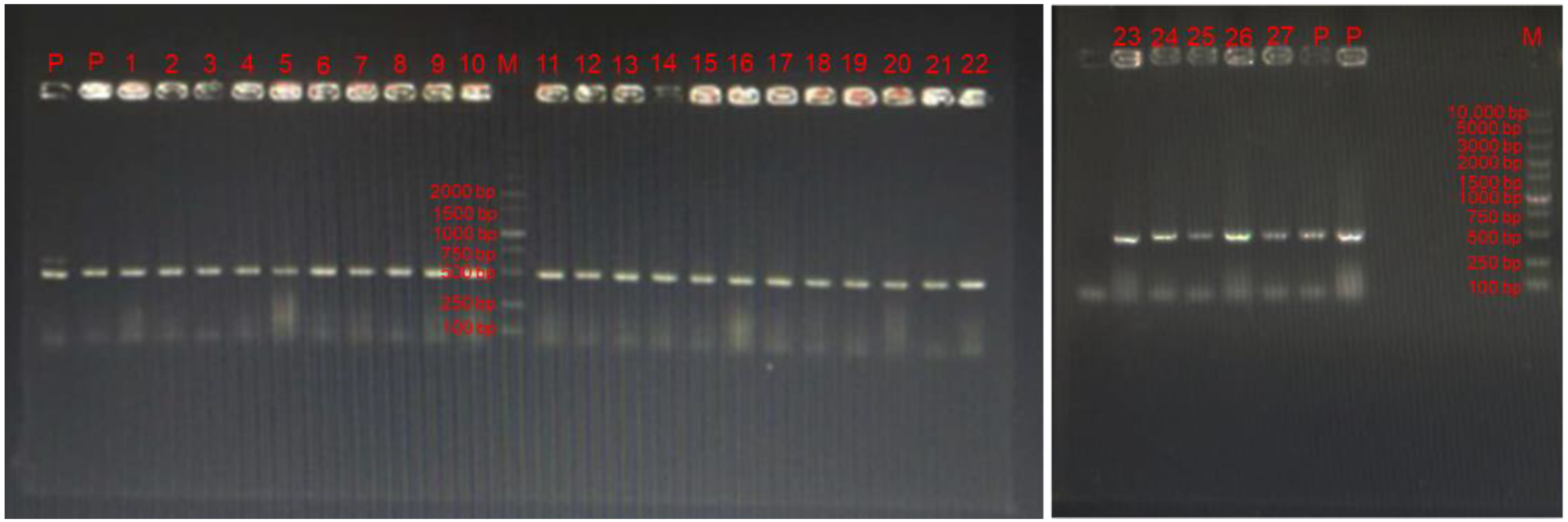
Figure 2.
The survival rates of the selected strains in simulated digestive tract conditions. All data were expressed as mean ± SD (n = 3 independent experiments). Statistical significance was defined as * p < 0.05; ** p < 0.01; *** p < 0.001. “ns”: no significance. (A) Differences of selected strains in artificial salvia and gastric fluid compared with reference strain LGG. (B) Differences of selected strains in simulated digestive tract compared with LGG.
Figure 2.
The survival rates of the selected strains in simulated digestive tract conditions. All data were expressed as mean ± SD (n = 3 independent experiments). Statistical significance was defined as * p < 0.05; ** p < 0.01; *** p < 0.001. “ns”: no significance. (A) Differences of selected strains in artificial salvia and gastric fluid compared with reference strain LGG. (B) Differences of selected strains in simulated digestive tract compared with LGG.

Figure 3.
The adhesion ability of the selected strains to Caco-2 cells in vitro. All data were expressed as mean ± SD (n = 3 independent experiments). Statistical significance was defined as * p < 0.05; *** p < 0.001. Reported statistical significance refers to comparisons with LGG.
Figure 3.
The adhesion ability of the selected strains to Caco-2 cells in vitro. All data were expressed as mean ± SD (n = 3 independent experiments). Statistical significance was defined as * p < 0.05; *** p < 0.001. Reported statistical significance refers to comparisons with LGG.

Figure 4.
Selected probiotic LAB (L. gasseri FWJL-4, L. plantarum Fjias-5 and L. rhamnosus FSJ-13) show anti-inflammatory effects in LPS-treated RAW 264.7 murine macrophages. (A) The mRNA expression level of TNFα.(B) The mRNA expression level of IL-6. (C) The mRNA expression level of IL-1β. (D) The mRNA expression level of IL-10. All data were expressed as mean ± SD (n = 3 independent experiments). Statistical significance was defined as * p < 0.05; *** p < 0.001. Reported statistical significance refers to comparisons with cell-treated with LPS.
Figure 4.
Selected probiotic LAB (L. gasseri FWJL-4, L. plantarum Fjias-5 and L. rhamnosus FSJ-13) show anti-inflammatory effects in LPS-treated RAW 264.7 murine macrophages. (A) The mRNA expression level of TNFα.(B) The mRNA expression level of IL-6. (C) The mRNA expression level of IL-1β. (D) The mRNA expression level of IL-10. All data were expressed as mean ± SD (n = 3 independent experiments). Statistical significance was defined as * p < 0.05; *** p < 0.001. Reported statistical significance refers to comparisons with cell-treated with LPS.

Figure 5.
Selected probiotic LAB (L. gasseri FWJL-4, L. plantarum Fjias-5 and L. rhamnosus FSJ-13) enhance antioxidant enzymes and reduce H2O2 production in LPS-treated RAW 264.7 murine macrophages. (A) The enzymatic activities of SOD in different groups. (B) The enzymatic activity of GPx in different groups. (C) The ratio of GSH/GSSG in different groups. (D) The generation of cellular H2O2 at 2 h, 4 h, 6 h, 8 h, 10 h and 12 h in different groups. All data were expressed as mean ± SD (n = 3 independent experiments). Statistical significance was defined as * p < 0.05; ** p < 0.01; *** p < 0.001. “ns”: no significance. Reported statistical significance refers to comparisons with cell-treated with LPS.
Figure 5.
Selected probiotic LAB (L. gasseri FWJL-4, L. plantarum Fjias-5 and L. rhamnosus FSJ-13) enhance antioxidant enzymes and reduce H2O2 production in LPS-treated RAW 264.7 murine macrophages. (A) The enzymatic activities of SOD in different groups. (B) The enzymatic activity of GPx in different groups. (C) The ratio of GSH/GSSG in different groups. (D) The generation of cellular H2O2 at 2 h, 4 h, 6 h, 8 h, 10 h and 12 h in different groups. All data were expressed as mean ± SD (n = 3 independent experiments). Statistical significance was defined as * p < 0.05; ** p < 0.01; *** p < 0.001. “ns”: no significance. Reported statistical significance refers to comparisons with cell-treated with LPS.

Figure 6.
Selected probiotic LAB (L. gasseri FWJL-4, L. plantarum Fjias-5 and L. rhamnosus FSJ-13) induce Nrf2-mediated signaling pathways. All data were expressed as mean ± SD. * p < 0.05; ** p < 0.01; *** p < 0.001 were considered as statistical significance. Reported statistical significance refers to comparisons with cell-treated with LPS.
Figure 6.
Selected probiotic LAB (L. gasseri FWJL-4, L. plantarum Fjias-5 and L. rhamnosus FSJ-13) induce Nrf2-mediated signaling pathways. All data were expressed as mean ± SD. * p < 0.05; ** p < 0.01; *** p < 0.001 were considered as statistical significance. Reported statistical significance refers to comparisons with cell-treated with LPS.

Figure 7.
Proposed molecular mechanisms underlying the inhibitory effect of LAB (isolated from infant feces) on the activation of macrophages induced by LPS. LAB activates Nrf2/HO-1 pathway to regulate cellular redox status and reduce oxidative stress, which in turn inhibits inflammation. Nrf2: nuclear factor-erythroid factor 2-related factor 2; ARE: antioxidant responsive element; HO-1: heme oxygenase-1; SOD: superoxide dismutase; GPx: glutathione peroxidase; GSH/GSSG: glutathione/oxidized glutathione; IL-6: Interleukin-6; IL-1β: Interleukin-1β; TNFα: tumor necrosis factor α.
Figure 7.
Proposed molecular mechanisms underlying the inhibitory effect of LAB (isolated from infant feces) on the activation of macrophages induced by LPS. LAB activates Nrf2/HO-1 pathway to regulate cellular redox status and reduce oxidative stress, which in turn inhibits inflammation. Nrf2: nuclear factor-erythroid factor 2-related factor 2; ARE: antioxidant responsive element; HO-1: heme oxygenase-1; SOD: superoxide dismutase; GPx: glutathione peroxidase; GSH/GSSG: glutathione/oxidized glutathione; IL-6: Interleukin-6; IL-1β: Interleukin-1β; TNFα: tumor necrosis factor α.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Primers used for RT-qPCR.
| Gene | Sequence (5′-3′) | Product Size (bp) | Accession Number | |
|---|---|---|---|---|
| β-Actin | Forward | ATGACCCAAGCCGAGAAGG | 185 | NM_027493 |
| Reverse | CGGCCAAGTCTTAGAGTTGTTG | |||
| Tnfα | Forward | CCACGCTCTTCTGTCTACTG | 169 | NM_010851.2 |
| Reverse | ACTTGGTGGTTTGCTACGAC | |||
| Il-10 | Forward | GGACCAGCTGGACAACATACTGCTA | 80 | NM_010548.2 |
| Reverse | CCGATAAGGCTTGGCAACCCAAGT | |||
| Il-6 | Forward | GAGTCACAGAAGGAGTGGCTAAGG | 106 | NM_031168.1 |
| Reverse | CGCACTAGGTTTGCCGAGTAGATCT | |||
| Il-1β | Forward | TTGAAAGTCCACCTCCTTACAGA | 129 | NM_008756 |
| Reverse | CCGGATAAAAAGAGTACGCTGG |
Table 2.
a Results of the antimicrobial activity of selected isolates.
| Strains | Staphylococcus aureus ATCC 25923 | Salmonella enterica ATCC 14028 | Escherichia coli ATCC 25922 | Listeria monocytogenes ATCC 13932 | Enterococcus faecalis E27 |
|---|---|---|---|---|---|
| FRY-2 | 10.67 ± 0.5 | 14.3 ± 1 | 11.67 ± 0.5 | 12.7 ± 0.5 | 14.7 ± 1.53 |
| FXHB-6 | 11.33 ± 0 | 12.67 ± 1.1 | 10.3 ± 0.4 | 13.7 ± 0.5 | 15.7 ± 0.5 |
| FRY-3 | 8.7 ± 1.0 | 8.3 ± 0.5 | 10.7 ± 0.5 | 11.3 ± 0.5 | 15.3 ± 1.1 |
| FMM-1 | - | - | - | - | 11.3 ± 0.5 |
| FZL-21 | 11.7 ± 1.1 | 10.7 ± 0.5 | 12.3 ± 1.1 | 11 ± 1.0 | 17 ± 1.0 |
| Fjias-5 | 19.3 ± 1.0 | 14.33 ± 0.5 | 20.3 ± 0.5 | 19.67 ± 0.4 | 19.3 ± 1.1 |
| FWJL-4 | 14.67 ± 0.5 | 19.7 ± 1.1 | 19.33 ± 0.5 | 14.7 ± 0.5 | 21.7 ± 0.5 |
| FLY-17 | - | - | - | - | 10.7 ± 1.1 |
| FZL-2 | 16.67 ± 1.7 | 12.7 ± 1.1 | 15 ± 0 | 12.67 ± 0.5 | 15 ± 1.0 |
| FRY-6 | 15.3 ± 0.5 | 12.3 ± 1.1 | 11.67 ± 0.5 | 14.33 ± 0.5 | 18.7 ± 0.5 |
| FMM-7 | - | - | - | - | - |
| FSJ-1 | 12.3 ± 0.5 | 11.3 ± 0.5 | 10.6 ± 0.5 | 9.33 ± 0.5 | 17.3 ± 1.1 |
| FWJL-5 | 13.7 ± 0.58 | 17 ± 1.0 | 14.7 ± 0.5 | 13.6 ± 0.5 | 20.7 ± 1.1 |
| FHHY-7 | - | - | - | - | 15.3 ± 1.1 |
| FSJ-11 | 14.33 ± 1.0 | 15.7 ± 1.1 | 12.67 ± 0.5 | 18.3 ± 0.5 | 18.7 ± 1.5 |
| FSJ-13 | 18.7 ± 1.5 | 19.7 ± 0.5 | 18.3 ± 0.5 | 17.3 ± 1.1 | 20.7 ± 1.1 |
| FZSJ-7 | 11.3 ± 0.4 | 12.7 ± 0.5 | 14.33 ± 0.5 | 12.7 ± 0.5 | 18 ± 1.0 |
| FSJ-6 | - | - | - | 8.7 ± 1.1 | - |
| FXHB-2 | 12.67 ± 0.5 | 17 ± 1.0 | 19 ± 1.0 | 17.3 ± 1.15 | 18.67 ± 0.5 |
| FSJ-4X | 14.33 ± 0.5 | 17 ± 0.5 | 13.3 ± 0.5 | 17 ± 1.0 | 19 ± 1.0 |
| FLT6-11 | 8.67 ± 0.4 | - | - | 8.3 ± 0.5 | - |
| FXHB-2X | 9.3 ± 1.1 | 8.3 ± 0.5 | 16 ± 1.0 | 17.3 ± 1.1 | 15.3 ± 0.5 |
| FRY-4X | 9.7 ± 0.5 | 14.3 ± 1.15 | 13.7 ± 0.5 | 11 ± 1.0 | 20.7 ± 1.1 |
| FSJ-2 | 10.3 ± 0.4 | 9.7 ± 0.5 | 9 ± 1.0 | 8.7 ± 1.53 | 15.3 ± 0.5 |
| FHHY-1 | 16.3 ± 0.5 | 15 ± 1.0 | 12.7 ± 0.5 | 15.7 ± 0.5 | 20.67 ± 1.15 |
| FZYY-3 | 10.7 ± 0.5 | 9.67 ± 0.5 | 8.33 ± 0.5 | 10.7 ± 0.5 | 15.67 ± 0.5 |
| FQM-3X | - | - | - | - | - |
| Penicillin | 32.7 ± 0.5 | 30.3 ± 1.15 | 35.7 ± 0.5 | 33.3 ± 0.4 | 38.67 ± 0 |
a No inhibition (–).
Table 3.
Antibiotic resistance of tested strains.
| Strains | Name | a MIC (μg/mL) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| K | P | V | E | A | S | C | PO | ||
| FRY-2 | Lactobacillus gasseri | 16 | 4 | 4 | <2 | <2 | 2 | 2 | ≥1024 |
| FXHB-6 | Lactobacillus gasseri | 16 | 32 | 2 | <2 | <2 | 2 | 2 | ≥1024 |
| FRY-3 | Lactobacillus gasseri | 16 | 4 | 2 | <2 | 2 | 2 | 2 | 1024 |
| FZL-21 | Lactobacillus gasseri | 64 | 4 | 2 | <2 | 2 | 2 | 2 | ≥1024 |
| Fjias-5 | Lactiplantibacillus plantarum | 512 | <2 | 2 | <2 | <2 | 16 | 8 | ≥1024 |
| FRY-4X | Lactobacillus gasseri | <2 | 8 | 2 | <2 | <2 | 2 | <2 | ≥1024 |
| FWJL-4 | Lactobacillus gasseri | 64 | 8 | 2 | <2 | 8 | 16 | 2 | 1024 |
| FZL-2 | Lactobacillus gasseri | 64 | 16 | 4 | <2 | 64 | 2 | 2 | ≥1024 |
| FRY-6 | Lactobacillus gasseri | 64 | 16 | 8 | 8 | 4 | 8 | 2 | ≥1024 |
| FSJ-1 | Lacticaseibacillus rhamnosus | 512 | 512 | ≥1024 | 8 | 16 | 32 | 2 | ≥1024 |
| FWJL-5 | Lactobacillus gasseri | 64 | 32 | 2 | <2 | 8 | 2 | 2 | 1024 |
| FSJ-11 | Lacticaseibacillus rhamnosus | 512 | 2 | ≥1024 | 2 | <2 | 32 | <2 | ≥1024 |
| FSJ-13 | Lacticaseibacillus rhamnosus | ≥1024 | <2 | ≥1024 | 2 | 2 | 128 | <2 | ≥1024 |
| FZSJ-7 | Lactobacillus gasseri | 16 | 4 | ≥1024 | <2 | <2 | <2 | <2 | ≥1024 |
| FXHB-2 | Lactobacillus gasseri | 16 | 4 | ≥1024 | <2 | <2 | <2 | <2 | ≥1024 |
| FSJ-4X | Lacticaseibacillus rhamnosus | 32 | <2 | ≥1024 | 2 | <2 | 32 | <2 | ≥1024 |
| FXHB-2X | Lactobacillus gasseri | 128 | 4 | ≥1024 | <2 | 8 | 4 | <2 | ≥1024 |
| LGG | Lacticaseibacillus rhamnosus | 256 | 8 | ≥1024 | <2 | 32 | 64 | 8 | ≥1024 |
a K: kanamycin, P: penicillin, V: vancomycin, E: erythromycin, A: ampicillin, S: streptomycin, C: chloramphenicol, PO: polymyxin.
Table 4.
16S rRNA sequencing of candidate probiotics based on the percentage similarity of the sequence found in the GenBank database.
Table 4.
16S rRNA sequencing of candidate probiotics based on the percentage similarity of the sequence found in the GenBank database.
| Names | Sequencing Results | Identity | Accession Number |
|---|---|---|---|
| FRY-2 | Lactobacillus gasseri | 97.90% | MZ220367 |
| FXHB-6 | Lactobacillus gasseri | 99.79% | MZ220368 |
| Fjias-5 | Lactiplantibacillus plantarum | 99.93% | MZ220369 |
| FRY-4X | Lactobacillus gasseri | 99.79% | MZ220370 |
| FWJL-4 | Lactobacillus gasseri | 99.79% | MZ220371 |
| FZL-2 | Lactobacillus gasseri | 99.79% | MZ220372 |
| FRY-6 | Lactobacillus gasseri | 93.73% | MZ220373 |
| FSJ-13 | Lacticaseibacillus rhamnosus | 99.80% | MZ220374 |
| FZSJ-7 | Lactobacillus gasseri | 99.79% | MZ220375 |
| FXHB-2 | Lactobacillus gasseri | 99.86% | MZ220376 |
| FWJL-5 | Lactobacillus gasseri | 99.79% | MZ220377 |
| FSJ-4X | Lacticaseibacillus rhamnosus | 99.86% | MZ220378 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Li, B.; Pan, L.-L.; Sun, J. Novel Probiotic Lactic Acid Bacteria Were Identified from Healthy Infant Feces and Exhibited Anti-Inflammatory Capacities. Antioxidants 2022, 11, 1246. https://doi.org/10.3390/antiox11071246
AMA Style
Li B, Pan L-L, Sun J. Novel Probiotic Lactic Acid Bacteria Were Identified from Healthy Infant Feces and Exhibited Anti-Inflammatory Capacities. Antioxidants. 2022; 11(7):1246. https://doi.org/10.3390/antiox11071246
Chicago/Turabian StyleLi, Binbin, Li-Long Pan, and Jia Sun. 2022. "Novel Probiotic Lactic Acid Bacteria Were Identified from Healthy Infant Feces and Exhibited Anti-Inflammatory Capacities" Antioxidants 11, no. 7: 1246. https://doi.org/10.3390/antiox11071246
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.