
Generation and Physiology of Hydrogen Sulfide and Reactive Sulfur Species in Bacteria

Institute of Microbiology and College of Life Sciences, Zhejiang University, Hangzhou 310058, China
*
Author to whom correspondence should be addressed.
Antioxidants 2022, 11(12), 2487; https://doi.org/10.3390/antiox11122487
Submission received: 4 November 2022
/
Revised: 14 December 2022
/
Accepted: 15 December 2022
/
Published: 17 December 2022
(This article belongs to the Special Issue Reactive Sulfur Species in Microorganisms)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Sulfur is not only one of the most abundant elements on the Earth, but it is also essential to all living organisms. As life likely began and evolved in a hydrogen sulfide (H2S)-rich environment, sulfur metabolism represents an early form of energy generation via various reactions in prokaryotes and has driven the sulfur biogeochemical cycle since. It has long been known that H2S is toxic to cells at high concentrations, but now this gaseous molecule, at the physiological level, is recognized as a signaling molecule and a regulator of critical biological processes. Recently, many metabolites of H2S, collectively called reactive sulfur species (RSS), have been gradually appreciated as having similar or divergent regulatory roles compared with H2S in living organisms, especially mammals. In prokaryotes, even in bacteria, investigations into generation and physiology of RSS remain preliminary and an understanding of the relevant biological processes is still in its infancy. Despite this, recent and exciting advances in the fields are many. Here, we discuss abiotic and biotic generation of H2S/RSS, sulfur-transforming enzymes and their functioning mechanisms, and their physiological roles as well as the sensing and regulation of H2S/RSS.
1. Introduction
Sulfur, one of the most abundant elements on our planet, and an element that has six valence electrons, is essential to all living organisms. The term ‘reactive sulfur species (RSS)′, coined by Jacob and colleagues [1], has been in the general scientific vocabulary for more than two decades. Apparently, RSS, much like reactive oxygen species (ROS) and reactive nitrogen species (RNS), comprise chemically reactive molecules containing the element for which they are named, sulfur, oxygen, and nitrogen respectively [2]. Because of their highly reactive nature, the reactive species are prone to interact with cellular macromolecules, including nucleic acids, proteins, and lipids [3]. Not surprisingly, most, if not all, of these reactive species, all of which can be generated endogenously in the cell and arise abiotically in environments, are hostile to living cells at supraphysiological levels [2]. Despite this, in eukaryotes, especially mammals, a large majority of studies in fact center on the beneficial roles that these species play by mediating redox signaling and redox regulation [4]. It has been firmly established that these reactive species, as a signaling molecules, impact diverse processes including metabolism, vasodilation, neurotransmission, immunity, apoptosis, and cancer [2,5,6].
Compared to ROS and RNS, which have been extensively studied for decades, RSS are relatively new. The concept of RSS has evolved over time and is still far from unanimously held. The ‘narrow’ view defines RSS as the molecules produced from sequential one-electron oxidations of hydrogen sulfide (H2S), forming a thiyl (sulfhydryl) radical (HS•, the counterpart of HO• in ROS, the same below), hydrogen persulfide (H2S2, H2O2), and the supersulfide radical (S2•, O2•) [7] (Figure 1). In the ‘broad’ view, RSS refer collectively to reactive sulfur chemotypes (both organic and inorganic) that can react with, oxidize or reduce other molecules under physiological conditions [8,9] (Figure 1). Therefore, ‘reactive’ sulfur atoms can be found in compounds having sulfur of higher valence, such as sulfite (SO32−, +4) and sulfate (SO42−, +6), which have interchalcogen bonds, and thiosulfate (S2O32−, −1/+5 or 0/+4), in which sulfur atoms are catenated as in polysulfides (Figure 1). To date, the widely accepted notion about RSS, which we defined here as the ‘normal’ view, regards RSS as any molecules carrying reactive sulfur with a valence of −2, −1, or 0 [9,10]. In this sense, representative RSS include protein thiols (PSH) and persulfide (PSSH), low-molecule-weight (LMW) thiols (RSH) and persulfide (RSSH), hydrogen persulfide/polysulfide (H2Sn, n ≥ 2), polysulfides (RSS (n) R, n > 1), and sulfenic acids (RSOH) [10,11] (Figure 1).
Formation and metabolism of RSSs have been intensively studied in eukaryotes, especially mammals [10,12]. In these processes, H2S is the center molecule that is generated in the cytosol through degradation of thiol-containing molecules, cysteine (cys) in particular [9] (Figure 2A). Persulfide and polysulfide can then be formed from H2S through various non-enzymatic reactions involving ROS. While bacteria preserve these pathways to generate H2S, many of them can also do so through reduction of a variety of inorganic and organic compounds containing sulfur of high higher valence, releasing H2S as well as other RSS as the terminal products or intermediates [13,14]. In parallel, H2S can be oxidized by diverse prokaryotes, and even mitochondria, through a combination of abiotic and biotic reactions to diverse sulfur compounds of any possible valence, many of which are RSS [8,9]. As inorganic sulfur in the form of sulfate (SO42−) is extremely abundant in oceans, and metabolites in sulfur cycling are diverse, RSSs generated by microbes have profound impacts on all life and beyond [15].
Despite the difference in their functioning roles, the mechanism for biological activity of the reactive species is similar: modification of cellular targets. It is generally accepted that the reactive species interact with their specific cellular targets for redox signaling at physiological levels, whereas at elevated concentrations they modify macromolecules in a rather indiscriminate way, with interactions amounting to damaged enzymes and DNA, and even cell death [4]. This also holds true for RSS, although sulfur has been usually considered as the essential element constituting cellular antioxidant systems [16].
In recent years, a number of studies combining conventional genetic analysis and cutting-edge technologies have stressed the profound biological roles of RSS in mammalian as well as bacterial cells. While an understanding of the sensing, production, and physiological role of RSS in prokaryotes, bacteria in particular, is still in its infancy, it is clear that control of RSS homeostasis emerges as a promising means for therapeutic treatments for sulfur-related diseases, for maintenance of a balanced sulfur biogeochemical cycle, and for development of RSS-dependent biotechnologies. In this review, we take the notion of RSS defined by the ‘normal’ view, with an emphasis on small-molecule RSS. We discuss the origin of RSS and bacterial enzymes transforming sulfur compounds associated with RSS, as well as the functioning mechanisms of RSS and their biological impacts, and we highlight recent progress in areas of RSS sensing, which expands the understanding of RSS biology in bacteria.
2. Influence of Sulfur on the Origin of Life
Redox reactions, a result of electron imbalance mediated by energy, are the basis of all physiological activities in cells, such as respiration and photosynthesis. The original life began in an anoxic ferrous ocean 3.8 billion years ago (bya), likely as anoxygenic chemolithotrophs in environments associated with deep-sea hydrothermal vent systems [7,17,18]. As these environments were characterized by large amounts of sulfur and metals, iron in particular, it is believed that iron-sulfur clusters were the first electron-transfer units to be produced during chemical evolution [19]. In addition, sulfur now has been recognized to be a crucial element in the physiology of life’s common ancestor because some sulfur species, such as acetyl-coenzyme A, are essential intermediates in metabolism [20,21]. Moreover, H2S and diverse RSS, such as elemental sulfur, sulfite, thiosulfate, and polysulfides, rather than sulfate, which is the predominant inorganic sulfur compound on the current earth, were more prevalent in the ancient ocean and have played an critical role in shaping the sulfur cycle and life [22,23,24].
A large portion of contemporary living organisms, eukaryotes in particular, use O2 as the terminal acceptor for energy production. During oxygen respiration, ROS are generated endogenously in the cell. It has been postulated that antioxidant mechanisms, including superoxide dismutase, catalase, and peroxide reductase, evolved concomitantly with oxygen respiration [25]. However, the history of RSS is undoubtedly much longer than that of ROS, and more importantly, antioxidant mechanisms were present far earlier than the oxic ocean [26]. The anoxygenic photosynthesis appeared ~3–3.5 bya, several million years before the cyanobacterial existence, according to fossil evidence [27,28]. In the early photosynthetic organisms, their light-gathering antennae and processing machinery were not sophisticated enough to support oxidation of H2O, and instead, H2S was more likely to act as the electron donor [7,29,30]. From 2.3 billion years on, in the ‘great oxidation event (GOE)’, cyanobacteria had evolved ability to oxidize H2O and produced a large amount of oxygen via photosynthesis [31]. During that time, the atmospheric O2 fluctuated between 0%–2% for 1.7 billion years, while the oceans remained anoxic [17,29,30,32,33].
While the first eukaryotes appeared approximately 1.5 bya, it was not until 0.6 bya that the ocean finally became aerobic [25,34]. This time, called the ‘Boring Billion’, represents a period of geobiological stasis caused by prolonged nutrient, climate, atmospheric and tectonic stability [35]. Phylogenetic studies have shown that antioxidant enzymes were present in the last universal common ancestor, well before the emergence of cyanobacteria and the GOE [36]. These enzymes are present in all domains of life, even in anaerobic bacteria, suggesting that these enzymes are ancient and that the need for ROS protection is pervasive in non-aerobic environments [26,36,37,38,39,40,41,42]. Because of the chemical similarity between O and S, and the less electron-negativity and greater reactivity versatility of S, these enzymes have been proposed to be primarily used to resist RSS. In recent years, evidence supporting this proposal has emerged [26,43,44,45,46]. All of these insights have provided the new interpretation of RSS in biological evolution.
3. Transformation of Sulfur Compounds Associated with Amino Acid Metabolism in Bacteria
Prokaryotes, especially bacteria, are renowned for metabolic diversity, which endows these organisms with unparalleled capacity in the biotransformation of sulfur compounds [47]. Through microbial metabolic processes that transform the oxidation state of sulfur, a variety of inorganic and organic sulfur compounds are generated. H2S, the predominant inorganic sulfur compound in earth’s earlier days that serves as the foundation for other RSS, is well-known for driving photosynthesis and energy metabolism in sulfide-oxidizing and sulfate- or sulfite-reducing microorganisms [48,49,50]. Bacterial H2S production has been perceived as a metabolic side product, but recent studies have demonstrated that H2S is an important signaling molecule and can protect bacteria from antibiotic and oxidative stress [51,52]. In bacteria, H2S can be generated endogenously either through amino acid metabolism or reduction of compounds having sulfur of higher valence.
3.1. S2− Biogenesis through Amino Acid Metabolism
There are at least three enzymes playing roles in bacterial H2S endogenous synthesis by utilizing homologues of the mammalian sulfur metabolic network. Two enzymes, cystathionine β-synthase (CBS) and cystathionine γ-lyase (CSE), are involved in the transsulfuration pathway [53,54,55]. The third enzyme, mercaptopyruvate sulfurtransferase (MST), converts 3-mercaptopyruvate (3-MP), derived from the reaction of cys with keto acids catalyzed by cys aminotransferase (CAT), to pyruvate and persulfide for the release of H2S [56] (Figure 2).
CBS, located at the intersection of two amino acid metabolic pathways, the methionine cycle and transsulfuration pathway, is involved in the redirection of homocys from the methionine cycle toward glutathione (GSH) synthesis, finally influencing H2S biogenesis [53,54]. When it comes to the roles in the transsulfuration pathway, CBS catalyzes the β-replacement of L-serine or L-homocys, forming cystathionine and H2O [53,54,57,58]. It can also catalyze β-replacement of L-cys by H2O, by L-homocys, or by L-cys forming L-cystathionine, L-lanthionine, or L-serine, respectively, while releasing H2S in the process. As an enzyme displaying multiple functions in amino acid metabolism, CBS activity can be regulated in distinct ways, including heme-dependent allosteric regulation, adoMet-dependent allosteric regulation, and posttranslational covalent modifications [59]. Like CBS, CSE displays abundant substrate promiscuity and can catalyze the γ-elimination of cystathionine to give L-cys, α-ketobutyrate, and NH3·H2O, generating H2S [60] (Figure 2). In addition, CSE also produces pyruvate, Cys-SSH, homocys persulfide, etc. from L-cys and L-homocys [61,62]. Compared to CBS/CSE-dependent H2S synthesis, our knowledge of CAT/MST-dependent H2S biogenesis in bacteria is limited. It is known that transformation of 3-MP to pyruvate and H2S requires an intermediate of protein persulfide, E-SSH, which acts as a source of H2S under reducing conditions [56,63].
Generally, bacteria can encode either CBS/CSE or MST, but exceptions that carry all of these enzymes are increasingly found, such as γ-proteobacterium Shewanella oneidensis [64]. In addition to CBS/CSE/MST, several enzymes that contribute to H2S biogenesis have been identified. For example, Fusobacterium nucleatum has been identified to have four enzymes involved in H2S production, L-cys desulfhydrase, cys synthase and L-methionine γ-lyase, respectively [65,66,67,68]. In Escherichia coli, L-cys desulfhydrases and cys desulfurases also contribute to H2S production [69,70].
3.2. S2− Biogenesis through Assimilatory Reduction of Inorganic Sulfur Species
In addition to generation via amino acid metabolism, H2S can be produced in vast amounts through reduction of compounds having sulfur of higher valence. Here we discuss the reductive pathways that release H2S in the cytoplasm, which can be incorporated during biosynthesis of cys. Sulfate (SO42−), the most stable form of sulfur under current oxic conditions, is the largest sulfur pool on the Earth [71,72]. On this account, bacteria that reduce SO42− to HS− are no doubt the most important sulfur transformers for the biogeochemical sulfur cycle (Figure 2). In parallel, L-cys synthesis from inorganic sulfate is also the major mechanism for sulfur conversion into amino acids or organic compounds, and eventually serves as a structural block for proteins [73]. The first step of sulfate reduction, which occurs in the cytoplasm and is the same for all living bacteria having this capability, converts sulfate to sulfite with adenosine-5′-phosphosulfate-reductase (APS reductase) [74]. The steps followed can be either the assimilatory sulfite reduction or the dissimilatory sulfite reduction [75]. The latter occurs in sulfate-reducing bacteria (SRB), which are composed of morphologically and ecologically diverse but physiologically unified microbes, and which will be discussed later in detail. After SO42− is transported into the cytoplasm, it is activated by the ATP sulfurylase to APS, which is subsequently reduced to SO32− by the APS reductase. The reduction of sulfite to HS− through the assimilatory pathway, which was first uncovered in E. coli about half a decade ago, is catalyzed by the CysIJ complex [73]. The reduction costs three NADPH, generating HS−, which converges with O-acetyl-l-serine (OAS) on O-acetylserine sulfhydrylase A (CysK), a pyridoxal 5′-phosphate-dependent enzyme that catalyzes the final reaction of cys biosynthesis in bacteria [74,76].
In addition to sulfite, reduction of thiosulfate could also occur through both assimilatory and dissimilatory pathways. For assimilation, thiosulfate is up-taken into the cytoplasm of E. coli cells largely via the ATP-dependent transporter CysUWA, which also functions as a sulfate importer [77] (Figure 2). In addition, YeeE has recently been identified as a thiosulfate importer in E. coli [78]. In the cytoplasm, CysM catalyzes the reaction of thiosulfate with O-acetyl-L-serine (OAS) to form S-sulfo-L-cys, which is subsequently converted to cys upon reduction of the disulfide bond by glutaredoxin-like protein NrdH or glutaredoxin Grx1 [79,80]. Recently, a CysM-independent pathway for thiosulfate assimilation was identified, but its contribution is rather limited [81].
4. Dissimilatory Transformation of Inorganic Sulfur Compounds in Bacteria
Bacteria are the predominant force driving the biogeochemical sulfur cycle, especially in oceans [47]. Through microbial metabolic processes that transform the oxidation state of sulfur, a variety of inorganic and organic sulfur compounds are generated. Although organic sulfur has profound impacts on living organisms and ecology, it plays only a minor role in the biogeochemical sulfur cycle [74,82]. As a large portion of intermediates formed in the sulfur cycle are RSS, it is conceivable that they are collectively indispensable for the biogeochemical sulfur cycle. In addition, many bacteria can harness energy released from the redox reactions constituting the biogeochemical sulfur cycle to support growth in environments.
4.1. Reduction of S6+(SO42−)/S4+(SO32−)
In SRB, after the reduction of SO42− to SO32− completes in the cytoplasm, the dissimilatory sulfite reductase (Dsr) catalyzes the subsequent reaction that generates S2O32−, and releases HS− into the surroundings [75] (Figure 2). Intriguingly, although SRB are ubiquitous and Dsr has been used as the functional marker for many years, the mechanistic enigma of Dsr enzymes has been unveiled only recently [72,83,84]. For many years, Dsr enzymes have been believed to be composed of two proteins, DsrA and DsrB, in the form of hetero-tetramer [85]. Each A/B heterodimer harbors 2 siro-hemes and 2 [4Fe–4S] clusters that are presumably involved in electron transfer to HSO3− (Figure 3A). Now it is clear that DsrA and DsrB are the minimum requirement for sulfite reduction, and additional and liable subunits, DsrC in particular, are also involved in the process [85]. The current understanding of the mechanism is that the DsrAB complex catalyzes the reduction reaction, resulting in the production of S2O32− and formation of a persulfide bond with a cys residue on DsrC [85]. Subsequently, through a yet-unknown mechanism, DsrC expunges the bound sulfur as HS− and becomes oxidized, which subsequently returns back to the reduced form by accepting electrons from the cytoplasmic membrane for the next reaction [76]. In this way, DsrC couples the reduction reaction with energy conservation (Figure 2). This model also suggests more accessary proteins to be implicated in the sulfite reduction, at least a quinone dehydrogenase or equivalent at the cytoplasmic membrane that reduces the oxidized DsrC. It has been proposed that the multiple-component DsrMKJOP complex likely plays this role [72,74,86,87] (Figure 2).
Besides SRB, various bacteria unable to respire sulfate can still use extracellular sulfite as an electron acceptor to support growth. To date, two types of non-Dsr dissimilatory sulfite reductases that are not homologous to DsrAB have been identified and characterized, differing from each other in cofactors used for catalysis [72]. While the reductases from Salmonella enterica and Methanocaldococcus jannaschii are siroheme-dependent in the manner of DsrAB, species in Campylobacter, Sulfurospirillum, Wolinella, and Shewanella have evolved cytochrome (cyt) c-based sulfite-reducing enzyme [88,89].
4.2. Reduction of S2+(S4O62−)
It has been known for nearly a century that some bacteria, such as S. enterica, could grow by respiring tetrathionate as an electron acceptor [92,93,94] (Figure 4). The product of 2-electron reduction of tetrathionate is thiosulfate [72]. To date, some distinct types of tetrathionate reductases have been described, including Ttr of S. enterica and TsdA of C. jejuni [72,93] (Figure 3B,C). Ttr is composed of three subunits: TtrA contains a molybdopterin guanine dinucleotide cofactor and a [4Fe–4S] cluster, TtrB is associated with four [4Fe–4S] clusters, and TtrC is an integral membrane protein functioning to oxidize quinol (Figure 4). In addition, TsdA has been identified as a bidirectional enzyme that catalyzes the interconversion of tetrathionate and thiosulfate [14,95,96,97] (Figure 3B,C). For oxidation of thiosulfate, the enzyme is called thiosulfate dehydrogenase (Tsd), which will be discussed later in thiosulfate oxidation.
Although tetrathionate reduction contributes to the sulfur cycle less significantly than does sulfate/sulfite reduction, it is prevalent in the gut and is important for S. enterica during infection [94]. In the mammalian host, tetrathionate is formed in a large amount from oxidation of thiosulfate by ROS, which is generated during the oxidative burst upon intestinal inflammation. The resulting tetrathionate provides S. enterica with a selective growth advantage in the gut over the commensal microbiota lacking this capacity [94]. Interestingly, it has also been reported that the application of a relatively simple tetrathionate salt-based molecule exerts a protective effect against ischemia-reperfusion ROS-derived injuries in mammals [98].
4.3. Reduction of S0
Reduction of sulfur to HS− is carried out by sulfur reductase in a respiratory type of metabolism [99]. As zero-valent sulfur (ZVS), in addition to elemental sulfur, can be found in various sulfur compounds, such as thiosulfate and polysulfides, substrates of sulfur reductases are diverse. Notably, there are two definitions for valence of two sulfur atoms of thiosulfate: the broadly accepted version of 0/+4, and as appearing in some publications as −1/+5 [8,100].
Dissimilatory sulfur reductases are widespread in prokaryotes, but they are distinct between bacteria and archaea [99,101,102] (Figure 4). Bacterial sulfur/thiosulfate reductases, such as PhsABC of S. enterica and PsrABC of Wolinella succinogenes, are composed of three subunits, a catalytic unit (A), an iron–sulfur protein (B), and an integral membrane protein (C) that serves to anchor the other subunits on the membrane [103,104]. Despite this, these enzymes may not be identical, as PhsABC catalyzes reduction of elemental sulfur and the zero-valent sulfur of thiosulfate to S2−, while PsrABC is capable of reducing polysulfides (Sn2−, n > 2) to Sn-12− and HS− in a stepwise mode [103,105].
4.4. Oxidation of S2− and S0
Sulfide generated from sulfate reduction can be converted to more oxidized sulfur species, either biotically or abiotically, including sulfur, thiosulfate, sulfite, and sulfate, with sulfate making up the vast majority (>90%) [72,106] (Figure 2 and Figure 4). Like reduction, oxidation of sulfide in living cells can be coupled to energy conservation by supplying electrons to the respiratory electron transport chain [107]. Oxidation of sulfide to sulfane sulfur is the most important process for formation of RSS, such as hydrogen persulfide/polysulfide and organic persulfide/polysulfide. Sulfane sulfur is a common cellular component, maintained in a micromolar range, and changing with growth stages in bacteria [108,109]. Most cellular persulfides and polysulfides are generated from sulfide (S2−) oxidation by SQRs and flavocyt c sulfide dehydrogenases (FCSDs) [110,111], or LMW persulfides from the metabolism of L-cys by MST and cysteinyl-tRNA synthetase [112,113]. H2S oxidation can be catalyzed by the concerted actions of three enzymes: SQR, PDO, and Rhd [114,115]. The work model established with Gram-negative bacterial SQR systems suggests a series of sequential reactions, including oxidation of sulfide to polysulfide by membrane-bound SQR, formation of GSSH from spontaneous reaction between polysulfide and GSH, oxidation of the sulfane sulfur in GSSH to sulfite catalyzed by PDO, and formation of thiosulfate from spontaneous reaction between polysulfide and sulfite [116,117] (Figure 4). Rhd, a thiosulfate sulfurtransferase that tends to speed up the reaction of polysulfide with GSH to produce GSSH, is not essential, as the reaction can occur spontaneously [104,107]. Although wildly distributed, SQRs have been proposed as primarily serving as sulfide detoxifiers, as they are present in mitochondria of many eukaryotes [70] (Figure 2). While SORs are structurally diverse, they can be conveniently grouped into two types according to their PDO partners, a well-known member of 2His–1Asp mononuclear iron-containing enzyme superfamily [110,118,119,120]. Type I includes PDOs in mitochondria and heterotrophic bacteria, and therefore, SQRs working with this type of PDOs function in the cytoplasm (Figure 2). Type II is found in dissimilatory sulfur-oxidizing bacteria, such as Rhodobacter capsulatus. In this case, PDOs are located outside the cytoplasmic membrane, and, consistently, SQRs catalyze the reaction at the outer surface of the cytoplasmic membrane [121,122,123] (Figure 4). In some bacteria, PDO and Rhd are naturally fused into a single polypeptide, becoming a bifunctional enzyme [124,125]. In addition to PDO and Rhd, CstB also plays a role in sulfane sulfur (RSSH) oxidation, converting two equivalents of persulfide to thiosulfate as a final product [124].
Flavocyt c sulfide dehydrogenases (FCSDs) represent another group of sulfide oxidizing enzymes in bacteria [111,114,126] (Figure 4). FCSDs, widely distributed in diverse bacteria, comprise two subunits in the periplasm, a large sulfide-binding flavoprotein and a small cyt c [114]. The final production of sulfide oxidation catalyzed by FCSDs is polysulfide [111]. It should be noted that many bacteria are equipped with multiple sulfide-oxidizing systems, including both SQR and FCSD [127]. Although SQR is generally regarded as the predominant sulfide-oxidizing system, FCSD may confer cells carrying both systems advantages under certain growth conditions.
4.5. Oxidation of S0 and S2+
In sulfur-oxidizing (Sox) prokaryotes, the best-studied system for sulfur oxidation is the Sox multienzyme system located in the periplasm [72] (Figure 4). The Sox systems may have promiscuity for substrates as they have been reported to carry out oxidation of sulfide and sulfite in certain bacteria [114,128]. Nevertheless, the primary role that the Sox system plays is to catalyze oxidation of zero-valent sulfur (ZVS) from a variety of sulfur compounds [72]. Among them, thiosulfate is considered particularly important, as it is a main product of most S2−/S0 oxidation and fulfills a key role in the sulfur cycle [14]. In Paracoccus pantrophus, whose Sox system has been intensively studied, the sox gene cluster comprises at least two transcriptional units with fifteen genes, seven of which, soxXYZABCD, encode proteins essential for sulfur oxidation in vitro [49,129,130]. The Sox pathway initiates with the activation of unconjugated SoxYZ with thiosulfate involving both SoxAX and SoxB, resulting in a SoxYZ-S-sulfane adduct via intermediate SoxYZ-S-thiosulfonate [131,132,133] (Figure 4). Once activated, the oxidation cycle commences. A disulfide bond between the sulfane sulfur of thiosulfate and the persulfurated active site cys residue on the carrier arm of SoxYZ is formed by SoxAX, where heme 2 is an active site (Figure 3D left), generating SoxYZ-S-thioperoxosulfonate, which subsequently is converted to SoxYZ-S-thiosulfane after releasing a molecule of sulfate under the catalysis of SoxB. For catalysis, the carrier arm of SoxY has to be positioned at the substrate channel of SoxB, leading to formation of a disulfide bond between Cys151 of SoxY and Trp175 of SoxB (Figure 3D, right). This reaction catalyzed by SoxB also generates the SoxYZ-S-sulfane, which acts as the activated adduct to start a new round of thiosulfate oxidation [131,132].
In many organisms, this aerobic Sox pathway lacks certain components, depending on species, and can only oxidize sulfide to soluble elemental sulfur [49,134]. For example, Rhodobacter spp. converts sulfide to sulfur using a Sox system without SoxCD [114]. The resulting zero-valent sulfur can be either released to the surroundings or stored in the periplasm as sulfur globules, a subject which has recently been reviewed in detail by Dahl [72]. Although the chemical nature of the sulfur in the globules is still under debate, it is increasingly accepted that octasulfur (S8) in a nano-crystalline form may be the main representative [72,135]. In addition, the truncated Sox system may work in conjunction with other sulfur oxidation pathways to complete oxidation of thiosulfate to sulfate [70]. Among them, the cytoplasmic sulfur oxidation pathway involving TusA is the first to be identified. The TusA protein, which has been identified as a central element supplying and transferring sulfur as persulfide to a number of important biosynthetic pathways, is implicated in providing elemental sulfur imported from the extracellular space to DsrABC for oxidation to sulfite [119,136,137]. However, despite recent advances, the mechanisms underpinning the transport of sulfur into the cytoplasm and the TusA involvement have to await further exploration. Recently, two other pathways in sulfur-oxidizing prokaryotes have been identified to complement the truncated Sox system for oxidation of thiosulfate to sulfite. One is composed of the proteins resembling the HdrA, B, and C subunits of heterodisulfide reductase from methanogenic archaea [138,139]. This pathway may require TusA-like proteins as well, and, more importantly, a lipoate-binding protein [140]. Given the indispensability of the lipoate-binding protein, it has been suggested that reduction of lipoate to dihydrolipoate is a part of the reaction cycle [140]. The other is a mixture of SoxB, TsdA and cytoplasmic sulfur dioxygenase (SDOs) [100]. In this pathway, TsdA converts thiosulfate to tetrathionate, from which sulfone is released by SoxB to form zero-valent sulfur, and subsequently zero-valent sulfur is oxidized to sulfite by SDOs. It is worth noting that in many sulfur oxidizers, SDOs function independently to oxidize sulfane sulfur bound to GSH (GSSH, and GSSnH) [141,142].
Thiosulfate can also be oxidized to tetrathionate, a process identified in diverse prokaryotes [72] (Figure 4). Enzymatic systems capable of catalyzing this reaction include thiosulfate dehydrogenase (Tsd) and partial Sox complex, representing a good interpretation of the branched thiosulfate oxidation [143]. Tsd, a widespread and well-studied system, is commonly composed of two functional diheme cyt c subunits TsdA and TsdB, which may be fused into a single polypeptide in some bacteria, such as M. purpuratum [96,144]. TsdA and TsdB function as the catalytic subunit and the electron acceptor partner respectively [145]. As mentioned earlier, TsdA is in fact a bidirectional enzyme that also acts as a tetrathionate reductase in some bacteria, such as C. jejuni, in which TsdB is missing [95,97]. The crystal structure of TsdA reveals His/Cys iron coordination for Heme 1, the active site of the enzyme, to which a thiosulfate covalently can be bound via the cys [96,145,146]. The diheme cyt c TsdB, which may be fused to TsdA, is the electron-accepting unit for transport electrons abstracted from the oxidation to cyt c heme-copper oxidase (HCO) of the respiratory chain for energy generation, although the exception has been found [14,96] (Figure 3B,C). Comparison of the structure of C. jejuni TsdA predicted by AlphaFold to a structurally available counterpart catalyzing thiosulfate oxidation reveals a high level of structural similarity. However, C. jejuni TsdA has a long random coil in the N-terminus (Figure 3C). According to phylogenetic and genomics analyses, the enzymes identified as thiosulfate dehydrogenase are distributed more broadly [14,96,145] (Figure 4).Intriguingly, some bacteria, such as S. oneidensis, are equipped with independent enzymatic systems catalyzing oxidation and reduction of thiosulfate [14].
In addition, thiosulfate oxidation can occur as an intermediate step in the oxidation of reduced sulfur compounds to sulfate, called tetrathionate (S4O62−) intermediate (S4I) pathway [134,147,148]. In the S4I pathway, the conversion of thiosulfate to tetrathionate is catalyzed by thiosulfate:quinone oxidoreductase TQO [149]. Unlike Tsd, the TQO enzyme transfers electrons into the respiratory chain via quinones, and beyond this, little is known about the catalytic mechanism of the enzyme.
In prokaryotes, oxidation of tetrathionate to sulfate, whether it is a part of the S4I pathway or not, has been known for some time [148,149,150,151,152]. Tetrathionate hydrolase, TetH, is responsible for hydrolysis of tetrathionate to sulfate and disulfane monosulfonic acid (−S-S-SO3−) in the S4I pathway [122,150,153]. Disulfane monosulfonic acid is a highly reactive sulfur species, which either quickly decomposes to sulfur and thiosulfate or autocombines into long-chain sulfur compounds, ultimately resulting in the production of elemental sulfur and sulfite. Another enzyme known to oxidize tetrathionate is thiol dehydrotransferase from Advenella kashmirensis, which is independent of S4I [152]. This enzyme is able to catalyze two sequential reactions, the formation of tetrathionate from thiosulfate and the reaction of tetrathionate with reduced GSH. The resulting product, GSH sulfodisulfane adduct, can be fed into the thiosulfate oxidation cycle catalyzed by Sox [152].
4.6. Oxidation of S4+
Sulfite is a highly reactive nucleophile and could cause damage to DNA and proteins through reacting with disulfide bonds [154,155]. Although sulfite can be spontaneously oxidized to sulfate in oxic environments, in order to protect cells against sulfite-induced damage, virtually all forms of living organisms are capable of performing biotic oxidation of sulfite [155,156]. In addition, sulfite has recently been reported to be protective against oxidative stress induced by high concentrations of glutamate called oxytosis in mammals [157], which has recently been suggested to be equivalent to ferroptosis [158]. In bacteria, the prevailing route for sulfite oxidation, called the direct route, is carried out by molybdenum (Mo)-containing sulfite-oxidizing enzymes (SOEs) in the periplasm [159] (Figure 4). All of the SOEs possess a cis-dioxo MoO22+ center that is bound to a unique pterin, molybdopterin, which is essential for catalysis [160,161]. Bacterial SOE, commonly termed as sulfite dehydrogenase (SDH), differs from its eukaryotic counterpart, which is a single polypeptide enzyme, in that it requires a cyt c subunit as electron acceptor in addition to the Mo-containing catalytic subunit [156,162]. The electrons extracted from the sulfite oxidation process are transferred to the HCO oxidase of the respiratory chain via the cyt c subunit for energy generation [163,164].
Oxidation of sulfite to sulfate can also occur in the bacterial cytoplasm (Figure 2). The enzymatic complex, SoeABC, first identified in Allochromatium vinosum but universally present in bacteria, comprises cytoplasmic molybdoprotein SoeA and iron-sulfur protein SoeB, as well as membrane-bound SoeC [127,147]. In many bacteria, SoeABC appears to be the major sulfite-oxidizing agent. Less commonly, the oxidation of sulfite to sulfate can be achieved via reversing the sulfate reduction pathway [72,155].
5. Physiological Impacts of RSS
Investigations into the physiological roles of RSS in bacteria are primarily centered on H2S, largely due to its physiological and pharmacological effects as a redox signaling molecule in mammals [9]. Despite the beneficial role of H2S at low concentrations, at high concentrations (millimolar), it is toxic to virtually all living organisms [9]. H2S is a highly poisonous gas, as a cause of inhalational deaths second only to carbon monoxide (CO), and this reputation has been known for centuries [165]. All other RSSs exhibit similar concentration-dependent effects on the physiology of living organisms. Not surprisingly, elemental sulfur and inorganic polysulfides have been explored as a weaponry to inhibit various types of pathogenic and drug-resistant microorganisms [44,166].
Persulfidation of proteins by H2S has been repeatedly reported as an important means in signaling, but this process is still under debate because H2S per se cannot react directly with thiols or does so after autooxidation in a sufficiently rapid manner [8]. Nonetheless, it is clear that disulfide bonds within proteins can be easily subjected to persulfide modification through a nucleophilic attach by the sulfide anion [8]. This may be particularly significant in prokaryotes because such proteins are abundant in the oxidizing milieu of the periplasm where the sulfide anion generated biotically and abiotically is present [8]. Nonetheless, some other RSSs, especially sulfane sulfur, including those in persulfide and polysulfide forms, and elemental sulfur (S8), which are more active than H2S, have been shown to be the leading agents for protein persulfidation [167,168,169].
Previous work in mammals has demonstrated that protein persulfidation affects specific activity in either positive (activation) or negative (inactivation) way, modulating diverse biological processes [170,171,172]. Upon sulfane sulfur treatments, persulfidation of the Staphylococcus aureus proteome is widespread [173]. The treatments increase the number of the modified proteins by over 30%, from 238 to 305. In addition to proteins involved in transcriptional regulation and metabolism (will be discussed later in the section of RSS sensing), the study reveals many secreted virulence factors and two uncharacterized thioredoxin-like proteins.
5.1. Inhibition of Energy Conservation and Growth
The ultimate phenotype of H2S at high concentrations is growth inhibition, which has been seen from a variety of bacteria, such as E. coli, S. oneidensis, Acinetobacter baumanni, to name a few [64,174]. In humans, the primary target of H2S is the mitochondrial oxygen reductase, a cyt c-type HCO which carries out reduction of O2 to H2O using electrons from soluble cyt c [174,175]. HCOs are also commonly found in bacteria, but in different forms, including cyt aa3 oxidases (mitochondrial-like oxidases) as in Bacillus subtilis and S. aureus, cyt bo3 oxidases as in E. coli, and cyt cbb3 as in S. oneidensis and Pseudomonas aeruginosa [100]. While all of HCOs employ the proton-pumping mechanism during oxygen reduction to generate energy efficiently, they are highly sensitive to a variety of small molecules, including H2S, CO, HCN, and nitrite/nitric oxide (NO) [100,176,177,178,179].
H2S inhibits HCOs in a biphasic manner, non-competitive with respect to either O2 or cyt c (for cyt c-type HCOs) [180]. The first molecule of H2S (in the form of HS−) binds to CuB in either cupric or cuprous state, and then is transferred to the ferric heme a3, resulting in inactivation of the catalytic activity of the enzyme [180,181]. Subsequently, the second molecule of HS− interacts with the enzyme to form the final protein-inhibitor adduct, in which heme a and CuA likely stay reduced [180]. During the reaction, H2S is probably oxidized to persulfide species. Unlike CO or NO, H2S may not react with fully reduced enzyme, and thus HCOs in the oxidized or mixed valence states are probably involved in both inhibitory and metabolic reactions with sulfide [180].
Similarly, H2S rapidly and effectively inhibits the activity of quinol-type HCOs, such as E. coli bo3 [182]. The apparent half-maximal inhibitory concentrations of H2S for cyt c-type and quinol-type HCOs are comparable, 0.55 and 1.1 μM, respectively [182,183]. Interestingly, the inhibition of the bo3 oxidase is fully reversible [182]. Although such an observation was not reported before with cyt c-type HCOs, it is probably common to all HCOs.
The inhibitory effect of H2S on bacterial growth is well recognized, but the molecule usually does not kill bacterial cells, contrasting with its lethality for mammals. One explanation would be that prokaryotes commonly carry additional terminal oxidases, the bd-type quinol oxidases in particular [179]. The bd oxidase lacks a counterpart proton-pumping mechanism as in HCOs and is not efficient in generation of proton motive force [184,185]. Consequently, unlike HCO, whose main role is to conserve energy, the bd oxidase endow bacterial cells with resistance to various harmful molecules, including H2S [179,186,187,188]. It has been found that the bd oxidases of E. coli remain active in the presence of H2S at 58 μM, indicating that the bd oxidases are insensitive to H2S [182,188]. Moreover, the difference of the oxidases in resistance to H2S may have profound impacts in bacterial physiology. It has been found recently that endogenous H2S alters energy metabolism and growth of Mycobacterium species by modulating activities of different oxidases [189,190].
5.2. Role of H2S in Oxidative Stress Response and Immune Response
H2S alone exerts little bactericidal activity against bacterial cells, but when combined with hydrogen peroxide (H2O2), it mediates a dramatic increase in cytotoxicity [64] (Figure 5A). This observation suggests that the cellular targets subjected to inhibition of H2S are beyond HCOs in bacteria. One of the key targets is catalases, most of which are hemoproteins primarily responsible for scavenging H2O2 [191,192]. It is well established that H2S can interact with several metals, including iron, copper, nickel, and zinc [193]. In the presence of H2O2, the interaction of H2S with the heme iron complex of hemoproteins leads to formation of sulfheme (sulfur-incorporated porphyrin) iron complex, resulting in loss of enzymatic activity [194,195]. Without protection of catalases, the bacterial cells are quickly killed by H2O2 [64,196,197] (Figure 5A). It is worth mentioning that an additional mechanism underlying the inhibition of catalases by H2S has been identified in plants, which is protein persulfidation, a result of the interaction between H2S and thiol groups of targets [198].
Interestingly, the combinatorial redox action of H2S and H2O2 that promotes cytotoxicity is just one side of the story. A contrasting effect has been seen, even if the timing of application is different (Figure 5A). When applied sequentially, H2S, like NO, acts as a molecule protecting E. coli, Staphylococcus, and Bacillus from ROS-mediated killing [51,199,200]. A similar phenomenon has also been observed from the combination of NO and H2O2 against a variety of bacteria [51,64]. Under this circumstance, bacterial cells promptly activate OxyR (or its equivalent), the master transcriptional regulating modulating cellular oxidative stress response. As a result, production of all sorts of ROS scavengers and damage-control proteins is drastically enhanced, leading to an increase in the bacterial tolerance to ROS [64,201] (Figure 5A). Given that oxidative stress is intertwined with antibiotic susceptibility, endogenous H2S mediates the sensitivity of various bacteria to a range of antibiotic agents [51]
Most recently, H2S has been recognized as playing a crucial role in human health by regulating immune response against bacterial pathogens. On one hand, endogenous H2S produced by cells of E. coli and S. aureus at elevated levels increases the resistance to immune-mediated killing [202]. Consistently, inhibitors of bacterial H2S biogenesis potentiate bactericidal antibiotics against P. aeruginosa and S. aureus in vitro and in mouse models of infection [203]. On the other hand, H2S released by host cells also affects pathogenesis [204]. In mice lacking CSE, cells of bacterial pathogen Mycobacterium tuberculosis survive less and have reduced colony forming units. This compromised pathogenesis by host H2S is a result of altered central metabolism, including glycolysis and the pentose phosphate pathway [204]. While many questions remain unanswered, all of these new findings manifest that H2S homeostasis during pathogenesis is critically linked to immunometabolism.
The physiological overlap and/or cross-talk between RSS and RNS, especially H2S and nitric oxide (NO), has come to the frontline of research in recent years and is reviewed in detail by Ivanovic-Burmazovic and Filipovic [205]. This is conceivable, as all of these compounds impact protein activity through thiol modifications, either synergistically or antagonistically [206]. In accord with this, thionitrous acid (HSNO), the smallest S-nitrosothiol formed from reaction of sulfide with low-molecular weight and/or protein S-nitrosothiols, including NO, is found to be the key molecule in cell signaling [207,208,209]. In mammals, it has been reported that the interaction of H2S and NO produces polysulfides (H2Sn), which serves as an activating molecule for TRPA1 channels [210]. In addition, organic thiols (RSH) can react with nitrite (NO2–) to form organic nitrosothiols (RSNO), which readily react with H2S at acidic pH to form a mixture of polysulfides [211]. Physiologically, nitroxyl donors via formation of HSNO and its cousins, such as nitrosopersulfide (SSNO–), could elevate cellular levels of RSS in S. aureus [212,213]. Despite these insights, impacts of the interplay of RSS and RNS on the biology of prokaryotes remain largely unknown.
5.3. Role in Metal Reduction
Electroactive microorganisms known to date, mostly bacteria, are renowned for their respiratory versatility, are capable of respiring an array of diverse chemicals as electron acceptors, and play a critical role in the biogeochemical cycle of elements, including metals, such as iron [214,215,216]. As a large portion of minerals are insoluble, these microorganisms have evolved multiple strategies for extracellular electron transfer, through which the electrons abstracted from oxidation of electron donors are transferred to multivalent metal ions within minerals [214]. Many bacteria, among which S. oneidensis is best studied, use porin-cyt c complexes to transport electrons from the cytoplasmic membrane to the outer cell surface. Nanowires, electrically conductive pili in which cyt c proteins are stacked, as best illustrated in Geobacter sulfurreducens, represent another strategy; while this electron transport machinery is found less commonly, it could deliver electrons to minerals at a distance. Additionally, soluble electron shuttles can be utilized alone or in conjunction with the first two means [147,215,217,218].
Despite these effective strategies, metal reduction in nature by electroactive bacteria could be drastically affected by environmental parameters. These microorganisms, including research models S. oneidensis and G. sulfurreducens, usually have exceptional ability to transform sulfur species. For example, S. oneidensis is able to not only reduce sulfite, thiosulfate, tetrathionate, and elemental sulfur to H2S, but also to oxidize thiosulfate to tetrathionate [14,64,89,108,219]. Sulfur species, generated from sulfur recycling in natural systems through a number of oxidation, reduction and disproportionation reactions both biotically and abiotically, have been demonstrated to be indispensable players in metal reduction [220,221,222,223]. Center to this sulfur-mediated metal reduction is HS−, which is produced by microbial reduction of various sulfur species and reacts with Fe3+ to form FeS (Figure 5B). This abiotic reaction between HS− and Fe3+ not only accounts for reduction of a large quantity of iron oxides but also offers an explanation of the low concentrations of HS− in aquifers, in addition to the oxidation to sulfate [220,224]. Given that these intertwined reactions of the sulfur and iron cycle are in part biological, electroactive microorganisms that maintain both iron and sulfur metabolic pathways may have an impact in ecology more substantially than previously expected.
6. RSS Sensing and Regulation
In order to survive and thrive in their natural niches, bacteria have evolved diverse adaptive strategies to swiftly respond to various adverse stress conditions, and transcriptional regulation is key for this adaptation [225]. As signal molecules, RSS tend to trigger modification of specific targets via protein persulfidation and subsequently affect the down-stream biological processes. To combat RSS at high concentrations, cells have various enzymes and regulatory systems to protect against excessive levels of modification [43]. A common set of proteins capable of cleaning or reducing the modification include thioredoxin and/or peroxiredoxin, and this scenario appears to be conserved in bacteria [46,226,227]. In Synechococcus sp. PCC7002, all of six peroxiredoxins are induced by S8, and peroxiredoxin I (PrxI) has been experimentally demonstrated to be able to reduce S8 to H2S [46]. To regulate the expression of the genes encoding common sulfide detoxification or repairing enzymes, a variety of mechanisms governing cellular responses to RSS have been identified in bacteria, among which transcriptional regulation by DNA-binding proteins is the most prevalent and important.
In S. aureus, cstR (CsoR-like sulfurtransferase repressor), located in cts operon, is the first gene reported to be involved in persulfide-responsive transcriptional regulation, which modulates the expression of five other cts genes to function [228,229]. CstR is an all-helical protein characterized by a disc-shaped D2-symmetric or pseudosymmetric tetrameric architecture and homotetrameric in solution, harboring four peripheral dithiol sensing sites (Figure 6, left). This regulator responds to cellular sulfide stress through sensing LMW persulfides and inorganic polysulfides (Sn2–), forming a mixture of di-, tri-, and tetra-sulfide interprotomer linkages between Cys31 and Cys60 [59,213,228]. Despite this, the information about DNA recognition and the changes upon inducer binding in CstR, along with other members of the CsoR family, which was defined recently, is rather limited [230]. CstR in bacteria, especially in the intestinal ones, such as Enterococcus faecalis, has been proved important to sustain sulfur-homeostasis as the intestinal flora is the major producer of L-homocys, one of the constituents of the LMW thiol pool [213,231,232].
Another sulfide-related regulator identified to date is SqrR (sulfide:quinone redcutase repressor) of R. capsulatus, a member of the ArsR family of bacterial repressors, which are widely distributed in Proteobacteria [225,233]. Experiments in vitro showed SqrR forms a dimer (Figure 6 right) and senses sulfide via an intramolecular tetrasulfide-bond formed between two conserved cys residues, Cys41 and Cys107 [234,235]. A homologue of SqrR, BigR (Biofilm growth-associated repressor), characterized in Xylella fastidiosa [236,237,238] and Acinetobacter baumannii [227], regulates a secondary RSS detoxification system [227,239]. BigR is responsive to H2S via forming a disulfide bond between Cys42 and Cys108 [234]. PigS, another member of the ArsR family [239], regulates part of the PigP regulon involved in the biosynthesis of the antibiotic prodigiosin (Pig), implying a regulatory connection between antibiotic biosynthesis and H2S/RSS homeostasis in Serratia [59]. The pigS gene per se is directly under transcriptional regulation of PigP, a novel master regulator that is implicated in secondary metabolism in Serratia and whose homologues are found only in a very limited group of Enterobacteriaceae [59]. Additionally, FisR (Fis family transcriptional regulator) acts as a RSS-responsive activator in Cupriavidus pinatubonensis, sensing RSS via forming disulfide and tetrasulfide cross-links among three conserved cys residues [235,240].
To date, a large number of studies have demonstrated physiological cross-talks between ROS and H2S/RSS [59,173,241,242]. Conceivably, the systems responding to ROS might also be able to sense RSS. OxyR, the master transcriptional regulator mediating cellular responses to H2O2-induced oxidative stresses in a variety of bacteria, has been found to be able to sense sulfane sulfur [43]. Most recent investigations have revealed that LasR and OhrR of P. aeruginosa, the primary transcriptional regulators in quorum sensing and the response to organic peroxides respectively, can also sense sulfane sulfur [243,244]. Thus, RSSs, especially sulfane sulfur, represent a group of new signaling factors that are involved in diverse biological processes, whose regulation is dependent on both RSS-specific regulators and global regulators primarily responsive to other environmental cues.
7. Concluding Remarks
In this review, we have summarized recent advances in generation of H2S and its derivative RSS via amino acid metabolism and biological transformation of inorganic sulfur species, their influence on the physiology, and how they are perceived in bacteria. In both higher organisms and prokaryotes, the amino acid metabolism as the source of H2S has been regarded to be the most critical one for the beneficial roles of H2S, signaling in particular. On the contrary, avast amount of H2S and RSS can be produced through biological transformation of inorganic sulfur compounds, which not only predominantly drives the global sulfur biogeochemical cycle, but also imposes a threat to cells. It is clear that the processes with sulfur compounds as electron donors and/or acceptors are early contributors to energetics prior to O2. To date, we have known a great deal about the enzymes that catalyze most, if not all, of steps of the transformation. Nevertheless, new enzymes and novel functioning mechanisms of known enzymes are continuingly revealed, especially in recent years.
The inherent chemical reactivity of H2S and RSS, especially when in conjunction with other reactive species, challenges development of all lifeforms. Redox-sensitive proteins are primary targets of H2S and RSS because of their susceptibility to modification by the formation of protein-inhibitor adducts. Consequently, the redox status of the cell, and the factors accounting for maintaining redox homeostasis, including those involved in combating ROS and RNS stresses, are particularly important in promoting or antagonizing the effects of H2S and RSS. In recent years, multiple lines of evidence have substantiated that bacterial cells have developed a sophisticated system to maintain homeostasis of H2S and RSS, many members of which are transcriptionally responsive to H2S and RSS. Furthermore, recent advances also convince us to consider H2S and RSS as a promising drug in promoting the effectiveness of antimicrobials and beyond. Nonetheless, the field of RSS chemical biology is still in its infancy, with many more questions open than answered. This may be particularly true with prokaryotes, as they are so diverse in phylogeny and metabolism. For instance, cellular targets of RSS are likely different in bacteria, even at the level of species and strain. Thus, with the availability of enormous amounts of genomic sequences and data in various omics, bacteria are no doubt a repertoire of living organisms for unraveling new mechanisms of RSS formation and metabolism impacting cellular signaling and function.
Author Contributions
S.H., Y.L. and H.G. wrote the manuscript. H.G. finalized the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by National Natural Science Foundation of China grants 31930003 and 41976087.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Giles, G.I.; Tasker, K.M.; Jacob, C. Hypothesis: The role of reactive sulfur species in oxidative stress. Free Radic. Biol. Med. 2001, 31, 1279–1283. [Google Scholar] [CrossRef] [PubMed]
- Sies, H.; Jones, D.P. Reactive oxygen species (ROS) as pleiotropic physiological signalling agents. Nat. Rev. Mol. Cell Biol. 2020, 21, 363–383. [Google Scholar] [CrossRef] [PubMed]
- Herb, M.; Schramm, M. Functions of ROS in macrophages and antimicrobial Immunity. Antioxidants 2021, 10, 313. [Google Scholar] [CrossRef]
- Forman, H.J.; Zhang, H. Targeting oxidative stress in disease: Promise and limitations of antioxidant therapy. Nat. Rev. Drug Discov. 2021, 20, 689–709. [Google Scholar] [CrossRef]
- Sies, H. Chapter 13—Oxidative Stress: Eustress and Distress in Redox Homeostasis. In Stress: Physiology, Biochemistry, and Pathology; Fink, G., Ed.; Academic Press: Cambridge, MA, USA, 2019; pp. 153–163. [Google Scholar]
- Khattak, S.; Rauf, M.A.; Khan, N.H.; Zhang, Q.-Q.; Chen, H.-J.; Muhammad, P.; Ansari, M.A.; Alomary, M.N.; Jahangir, M.; Zhang, C.-Y.; et al. Hydrogen sulfide biology and its role in cancer. Molecules 2022, 27, 3389. [Google Scholar] [CrossRef]
- Olson, K.R. Are reactive sulfur species the new reactive oxygen species? Antioxid. Redox Signal. 2020, 33, 1125–1142. [Google Scholar] [CrossRef]
- Mishanina, T.V.; Libiad, M.; Banerjee, R. Biogenesis of reactive sulfur species for signaling by hydrogen sulfide oxidation pathways. Nat. Chem. Biol. 2015, 11, 457–464. [Google Scholar] [CrossRef] [Green Version]
- Filipovic, M.R.; Zivanovic, J.; Alvarez, B.; Banerjee, R. Chemical biology of H2S Signaling through persulfidation. Chem. Rev. 2018, 118, 1253–1337. [Google Scholar] [CrossRef]
- Kolluru, G.K.; Shen, X.; Kevil, C.G. Reactive sulfur species. Arter. Thromb. Vasc. Biol. 2020, 40, 874–884. [Google Scholar] [CrossRef]
- Lau, N.; Pluth, M.D. Reactive sulfur species (RSS): Persulfides, polysulfides, potential, and problems. Curr. Opin. Chem. Biol. 2019, 49, 1–8. [Google Scholar] [CrossRef]
- Gruhlke, M.C.H.; Slusarenko, A.J. The biology of reactive sulfur species (RSS). Plant Physiol. Biochem. 2012, 59, 98–107. [Google Scholar] [CrossRef]
- Moran, M.A.; Durham, B.P. Sulfur metabolites in the pelagic ocean. Nat. Rev. Microbiol. 2019, 17, 665–678. [Google Scholar] [CrossRef] [PubMed]
- Yu, Q.; Sun, W.; Gao, H. Thiosulfate oxidation in sulfur-reducing Shewanella oneidensis and its unexpected influences on the cytochrome c content. Environ. Microbiol. 2021, 23, 7056–7072. [Google Scholar] [CrossRef]
- Landa, M.; Burns, A.S.; Durham, B.P.; Esson, K.; Nowinski, B.; Sharma, S.; Vorobev, A.; Nielsen, T.; Kiene, R.P.; Moran, M.A. Sulfur metabolites that facilitate oceanic phytoplankton–bacteria carbon flux. ISME J. 2019, 13, 2536–2550. [Google Scholar] [CrossRef]
- Lu, J.; Holmgren, A. The thioredoxin antioxidant system. Free Radic. Biol. Med. 2014, 66, 75–87. [Google Scholar] [CrossRef]
- Olson, K.R. Reactive oxygen species or reactive sulfur species: Why we should consider the latter. J. Exp. Biol. 2020, 223, jeb196352. [Google Scholar] [CrossRef]
- Sojo, V.; Herschy, B.; Whicher, A.; Camprubí, E.; Lane, N. The origin of life in alkaline hydrothermal vents. Astrobiology 2016, 16, 181–197. [Google Scholar] [CrossRef]
- Garcia, P.S.; D’Angelo, F.; Ollagnier de Choudens, S.; Dussouchaud, M.; Bouveret, E.; Gribaldo, S.; Barras, F. An early origin of iron–sulfur cluster biosynthesis machineries before Earth oxygenation. Nat. Ecol. Evol. 2022, 6, 1564–1572. [Google Scholar] [CrossRef]
- Nitschke, W.; Russell, M.J. Beating the acetyl coenzyme A-pathway to the origin of life. Philos. Trans. R. Soc. B 2013, 368, 20120258. [Google Scholar] [CrossRef] [Green Version]
- Ritson, D.J. A cyanosulfidic origin of the Krebs cycle. Sci. Adv. 2021, 7, eabh3981. [Google Scholar] [CrossRef]
- Wacey, D.; Kilburn, M.R.; Saunders, M.; Cliff, J.; Brasier, M.D. Microfossils of sulphur-metabolizing cells in 3.4-billion-year-old rocks of Western Australia. Nat. Geosci. 2011, 4, 698–702. [Google Scholar] [CrossRef]
- Philippot, P.; Van Zuilen, M.; Lepot, K.; Thomazo, C.; Farquhar, J.; Van Kranendonk, M.J. Early archaean microorganisms preferred elemental sulfur, not sulfate. Science 2007, 317, 1534–1537. [Google Scholar] [CrossRef] [PubMed]
- Marin, J.; Battistuzzi, F.U.; Brown, A.C.; Hedges, S.B. The timetree of prokaryotes: New insights into their evolution and speciation. Mol. Biol. Evol. 2016, 34, 437–446. [Google Scholar] [CrossRef] [PubMed]
- Jones, D.P.; Sies, H. The redox code. Antioxid. Redox Signal. 2015, 23, 734–746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olson, K.R.; Gao, Y.; DeLeon, E.R.; Arif, M.; Arif, F.; Arora, N.; Straub, K.D. Catalase as a sulfide-sulfur oxido-reductase: An ancient (and modern?) regulator of reactive sulfur species (RSS). Redox Biol. 2017, 12, 325–339. [Google Scholar] [CrossRef]
- Schmidt, A. Sulfur metabolism in Cyanobacteria. Methods Enzymol. 1988, 167, 572–583. [Google Scholar]
- Schopf, J.W. Disparate rates, differing fates: Tempo and mode of evolution changed from the Precambrian to the Phanerozoic. Proc. Natl. Acad. Sci. USA 1994, 91, 6735–6742. [Google Scholar] [CrossRef] [Green Version]
- Holland, H.D. Volcanic gases, black smokers, and the Great Oxidation Event. Geochim. Cosmochim. Acta 2002, 66, 3811–3826. [Google Scholar] [CrossRef]
- Planavsky, N.J.; Bekker, A.; Hofmann, A.; Owens, J.D.; Lyons, T.W. Sulfur record of rising and falling marine oxygen and sulfate levels during the Lomagundi event. Proc. Natl. Acad. Sci. USA 2012, 109, 18300–18305. [Google Scholar] [CrossRef] [Green Version]
- Poulton, S.W.; Bekker, A.; Cumming, V.M.; Zerkle, A.L.; Canfield, D.E.; Johnston, D.T. A 200-million-year delay in permanent atmospheric oxygenation. Nature 2021, 592, 232–236. [Google Scholar] [CrossRef]
- Lyons, T.W.; Reinhard, C.T.; Planavsky, N.J. The rise of oxygen in Earth’s early ocean and atmosphere. Nature 2014, 506, 307–315. [Google Scholar] [CrossRef] [PubMed]
- Schopf, J.W. Geological evidence of oxygenic photosynthesis and the biotic response to the 2400–2200 Ma “Great Oxidation Event”. Biochemistry 2014, 79, 165–177. [Google Scholar] [CrossRef] [PubMed]
- Kurland, C.G.; Andersson, S.G.E. Origin and evolution of the mitochondrial proteome. Microbiol. Mol. Biol. Rev. 2000, 64, 786–820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukherjee, I.; Large, R.R.; Corkrey, R.; Danyushevsky, L.V. The Boring Billion, a slingshot for complex life on earth. Sci. Rep. 2018, 8, 4432. [Google Scholar] [CrossRef] [Green Version]
- Case, A.J. On the origin of superoxide dismutase: An evolutionary perspective of superoxide-mediated redox signaling. Antioxidants 2017, 6, 82. [Google Scholar] [CrossRef] [Green Version]
- Zelko, I.N.; Mariani, T.J.; Folz, R.J. Superoxide dismutase multigene family: A comparison of the CuZn-SOD (SOD1), Mn-SOD (SOD2), and EC-SOD (SOD3) gene structures, evolution, and expression. Free Radic. Biol. Med. 2002, 33, 337–349. [Google Scholar] [CrossRef]
- Zamocky, M.; Gasselhuber, B.; Furtmuller, P.G.; Obinger, C. Molecular evolution of hydrogen peroxide degrading enzymes. Arch. Biochem. Biophys. 2012, 525, 131–144. [Google Scholar] [CrossRef] [Green Version]
- Inupakutika, M.A.; Sengupta, S.; Devireddy, A.R.; Azad, R.K.; Mittler, R. The evolution of reactive oxygen species metabolism. J. Exp. Bot. 2016, 67, 5933–5943. [Google Scholar] [CrossRef] [Green Version]
- Dibrova, D.V.; Cherepanov, D.A.; Galperin, M.Y.; Skulachev, V.P.; Mulkidjanian, A.Y. Evolution of cytochrome bc complexes: From membrane-anchored dehydrogenases of ancient bacteria to triggers of apoptosis in vertebrates. Biochim. Biophys. Acta-Bioenerg. 2013, 1827, 1407–1427. [Google Scholar] [CrossRef] [Green Version]
- Grace, S.C. Phylogenetic distribution of superoxide-dismutase supports an endosymbiotic origin for chloroplasts and mitochondria. Life Sci. 1990, 47, 1875–1886. [Google Scholar] [CrossRef]
- Miller, A.F. Superoxide dismutases: Ancient enzymes and new insights. Febs Lett. 2012, 586, 585–595. [Google Scholar] [CrossRef] [PubMed]
- Hou, N.; Yan, Z.; Fan, K.; Li, H.; Zhao, R.; Xia, Y.; Xun, L.; Liu, H. OxyR senses sulfane sulfur and activates the genes for its removal in Escherichia coli. Redox Biol. 2019, 26, 101293. [Google Scholar] [CrossRef] [PubMed]
- Sato, I.; Shimatani, K.; Fujita, K.; Abe, T.; Shimizu, M.; Fujii, T.; Hoshino, T.; Takaya, N. Glutathione reductase/glutathione is responsible for cytotoxic elemental sulfur tolerance via polysulfide shuttle in fungi. J. Biol. Chem. 2011, 286, 20283–20291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dóka, É.; Pader, I.; Bíró, A.; Johansson, K.; Cheng, Q.; Ballagó, K.; Prigge, J.R.; Pastor-Flores, D.; Dick, T.P.; Schmidt, E.E.; et al. A novel persulfide detection method reveals protein persulfide- and polysulfide-reducing functions of thioredoxin and glutathione systems. Sci. Adv. 2016, 2, e1500968. [Google Scholar] [CrossRef] [Green Version]
- Liu, D.; Chen, J.; Wang, Y.; Meng, Y.; Li, Y.; Huang, R.; Xia, Y.; Liu, H.; Jiao, N.; Xun, L.; et al. Synechococcus sp. PCC7002 uses peroxiredoxin to cope with reactive sulfur species stress. mBio 2022, 13, e01039-22. [Google Scholar] [CrossRef]
- van Vliet, D.M.; von Meijenfeldt, F.A.B.; Dutilh, B.E.; Villanueva, L.; Sinninghe Damsté, J.S.; Stams, A.J.M.; Sánchez-Andrea, I. The bacterial sulfur cycle in expanding dysoxic and euxinic marine waters. Environ. Microbiol. 2021, 23, 2834–2857. [Google Scholar] [CrossRef]
- Bronstein, M.; Schütz, M.; Hauska, G.; Padan, E.; Shahak, Y. Cyanobacterial sulfide-quinone reductase: Cloning and heterologous expression. J. Bacteriol. 2000, 182, 3336–3344. [Google Scholar] [CrossRef] [Green Version]
- Friedrich, C.G.; Bardischewsky, F.; Rother, D.; Quentmeier, A.; Fischer, J. Prokaryotic sulfur oxidation. Curr. Opin. Microbiol. 2005, 8, 253–259. [Google Scholar] [CrossRef]
- Vigneron, A.; Cruaud, P.; Alsop, E.; de Rezende, J.R.; Head, I.M.; Tsesmetzis, N. Beyond the tip of the iceberg; a new view of the diversity of sulfite- and sulfate-reducing microorganisms. ISME J. 2018, 12, 2096–2099. [Google Scholar] [CrossRef]
- Shatalin, K.; Shatalina, E.; Mironov, A.; Nudler, E. H2S: A universal defense against antibiotics in bacteria. Science 2011, 334, 986–990. [Google Scholar] [CrossRef]
- Mironov, A.; Seregina, T.; Nagornykh, M.; Luhachack, L.G.; Korolkova, N.; Lopes, L.E.; Kotova, V.; Zavilgelsky, G.; Shakulov, R.; Shatalin, K.; et al. Mechanism of H2S-mediated protection against oxidative stress in Escherichia coli. Proc. Natl. Acad. Sci. USA 2017, 114, 6022–6027. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Jhee, K.-H.; Kruger, W.D. Production of the neuromodulator H2S by cystathionine beta-synthase via the condensation of cysteine and homocysteine. J. Biol. Chem. 2004, 279, 52082–52086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, S.; Padovani, D.; Leslie, R.A.; Chiku, T.; Banerjee, R. Relative contributions of cystathionine beta-synthase and gamma-cystathionase to H2S biogenesis via alternative trans-sulfuration reactions. J. Biol. Chem. 2009, 284, 22457–22466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, S.; Banerjee, R. PLP-dependent H2S biogenesis. Biochim. Biophys. Acta 2011, 1814, 1518–1527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimura, Y.; Koike, S.; Shibuya, N.; Lefer, D.; Ogasawara, Y.; Kimura, H. 3-Mercaptopyruvate sulfurtransferase produces potential redox regulators cysteine- and glutathione-persulfide (Cys-SSH and GSSH) together with signaling molecules H2S2, H2S3 and H2S. Sci. Rep. 2017, 7, 10459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braunstein, A.E.; Goryachenkova, E.V.; Lac, N.D. Reactions catalysed by serine sulfhydrase from chicken liver. Biochim. Biophys. Acta 1969, 171, 366–368. [Google Scholar] [CrossRef] [PubMed]
- Chiku, T.; Padovani, D.; Zhu, W.; Singh, S.; Vitvitsky, V.; Banerjee, R. H2S biogenesis by human cystathionine gamma-lyase leads to the novel sulfur metabolites lanthionine and homolanthionine and is responsive to the grade of hyperhomocysteinemia. J. Biol. Chem. 2009, 284, 11601–11612. [Google Scholar] [CrossRef] [Green Version]
- Walsh, B.J.C.; Giedroc, D.P. H2S and reactive sulfur signaling at the host-bacterial pathogen interface. J. Biol. Chem. 2020, 295, 13150–13168. [Google Scholar] [CrossRef]
- Zhu, W.; Lin, A.; Banerjee, R. Kinetic properties of polymorphic variants and pathogenic mutants in human cystathionine gamma-lyase. Biochemistry 2008, 47, 6226–6232. [Google Scholar] [CrossRef] [Green Version]
- Ida, T.; Sawa, T.; Ihara, H.; Tsuchiya, Y.; Watanabe, Y.; Kumagai, Y.; Suematsu, M.; Motohashi, H.; Fujii, S.; Matsunaga, T.; et al. Reactive cysteine persulfides and S-polythiolation regulate oxidative stress and redox signaling. Proc. Natl. Acad. Sci. USA 2014, 111, 7606–7611. [Google Scholar] [CrossRef] [Green Version]
- Yadav, P.K.; Martinov, M.; Vitvitsky, V.; Seravalli, J.; Wedmann, R.; Filipovic, M.R.; Banerjee, R. Biosynthesis and reactivity of cysteine persulfides in signaling. J. Am. Chem. Soc. 2016, 138, 289–299. [Google Scholar] [CrossRef] [PubMed]
- Yadav, P.K.; Yamada, K.; Chiku, T.; Koutmos, M.; Banerjee, R. Structure and kinetic analysis of H2S production by human mercaptopyruvate sulfurtransferase. J. Biol. Chem. 2013, 288, 20002–20013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, G.; Li, N.; Mao, Y.; Zhou, G.; Gao, H. Endogenous generation of hydrogen sulfide and its regulation in Shewanella oneidensis. Front. Microbiol. 2015, 6, 374. [Google Scholar] [CrossRef] [PubMed]
- Fukamachi, H.; Nakano, Y.; Yoshimura, M.; Koga, T. Cloning and characterization of the L-cysteine desulfhydrase gene of Fusobacterium nucleatum. FEMS Microbiol. Lett. 2002, 215, 75–80. [Google Scholar] [CrossRef]
- Yoshida, Y.; Ito, S.; Kamo, M.; Kezuka, Y.; Tamura, H.; Kunimatsu, K.; Kato, H. Production of hydrogen sulfide by two enzymes associated with biosynthesis of homocysteine and lanthionine in Fusobacterium nucleatum subsp. nucleatum ATCC 25586. Microbiology 2010, 156, 2260–2269. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, Y.; Suwabe, K.; Nagano, K.; Kezuka, Y.; Kato, H.; Yoshimura, F. Identification and enzymic analysis of a novel protein associated with production of hydrogen sulfide and L-serine from L-cysteine in Fusobacterium nucleatum subsp. nucleatum ATCC 25586. Microbiology 2011, 157, 2164–2171. [Google Scholar] [CrossRef] [Green Version]
- Suwabe, K.; Yoshida, Y.; Nagano, K.; Yoshimura, F. Identification of an L-methionine γ-lyase involved in the production of hydrogen sulfide from L-cysteine in Fusobacterium nucleatum subsp. nucleatum ATCC 25586. Microbiology 2011, 157, 2992–3000. [Google Scholar] [CrossRef] [Green Version]
- Li, K.; Xin, Y.; Xuan, G.; Zhao, R.; Liu, H.; Xia, Y.; Xun, L. Escherichia coli uses separate enzymes to produce H2S and reactive sulfane sulfur from L-cysteine. Front. Microbiol. 2019, 10, 298. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Guo, X.; Li, H.; Qi, H.; Qian, J.; Yan, S.; Shi, J.; Niu, W. Hydrogen sulfide from cysteine desulfurase, not 3-mercaptopyruvate sulfurtransferase, contributes to sustaining cell growth and bioenergetics in E. coli under anaerobic conditions. Front. Microbiol. 2019, 10, 2357. [Google Scholar] [CrossRef]
- Fakhraee, M.; Hancisse, O.; Canfield, D.E.; Crowe, S.A.; Katsev, S. Proterozoic seawater sulfate scarcity and the evolution of ocean–atmosphere chemistry. Nat. Geosci. 2019, 12, 375–380. [Google Scholar] [CrossRef]
- Dahl, C. A biochemical view on the biological sulfur cycle. In Environmental Technologies to Treat Sulfur Pollution: Principles and Engineering; Lens, P.N.L., Ed.; IWA Publishing, 2020. [Google Scholar]
- Kushkevych, I.; Cejnar, J.; Treml, J.; Dordević, D.; Kollar, P.; Vítězová, M. Recent advances in metabolic pathways of sulfate reduction in intestinal bacteria. Cells 2020, 9, 698. [Google Scholar] [CrossRef] [PubMed]
- Fike, D.A.; Bradley, A.S.; Rose, C.V. Rethinking the ancient sulfur cycle. Annu. Rev. Earth Planet Sci. 2015, 43, 593–622. [Google Scholar] [CrossRef] [Green Version]
- Plugge, C.; Zhang, W.; Scholten, J.; Stams, A. Metabolic flexibility of sulfate-reducing bacteria. Front. Microbiol. 2011, 2, 81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, A.A.; Venceslau, S.S.; Grein, F.; Leavitt, W.D.; Dahl, C.; Johnston, D.T.; Pereira, I.A. A protein trisulfide couples dissimilatory sulfate reduction to energy conservation. Science 2015, 350, 1541–1545. [Google Scholar] [CrossRef] [PubMed]
- Aguilar-Barajas, E.; Díaz-Pérez, C.; Ramírez-Díaz, M.I.; Riveros-Rosas, H.; Cervantes, C. Bacterial transport of sulfate, molybdate, and related oxyanions. BioMetals 2011, 24, 687–707. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, Y.; Yoshikaie, K.; Takeuchi, A.; Ichikawa, M.; Mori, T.; Uchino, S.; Sugano, Y.; Hakoshima, T.; Takagi, H.; Nonaka, G.; et al. Crystal structure of a YeeE/YedE family protein engaged in thiosulfate uptake. Sci. Adv. 2020, 6, eaba7637. [Google Scholar] [CrossRef]
- Tai, C.-H.; Cook, P.F. Pyridoxal 5‘-phosphate-dependent α,β-elimination reactions: mechanism of O-acetylserine sulfhydrylase. Acc. Chem. Res. 2001, 34, 49–59. [Google Scholar] [CrossRef]
- Nakatani, T.; Ohtsu, I.; Nonaka, G.; Wiriyathanawudhiwong, N.; Morigasaki, S.; Takagi, H. Enhancement of thioredoxin/glutaredoxin-mediated L-cysteine synthesis from S-sulfocysteine increases L-cysteine production in Escherichia coli. Microb. Cell Fact. 2012, 11, 62. [Google Scholar] [CrossRef] [Green Version]
- Kawano, Y.; Onishi, F.; Shiroyama, M.; Miura, M.; Tanaka, N.; Oshiro, S.; Nonaka, G.; Nakanishi, T.; Ohtsu, I. Improved fermentative l-cysteine overproduction by enhancing a newly identified thiosulfate assimilation pathway in Escherichia coli. Appl. Microbiol. Biotechnol. 2017, 101, 6879–6889. [Google Scholar] [CrossRef]
- Callbeck, C.M.; Canfield, D.E.; Kuypers, M.M.M.; Yilmaz, P.; Lavik, G.; Thamdrup, B.; Schubert, C.J.; Bristow, L.A. Sulfur cycling in oceanic oxygen minimum zones. Limnol. Oceanogr. 2021, 66, 2360–2392. [Google Scholar] [CrossRef]
- Müller, A.L.; Kjeldsen, K.U.; Rattei, T.; Pester, M.; Loy, A. Phylogenetic and environmental diversity of DsrAB-type dissimilatory (bi)sulfite reductases. ISME J. 2015, 9, 1152–1165. [Google Scholar] [CrossRef] [PubMed]
- Colman, D.R.; Labesse, G.; Swapna, G.V.T.; Stefanakis, J.; Montelione, G.T.; Boyd, E.S.; Royer, C.A. Structural evolution of the ancient enzyme, dissimilatory sulfite reductase. Proteins Struct. Funct. Bioinf. 2022, 90, 1331–1345. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, T.F.; Vonrhein, C.; Matias, P.M.; Venceslau, S.S.; Pereira, I.A.; Archer, M. The crystal structure of Desulfovibrio vulgaris dissimilatory sulfite reductase bound to DsrC provides novel insights into the mechanism of sulfate respiration. J. Biol. Chem. 2008, 283, 34141–34149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grein, F.; Pereira, I.A.C.; Dahl, C. Biochemical characterization of individual components of the Allochromatium vinosum DsrMKJOP transmembrane complex aids understanding of complex function in vivo. J. Bacteriol. 2010, 192, 6369–6377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venceslau, S.S.; Stockdreher, Y.; Dahl, C.; Pereira, I.A.C. The “bacterial heterodisulfide” DsrC is a key protein in dissimilatory sulfur metabolism. Biochim. Biophys. Acta (BBA) Bioenerg. 2014, 1837, 1148–1164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simon, J.; Kroneck, P.M.H. Chapter Two—Microbial Sulfite Respiration. In Advances in Microbial Physiology; Poole, R.K., Ed.; Academic Press: Cambridge, MA, USA, 2013; Volume 62, pp. 45–117. [Google Scholar]
- Shirodkar, S.; Reed, S.; Romine, M.; Saffarini, D. The octahaem SirA catalyses dissimilatory sulfite reduction in Shewanella oneidensis MR-1. Environ. Microbiol. 2011, 13, 108–115. [Google Scholar] [CrossRef]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Zidek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]
- Varadi, M.; Anyango, S.; Deshpande, M.; Nair, S.; Natassia, C.; Yordanova, G.; Yuan, D.; Stroe, O.; Wood, G.; Laydon, A.; et al. AlphaFold Protein Structure Database: Massively expanding the structural coverage of protein-sequence space with high-accuracy models. Nucleic Acids Res. 2022, 50, D439–D444. [Google Scholar] [CrossRef]
- Barrett, E.L.; Clark, M.A. Tetrathionate reduction and production of hydrogen sulfide from thiosulfate. Microbiol. Rev. 1987, 51, 192–205. [Google Scholar] [CrossRef]
- Hensel, M.; Hinsley, A.P.; Nikolaus, T.; Sawers, G.; Berks, B.C. The genetic basis of tetrathionate respiration in Salmonella typhimurium. Mol. Microbiol. 1999, 32, 275–287. [Google Scholar] [CrossRef] [Green Version]
- Winter, S.E.; Thiennimitr, P.; Winter, M.G.; Butler, B.P.; Huseby, D.L.; Crawford, R.W.; Russell, J.M.; Bevins, C.L.; Adams, L.G.; Tsolis, R.M.; et al. Gut inflammation provides a respiratory electron acceptor for Salmonella. Nature 2010, 467, 426–429. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.-W.; Denkmann, K.; Kosciow, K.; Dahl, C.; Kelly, D.J. Tetrathionate stimulated growth of Campylobacter jejuni identifies a new type of bi-functional tetrathionate reductase (TsdA) that is widely distributed in bacteria. Mol. Microbiol. 2013, 88, 173–188. [Google Scholar] [CrossRef] [PubMed]
- Kurth, J.M.; Brito, J.A.; Reuter, J.; Flegler, A.; Koch, T.; Franke, T.; Klein, E.M.; Rowe, S.F.; Butt, J.N.; Denkmann, K.; et al. Electron accepting units of the diheme cytochrome c TsdA, a bifunctional thiosulfate dehydrogenase/tetrathionate reductase. J. Biol. Chem. 2016, 291, 24804–24818. [Google Scholar] [CrossRef] [Green Version]
- Jenner, L.P.; Kurth, J.M.; van Helmont, S.; Sokol, K.P.; Reisner, E.; Dahl, C.; Bradley, J.M.; Butt, J.N.; Cheesman, M.R. Heme ligation and redox chemistry in two bacterial thiosulfate dehydrogenase (TsdA) enzymes. J. Biol. Chem. 2019, 294, 18002–18014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dyson, A.; Dal-Pizzol, F.; Sabbatini, G.; Lach, A.B.; Galfo, F.; Dos Santos Cardoso, J.; Pescador Mendonça, B.; Hargreaves, I.; Bollen Pinto, B.; Bromage, D.I.; et al. Ammonium tetrathiomolybdate following ischemia/reperfusion injury: Chemistry, pharmacology, and impact of a new class of sulfide donor in preclinical injury models. PLoS Med. 2017, 14, e1002310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barton, L.L.; Fauque, G.D. Biochemistry, physiology and biotechnology of sulfate-reducing bacteria. Adv. Appl. Microbiol. 2009, 68, 41–98. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, R.; Xi, S.; Cai, R.; Zhang, X.; Sun, C. A novel bacterial thiosulfate oxidation pathway provides a new clue about the formation of zero-valent sulfur in deep sea. ISME J. 2020, 14, 2261–2274. [Google Scholar] [CrossRef]
- Laska, S.; Lottspeich, F.; Kletzin, A. Membrane-bound hydrogenase and sulfur reductase of the hyperthermophilic and acidophilic archaeon Acidianus ambivalens. Microbiology 2003, 149, 2357–2371. [Google Scholar] [CrossRef] [Green Version]
- Chernyh, N.A.; Neukirchen, S.; Frolov, E.N.; Sousa, F.L.; Miroshnichenko, M.L.; Merkel, A.Y.; Pimenov, N.V.; Sorokin, D.Y.; Ciordia, S.; Mena, M.C.; et al. Dissimilatory sulfate reduction in the archaeon ‘Candidatus Vulcanisaeta moutnovskia’ sheds light on the evolution of sulfur metabolism. Nat. Microbiol. 2020, 5, 1428–1438. [Google Scholar] [CrossRef]
- Jormakka, M.; Yokoyama, K.; Yano, T.; Tamakoshi, M.; Akimoto, S.; Shimamura, T.; Curmi, P.; Iwata, S. Molecular mechanism of energy conservation in polysulfide respiration. Nat. Struct. Mol. Biol. 2008, 15, 730–737. [Google Scholar] [CrossRef] [Green Version]
- Heinzinger, N.K.; Fujimoto, S.Y.; Clark, M.A.; Moreno, M.S.; Barrett, E.L. Sequence analysis of the phs operon in Salmonella typhimurium and the contribution of thiosulfate reduction to anaerobic energy metabolism. J. Bacteriol. 1995, 177, 2813–2820. [Google Scholar] [CrossRef] [PubMed]
- Burns, J.L.; DiChristina, T.J. Anaerobic respiration of elemental sulfur and thiosulfate by Shewanella oneidensis MR-1 requires psrA, a homolog of the phsA gene of Salmonella enterica serovar typhimurium LT2. Appl. Environ. Microbiol. 2009, 75, 5209–5217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zerkle, A.L.; Farquhar, J.; Johnston, D.T.; Cox, R.P.; Canfield, D.E. Fractionation of multiple sulfur isotopes during phototrophic oxidation of sulfide and elemental sulfur by a green sulfur bacterium. Geochim. Cosmochim. Acta 2009, 73, 291–306. [Google Scholar] [CrossRef]
- Libiad, M.; Yadav, P.K.; Vitvitsky, V.; Martinov, M.; Banerjee, R. Organization of the human mitochondrial hydrogen sulfide oxidation pathway. J. Biol. Chem. 2014, 289, 30901–30910. [Google Scholar] [CrossRef] [Green Version]
- Toohey, J.I. Sulfur signaling: Is the agent sulfide or sulfane? Anal. Biochem. 2011, 413, 1–7. [Google Scholar] [CrossRef]
- Xuan, G.; Lü, C.; Xu, H.; Chen, Z.; Li, K.; Liu, H.; Liu, H.; Xia, Y.; Xun, L. Sulfane sulfur is an intrinsic signal activating MexR-regulated antibiotic resistance in Pseudomonas aeruginosa. Mol. Microbiol. 2020, 114, 1038–1048. [Google Scholar] [CrossRef]
- Xin, Y.; Liu, H.; Cui, F.; Liu, H.; Xun, L. Recombinant Escherichia coli with sulfide:quinone oxidoreductase and persulfide dioxygenase rapidly oxidises sulfide to sulfite and thiosulfate via a new pathway. Environ. Microbiol. 2016, 18, 5123–5136. [Google Scholar] [CrossRef]
- Lü, C.; Xia, Y.; Liu, D.; Zhao, R.; Gao, R.; Liu, H.; Xun, L. Cupriavidus necator H16 uses flavocytochrome c sulfide dehydrogenase to oxidize self-produced and added sulfide. Appl Environ. Microbiol. 2017, 83, e01610-17. [Google Scholar] [CrossRef] [Green Version]
- Akaike, T.; Ida, T.; Wei, F.-Y.; Nishida, M.; Kumagai, Y.; Alam, M.M.; Ihara, H.; Sawa, T.; Matsunaga, T.; Kasamatsu, S.; et al. Cysteinyl-tRNA synthetase governs cysteine polysulfidation and mitochondrial bioenergetics. Nat. Commun. 2017, 8, 1177. [Google Scholar] [CrossRef] [Green Version]
- Nagahara, N.; Koike, S.; Nirasawa, T.; Kimura, H.; Ogasawara, Y. Alternative pathway of H2S and polysulfides production from sulfurated catalytic-cysteine of reaction intermediates of 3-mercaptopyruvate sulfurtransferase. Biochem. Biophys. Res. Commun. 2018, 496, 648–653. [Google Scholar] [CrossRef]
- Frigaard, N.-U.; Dahl, C. Sulfur metabolism in phototrophic sulfur bacteria. In Advances in Microbial Physiology; Poole, R.K., Ed.; Academic Press: Cambridge, MA, USA, 2008; Volume 54, pp. 103–200. [Google Scholar]
- Xia, Y.; Lü, C.; Hou, N.; Xin, Y.; Liu, J.; Liu, H.; Xun, L. Sulfide production and oxidation by heterotrophic bacteria under aerobic conditions. ISME J. 2017, 11, 2754–2766. [Google Scholar] [CrossRef] [PubMed]
- Kabil, O.; Banerjee, R. Characterization of patient mutations in human persulfide dioxygenase (ETHE1) involved in H2S catabolism. J. Biol. Chem. 2012, 287, 44561–44567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sattler, S.A.; Wang, X.; Lewis, K.M.; DeHan, P.J.; Park, C.-M.; Xin, Y.; Liu, H.; Xian, M.; Xun, L.; Kang, C. Characterizations of two bacterial persulfide dioxygenases of the metallo-β-lactamase superfamily. J. Biol. Chem. 2015, 290, 18914–18923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marcia, M.; Ermler, U.; Peng, G.; Michel, H. A new structure-based classification of sulfide:quinone oxidoreductases. Proteins Struct. Funct. Bioinf. 2010, 78, 1073–1083. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.-J.; Stockdreher, Y.; Koch, T.; Sun, S.-T.; Fan, Z.; Josten, M.; Sahl, H.-G.; Wang, Q.; Luo, Y.-M.; Liu, S.-J.; et al. Thiosulfate transfer mediated by DsrE/TusA homologs from acidothermophilic sulfur-oxidizing archaeon metallosphaera cuprina. J. Biol. Chem. 2014, 289, 26949–26959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pettinati, I.; Brem, J.; McDonough, M.A.; Schofield, C.J. Crystal structure of human persulfide dioxygenase: Structural basis of ethylmalonic encephalopathy. Hum. Mol. Genet. 2015, 24, 2458–2469. [Google Scholar] [CrossRef] [Green Version]
- Schütz, M.; Maldener, I.; Griesbeck, C.; Hauska, G. Sulfide-Quinone Reductase from Rhodobacter capsulatus: Requirement for growth, periplasmic localization, and extension of gene sequence analysis. J. Bacteriol. 1999, 181, 6516–6523. [Google Scholar] [CrossRef] [Green Version]
- Mangold, S.; Valdés, J.; Holmes, D.; Dopson, M. Sulfur metabolism in the extreme acidophile Acidithiobacillus caldus. Front. Microbiol. 2011, 2, 17. [Google Scholar] [CrossRef] [Green Version]
- Gao, R.; Liu, H.; Xun, L. Cytoplasmic localization of sulfide:quinone oxidoreductase and persulfide dioxygenase of Cupriavidus pinatubonensis JMP134. Appl. Environ. Microbiol. 2017, 83, e01820-17. [Google Scholar] [CrossRef] [Green Version]
- Shen, J.; Keithly, M.E.; Armstrong, R.N.; Higgins, K.A.; Edmonds, K.A.; Giedroc, D.P. Staphylococcus aureus CstB is a novel multidomain persulfide dioxygenase-sulfurtransferase involved in hydrogen sulfide detoxification. Biochemistry 2015, 54, 4542–4554. [Google Scholar] [CrossRef] [Green Version]
- Motl, N.; Skiba, M.A.; Kabil, O.; Smith, J.L.; Banerjee, R. Structural and biochemical analyses indicate that a bacterial persulfide dioxygenase-rhodanese fusion protein functions in sulfur assimilation. J. Biol. Chem. 2017, 292, 14026–14038. [Google Scholar] [CrossRef] [PubMed]
- Kostanjevecki, V.; Brigé, A.; Meyer, T.E.; Cusanovich, M.A.; Guisez, Y.; Beeumen, J.V. A membrane-bound flavocytochrome c -Sulfide dehydrogenase from the purple phototrophic sulfur bacterium Ectothiorhodospira vacuolata. J. Bacteriol. 2000, 182, 3097–3103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dahl, C.; Franz, B.; Hensen, D.; Kesselheim, A.; Zigann, R. Sulfite oxidation in the purple sulfur bacterium Allochromatium vinosum: Identification of SoeABC as a major player and relevance of SoxYZ in the process. Microbiology 2013, 159, 2626–2638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Appia-Ayme, C.; Little, P.J.; Matsumoto, Y.; Leech, A.P.; Berks, B.C. Cytochrome complex essential for photosynthetic oxidation of both thiosulfate and sulfide in Rhodovulum sulfidophilum. J. Bacteriol. 2001, 183, 6107–6118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wodara, C.; Bardischewsky, F.; Friedrich, C.G. Cloning and characterization of sulfite dehydrogenase, two c-type cytochromes, and a flavoprotein of Paracoccus denitrificans GB17: Essential role of sulfite dehydrogenase in lithotrophic sulfur oxidation. J. Bacteriol. 1997, 179, 5014–5023. [Google Scholar] [CrossRef] [Green Version]
- Friedrich, C.G.; Rother, D.; Bardischewsky, F.; Quentmeier, A.; Fischer, J. Oxidation of reduced inorganic sulfur compounds by bacteria: Emergence of a common mechanism? Appl. Environ. Microbiol. 2001, 67, 2873–2882. [Google Scholar] [CrossRef] [Green Version]
- Sauve, V.; Roversi, P.; Leath, K.J.; Garman, E.F.; Antrobus, R.; Lea, S.M.; Berks, B.C. Mechanism for the hydrolysis of a sulfur-sulfur bond based on the crystal structure of the thiosulfohydrolase SoxB. J. Biol. Chem. 2009, 284, 21707–21718. [Google Scholar] [CrossRef] [Green Version]
- Grabarczyk, D.B.; Chappell, P.E.; Johnson, S.; Stelzl, L.S.; Lea, S.M.; Berks, B.C. Structural basis for specificity and promiscuity in a carrier protein/enzyme system from the sulfur cycle. Proc. Natl. Acad. Sci. USA 2015, 112, E7166–E7175. [Google Scholar] [CrossRef] [Green Version]
- Grabarczyk, D.B.; Berks, B.C. Intermediates in the Sox sulfur oxidation pathway are bound to a sulfane conjugate of the carrier protein SoxYZ. PLoS ONE 2017, 12, e0173395. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, W.; Dam, B. Biochemistry and molecular biology of lithotrophic sulfur oxidation by taxonomically and ecologically diverse bacteria and archaea. FEMS Microbiol. Rev. 2009, 33, 999–1043. [Google Scholar] [CrossRef] [Green Version]
- Maki, J.S. Bacterial intracellular sulfur globules: Structure and function. J. Mol. Microbiol. Biotechnol. 2013, 23, 270–280. [Google Scholar] [CrossRef]
- Tanabe, T.S.; Leimkühler, S.; Dahl, C. Chapter seven—The functional diversity of the prokaryotic sulfur carrier protein TusA. In Advances in Microbial Physiology; Poole, R.K., Ed.; Academic Press: Cambridge, MA, USA, 2019; Volume 75, pp. 233–277. [Google Scholar]
- Löffler, M.; Feldhues, J.; Venceslau, S.S.; Kammler, L.; Grein, F.; Pereira, I.A.C.; Dahl, C. DsrL mediates electron transfer between NADH and rDsrAB in Allochromatium vinosum. Environ. Microbiol. 2020, 22, 783–795. [Google Scholar] [CrossRef] [PubMed]
- Koch, T.; Dahl, C. A novel bacterial sulfur oxidation pathway provides a new link between the cycles of organic and inorganic sulfur compounds. ISME J. 2018, 12, 2479–2491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, H.; Gao, M.; Zhou, H.; Skolnick, J. The crystal structure of a tetrahydrofolate-bound dihydrofolate reductase reveals the origin of slow product release. Commun. Biol. 2018, 1, 226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ernst, C.; Kayastha, K.; Koch, T.; Venceslau, S.S.; Pereira, I.A.C.; Demmer, U.; Ermler, U.; Dahl, C. Structural and spectroscopic characterization of a HdrA-like subunit from Hyphomicrobium denitrificans. FEBS J. 2021, 288, 1664–1678. [Google Scholar] [CrossRef] [PubMed]
- Rohwerder, T.; Sand, W. The sulfane sulfur of persulfides is the actual substrate of the sulfur-oxidizing enzymes from Acidithiobacillus and Acidiphilium spp. Microbiology 2003, 149, 1699–1710. [Google Scholar] [CrossRef] [Green Version]
- Wu, W.; Pang, X.; Lin, J.; Liu, X.; Wang, R.; Lin, J.; Chen, L. Discovery of a new subgroup of sulfur dioxygenases and characterization of sulfur dioxygenases in the sulfur metabolic network of Acidithiobacillus caldus. PLoS ONE 2017, 12, e0183668. [Google Scholar] [CrossRef] [Green Version]
- Hensen, D.; Sperling, D.; Trüper, H.G.; Brune, D.C.; Dahl, C. Thiosulphate oxidation in the phototrophic sulphur bacterium Allochromatium vinosum. Mol. Microbiol. 2006, 62, 794–810. [Google Scholar] [CrossRef]
- Denkmann, K.; Grein, F.; Zigann, R.; Siemen, A.; Bergmann, J.; van Helmont, S.; Nicolai, A.; Pereira, I.A.; Dahl, C. Thiosulfate dehydrogenase: A widespread unusual acidophilic c-type cytochrome. Environ. Microbiol. 2012, 14, 2673–2688. [Google Scholar] [CrossRef]
- Brito, J.A.; Denkmann, K.; Pereira, I.A.; Archer, M.; Dahl, C. Thiosulfate dehydrogenase (TsdA) from Allochromatium vinosum: Structural and functional insights into thiosulfate oxidation. J. Biol. Chem. 2015, 290, 9222–9238. [Google Scholar] [CrossRef] [Green Version]
- Grabarczyk, D.B.; Chappell, P.E.; Eisel, B.; Johnson, S.; Lea, S.M.; Berks, B.C. Mechanism of thiosulfate oxidation in the SoxA family of cysteine-ligated cytochromes. J. Biol. Chem. 2015, 290, 9209–9221. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Lin, J.-Q.; Liu, X.-M.; Pang, X.; Zhang, C.-J.; Yang, C.-L.; Gao, X.-Y.; Lin, C.-M.; Li, Y.-Q.; Li, Y.; et al. Sulfur oxidation in the Acidophilic Autotrophic Acidithiobacillus spp. Front. Microbiol. 2019, 9, 3290. [Google Scholar] [CrossRef] [PubMed]
- Dam, B.; Mandal, S.; Ghosh, W.; Das Gupta, S.K.; Roy, P. The S4-intermediate pathway for the oxidation of thiosulfate by the chemolithoautotroph tetrathiobacter kashmirensis and inhibition of tetrathionate oxidation by sulfite. Res. Microbiol. 2007, 158, 330–338. [Google Scholar] [CrossRef] [PubMed]
- Rameez, M.J.; Pyne, P.; Mandal, S.; Chatterjee, S.; Alam, M.; Bhattacharya, S.; Mondal, N.; Sarkar, J.; Ghosh, W. Two pathways for thiosulfate oxidation in the alphaproteobacterial chemolithotroph Paracoccus thiocyanatus SST. Microbiol. Res. 2020, 230, 126345. [Google Scholar] [CrossRef]
- Bugaytsova, Z.; Lindström, E.B. Localization, purification and properties of a tetrathionate hydrolase from Acidithiobacillus caldus. Eur. J. Biochem. 2004, 271, 272–280. [Google Scholar] [CrossRef] [Green Version]
- Kikumoto, M.; Nogami, S.; Kanao, T.; Takada, J.; Kamimura, K. Tetrathionate-forming thiosulfate dehydrogenase from the acidophilic, chemolithoautotrophic bacterium Acidithiobacillus ferrooxidans. Appl. Environ. Microbiol. 2013, 79, 113–120. [Google Scholar] [CrossRef] [Green Version]
- Pyne, P.; Alam, M.; Rameez, M.J.; Mandal, S.; Sar, A.; Mondal, N.; Debnath, U.; Mathew, B.; Misra, A.K.; Mandal, A.K.; et al. Homologs from sulfur oxidation (Sox) and methanol dehydrogenation (Xox) enzyme systems collaborate to give rise to a novel pathway of chemolithotrophic tetrathionate oxidation. Mol. Microbiol. 2018, 109, 169–191. [Google Scholar] [CrossRef] [Green Version]
- Protze, J.; Müller, F.; Lauber, K.; Naß, B.; Mentele, R.; Lottspeich, F.; Kletzin, A. An Extracellular Tetrathionate Hydrolase from the Thermoacidophilic Archaeon Acidianus Ambivalens with an Activity Optimum at pH 1. Front. Microbiol. 2011, 2, 68. [Google Scholar] [CrossRef] [Green Version]
- Feng, C.; Tollin, G.; Enemark, J.H. Sulfite oxidizing enzymes. Biochim. Biophys. Acta 2007, 1774, 527–539. [Google Scholar] [CrossRef]
- Kappler, U. Bacterial sulfite-oxidizing enzymes. Biochim. Biophys. Acta 2011, 1807, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Kappler, U. Bacterial Sulfite-Oxidizing Enzymes—Enzymes for Chemolithotrophs Only. In Microbial Sulfur Metabolism; Dahl, C., Friedrich, C.G., Eds.; Springer: Berlin/Heidelberg, Germany, 2008; pp. 151–169. [Google Scholar]
- Kimura, Y.; Shibuya, N.; Kimura, H. Sulfite protects neurons from oxidative stress. Br. J. Pharmacol 2019, 176, 571–582. [Google Scholar] [CrossRef] [PubMed]
- Maher, P.; Currais, A.; Schubert, D. Using the oxytosis/ferroptosis pathway to understand and treat age-associated neurodegenerative diseases. Cell Chem. Biol. 2020, 27, 1456–1471. [Google Scholar] [CrossRef] [PubMed]
- Kappler, U.; Dahl, C. Enzymology and molecular biology of prokaryotic sulfite oxidation. FEMS Microbiol. Lett. 2001, 203, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Kappler, U.; Enemark, J.H. Sulfite-oxidizing enzymes. J. Biol. Inorg. Chem. 2015, 20, 253–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hille, R.; Hall, J.; Basu, P. The mononuclear molybdenum enzymes. Chem. Rev. 2014, 114, 3963–4038. [Google Scholar] [CrossRef] [Green Version]
- Schrader, N.; Fischer, K.; Theis, K.; Mendel, R.R.; Schwarz, G.; Kisker, C. The crystal structure of plant sulfite oxidase provides insights into sulfite oxidation in plants and animals. Structure 2003, 11, 1251–1263. [Google Scholar] [CrossRef] [Green Version]
- Low, L.; Kilmartin, J.; Bernhardt, P.; Kappler, U. How are “atypical” sulfite dehydrogenases linked to cell metabolism? interactions between the SorT sulfite dehydrogenase and small redox proteins. Front. Microbiol. 2011, 2, 58. [Google Scholar] [CrossRef] [Green Version]
- McGrath, A.P.; Laming, E.L.; Casas Garcia, G.P.; Kvansakul, M.; Guss, J.M.; Trewhella, J.; Calmes, B.; Bernhardt, P.V.; Hanson, G.R.; Kappler, U.; et al. Structural basis of interprotein electron transfer in bacterial sulfite oxidation. eLife 2015, 4, e09066. [Google Scholar] [CrossRef]
- Truong, D.H.; Eghbal, M.A.; Hindmarsh, W.; Roth, S.H.; O’Brien, P.J. Molecular mechanisms of hydrogen sulfide toxicity. Drug Metab. Rev. 2006, 38, 733–744. [Google Scholar] [CrossRef]
- Xu, Z.; Qiu, Z.; Liu, Q.; Huang, Y.; Li, D.; Shen, X.; Fan, K.; Xi, J.; Gu, Y.; Tang, Y.; et al. Converting organosulfur compounds to inorganic polysulfides against resistant bacterial infections. Nat. Commun. 2018, 9, 3713. [Google Scholar] [CrossRef] [Green Version]
- Greiner, R.; Pálinkás, Z.; Bäsell, K.; Becher, D.; Antelmann, H.; Nagy, P.; Dick, T.P. Polysulfides link H2S to protein thiol oxidation. Antioxid. Redox Signal. 2013, 19, 1749–1765. [Google Scholar] [CrossRef] [PubMed]
- Kimura, H. Signaling molecules: Hydrogen sulfide and polysulfide. Antioxid. Redox Signal. 2015, 22, 362–376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iciek, M.; Kowalczyk-Pachel, D.; Bilska-Wilkosz, A.; Kwiecień, I.; Górny, M.; Włodek, L. S-sulfhydration as a cellular redox regulation. Biosci. Rep. 2015, 36, e00304. [Google Scholar] [CrossRef]
- Krishnan, N.; Fu, C.; Pappin, D.J.; Tonks, N.K. H2S-induced sulfhydration of the phosphatase PTP1B and its role in the endoplasmic reticulum stress response. Sci. Signal. 2011, 4, ra86. [Google Scholar] [CrossRef] [Green Version]
- Vandiver, M.S.; Paul, B.D.; Xu, R.; Karuppagounder, S.; Rao, F.; Snowman, A.M.; Seok Ko, H.; Lee, Y.I.; Dawson, V.L.; Dawson, T.M.; et al. Sulfhydration mediates neuroprotective actions of parkin. Nat. Commun. 2013, 4, 1626. [Google Scholar] [CrossRef] [Green Version]
- Ju, Y.; Untereiner, A.; Wu, L.; Yang, G. H2S-induced S-sulfhydration of pyruvate carboxylase contributes to gluconeogenesis in liver cells. Biochim. Biophys. Acta 2015, 1850, 2293–2303. [Google Scholar] [CrossRef]
- Peng, H.; Shen, J.; Edmonds, K.A.; Luebke, J.L.; Hickey, A.K.; Palmer, L.D.; Chang, F.J.; Bruce, K.A.; Kehl-Fie, T.E.; Skaar, E.P.; et al. Sulfide homeostasis and nitroxyl intersect via formation of reactive sulfur species in Staphylococcus aureus. mSphere 2017, 2, e00082-17. [Google Scholar] [CrossRef] [Green Version]
- Borisov, V.B.; Forte, E. Impact of hydrogen sulfide on mitochondrial and bacterial bioenergetics. Int. J. Mol. Sci. 2021, 22, 12688. [Google Scholar] [CrossRef]
- Sousa, F.L.; Alves, R.J.; Ribeiro, M.A.; Pereira-Leal, J.B.; Teixeira, M.; Pereira, M.M. The superfamily of heme–copper oxygen reductases: Types and evolutionary considerations. Biochim. Biophys. Acta 2012, 1817, 629–637. [Google Scholar] [CrossRef] [Green Version]
- Fu, H.; Chen, H.; Wang, J.; Zhou, G.; Zhang, H.; Zhang, L.; Gao, H. Crp-dependent cytochrome bd oxidase confers nitrite resistance to Shewanella oneidensis. Environ. Microbiol. 2013, 15, 2198–2212. [Google Scholar] [CrossRef]
- Zhou, G.; Yin, J.; Chen, H.; Hua, Y.; Sun, L.; Gao, H. Combined effect of loss of the caa3 oxidase and Crp regulation drives Shewanella to thrive in redox-stratified environments. ISME J. 2013, 7, 1752–1763. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Xie, P.; Huang, Y.; Gao, H. Complex interplay of heme-copper oxidases with nitrite and nitric oxide. Int. J. Mol. Sci. 2022, 23, 979. [Google Scholar] [CrossRef]
- Borisov, V.B.; Siletsky, S.A.; Nastasi, M.R.; Forte, E. ROS defense systems and terminal oxidases in bacteria. Antioxidants 2021, 10, 839. [Google Scholar] [CrossRef] [PubMed]
- Nicholls, P.; Marshall, D.C.; Cooper, C.E.; Wilson, M.T. Sulfide inhibition of and metabolism by cytochrome c oxidase. Biochem. Soc. Trans. 2013, 41, 1312–1316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hill, B.C.; Woon, T.C.; Nicholls, P.; Peterson, J.; Greenwood, C.; Thomson, A.J. Interactions of sulphide and other ligands with cytochrome c oxidase. an electron-paramagnetic-resonance study. Biochem. J. 1984, 224, 591–600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forte, E.; Borisov, V.B.; Falabella, M.; Colaço, H.G.; Tinajero-Trejo, M.; Poole, R.K.; Vicente, J.B.; Sarti, P.; Giuffrè, A. The terminal oxidase cytochrome bd promotes sulfide-resistant bacterial respiration and growth. Sci. Rep. 2016, 6, 23788. [Google Scholar] [CrossRef] [Green Version]
- Nicholls, P.; Kim, J.-K. Sulphide as an inhibitor and electron donor for the cytochrome c oxidase system. Can. J. Biochem. 1982, 60, 613–623. [Google Scholar] [CrossRef]
- Bertsova, Y.V.; Bogachev, A.V.; Skulachev, V.P. Generation of protonic potential by the bd-type quinol oxidase of Azotobacter vinelandii. FEBS Lett. 1997, 414, 369–372. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Luo, Q.; Yin, J.; Gao, T.; Gao, H. Evidence for the requirement of CydX in function but not assembly of the cytochrome bd oxidase in Shewanella oneidensis. Biochim. Biophys. Acta 2015, 1850, 318–328. [Google Scholar] [CrossRef]
- Mason, M.G.; Shepherd, M.; Nicholls, P.; Dobbin, P.S.; Dodsworth, K.S.; Poole, R.K.; Cooper, C.E. Cytochrome bd confers nitric oxide resistance to Escherichia coli. Nat. Chem. Biol. 2009, 5, 94–96. [Google Scholar] [CrossRef]
- Giuffrè, A.; Borisov, V.B.; Mastronicola, D.; Sarti, P.; Forte, E. Cytochrome bd oxidase and nitric oxide: From reaction mechanisms to bacterial physiology. FEBS Lett. 2012, 586, 622–629. [Google Scholar] [CrossRef] [PubMed]
- Korshunov, S.; Imlay, K.R.C.; Imlay, J.A. The cytochrome bd oxidase of Escherichia coli prevents respiratory inhibition by endogenous and exogenous hydrogen sulfide. Mol. Microbiol. 2016, 101, 62–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saini, V.; Chinta, K.C.; Reddy, V.P.; Glasgow, J.N.; Stein, A.; Lamprecht, D.A.; Rahman, M.A.; Mackenzie, J.S.; Truebody, B.E.; Adamson, J.H.; et al. Hydrogen sulfide stimulates Mycobacterium tuberculosis respiration, growth and pathogenesis. Nat. Commun. 2020, 11, 557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kunota, T.T.R.; Rahman, M.A.; Truebody, B.E.; Mackenzie, J.S.; Saini, V.; Lamprecht, D.A.; Adamson, J.H.; Sevalkar, R.R.; Lancaster, J.R.; Berney, M.; et al. Mycobacterium tuberculosis H2S functions as a sink to modulate central metabolism, bioenergetics, and drug susceptibility. Antioxidants 2021, 10, 1285. [Google Scholar] [CrossRef]
- Mishra, S.; Imlay, J. Why do bacteria use so many enzymes to scavenge hydrogen peroxide? Arch. Biochem. Biophys. 2012, 525, 145–160. [Google Scholar] [CrossRef] [Green Version]
- Palma, J.M.; Mateos, R.M.; López-Jaramillo, J.; Rodríguez-Ruiz, M.; González-Gordo, S.; Lechuga-Sancho, A.M.; Corpas, F.J. Plant catalases as NO and H2S targets. Redox Biol. 2020, 34, 101525. [Google Scholar] [CrossRef]
- Mantle, D.; Yang, G. Hydrogen sulfide and metal interaction: The pathophysiological implications. Mol. Cell. Biochem. 2022, 477, 2235–2248. [Google Scholar] [CrossRef]
- Shimizu, T. Revisit of the interactions between hydrogen sulfide and heme proteins. Curr. Chem. Biol. 2013, 7, 207–212. [Google Scholar] [CrossRef]
- Jensen, B.; Fago, A. Reactions of ferric hemoglobin and myoglobin with hydrogen sulfide under physiological conditions. J. Inorg. Biochem. 2018, 182, 133–140. [Google Scholar] [CrossRef]
- Fu, L.-H.; Wei, Z.-Z.; Hu, K.-D.; Hu, L.-Y.; Li, Y.-H.; Chen, X.-Y.; Han, Z.; Yao, G.-F.; Zhang, H. Hydrogen sulfide inhibits the growth of Escherichia coli through oxidative damage. J. Microbiol. 2018, 56, 238–245. [Google Scholar] [CrossRef]
- Ng, S.Y.; Ong, K.X.; Surendran, S.T.; Sinha, A.; Lai, J.J.H.; Chen, J.; Liang, J.; Tay, L.K.S.; Cui, L.; Loo, H.L.; et al. Hydrogen sulfide sensitizes Acinetobacter baumannii to killing by antibiotics. Front. Microbiol. 2020, 11, 1875. [Google Scholar] [CrossRef] [PubMed]
- Corpas, F.J.; Barroso, J.B.; González-Gordo, S.; Muñoz-Vargas, M.A.; Palma, J.M. Hydrogen sulfide: A novel component in Arabidopsis peroxisomes which triggers catalase inhibition. J. Integr. Plant Biol. 2019, 61, 871–883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gusarov, I.; Nudler, E. NO-mediated cytoprotection: Instant adaptation to oxidative stress in bacteria. Proc. Natl. Acad. Sci. USA 2005, 102, 13855–13860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gusarov, I.; Shatalin, K.; Starodubtseva, M.; Nudler, E. Endogenous nitric oxide protects bacteria against a wide spectrum of antibiotics. Science 2009, 325, 1380–1384. [Google Scholar] [CrossRef] [Green Version]
- Agashe, P.; Kuzminov, A. Catalase inhibition by nitric oxide potentiates hydrogen peroxide to trigger catastrophic chromosome fragmentation in Escherichia coli. Genetics 2021, 218, iyab057. [Google Scholar] [CrossRef] [PubMed]
- Toliver-Kinsky, T.; Cui, W.; Törö, G.; Lee, S.J.; Shatalin, K.; Nudler, E.; Szabo, C. H2S, a bacterial defense mechanism against the host immune response. Infect. Immun. 2018, 87, e00272-18. [Google Scholar] [PubMed] [Green Version]
- Rahman, M.A.; Cumming, B.M.; Addicott, K.W.; Pacl, H.T.; Russell, S.L.; Nargan, K.; Naidoo, T.; Ramdial, P.K.; Adamson, J.H.; Wang, R.; et al. Hydrogen sulfide dysregulates the immune response by suppressing central carbon metabolism to promote tuberculosis. Proc. Natl. Acad. Sci. USA 2020, 117, 6663–6674. [Google Scholar] [CrossRef] [Green Version]
- Shatalin, K.; Nuthanakanti, A.; Kaushik, A.; Shishov, D.; Peselis, A.; Shamovsky, I.; Pani, B.; Lechpammer, M.; Vasilyev, N.; Shatalina, E.; et al. Inhibitors of bacterial H2S biogenesis targeting antibiotic resistance and tolerance. Science 2021, 372, 1169–1175. [Google Scholar] [CrossRef]
- Ivanovic-Burmazovic, I.; Filipovic, M.R. Saying NO to H2S: A story of HNO, HSNO, and SSNO–. Inorg. Chem. 2019, 58, 4039–4051. [Google Scholar] [CrossRef] [Green Version]
- Paul, B.D.; Snyder, S.H. H2S signalling through protein sulfhydration and beyond. Nat. Rev. Mol. Cell Biol. 2012, 13, 499–507. [Google Scholar] [CrossRef]
- Filipovic, M.R.; Miljkovic, J.L.; Nauser, T.; Royzen, M.; Klos, K.; Shubina, T.; Koppenol, W.H.; Lippard, S.J.; Ivanović-Burmazović, I. Chemical characterization of the smallest S-nitrosothiol, HSNO; cellular cross-talk of H2S and S-nitrosothiols. J. Am. Chem. Soc. 2012, 134, 12016–12027. [Google Scholar] [CrossRef] [PubMed]
- Cortese-Krott, M.M.; Kuhnle, G.G.C.; Dyson, A.; Fernandez, B.O.; Grman, M.; DuMond, J.F.; Barrow, M.P.; McLeod, G.; Nakagawa, H.; Ondrias, K.; et al. Key bioactive reaction products of the NO/H2S interaction are S/N-hybrid species, polysulfides, and nitroxyl. Proc. Natl. Acad. Sci. USA 2015, 112, E4651–E4660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bailey, T.S.; Henthorn, H.A.; Pluth, M.D. The intersection of NO and H2S: Persulfides generate NO from nitrite through polysulfide formation. Inorg. Chem. 2016, 55, 12618–12625. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, R.; Koike, S.; Takano, Y.; Shibuya, N.; Kimura, Y.; Hanaoka, K.; Urano, Y.; Ogasawara, Y.; Kimura, H. Polysulfides (H2Sn) produced from the interaction of hydrogen sulfide (H2S) and nitric oxide (NO) activate TRPA1 channels. Sci. Rep. 2017, 7, 45995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, T.; Ono, K.; Tsutsuki, H.; Ihara, H.; Islam, W.; Akaike, T.; Sawa, T. Enhanced cellular polysulfides negatively regulate TLR4 signaling and mitigate lethal endotoxin shock. Cell Chem. Biol. 2019, 26, 686–698.e4. [Google Scholar] [CrossRef] [PubMed]
- Peng, H.; Zhang, Y.; Palmer, L.D.; Kehl-Fie, T.E.; Skaar, E.P.; Trinidad, J.C.; Giedroc, D.P. Hydrogen sulfide and reactive sulfur species impact proteome S-sulfhydration and global virulence regulation in Staphylococcus aureus. ACS Infect. Dis. 2017, 3, 744–755. [Google Scholar] [CrossRef] [Green Version]
- Shen, J.; Walsh, B.J.C.; Flores-Mireles, A.L.; Peng, H.; Zhang, Y.; Zhang, Y.; Trinidad, J.C.; Hultgren, S.J.; Giedroc, D.P. Hydrogen sulfide sensing through reactive sulfur species (RSS) and nitroxyl (HNO) in Enterococcus faecalis. ACS Chem. Biol. 2018, 13, 1610–1620. [Google Scholar] [CrossRef]
- Shi, L.; Dong, H.; Reguera, G.; Beyenal, H.; Lu, A.; Liu, J.; Yu, H.-Q.; Fredrickson, J.K. Extracellular electron transfer mechanisms between microorganisms and minerals. Nat. Rev. Microbiol. 2016, 14, 651–662. [Google Scholar] [CrossRef]
- Lovley, D.R.; Holmes, D.E. Electromicrobiology: The ecophysiology of phylogenetically diverse electroactive microorganisms. Nat. Rev. Microbiol. 2022, 20, 5–19. [Google Scholar] [CrossRef]
- Fredrickson, J.K.; Romine, M.F.; Beliaev, A.S.; Auchtung, J.M.; Driscoll, M.E.; Gardner, T.S.; Nealson, K.H.; Osterman, A.L.; Pinchuk, G.; Reed, J.L.; et al. Towards environmental systems biology of Shewanella. Nat. Rev. Microbiol. 2008, 6, 592–603. [Google Scholar] [CrossRef]
- Zhao, J.; Li, F.; Cao, Y.; Zhang, X.; Chen, T.; Song, H.; Wang, Z. Microbial extracellular electron transfer and strategies for engineering electroactive microorganisms. Biotechnol. Adv. 2021, 53, 107682. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.; Srikanth, V.; Salazar-Morales, A.I.; Jain, R.; O’Brien, J.P.; Yi, S.M.; Soni, R.K.; Samatey, F.A.; Yalcin, S.E.; Malvankar, N.S. Structure of Geobacter pili reveals secretory rather than nanowire behaviour. Nature 2021, 597, 430–434. [Google Scholar] [CrossRef] [PubMed]
- Myers, C.R.; Nealson, K.H. Bacterial manganese reduction and growth with manganese oxide as the sole electron acceptor. Science 1988, 240, 1319–1321. [Google Scholar] [CrossRef] [PubMed]
- Canfield, D.E.; Stewart, F.J.; Thamdrup, B.; De Brabandere, L.; Dalsgaard, T.; Delong, E.F.; Revsbech, N.P.; Ulloa, O. A cryptic sulfur cycle in oxygen-minimum-zone waters off the chilean coast. Science 2010, 330, 1375–1378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osorio, H.; Mangold, S.; Denis, Y.; Ñancucheo, I.; Esparza, M.; Johnson, D.B.; Bonnefoy, V.; Dopson, M.; Holmes, D.S. Anaerobic sulfur metabolism coupled to dissimilatory iron reduction in the extremophile Acidithiobacillus ferrooxidans. Appl. Environ. Microbiol. 2013, 79, 2172–2181. [Google Scholar] [CrossRef] [Green Version]
- Flynn, T.M.; O’Loughlin, E.J.; Mishra, B.; DiChristina, T.J.; Kemner, K.M. Sulfur-mediated electron shuttling during bacterial iron reduction. Science 2014, 344, 1039–1042. [Google Scholar] [CrossRef]
- Hansel, C.M.; Lentini, C.J.; Tang, Y.; Johnston, D.T.; Wankel, S.D.; Jardine, P.M. Dominance of sulfur-fueled iron oxide reduction in low-sulfate freshwater sediments. ISME J. 2015, 9, 2400–2412. [Google Scholar] [CrossRef] [PubMed]
- Poser, A.; Lohmayer, R.; Vogt, C.; Knoeller, K.; Planer-Friedrich, B.; Sorokin, D.; Richnow, H.-H.; Finster, K. Disproportionation of elemental sulfur by haloalkaliphilic bacteria from soda lakes. Extremophiles 2013, 17, 1003–1012. [Google Scholar] [CrossRef] [PubMed]
- Diez, C.M.P.; Juncos, M.J.; Dujovne, M.V.; Capdevila, D.A. Bacterial transcriptional regulators: A road map for functional, structural, and biophysical characterization. Int. J. Mol. Sci. 2022, 23, 2179. [Google Scholar] [CrossRef]
- Peng, H.; Zhang, Y.; Trinidad, J.C.; Giedroc, D.P. Thioredoxin profiling of multiple thioredoxin-like proteins in Staphylococcus aureus. Front. Microbiol. 2018, 9, 2385. [Google Scholar] [CrossRef]
- Walsh, B.J.C.; Wang, J.; Edmonds, K.A.; Palmer, L.D.; Zhang, Y.; Trinidad, J.C.; Skaar, E.P.; Giedroc, D.P. The response of Acinetobacter baumannii to hydrogen sulfide reveals two independent persulfide-sensing systems and a connection to biofilm regulation. mBio 2020, 11, e01254-20. [Google Scholar] [CrossRef] [PubMed]
- Luebke, J.L.; Shen, J.; Bruce, K.E.; Kehl-Fie, T.E.; Peng, H.; Skaar, E.P.; Giedroc, D.P. The CsoR-like sulfurtransferase repressor (CstR) is a persulfide sensor in Staphylococcus aureus. Mol. Microbiol. 2014, 94, 1343–1360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grossoehme, N.; Kehl-Fie, T.E.; Ma, Z.; Adams, K.W.; Cowart, D.M.; Scott, R.A.; Skaar, E.P.; Giedroc, D.P. Control of copper resistance and inorganic sulfur metabolism by paralogous regulators in Staphylococcus aureus. J. Biol. Chem. 2011, 286, 13522–13531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, T.; Ramesh, A.; Ma, Z.; Ward, S.K.; Zhang, L.; George, G.N.; Talaat, A.M.; Sacchettini, J.C.; Giedroc, D.P. CsoR is a novel Mycobacterium tuberculosis copper-sensing transcriptional regulator. Nat. Chem. Biol. 2007, 3, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Bauchart-Thevret, C.; Stoll, B.; Burrin, D.G. Intestinal metabolism of sulfur amino acids. Nutr. Res. Rev. 2009, 22, 175–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riedijk, M.A.; Stoll, B.; Chacko, S.; Schierbeek, H.; Sunehag, A.L.; van Goudoever, J.B.; Burrin, D.G. Methionine transmethylation and transsulfuration in the piglet gastrointestinal tract. Proc. Natl. Acad. Sci. USA 2007, 104, 3408–3413. [Google Scholar] [CrossRef] [Green Version]
- Busenlehner, L.S.; Pennella, M.A.; Giedroc, D.P. The SmtB/ArsR family of metalloregulatory transcriptional repressors: Structural insights into prokaryotic metal resistance. FEMS Microbiol. Rev. 2003, 27, 131–143. [Google Scholar] [CrossRef] [Green Version]
- Shimizu, T.; Shen, J.; Fang, M.; Zhang, Y.; Hori, K.; Trinidad, J.C.; Bauer, C.E.; Giedroc, D.P.; Masuda, S. Sulfide-responsive transcriptional repressor SqrR functions as a master regulator of sulfide-dependent photosynthesis. Proc. Natl. Acad. Sci. USA 2017, 114, 2355–2360. [Google Scholar] [CrossRef] [Green Version]
- Shimizu, T.; Masuda, S. Persulphide-responsive transcriptional regulation and metabolism in bacteria. J. Biochem. 2020, 167, 125–132. [Google Scholar] [CrossRef]
- de Lira, N.P.V.; Pauletti, B.A.; Marques, A.C.; Perez, C.A.; Caserta, R.; de Souza, A.A.; Vercesi, A.E.; Paes Leme, A.F.; Benedetti, C.E. BigR is a sulfide sensor that regulates a sulfur transferase/dioxygenase required for aerobic respiration of plant bacteria under sulfide stress. Sci. Rep. 2018, 8, 3508. [Google Scholar] [CrossRef] [Green Version]
- Barbosa, R.L.; Benedetti, C.E. BigR, a transcriptional repressor from plant-associated bacteria, regulates an operon implicated in biofilm growth. J. Bacteriol. 2007, 189, 6185–6194. [Google Scholar] [CrossRef]
- Guimaraes, B.G.; Barbosa, R.L.; Soprano, A.S.; Campos, B.M.; de Souza, T.A.; Tonoli, C.C.; Leme, A.F.; Murakami, M.T.; Benedetti, C.E. Plant pathogenic bacteria utilize biofilm growth-associated repressor (BigR), a novel winged-helix redox switch, to control hydrogen sulfide detoxification under hypoxia. J. Biol. Chem. 2011, 286, 26148–26157. [Google Scholar] [CrossRef] [Green Version]
- Gristwood, T.; McNeil, M.B.; Clulow, J.S.; Salmond, G.P.; Fineran, P.C. PigS and PigP regulate prodigiosin biosynthesis in Serratia via differential control of divergent operons, which include predicted transporters of sulfur-containing molecules. J. Bacteriol. 2011, 193, 1076–1085. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Li, J.; Lu, C.; Xia, Y.; Xin, Y.; Liu, H.; Xun, L.; Liu, H. FisR activates sigma(54) -dependent transcription of sulfide-oxidizing genes in Cupriavidus pinatubonensis JMP134. Mol. Microbiol. 2017, 105, 373–384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benchoam, D.; Cuevasanta, E.; Moller, M.N.; Alvarez, B. Hydrogen sulfide and persulfides oxidation by biologically relevant oxidizing species. Antioxidants 2019, 8, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ezerina, D.; Takano, Y.; Hanaoka, K.; Urano, Y.; Dick, T.P. N-acetyl cysteine functions as a fast-acting antioxidant by triggering intracellular H2S and sulfane sulfur production. Cell Chem. Biol. 2018, 25, 447–459.e4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xuan, G.; Lv, C.; Xu, H.; Li, K.; Liu, H.; Xia, Y.; Xun, L. Sulfane sulfur regulates LasR-mediated quorum sensing and virulence in Pseudomonas aeruginosa PAO1. Antioxidants 2021, 10, 1498. [Google Scholar] [CrossRef]
- Xu, H.; Xuan, G.; Liu, H.; Liu, H.; Xia, Y.; Xun, L. Sulfane sulfur is an intrinsic signal for the organic peroxide sensor OhrR of Pseudomonas aeruginosa. Antioxidants 2022, 11, 1667. [Google Scholar] [CrossRef]
Figure 1.
Structures of some RSS chemotypes in different views. The red (−2), yellow (−1), gray (0), purple (+1), green (+4), light blue (+5) and dark blue (+6) rectangles are used to designate the valence states of sulfur, as specified.
Figure 1.
Structures of some RSS chemotypes in different views. The red (−2), yellow (−1), gray (0), purple (+1), green (+4), light blue (+5) and dark blue (+6) rectangles are used to designate the valence states of sulfur, as specified.
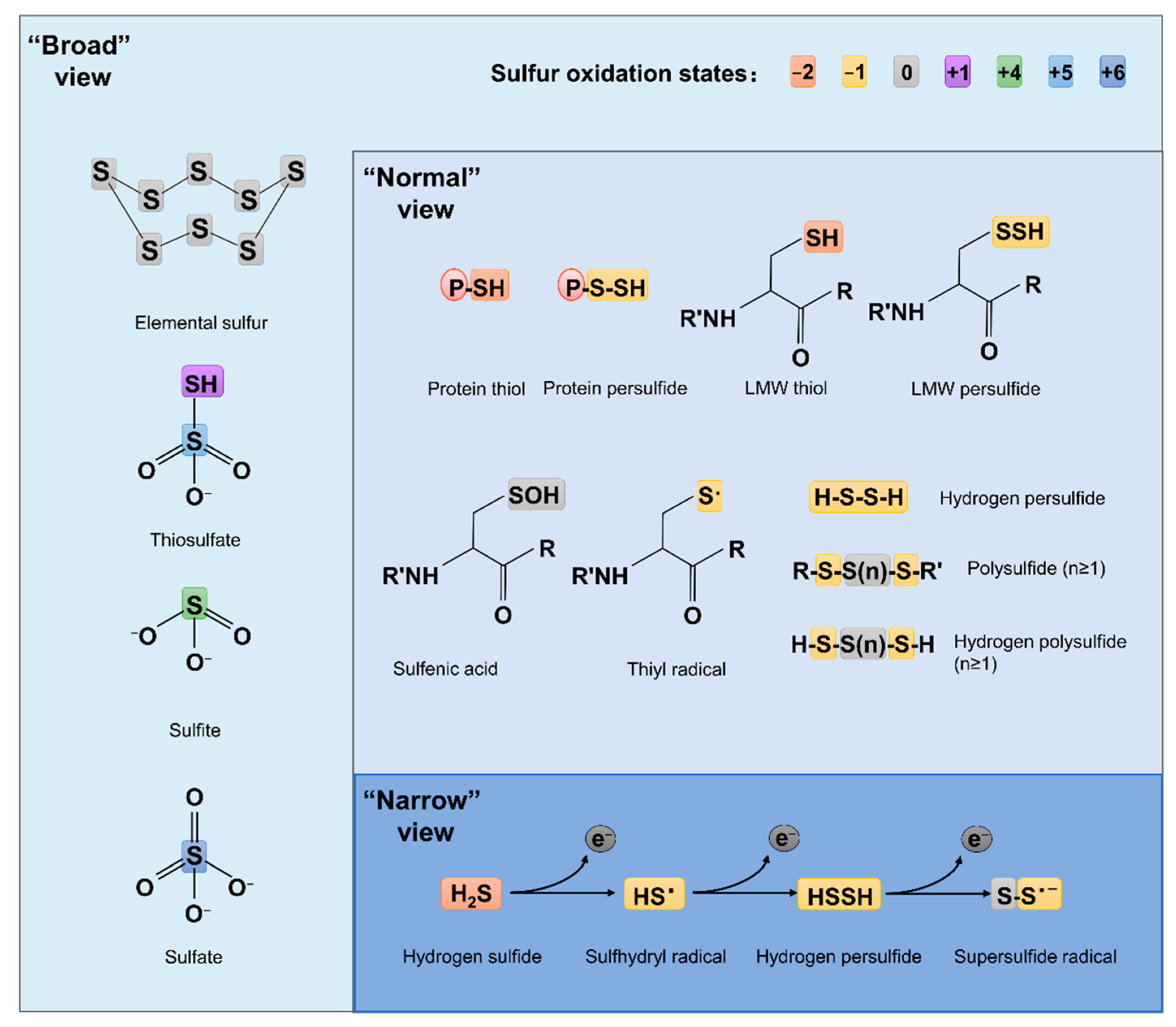
Figure 2.
Pathways for bacterial sulfur metabolism in the cytoplasm. H2S biogenesis through amino acid metabolism: generation of H2S from homocys can be catalyzed by cystathionine β-synthase (CBS) and cystathionine γ-lyase (CSE). Cys aminotransferase (CAT) catalyzes the formation of 3-mercaptopyruvate (3-MP) from cys, and then, mercaptopyruvate sulfurtransferase (MST) converts 3-MP to H2S. H2S biogenesis occurs through assimilatory sulfate reduction (ASR) and dissimilatory sulfate reduction (DSR) of inorganic sulfur species; the latter only occurs in sulfate-reducing bacteria (SRB), catalyzing SO32− to HS− through dissimilatory sulfite reductase (Dsr). For ASR pathway, SO42−, which imported from ATP-dependent transporter CysUWA, is catalyzed and converted to HS− by a series of enzymes, including Sat, CysC, CysH, and CysIJ. S2O32− can also be reduced and used to synthesize cys by CysM and NrdH/Grx1. H2S catabolism: H2S binds to sulfur quinone oxidoreductase (SQR), and goes through a series of sequential reactions, including oxidation of sulfide to polysulfide by membrane-bound SQR, formation of GSSH from reaction involving with rhodanese (Rhd), oxidation of the sulfane sulfur in GSSH to SO32− catalyzed by persulfide dioxygenase (PDO), formation of S2O32− from spontaneous reaction between polysulfide and SO32−, and formation of SO42− either spontaneously or catalyzed by various enzymes, transferring two electrons via quinone into the electron transport chain.
Figure 2.
Pathways for bacterial sulfur metabolism in the cytoplasm. H2S biogenesis through amino acid metabolism: generation of H2S from homocys can be catalyzed by cystathionine β-synthase (CBS) and cystathionine γ-lyase (CSE). Cys aminotransferase (CAT) catalyzes the formation of 3-mercaptopyruvate (3-MP) from cys, and then, mercaptopyruvate sulfurtransferase (MST) converts 3-MP to H2S. H2S biogenesis occurs through assimilatory sulfate reduction (ASR) and dissimilatory sulfate reduction (DSR) of inorganic sulfur species; the latter only occurs in sulfate-reducing bacteria (SRB), catalyzing SO32− to HS− through dissimilatory sulfite reductase (Dsr). For ASR pathway, SO42−, which imported from ATP-dependent transporter CysUWA, is catalyzed and converted to HS− by a series of enzymes, including Sat, CysC, CysH, and CysIJ. S2O32− can also be reduced and used to synthesize cys by CysM and NrdH/Grx1. H2S catabolism: H2S binds to sulfur quinone oxidoreductase (SQR), and goes through a series of sequential reactions, including oxidation of sulfide to polysulfide by membrane-bound SQR, formation of GSSH from reaction involving with rhodanese (Rhd), oxidation of the sulfane sulfur in GSSH to SO32− catalyzed by persulfide dioxygenase (PDO), formation of S2O32− from spontaneous reaction between polysulfide and SO32−, and formation of SO42− either spontaneously or catalyzed by various enzymes, transferring two electrons via quinone into the electron transport chain.
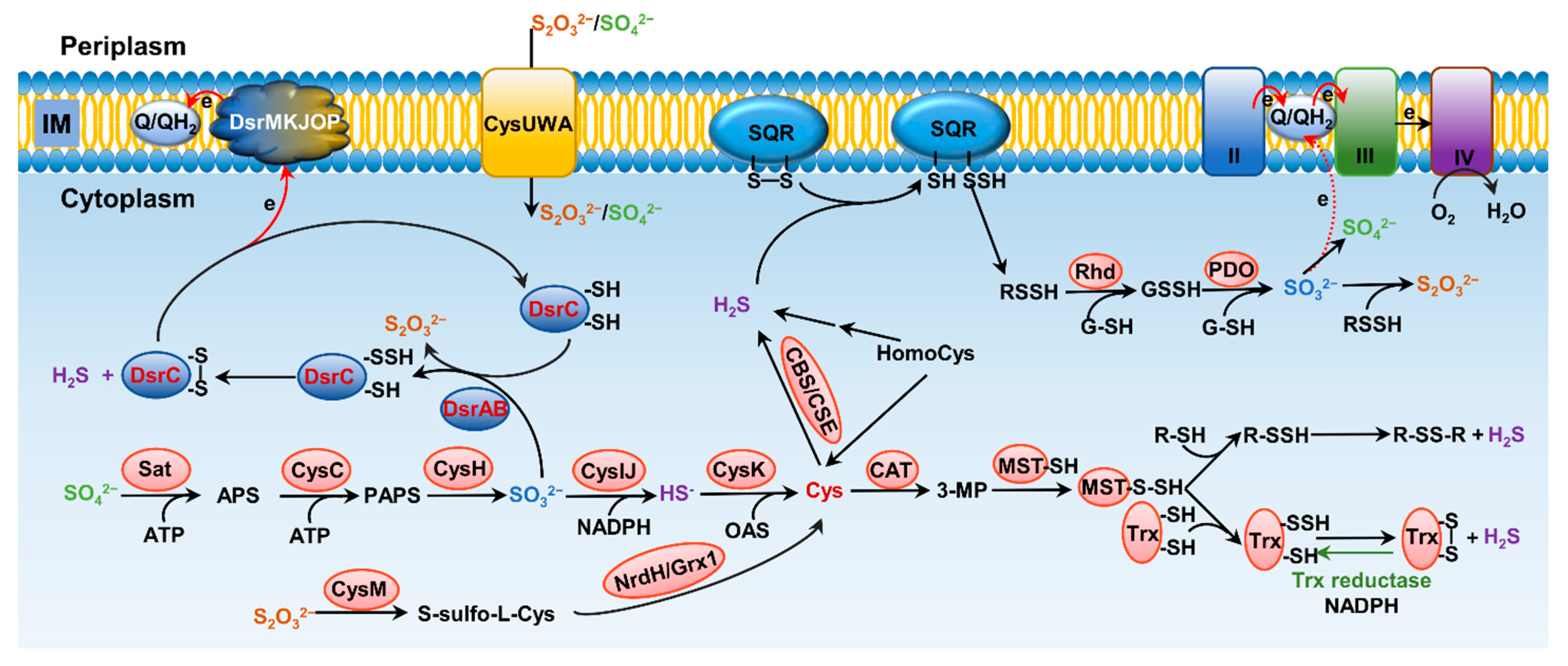
Figure 3.
Structure of representative enzymes involved in transformation of sulfur compounds in bacteria. (A) Overall structure of the Dsr A2B2 heterotetramer of Desulfovibrio vulgaris (PDB ID: 2V4J), green; Archaeoglobus fulgidus (PDB ID: 3MM5), yellow; Desulfovibrio gigas (PDB ID: 3OR1), pink; and Desulfoicrobrium norvegicum (PDB ID: 2XSJ), blue. Heme ligands are shown in ball-stick model. (B) Structure of TsdA of Marichromatium purpuratum (PDB ID: 5LO9; violet) and of Allochromatium vinosum (PDB ID: 4WQ7; orange). Heme ligands are shown in ball-stick model. Expanded regions show the ligands to hemes in active centers. (C) TsdA of Campylobacter jejuni (orange, prepared from AlaphaFold database [90,91]) is aligned and superimposed onto TsdAB of M. purpuratum (PDB ID: 5LO9), which are in violet and yellow, respectively. Heme ligands are shown in ball-stick model. Expanded region shows the ligands to heme 2. (D) Structure of the SoxXA (PDB ID: 2C1D) (left) and SoxYZ-B (PDB ID: 4UWQ) (right) complexes. Heme ligands are shown in ball-stick model. Expanded region shows the active site positioning at the substrate channel of SoxB.
Figure 3.
Structure of representative enzymes involved in transformation of sulfur compounds in bacteria. (A) Overall structure of the Dsr A2B2 heterotetramer of Desulfovibrio vulgaris (PDB ID: 2V4J), green; Archaeoglobus fulgidus (PDB ID: 3MM5), yellow; Desulfovibrio gigas (PDB ID: 3OR1), pink; and Desulfoicrobrium norvegicum (PDB ID: 2XSJ), blue. Heme ligands are shown in ball-stick model. (B) Structure of TsdA of Marichromatium purpuratum (PDB ID: 5LO9; violet) and of Allochromatium vinosum (PDB ID: 4WQ7; orange). Heme ligands are shown in ball-stick model. Expanded regions show the ligands to hemes in active centers. (C) TsdA of Campylobacter jejuni (orange, prepared from AlaphaFold database [90,91]) is aligned and superimposed onto TsdAB of M. purpuratum (PDB ID: 5LO9), which are in violet and yellow, respectively. Heme ligands are shown in ball-stick model. Expanded region shows the ligands to heme 2. (D) Structure of the SoxXA (PDB ID: 2C1D) (left) and SoxYZ-B (PDB ID: 4UWQ) (right) complexes. Heme ligands are shown in ball-stick model. Expanded region shows the active site positioning at the substrate channel of SoxB.

Figure 4.
Pathways for bacterial sulfur transformation in the periplasm. Oxidation of S2− and S0: Flavocyt c sulfide dehydrogenases (FCSDs) can oxidize H2S to the final product polysulfide. In dissimilatory sulfur-oxidizing bacteria, Rhds and PDOs are located in the periplasm, and the oxidation of H2S is catalyzed by SQRs, which consistently expose the reaction to the periplasm space. Oxidation of S0 and S2+: Unconjugated SoxYZ is catalyzed by SoxAX with S2O32−, generating SoxYZ-S-S-SO3−, which is subsequently converted to SoxYZ-S-S−, releasing one molecular of SO42− under the catalysis of SoxB. Reduction of S2+: TtrABC and TsdAB are responsible for interconversion of S4O62− and S2O32−. Reduction of S0: PhsABC and PsrABC are supposed to participate in the reduction of S0 (Sn2−) and S0 (S2O32−) to HS−.
Figure 4.
Pathways for bacterial sulfur transformation in the periplasm. Oxidation of S2− and S0: Flavocyt c sulfide dehydrogenases (FCSDs) can oxidize H2S to the final product polysulfide. In dissimilatory sulfur-oxidizing bacteria, Rhds and PDOs are located in the periplasm, and the oxidation of H2S is catalyzed by SQRs, which consistently expose the reaction to the periplasm space. Oxidation of S0 and S2+: Unconjugated SoxYZ is catalyzed by SoxAX with S2O32−, generating SoxYZ-S-S-SO3−, which is subsequently converted to SoxYZ-S-S−, releasing one molecular of SO42− under the catalysis of SoxB. Reduction of S2+: TtrABC and TsdAB are responsible for interconversion of S4O62− and S2O32−. Reduction of S0: PhsABC and PsrABC are supposed to participate in the reduction of S0 (Sn2−) and S0 (S2O32−) to HS−.

Figure 5.
Physiological impacts of hydrogen sulfide. (A) H2S in oxidative stress. Upon oxidative stress, macromolecules such as DNA and proteins are damaged primarily by •OH, which is generated from the interaction of Fe2+ and H2O2. H2S is a strong inhibitor of hemoproteins, catalase (CAT) in particular. With CAT inhibited, cells are unable to promptly decompose H2O2, leading to increased sensitivity to H2O2 killing. However, the prolonged presence of H2O2 activates OxyR, the master regulator in response to oxidative stress, which in turn induces expression of genes under its control, including CAT. As a result, cells gain an increased resistance against H2O2. In addition, it has been suggested that H2S from endogenous and exogenous sources may offer protection against oxidative stress by sequestering free Fe2+ intracellularly. (B) H2S in metal reduction. Microbial reduction of SO42− and elemental S0 to S2−, which catalyzes abiotic reduction of Fe3+ of Fe(III)oxide and forms FeS with the resulting Fe2+.
Figure 5.
Physiological impacts of hydrogen sulfide. (A) H2S in oxidative stress. Upon oxidative stress, macromolecules such as DNA and proteins are damaged primarily by •OH, which is generated from the interaction of Fe2+ and H2O2. H2S is a strong inhibitor of hemoproteins, catalase (CAT) in particular. With CAT inhibited, cells are unable to promptly decompose H2O2, leading to increased sensitivity to H2O2 killing. However, the prolonged presence of H2O2 activates OxyR, the master regulator in response to oxidative stress, which in turn induces expression of genes under its control, including CAT. As a result, cells gain an increased resistance against H2O2. In addition, it has been suggested that H2S from endogenous and exogenous sources may offer protection against oxidative stress by sequestering free Fe2+ intracellularly. (B) H2S in metal reduction. Microbial reduction of SO42− and elemental S0 to S2−, which catalyzes abiotic reduction of Fe3+ of Fe(III)oxide and forms FeS with the resulting Fe2+.

Figure 6.
Structures and responding mechanism of CstR to RSS. Left, CstR (PDB ID: 7MQ2), which exists as homotetramer. Right, SqrR (PDB: 6O8N), which exists as homodimer. Expanded regions show the mechanism of activation. Both regulators use multiple cys thiols for sensing RSS by forming di-, tri-, and tetra-sulfide bonds.
Figure 6.
Structures and responding mechanism of CstR to RSS. Left, CstR (PDB ID: 7MQ2), which exists as homotetramer. Right, SqrR (PDB: 6O8N), which exists as homodimer. Expanded regions show the mechanism of activation. Both regulators use multiple cys thiols for sensing RSS by forming di-, tri-, and tetra-sulfide bonds.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Han, S.; Li, Y.; Gao, H. Generation and Physiology of Hydrogen Sulfide and Reactive Sulfur Species in Bacteria. Antioxidants 2022, 11, 2487. https://doi.org/10.3390/antiox11122487
AMA Style
Han S, Li Y, Gao H. Generation and Physiology of Hydrogen Sulfide and Reactive Sulfur Species in Bacteria. Antioxidants. 2022; 11(12):2487. https://doi.org/10.3390/antiox11122487
Chicago/Turabian StyleHan, Sirui, Yingxi Li, and Haichun Gao. 2022. "Generation and Physiology of Hydrogen Sulfide and Reactive Sulfur Species in Bacteria" Antioxidants 11, no. 12: 2487. https://doi.org/10.3390/antiox11122487
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.