
Flaxseed Ethanol Extracts’ Antitumor, Antioxidant, and Anti-Inflammatory Potential

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Cell Cultures
2.2. Plant Extract Preparation
2.3. Phytochemical Analysis
2.3.1. Total Polyphenols Content
2.3.2. Total Flavonoids Content
2.3.3. Identification and Quantification of Polyphenolic Compounds by HPLC-DAD-ESI MS
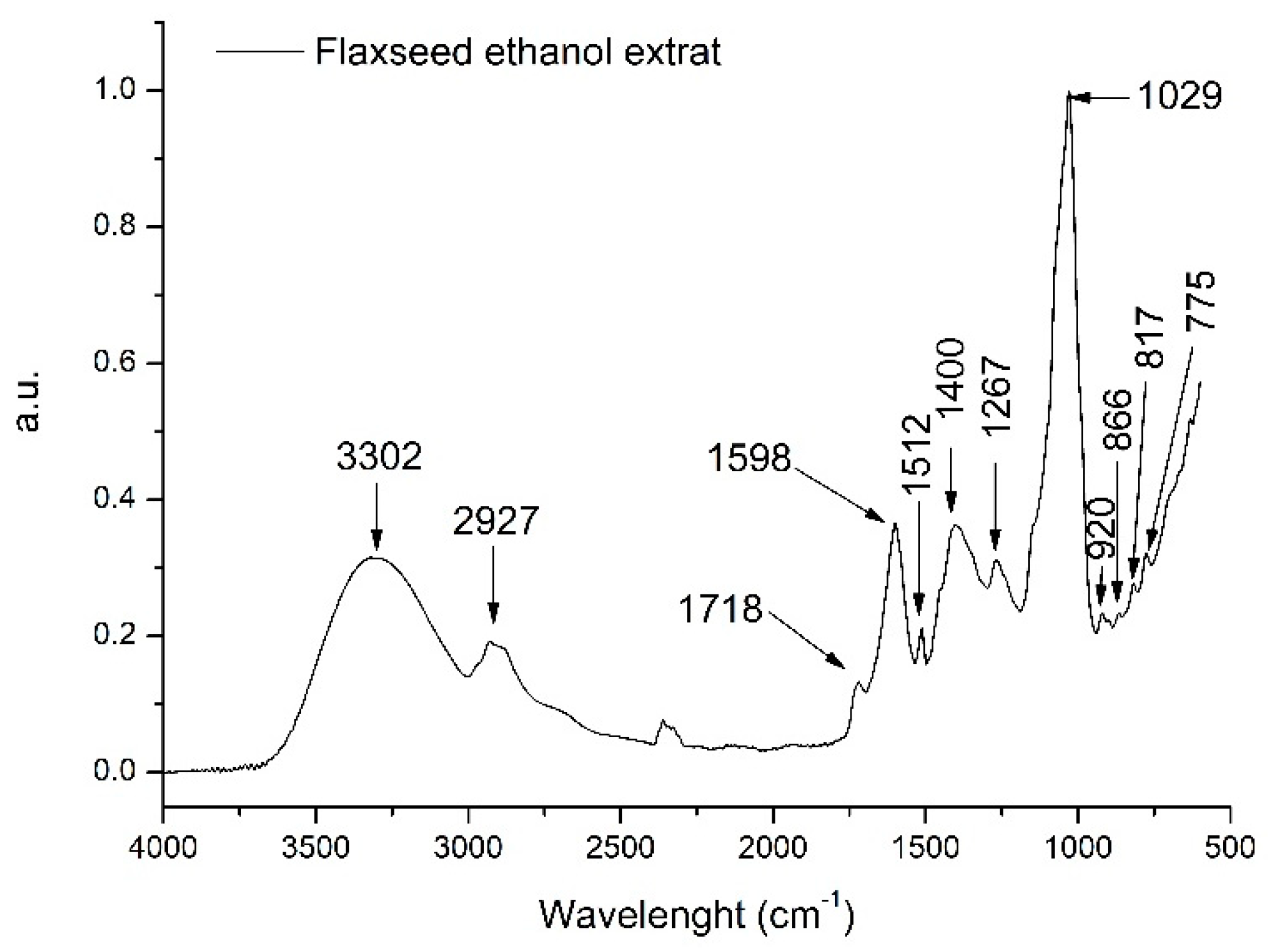
2.3.4. Identification and Quantification of Polyphenolic Compounds by FTIR
2.4. In Vitro Antioxidant Activity Analysis
2.5. Cell Cultures and Cytotoxicity Test
2.5.1. Cell Lines
2.5.2. MTT Viability Assay
2.6. Experimental Design
2.6.1. Animal Subjects
2.6.2. Experimental Protocol
2.6.3. EAC Tumour Cell Viability
2.7. In Vivo Oxidative Stress and Anti-Inflammatory Markers
2.8. Statistical Analysis
3. Results
3.1. Phytochemical Analysis
3.2. In Vitro Antioxidant Activity Analysis
3.2.1. DPPH, FRAP, H2O2, and NO Analysis
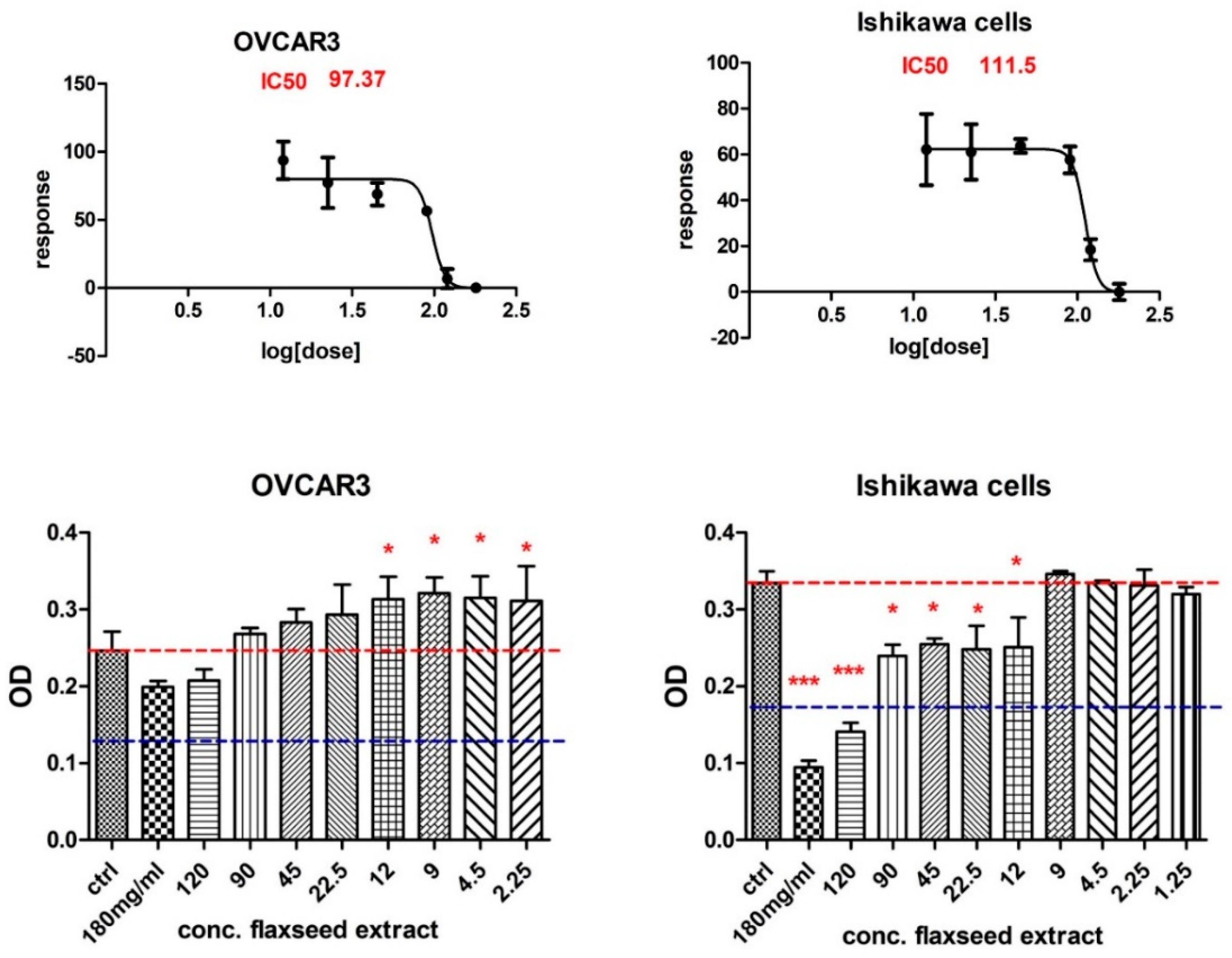
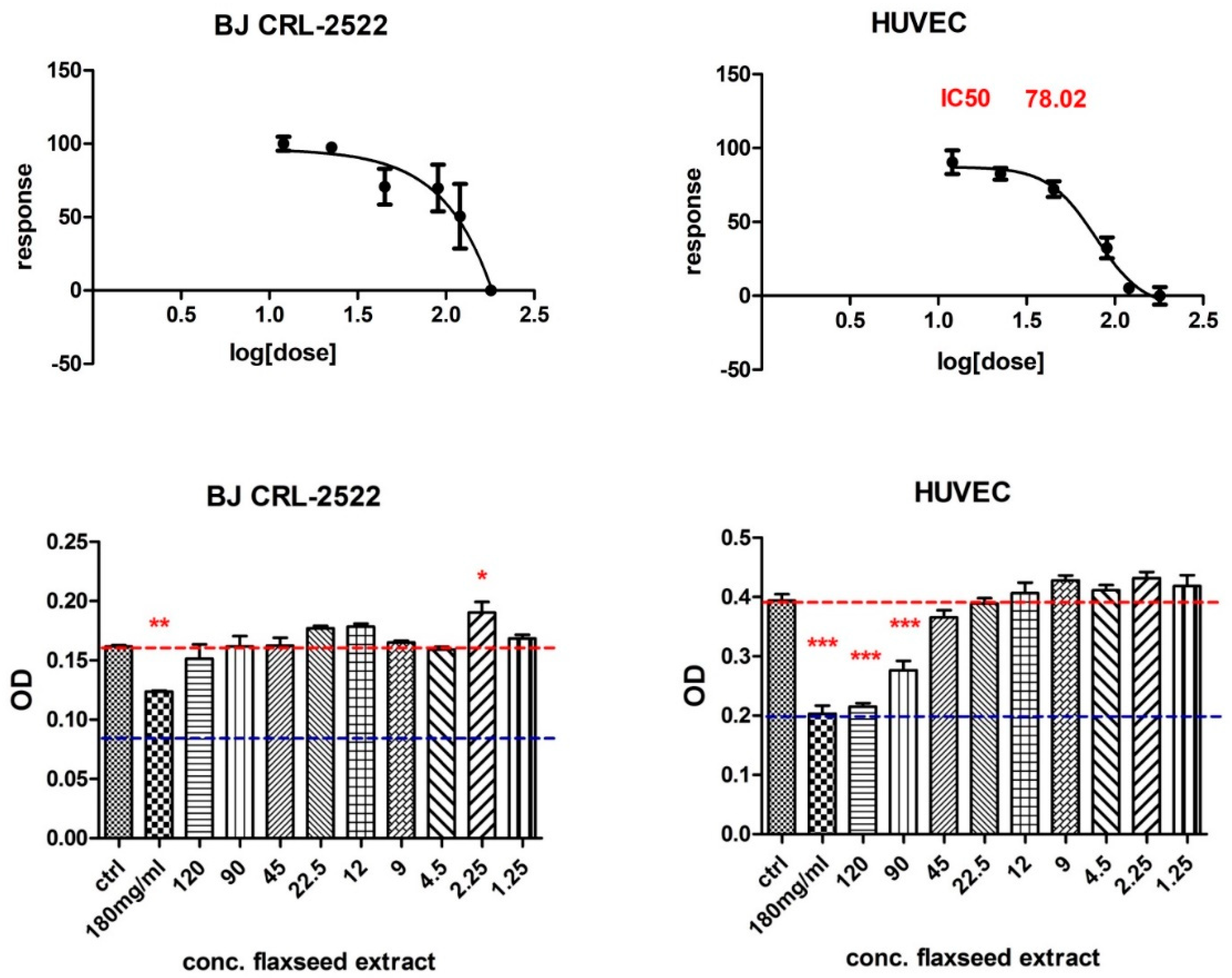
3.2.2. MTT Cytotoxicity Test
3.3. Flaxseeds Extract Treatment Effects on the Body Weight, Ascites Volume, and Viable/Nonviable Cell Count in EAC-Bearing Mice
3.4. In Vivo Antioxidative and Anti-Inflammatory Effect
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Santhi, R.; Annapurani, S. Preliminary evaluation of In vitro and In vivo antioxidative and antitumor activities of flavonoid extract of Tabernaemontana divaricata leaves in Ehrlich’s lymphoma and Dalton’s lymphoma ascites model. J. Cancer Res. Ther. 2020, 16, 78–87. [Google Scholar] [CrossRef]
- Kabir, M.; Al-Noman, A.; Dash, B.K.; Hasan, M.; Akhter, S.; Rahman, M. Trema orientalis (Linn.) leaves promotes anticancer activity in Ehrlich ascites carcinoma (EAC) in Swiss albino mice. J. Basic Clin. Physiol. Pharmacol. 2020, 31, 1–12. [Google Scholar] [CrossRef]
- Barhoi, D.; Upadhaya, P.; Barbhuiya, S.N.; Giri, A.; Giri, S. Aqueous Extract of Moringa oleifera Exhibit Potential Anticancer Activity and can be Used as a Possible Cancer Therapeutic Agent: A Study Involving In Vitro and In Vivo Approach. J. Am. Coll. Nutr. 2021, 40, 70–85. [Google Scholar] [CrossRef]
- Donia, T.I.K.; Gerges, M.N.; Mohamed, T.M. Amelioration effect of Egyptian sweet orange hesperidin on Ehrlich ascites carcinoma (EAC) bearing mice. Chem. Biol. Interact. 2018, 285, 76–84. [Google Scholar] [CrossRef]
- Santos, J.; Brito, M.; Ferreira, R.; Moura, A.P.; Sousa, T.; Batista, T.; Mangueira, V.; Leite, F.; Cruz, R.; Vieira, G.; et al. Th1-biased immunomodulation and in vivo antitumor effect of a novel piperine analogue. Int. J. Mol. Sci. 2018, 19, 2594. [Google Scholar] [CrossRef] [Green Version]
- Mohamed, A.S. Echinochrome Exhibits Antitumor Activity against Ehrlich Ascites Carcinoma in Swiss Albino Mice. Nutr. Cancer 2021, 73, 124–132. [Google Scholar] [CrossRef]
- Biswas, S.K. Does the Interdependence between Oxidative Stress and Inflammation Explain the Antioxidant Paradox? Oxid. Med. Cell. Longev. 2016, 2016, 5698931. [Google Scholar] [CrossRef] [Green Version]
- Buckner, A.L.; Buckner, C.A.; Montaut, S.; Lafrenie, R.M. Treatment with flaxseed oil induces apoptosis in cultured malignant cells. Heliyon 2019, 5, e02251. [Google Scholar] [CrossRef] [Green Version]
- Dikshit, A.; Filho, M.A.G.; Eilati, E.; McGee, S.; Small, C.; Gao, C.; Klug, T.; Hales, D.B. Flaxseed reduces the pro-carcinogenic micro-environment in the ovaries of normal hens by altering the PG and oestrogen pathways in a dose-dependent manner. Br. J. Nutr. 2015, 113, 1384–1395. [Google Scholar] [CrossRef] [Green Version]
- Mareti, E.; Abatzi, C.; Vavilis, D.; Lambrinoudaki, I.; Goulis, D.G. Effect of oral phytoestrogens on endometrial thickness and breast density of perimenopausal and postmenopausal women: A systematic review and meta-analysis. Maturitas 2019, 124, 81–88. [Google Scholar] [CrossRef]
- Liu, H.X.; Wang, Y.; Lu, Q.; Yang, M.Z.; Fan, G.W.; Karas, R.H.; Gao, X.M.; Zhu, Y. Bidirectional regulation of angiogenesis by phytoestrogens through estrogen receptor-mediated signaling networks. Chin. J. Nat. Med. 2016, 14, 241–254. [Google Scholar] [CrossRef]
- Imran, M.; Ahmad, N.; Anjum, F.M.; Khan, M.K.; Mushtaq, Z.; Nadeem, M.; Hussain, S. Potential protective properties of flax lignan secoisolariciresinol diglucoside. Nutr. J. 2015, 14, 71. [Google Scholar] [CrossRef] [Green Version]
- Parikh, M.; Pierce, G.N. Dietary flaxseed: What we know and don’t know about its effects on cardiovascular disease. Can. J. Physiol. Pharmacol. 2019, 81, 75–81. [Google Scholar] [CrossRef]
- Kanikowska, D.; Korybalska, K.; Mickiewicz, A.; Rutkowski, R.; Kuchta, A.; Sato, M.; Kreft, E.; Fijałkowski, M.; Gruchała, M.; Jankowski, M.; et al. Flaxseed (Linum usitatissimum L.) supplementation in patients undergoing lipoprotein apheresis for severe hyperlipidemia—A pilot study. Nutrients 2020, 12, 1137. [Google Scholar] [CrossRef] [Green Version]
- Garros, L.; Drouet, S.; Corbin, C.; Decourtil, C.; Fidel, T.; De Lacour, J.L.; Leclerc, E.A.; Renouard, S.; Tungmunnithum, D.; Doussot, J.; et al. Insight into the influence of cultivar type, cultivation year, and site on the lignans and related phenolic profiles, and the health-promoting antioxidant potential of flax (Linum Usitatissimum L.) seeds. Molecules 2018, 23, 2636. [Google Scholar] [CrossRef] [Green Version]
- Křížová, L.; Dadáková, K.; Kašparovská, J.; Kašparovský, T. Isoflavones. Molecules 2019, 24, 1076. [Google Scholar] [CrossRef] [Green Version]
- Basu, P.; Maier, C. Phytoestrogens and breast cancer: In vitro anticancer activities of isoflavones, lignans, coumestans, stilbenes and their analogs and derivatives. Biomed. Pharmacother. 2018, 107, 1648–1666. [Google Scholar] [CrossRef]
- Balea, Ş.S.; Pârvu, A.E.; Pârvu, M.; Vlase, L.; Dehelean, C.A.; Pop, T.I. Antioxidant, Anti-Inflammatory and Antiproliferative Effects of the Vitis vinifera L. var. Fetească Neagră and Pinot Noir Pomace Extracts. Front. Pharmacol. 2020, 11, 990. [Google Scholar] [CrossRef]
- Toiu, A.; Mocan, A.; Vlase, L.; Pârvu, A.E.; Vodnar, D.C.; Gheldiu, A.M.; Moldovan, C.; Oniga, I. Phytochemical composition, antioxidant, antimicrobial and in vivo anti-inflammatory activity of traditionally used Romanian Ajuga laxmannii (Murray) Benth. (“nobleman’s beard”—barba împăratului). Front. Pharmacol. 2018, 9, 7. [Google Scholar] [CrossRef] [Green Version]
- Pop, R.M.; Puia, I.C.; Puia, A.; Chedea, V.S.; Leopold, N.; Bocsan, I.C.; Buzoianu, A.D. Characterization of Trametes versicolor: Medicinal Mushroom with Important Health Benefits. Not. Bot. Horti Agrobot. Cluj-Napoca 2018, 46, 343–349. [Google Scholar] [CrossRef] [Green Version]
- Fernandes, R.P.P.; Trindade, M.A.; Tonin, F.G.; Lima, C.G.; Pugine, S.M.P.; Munekata, P.E.S.; Lorenzo, J.M.; de Melo, M.P. Evaluation of antioxidant capacity of 13 plant extracts by three different methods: Cluster analyses applied for selection of the natural extracts with higher antioxidant capacity to replace synthetic antioxidant in lamb burgers. J. Food Sci. Technol. 2016, 53, 451–460. [Google Scholar] [CrossRef] [Green Version]
- Keser, S.; Celik, S.; Turkoglu, S.; Yilmaz, Ö.; Turkoglu, I. Hydrogen Peroxide Radical Scavenging and Total Antioxidant Activity of Hawthorn. Chem. J. 2012, 2, 912. [Google Scholar]
- Ho, E.; Karimi Galougahi, K.; Liu, C.C.; Bhindi, R.; Figtree, G.A. Biological markers of oxidative stress: Applications to cardiovascular research and practice. Redox Biol. 2013, 1, 483–491. [Google Scholar] [CrossRef] [Green Version]
- Taukoorah, U.; Mahomoodally, M.F. Crude Aloe vera Gel Shows Antioxidant Propensities and Inhibits Pancreatic Lipase and Glucose Movement in Vitro. Adv. Pharmacol. Sci. 2016, 2016, 3720850. [Google Scholar] [CrossRef] [Green Version]
- Ciortea, R.; Şuşman, S.; Măluţan, A.M.; Berceanu, C.; Mocan-Hognogi, R.F.; Bucuri, C.E.; Soriţău, O.; Neagoe, I.; Mihu, D. Mesenchymal stem cells derived from adipose tissue and Ishikawa cells co-culture highlight the role of adiponectin in endometrial cancer pathogenesis. Rom. J. Morphol. Embryol. 2018, 59, 1165–1172. [Google Scholar]
- Cenariu, D.; Fischer-Fodor, E.; Țigu, A.B.; Bunea, A.; Virág, P.; Perde-Schrepler, M.; Toma, V.A.; Mocan, A.; Berindan-Neagoe, I.; Pintea, A.; et al. Zeaxanthin-rich extract from superfood lycium barbarum selectively modulates the cellular adhesion and mapk signaling in melanoma versus normal skin cells in vitro. Molecules 2021, 26, 333. [Google Scholar] [CrossRef]
- Pallag, A.; Filip, G.A.; Olteanu, D.; Clichici, S.; Baldea, I.; Jurca, T.; Micle, O.; Vicas, L.; Marian, E.; Soriţău, O.; et al. Equisetum arvense L. extract induces antibacterial activity and modulates oxidative stress, inflammation, and apoptosis in endothelial vascular cells exposed to hyperosmotic stress. Oxid. Med. Cell. Longev. 2018, 2018, 3060525. [Google Scholar] [CrossRef] [Green Version]
- Alkhatiba, M.H.; Alyamania, S.A.; Abdu, F. Incorporation of methotrexate into coconut oil nanoemulsion potentiates its antiproliferation activity and attenuates its oxidative stress. DRUG Deliv. 2020, 27, 22–430. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.; Wan, M.; Xu, D.; Pan, D.; Xia, H.; Yang, L.; Sun, G. Flaxseed powder attenuates non-alcoholic steatohepatitis via modulation of gut microbiota and bile acid metabolism through gut–liver axis. Int. J. Mol. Sci. 2021, 22, 10858. [Google Scholar] [CrossRef]
- Parikh, M.; Maddaford, T.G.; Austria, J.A.; Aliani, M.; Netticadan, T.; Pierce, G.N. Dietary Flaxseed as a Strategy for Improving Human Health. Nutrients 2019, 11, 1171. [Google Scholar] [CrossRef] [Green Version]
- Erel, O. A novel automated direct measurement method for total antioxidant capacity using a new generation, more stable ABTS radical cation. Clin. Biochem. 2004, 37, 277–285. [Google Scholar] [CrossRef]
- Erel, O. A new automated colorimetric method for measuring total oxidant status. Clin. Biochem. 2005, 38, 1103–1111. [Google Scholar] [CrossRef]
- Harma, M.; Harma, M.; Erel, O. Increased oxidative stress in patients with hydatidiform mole. Swiss Med. Wkly. 2003, 133, 563–566. [Google Scholar]
- Erel, O.; Neselioglu, S. A novel and automated assay for thiol/disulphide homeostasis. Clin. Biochem. 2014, 47, 326–332. [Google Scholar] [CrossRef]
- Draper, H.H.; Squires, E.J.; Mahmoodi, H.; Wu, J.; Agarwal, S.; Hadley, M. A comparative evaluation of thiobarbituric acid methods for the determination of malondialdehyde in biological materials. Free Radic. Biol. Med. 1993, 15, 353–363. [Google Scholar] [CrossRef]
- Pfingstgraf, I.O.; Taulescu, M.; Pop, R.M.; Orăsan, R.; Vlase, L.; Uifalean, A.; Todea, D.; Alexescu, T.; Toma, C.; Pârvu, A.E. Protective effects of taraxacum officinale L. (dandelion) root extract in experimental acute on chronic liver failure. Antioxidants 2021, 10, 504. [Google Scholar] [CrossRef]
- Sharbidre, A.; Kasote, D. Synthesis of Silver Nanoparticles Using Flaxseed Hydroalcoholic Extract and its Antimicrobial Activity. Curr. Biotechnol. 2013, 2, 162–166. [Google Scholar] [CrossRef]
- Minevich, I.E.; Osipova, L.L.; Nechiporenko, A.P.; Melnikova, M.I.; Tsyganova, T.B. IR-spectroscopy of polysaccharide flaxseed (Linum usitatissimum L.) products. Foods Raw Mater. 2019, 7, 274–282. [Google Scholar] [CrossRef]
- Hong, T.; Yin, J.-Y.; Nie, S.-P.; Xie, M.-Y. Applications of infrared spectroscopy in polysaccharide structural analysis: Progress, challenge and perspective. Food Chem. X 2021, 12, 100168. [Google Scholar] [CrossRef]
- Okur, İ.; Baltacıoğlu, C.; Ağçam, E.; Baltacıoğlu, H.; Alpas, H. Evaluation of the Effect of Different Extraction Techniques on Sour Cherry Pomace Phenolic Content and Antioxidant Activity and Determination of Phenolic Compounds by FTIR and HPLC. Waste Biomass Valoriz. 2019, 10, 3545–3555. [Google Scholar] [CrossRef]
- Colalto, C. What phytotherapy needs: Evidence-based guidelines for better clinical practice. Phyther. Res. 2018, 32, 413–425. [Google Scholar] [CrossRef]
- Efferth, T.; Saeed, M.E.M.; Mirghani, E.; Alim, A. Integration of phytochemicals and phytotherapy into cancer precision medicine. Oncotarget 2017, 8, 50284–50304. [Google Scholar] [CrossRef] [Green Version]
- Kwesi, C.; Dzuvor, O.; Taylor, J.T.; Acquah, C.; Pan, S.; Agyei, D. Bioprocessing of Functional Ingredients from Flaxseed. Molecules 2018, 23, 2444. [Google Scholar] [CrossRef] [Green Version]
- Dobrowolska, K.; Regulska-ilow, B. Review Article the Legitimacy of Using Dietary Supplement Diglycoside Secoisolariciresinol (SDG) from Flaxseed in Cancer. Rocz. Panstw. Zakl. Hig. 2021, 72, 9–20. [Google Scholar] [CrossRef]
- Brodowska, K.; Catthoor, R.; Brodowska, A.J.; Symonowicz, M.; Ska, E.B.L. A comparison of antioxidant properties of extracts from defatted and non-defatted flax (Linum usitatissimum) seeds. Albanian J. Agric. Sci. 2014, 13, 16–23. [Google Scholar]
- Oomah, B.D.; Mazza, G.; Kenaschuk, E.O. Flavonoid content of flaxseed. Influence of cultivar and environment. Euphytica 1996, 90, 163–167. [Google Scholar] [CrossRef]
- Hsieh, C.-J.; Hsu, Y.-L.; Huang, Y.-F.; Tsai, E.-M. Molecular Mechanisms of Anticancer Effects of Phytoestrogens in Breast Cancer. Curr. Protein Pept. Sci. 2018, 19, 323–332. [Google Scholar] [CrossRef]
- Senthilkumar, H.A.; Fata, J.E.; Kennelly, E.J. Phytoestrogens: The current state of research emphasizing breast pathophysiology. Phyther. Res. 2018, 32, 1707–1719. [Google Scholar] [CrossRef]
- Viggiani, M.T.; Polimeno, L.; Di Leo, A.; Barone, M. Phytoestrogens: Dietary intake, bioavailability, and protective mechanisms against colorectal neoproliferative lesions. Nutrients 2019, 11, 1709. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, C.; Cagide, F.; Teixeira, J.; Amorim, R.; Sequeira, L.; Mesiti, F.; Silva, T.; Garrido, J.; Remião, F.; Vilar, S.; et al. Hydroxybenzoic acid derivatives as dual-target ligands: Mitochondriotropic antioxidants and cholinesterase inhibitors. Front. Chem. 2018, 6, 126. [Google Scholar] [CrossRef] [Green Version]
- Prudencio, A.; Faig, J.J.; Song, M.J.; Uhrich, K.E. Phenolic Acid-based Poly(anhydride-esters) as Antioxidant Biomaterials. Macromol. Biosci. 2016, 16, 214–222. [Google Scholar] [CrossRef] [Green Version]
- Verma, S.; Singh, A.; Mishra, A. Gallic acid: Molecular rival of cancer. Environ. Toxicol. Pharmacol. 2013, 35, 473–485. [Google Scholar] [CrossRef]
- Ekeuku, S.O.; Pang, K.L.; Chin, K.Y. Effects of caffeic acid and its derivatives on bone: A systematic review. Drug Des. Devel. Ther. 2021, 15, 259–275. [Google Scholar] [CrossRef]
- Zielińska, D.; Zieliński, H.; Laparra-Llopis, J.M.; Szawara-Nowak, D.; Honke, J.; Giménez-Bastida, J.A. Caffeic acid modulates processes associated with intestinal inflammation. Nutrients 2021, 13, 554. [Google Scholar] [CrossRef]
- Chen, C. Sinapic acid and its derivatives as medicine in oxidative stress-induced diseases and aging. Oxid. Med. Cell. Longev. 2016, 2016, 3571614. [Google Scholar] [CrossRef] [Green Version]
- Pandi, A.; Kalappan, V.M. Pharmacological and therapeutic applications of Sinapic acid—An updated review. Mol. Biol. Rep. 2021, 48, 3733–3745. [Google Scholar] [CrossRef]
- Zduńska, K.; Dana, A.; Kolodziejczak, A.; Rotsztejn, H. Antioxidant properties of ferulic acid and its possible application. Skin Pharmacol. Physiol. 2018, 31, 332–336. [Google Scholar] [CrossRef]
- Chaudhary, A.; Jaswal, V.S.; Choudhary, S.; Sonika; Sharma, A.; Beniwal, V.; Tuli, H.S.; Sharma, S. Ferulic Acid: A Promising Therapeutic Phytochemical and Recent Patents Advances. Recent Pat. Inflamm. Allergy Drug Discov. 2019, 13, 115–123. [Google Scholar] [CrossRef]
- Eilati, E.; Hales, K.; Zhuge, Y.; Ansenberger Fricano, K.; Yu, R.; van Breemen, R.B.; Hales, D.B. Flaxseed enriched diet-mediated reduction in ovarian cancer severity is correlated to the reduction of prostaglandin E2 in laying hen ovaries. Prostaglandins Leukot. Essent. Fat. Acids 2013, 89, 179–187. [Google Scholar] [CrossRef] [Green Version]
- Hasan, I.; Rahman, S.N.; Islam, M.M.; Ghosh, S.K.; Mamun, M.R.; Uddin, M.B.; Shaha, R.K.; Kabir, S.R. A N-acetyl-D-galactosamine-binding lectin from Amaranthus gangeticus seeds inhibits biofilm formation and Ehrlich ascites carcinoma cell growth in vivo in mice. Int. J. Biol. Macromol. 2021, 181, 928–936. [Google Scholar] [CrossRef]
- Lisboa, T.; Silva, D.; Duarte, S.; Ferreira, R.; Andrade, C.; Lopes, A.L.; Ribeiro, J.; Farias, D.; Moura, R.; Reis, M.; et al. Toxicity and Antitumor Activity of a Thiophene‐Acridine Hybrid. Molecules 2020, 25, 64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kar, B.; Kumar, R.B.S.; Bala, A.; Dolai, N.; Mazumder, U.K.; Haldar, P.K. Evaluation of antitumor activity of mimusops elengi leaves on Ehrlich’s ascites carcinoma-treated mice. J. Diet. Suppl. 2012, 9, 166–177. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Baby, D.; Rajguru, J.P.; Patil, P.B.; Thakkannavar, S.S.; Pujari, V.B. Inflammation and cancer. Ann. Afr. Med. 2019, 18, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Mansour, M.A.; Ibrahim, W.M.; Salama, M.M.; Salama, A.F. Dual inhibition of glycolysis and autophagy as a therapeutic strategy in the treatment of Ehrlich ascites carcinoma. J. Biochem. Mol. Toxicol. 2020, 34, e224. [Google Scholar] [CrossRef] [PubMed]
- Sharma, J.N.; Al-Omran, A.; Parvathy, S.S. Role of nitric oxide in inflammatory diseases. Inflammopharmacology 2007, 15, 252–259. [Google Scholar] [CrossRef] [PubMed]
- Boora, F.; Chirisa, E.; Mukanganyama, S. Evaluation of Nitrite Radical Scavenging Properties of Selected Zimbabwean Plant Extracts and Their Phytoconstituents. J. Food Process. 2014, 2014, 918018. [Google Scholar] [CrossRef] [Green Version]
- Omari, N.E.; Menyiy, N.E.; Zengin, G.; Goh, B.H.; Gallo, M.; Montesano, D.; Naviglio, D.; Bouyahya, A. Anticancer and anti-inflammatory effects of tomentosin: Cellular and molecular mechanisms. Separations 2021, 8, 207. [Google Scholar] [CrossRef]
- Huang, W.; Hou, S. Decreased Human Platelet Activation and Mouse Pulmonary Thrombosis by Rutaecarpine and Comparison of the Relative Effectiveness with BAY11-7082: Crucial Signals of p38-NF- κB. Molecules 2021, 27, 476. [Google Scholar] [CrossRef]
- Arulselvan, P.; Fard, M.T.; Tan, W.S.; Gothai, S.; Fakurazi, S.; Norhaizan, M.E.; Kumar, S.S. Role of Antioxidants and Natural Products in Inflammation. Oxid. Med. Cell. Longev. 2016, 2016, 5276130. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No | Retention Time Rt (min) | UV λmax (nm) | [M+H]+ (m/z) | Tentative Identification | Subclass | Quantity * μg/mL |
|---|---|---|---|---|---|---|
| 1 | 3.37 | 270 | 138 | Hydroxybenzoic acid | Hydroxybenzoic acid | 107.629 |
| 2 | 4.75 | 280 | 171 | Gallic acid | Hydroxybenzoic acid | 18.219 |
| 3 | 10.43 | 280 | 199 | Syringic acid | Hydroxybenzoic acid | 37.432 |
| 4 | 12.87 | 320 | 181 | Caffeic acid | Hydroxycinnamic acid | 13.374 |
| 5 | 14.13 | 280 | 687 | Secoisolariciresinol- diglucoside (SDG) | Lignan | 5.193 |
| 6 | 16.33 | 320 | 225 | Sinapic acid | Hydroxycinnamic acid | 6.543 |
| 7 | 17.16 | 321 | 195 | Ferulic acid | Hydroxycinnamic acid | 5.426 |
| 8 | 19.72 | 280 | 360 | Secoisolariciresinol (SECO) | Lignan | 210.796 |
| 9 | 22.89 | 280 | 358 | Pinoresinol | Lignan | 10.772 |
| 10 | 23.83 | 280 | 358 | Matairesinol | Lignan | 1.569 |
| Sample | DPPH | FRAP | H2O2 | NO |
|---|---|---|---|---|
| FLAX IC50(µg TE/g d.w. plant material) | 39.07 ± 2.84 | 64.57 ± 12.09 | 14.33 ± 3.86 | 57.72 ± 3.58 |
| TROLOX IC50(µg /mL) | 11.18 ± 1.22 | 10.21 ± 0.99 | 10.77 ± 2.43 | 86.82 ± 12.49 |
| p | 0.01 | 0.001 | 0.05 | 0.001 |
| Samples | BW Gain (%) | Ascites (mL) | Viable Cell (%) | Non-Viable Cell (%) |
|---|---|---|---|---|
| CONTROL | 11.02 ± 1.57 | - | - | - |
| EAC | 32.55 ± 3.29 ### | 9.13 ± 1.35 | 93.68 ± 2.97 | 6.32 ± 0.94 |
| EAC-FLAX | 19.57 ± 4.77 ### | 7.40 ± 1.83 * | 85.16 ± 3.70 * | 14.84 ± 1.08 *** |
| Samples | TAC (mmol TROLOX Equiv./L) | TOS (µM H2O2 Equiv./L) | OSI | SH (mM GSH/L) | MDA (nM/L) |
|---|---|---|---|---|---|
| CONTROL | 1.093 ± 0.002 | 4.20 ± 0.90 | 3.84 ± 0.63 | 351.00 ± 59.10 | 2.64 ± 0.31 |
| EAC | 1.088 ± 0.001 # | 7.22 ± 1.42 ## | 6.73 ± 0.38 ## | 360.78 ± 38.35 ## | 5.41 ± 0.26 ## |
| EAC-FLAX | 1.089 ± 0.001 | 5.77 ± 0.86 * | 5.29 ± 0.16 | 563.25 ± 103.22 *** | 3.87 ± 0.40 *** |
| Samples | TAC (mmol Trolox Equiv/L) | TOS (µM H2O2 Equiv/L) | OSI | SH (mM GSH/L) | MDA (nM/L) |
|---|---|---|---|---|---|
| CONTROL | 1.099 ± 0.001 | 12.39 ± 1.22 | 11.42 ± 1.52 | 410.00 ± 38.09 | 2.37 ± 0.21 |
| EAC | 1.084 ± 0.001 ## | 18.90 ± 4.16 ## | 17.33 ± 3.52 # | 201.67 ± 52.99 ## | 3.36 ± 0.36 ## |
| EAC-FLAX | 1.098 ± 0.003 | 16.70 ± 2.01 * | 15.21 ± 2.18 | 306.50 ± 59.35 ** | 3.18 ± 0.23 * |
| Samples | NOx (ng/mL) | 3NT (ng/mL) | NF-kB (ng/mL) |
|---|---|---|---|
| CONTROL | 26.42 ± 3.54 | 19.84 ± 3.89 | 1.9 ± 0.15 |
| EAC | 39.54 ± 5.21 ## | 41.05 ± 10.09 ### | 3.70 ± 0.67 ### |
| EAC-FLAX | 31.02 ± 3.38 ** | 22.16 ± 9.87 ** | 1.98 ± 0.09 ** |
| Samples | NOx (ng/mL) | 3NT (ng/mL) | NF-kB (ng/mL) |
|---|---|---|---|
| CONTROL | 7.54 ± 0.04 | 18.18 ± 2.72 | 2.15 ± 0.17 |
| EAC | 17.20 ± 5.52 ### | 52.16 ± 8.71 ### | 5.44 ± 0.12 ## |
| EAC-FLAX | 9.63 ± 2.85 ** | 30.53 ± 7.09 ** | 4.03 ± 0.39 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chera, E.I.; Pop, R.M.; Pârvu, M.; Sorițău, O.; Uifălean, A.; Cătoi, F.A.; Cecan, A.; Negoescu, A.G.; Achimaș-Cadariu, P.; Pârvu, A.E. Flaxseed Ethanol Extracts’ Antitumor, Antioxidant, and Anti-Inflammatory Potential. Antioxidants 2022, 11, 892. https://doi.org/10.3390/antiox11050892
Chera EI, Pop RM, Pârvu M, Sorițău O, Uifălean A, Cătoi FA, Cecan A, Negoescu AG, Achimaș-Cadariu P, Pârvu AE. Flaxseed Ethanol Extracts’ Antitumor, Antioxidant, and Anti-Inflammatory Potential. Antioxidants. 2022; 11(5):892. https://doi.org/10.3390/antiox11050892
Chicago/Turabian StyleChera, Elisabeta Ioana, Raluca Maria Pop, Marcel Pârvu, Olga Sorițău, Ana Uifălean, Florinela Adriana Cătoi, Andra Cecan, Andrada Gabriela Negoescu, Patriciu Achimaș-Cadariu, and Alina Elena Pârvu. 2022. "Flaxseed Ethanol Extracts’ Antitumor, Antioxidant, and Anti-Inflammatory Potential" Antioxidants 11, no. 5: 892. https://doi.org/10.3390/antiox11050892