Long-Term Environmental Monitoring in an Arctic Lake Polluted by Metals under Climate Change

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
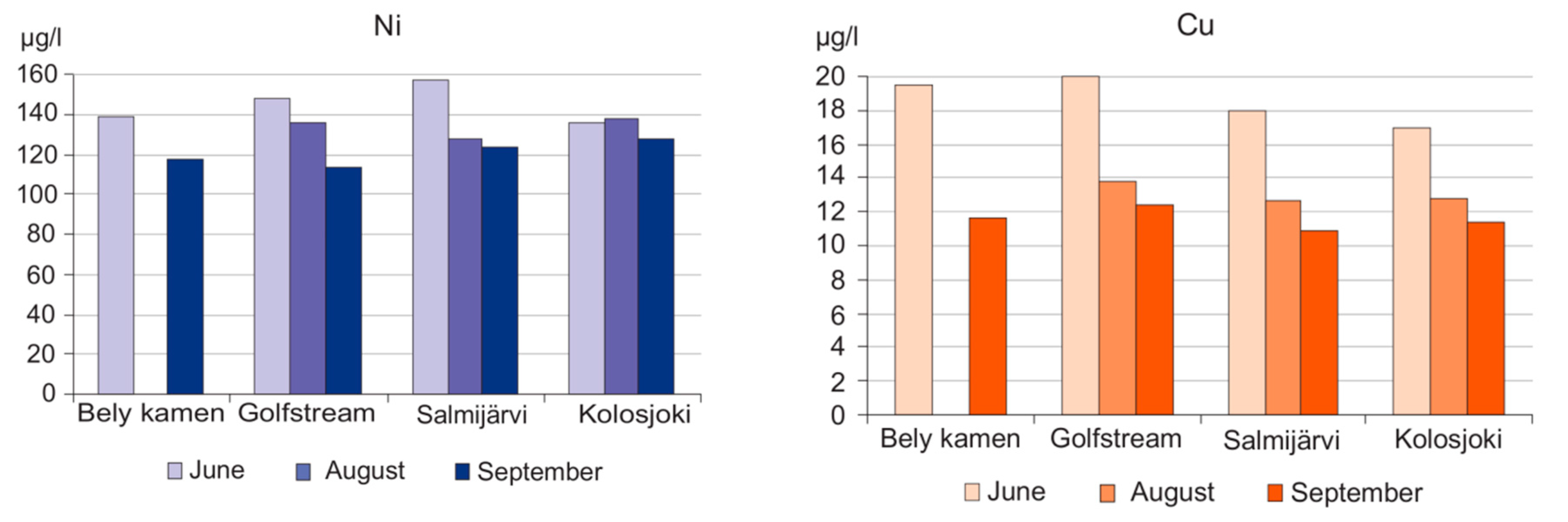
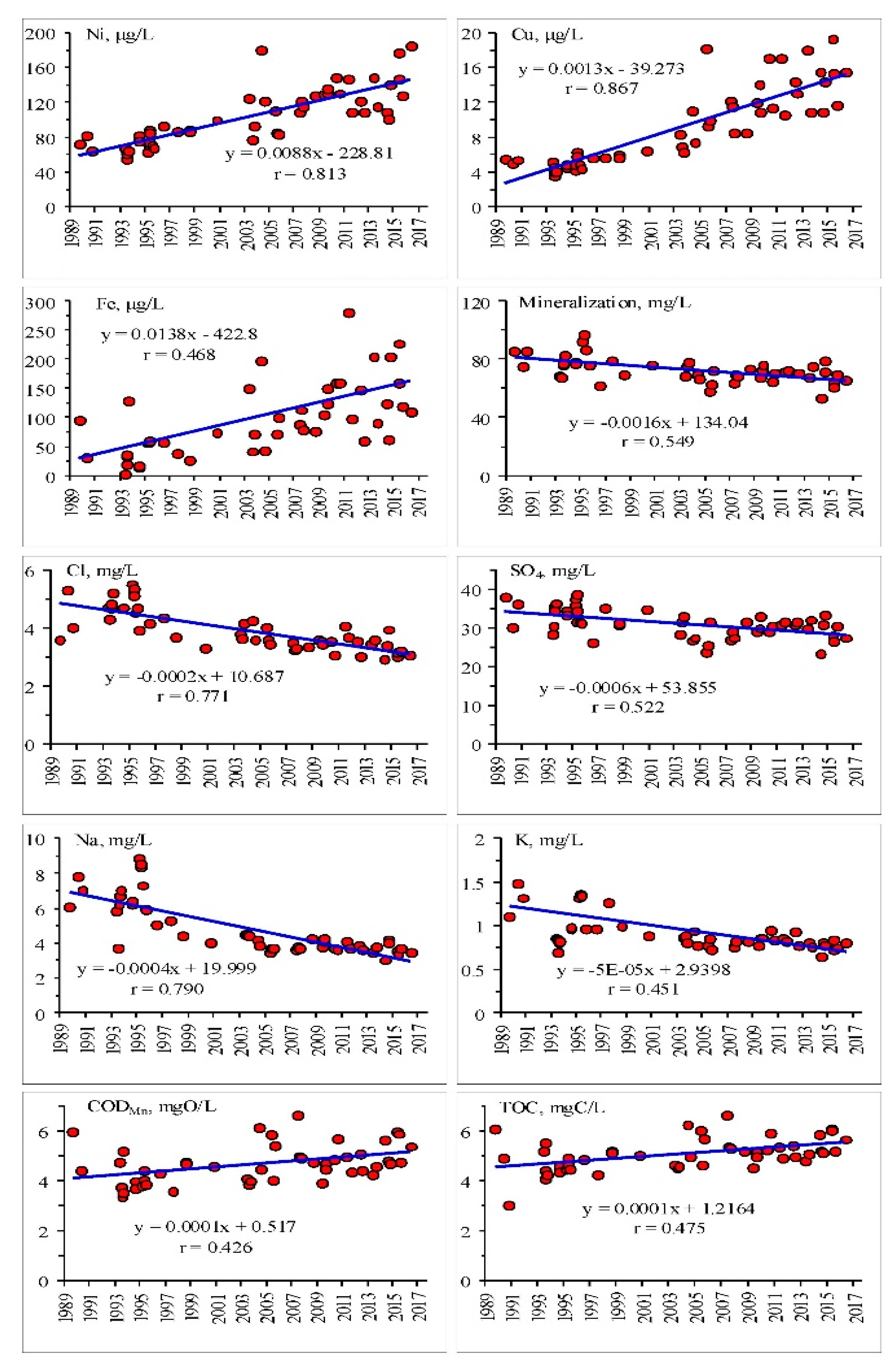
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Dauvalter, V.A.; Kashulin, N.A. Assessment of the ecological state of the Arctic freshwater system based on studies of heavy metals in bottom sediments. Geohimiya 2018, 8, 805–819. [Google Scholar]
- Kashulin, N.A. Fishes of Small Lakes of Northern Fennoscandia in Conditions of Aerotechnogenic Pollution; Izdatelstvo Kolskogo Nauchnogo Tsentra RAN: Apatity, Russia, 2004; 134p. [Google Scholar]
- Kashulin, N.A.; Dauvalter, V.A.; Kashulina, T.G.; Sandimirov, S.S.; Ratkin, N.E.; Kudryavtseva, L.P.; Koroleva, I.M.; Vandysh, O.I.; Mokrotovarova, O.I. Anthropogenic Changes of Lothic Ecosystems in the Murmansk Region; 1: Kovdor Area; Izdatelstvo Kolskogo Nauchnogo Tsentra RAN: Apatity, Russia, 2005; 234p. [Google Scholar]
- Kashulin, N.A.; Dauvalter, V.A.; Sandimirov, S.S.; Ratkin, N.E.; Terentev, P.M.; Koroleva, I.M.; Vandysh, O.I.; Kudryavtseva, L.P. Anthropogenic Changes of Lothic Ecosystems in the Murmansk Region; 2: The Chuna Lake-river System under Conditions of Air Pollution; Izdatelstvo Kolskogo Nauchnogo Tsentra RAN: Apatity, Russia, 2007; 238p. [Google Scholar]
- Moiseenko, T.I.; Dauvalter, V.A.; Lukin, A.A.; Kudryavtseva, L.P.; Ilyashuk, B.P.; Ilyashuk, E.A.; Sandimirov, S.S.; Kagan, L.A.; Vandysh, O.I.; Sharov, A.N.; et al. Anthropogenic Modifications of the Imandra Lake Ecosystem; Nauka: Moscow, Russia, 2002; 398p. [Google Scholar]
- Kashulin, N.A.; Ratkin, N.E.; Dauvalter, V.A.; Lukin, A.A. Impact of airborne pollution on the drainage area of subarctic. Chemosphere 2001, 42, 51–59. [Google Scholar] [CrossRef]
- Ylikörkkö, J.; Christensen, G.N.; Kashulin, N.; Denisov, D.; Andersen, H.J.; Jelkänen, E. Environmental Challenges in the Joint Border Area, Reports 41; Centre for Economic Development, Transport and the Environment for Lapland: Kokkola, Finland, 2015; 165p. [Google Scholar]
- Amundsen, P.A.; Staldvik, F.; Lukin, A.; Kashulin, N.; Reshetnikov, Y.S.; Popova, O. Ecology and Heavy Metal Contaminations in the Fish Communities of the Pasvik River System; Report; Norwegian College of Fishery Science, University of Tromsø: Tromso, Norway, 1993; 29p. [Google Scholar]
- Amundsen, P.A.; Staldvik, F.J.; Lukin, A.A.; Kashulin, N.A.; Popova, O.A.; Reshetnikov, Y.S. Heavy metal contamination in freshwater fish from the border region between Norway and Russia. Sci. Total Environ. 1997, 201, 211–224. [Google Scholar] [CrossRef]
- Lukin, A.A.; Dauvalter, V.A.; Kashulin, N.A.; Yakovlev, V.A.; Sharov, A.N.; Vandysh, O.I. Assessment of copper-nickel industry impact on a subarctic lake ecosystem. Sci. Total Environ. 2003, 306, 73–83. [Google Scholar] [CrossRef]
- Nøst, T.; Kashulin, N.; Schartau, A.K.L.; Lukin, A.; Berger, H.M.; Sharov, A. Impacts of Pollution on Freshwater Communities in the Border Region between Russia and Norway. III. Monitoring Lakes, 1990–1996; NINA Fagrapport: Tromsø, Norway, 1997; pp. 1–37. [Google Scholar]
- Kashulin, N.A.; Lukin, A.A.; Amundsen, P.-A. Subarctic Freshwater Fish as Bio-indicators of Technogenic Pollution; Izdatelstvo Kolskogo Nauchnogo Tsentra RAN: Apatity, Russia, 1999; 142p. [Google Scholar]
- Lukin, A.A.; Kashulin, N.A. Status of Ichthyofauna of Water Bodies in the Border Zone of the USSR and Norway; Izdatelstvo Kolskogo Nauchnogo Tsentra RAN: Apatity, Russia, 2001; 50p. [Google Scholar]
- Zubova, E.M. Linear Growth of European Whitefish Coregonus lavaretus (L.) in Anthropogenically Modified Reservoirs of the European Subarctic (Case Study of the Murmansk Region) (Dissertation Abstract); Perm Regional Research University: Perm, Russia, 2015; 28p. [Google Scholar]
- Zubova, E.M.; Kashulin, N.A.; Terentyev, P.M. Biotic sympatry of whitefish Coregonus lavaretus (L.) in Kuetsjarvi lake (Pasvik river system, Murmansk region). Princ. Ecol. 2019, 2, 3–20. [Google Scholar]
- Kashulin, N.A.; Sandimirov, S.S.; Dauvalter, V.A.; Terentyev, P.M.; Denisov, D.B. Ekologicheskiy Katalog Ozer Murmanskoy Oblasti. Severo-zapadnaya Chast’ Murmanskoy Oblasti i Prigranichnoy Territorii Sopredel’nykh Stran, Part II; Izdatelstvo Kolskogo Nauchnogo Tsentra RAN: Apatity, Russia, 2009; 226p. [Google Scholar]
- Moiseenko, T.; Mjeled, M.; Branchurd, T.; Brettum, P.; Dauvalter, V.; Kagan, L.; Kashulin, N.; Kudriavtseva, L.; Lukin, A.; Sandimirov, S.; et al. Pasvik River Watercourse, Barents region: Pollution Impacts and Ecological Responses Investigation in 1993; NIVA-report OR-3118; Institute of North Industrial Ecology Problems: Apatity, Russia; Norwegian Institute for Water Research (Norway): Oslo, Norway, 1994; pp. 1–87. [Google Scholar]
- American Public Health Association. Standard Method for Examination for Water and Wastewater, 20th ed.; American Public Health Association: Washington, DC, USA, 1999; 2671p. [Google Scholar]
- Dauvalter, V.A. Geoecology of Lake Sediments; MGTU: Murmansk, Russia, 2012; 242p. [Google Scholar]
- Håkanson, L. An ecological risk index for aquatic pollution control—A sedimentological approach. Water Res. 1980, 14, 975–1001. [Google Scholar] [CrossRef]
- Norwegian Institute for Water Research. Intercomparison–2016. 1630: pH, Conductivity, Alkalinity, NO3-N, Cl, SO4, Ca, Mg, Na, K, TOC, Al, Fe, Mn, Cd, Pb, Cu, Ni and Zn; ICP Waters report 129/2016; Report No. 7081; Norwegian Institute for Water Research: Oslo, Norway, 2016; 72p. [Google Scholar]
- State standard 17.1.3.07–82. Protection of the Environment. Hydrosphere. The Rules of Water Quality Control of Water Bodies; Water Quality Control, State Standards: Moscow, Russia, 2010. [Google Scholar]
- Balushkina, E.V.; Vinberg, G.G. The Relationship between the Length and Body Weight of Planktonic Crustaceans. Experimental and Field Studies of the Biological Foundations of Lake Productivity; Nauka: Leningrad, Russia, 1979; 318p. [Google Scholar]
- Alekseev, V.R.; Tsalolikhin, S.Y. (Eds.) Key of Zooplankton and Zoobenthos of Fresh Waters of European Russia. V.1. Zooplankton; KMK: Moscow, Russia; St. Petersburg, Russia, 2016; 495p. [Google Scholar]
- Аbakumov, V.A. (Ed.) Guidance on Methods of Hydrobiological Analysis of Surface Water and Sediments; Gidrometeoizdat: St. Petersburg, Russia, 1992; 318p. [Google Scholar]
- Ruttner-Kolisko, A. Suggestion for biomass calculation of planktonic rotifers. Arch. Hydrobiol. Ergebn. Limnol. 1977, 8, 71–78. [Google Scholar]
- Denisov, D.B. Algae communities’ ecological peculiarities in polytypic subarctic water objects. Vestnik Kol’skogo Nauchnogo Centra RAN 2010, 1, 48–55. [Google Scholar]
- Denisov, D.B.; Kashulin, N.A. The phytoplankton communities under the Kola nuclear power Plant impact (the Imandra Lake). Trudy Kolskogo Nauchnogo Tsentra RAN Priknadnaya Ekologiya Severa. Appl. Ecol. North 2013, 16, 70–96. [Google Scholar]
- Guiry, M.D.; Guiry, G.M. AlgaeBase, World-wide Electronic Publication; National University of Ireland: Galway, Ireland, 2019; Available online: http://www.algaebase.org (accessed on 16 December 2019).
- Sladeček, V. General Biological Scheme of Water Quality. Sanitary and Technical Hydrobiology; Nauka: Moscow, Russia, 1967; pp. 26–31. [Google Scholar]
- Pantle, R.; Buck, H. Die biologische Uberwachung der Gewasser und die Darstellung der Ergebnisse. Gas-und Wasserbach 1955, 96, 17–21. [Google Scholar]
- Barinova, S.S.; Medvedeva, L.A.; Anisimova, O.V. Biodiversity of Environmental Indicator Algae; Pilies Studio: Tel Aviv, Israel, 2006; 498p. [Google Scholar]
- UNESCO. Determination of Photosynthetic Pigments in Sea Water, Rep. of SCOP-UNESCO Working Group 17; UNESCO: Paris, France, 1966; pp. 9–18. [Google Scholar]
- Kitaev, S.P. Ecological Basis of Bioproductivity of Lakes of Different Natural Zones; Nauka: Moscow, Russia, 1984; 204p. [Google Scholar]
- Sharov, A.N. The Phytoplankton Structure in the Reservoirs of the Far North under Conditions of Technogenic Pollution (Dissertation); Izdatelstvo Kolskogo Nauchnogo Tsentra RAN: Apatity, Russia, 2000; 168p. [Google Scholar]
- Sharov, A.N. Phytoplankton of the Kola Peninsula; Karel’skij nauchnyj centr RAN: Petrozavodsk, Russia, 2004; 113p. [Google Scholar]
- Alekseev, V.R.; Tsalolikhin, S.Y. (Eds.) Key of Zooplankton and Zoobenthos of Fresh Waters of European Russia. V.2. Zoobenthos; KMK: Moscow, Russia; St. Petersburg, Russia, 2016; 457p. [Google Scholar]
- Tsalolikhin, S.Y. (Ed) Key to Freshwater Invertebrates in Russia and Adjacent Territories. V.4. Diptera Insects; Nauka: St. Petersburg, Russia, 2000; 997p. [Google Scholar]
- Tsalolikhin, S.Y. (Ed) Key to Freshwater Invertebrates in Russia and Adjacent Territories. V.5. Insects; Nauka: St. Petersburg, Russia, 2001; 825p. [Google Scholar]
- Merritt, R.W.; Cummins, K.W. An Introduction the Aquatic Insects of North America, 2nd ed.; Kendall/Hunt Publishing company Dubuque: Dubuque, IA, USA, 1984; 722p. [Google Scholar]
- Sidorov, G.P.; Reshetnikov, Y.S. Salmon Fish of the European Northeast; KMK: Moscow, Russia; St. Petersburg, Russia, 2014; 345p. [Google Scholar]
- Reshetnikov, Y.S.; Bogdanov, V.D. Features of reproduction of whitefish. Ichthyology 2011, 51, 502–525. [Google Scholar]
- Mineeva, N.M. Photopigments in the Water of the Volga Reservoirs; Nauka: Moscow, Russia, 2004; 158p. [Google Scholar]
- Kashulin, N.A.; Dauvalter, V.A.; Denisov, D.B.; Valkova, S.A.; Vandysh, O.I.; Terentjev, P.M.; Kashulin, A.N. Selected aspects of the current state of freshwater resources in the Murmansk Region, Russia. J. Environ. Sci. Health Part A 2017, 52, 921–929. [Google Scholar] [CrossRef] [PubMed]
- Goncharova, T.O.; Ivanova, A.L.; Kaplin, V.T.; Borisova, A.S.; Norina, A.M.; Titova, E.N.; Sorokina, E.F. Characterization of pollution of some water bodies in the Murmansk region by the effluents of ore processing plants. Gydrohimicheskiye Mater. 1973, 2, 120–138. [Google Scholar]
- Dauvalter, V. Concentrations of heavy metals in superficial lake sediments of Pechenga district, Murmansk region, Russia. Vatten 1992, 48, 141–145. [Google Scholar]
- Dauvalter, V.; Rognerud, S. Heavy metals pollution in sediment of the Pasvik River drainage. Chemosphere 2001, 42, 9–18. [Google Scholar] [CrossRef]
- Lanteigne, S.; Schindler, M.; McDonald, A.M.; Skeries, K.; Abdu, Y.; Mantha, N.M.; Murayama, M.; Hawthorne, F.C.; Hochella, M.F., Jr. Mineralogy and Weathering of Smelter-Derived Spherical Particles in Soils: Implications for the Mobility of Ni and Cu in the Surficial Environment. Water Air Soil Pollut. 2012, 223, 3619–3641. [Google Scholar] [CrossRef]
- Slukovsky, Z.I.; Dauvalter, V.A. Morphology and composition of technogenic particles of bottom sediments of Lake Nyudyavr, Murmansk region. Notes RMO 2019, 3, 102–117. [Google Scholar]
- Yakovlev, V.A.; Nyost, T.; Langeland, A. The State of the Fauna of Aquatic Invertebrates in the Border Regions of the USSR and Norway; Izd. Akademii nauk SSSR: Apatity, Russia, 1991; 54p. [Google Scholar]
- Amundsen, P.A.; Staldvik, F.J.; Reshetnikov, Y.S.; Kashulin, N.A.; Lukin, A.A.; Bøhn, T.; Sandlund, O.T.; Popova, O.A. Invasion of vendace (Coregonus albula) in a subarctic watercourse. Biol. Conserv. 1999, 88, 405–413. [Google Scholar] [CrossRef]
- Moiseenko, T.I. Theoretical Basis for Norming of Anthropogenic Loads on Water Bodies of the Subarctic; Izdatelstvo Kolskogo Nauchnogo Tsentra RAN: Apatity, Russia, 1997; 261p. [Google Scholar]
- Reshetnokov, Y.S. Ecology and Systematics of Whitefishes; Nauka: Moscow, Russia, 1980; 301p. [Google Scholar]
- Harrod, C.; Mallela, L.; Kahilainen, K. Phenotype-environment correlations in a putative whitefish adaptive radiation. J. Anim. Ecol. 2010, 79, 1057–1068. [Google Scholar] [CrossRef]
- Kahilainen, K.; Østbye, K. Morphological differentiation and resource polymorphism in three sympatric whitefish Coregonus lavaretus (L.) forms in a subarctic lake. J. Fish Biol. 2006, 68, 63–79. [Google Scholar] [CrossRef]
- Kahilainen, K.; Malinen, T.; Tuamaala, A.; Lehtonen, H. Diel and seasonal habitat and food segregation of three sympatric Coregonus lavaretus forms in a subarctic lake. J. Fish Biol. 2004, 64, 418–434. [Google Scholar] [CrossRef]
- Kahilainen, K.; Malinen, T.; Tuomaala, A.; Alajaervi, E.; Tolonen, A.; Lehtonen, H. Empirical evaluation of phenotype–environment correlation and trait utility with allopatric and sympatric whitefish, Coregonus lavaretus (L.), populations in subarctic lakes. Biol. J. Linn. Soc. 2007, 92, 561–572. [Google Scholar] [CrossRef] [Green Version]
- Kahilainen, K.; Malinen, T.; Lentonen, H. Polar light regime and piscivory govern diel vertical migrations of planktivorous fish and zooplankton in a subarctic lake. Ecol. Freshw. Fish 2009, 18, 481–490. [Google Scholar] [CrossRef]
- Kahilainen, K.; Patterson, W.; Sonninen, E.; Harrod, C.; Kiljunen, M. Adaptive Radiation along a Thermal gradient: Preliminary results of habitat use and respiration rate divergence among whitefish morphs. PLoS ONE 2014, 11, e112085. [Google Scholar] [CrossRef] [Green Version]
- Præbel, K.; Knudsen, R.; Siwertsson, A.; Karhunen, M.; Kahilainen, K.; Ovaskainen, O.; Ǿstbye, K.; Peruzzi, S.; Fevolden, S.E.; Amundsen, P.A. Ecological speciation in postglacial European whitefish: Rapid adaptive radiations into the littoral, pelagic, and profundal lake habitats. Ecol. Evol. Res. 2013, 15, 4970–4986. [Google Scholar] [CrossRef] [Green Version]
- Siwertsson, A.; Knudsen, R.; Amundsen, P.A. Temporal stability in gill raker numbers of subarctic European whitefish populations. Adv. Limnol. 2008, 63, 229–240. [Google Scholar]
- Siwertsson, A.; Knudsen, R.; Kahilainen, K.; Præbel, K.; Primicerio, R.; Amundsen, P.A. Sympatric divercification as influenced by ecological opportunity and historical contingency in a young species lineage of whitefish. Evol. Ecol. Res. 2010, 12, 929–947. [Google Scholar]
- Østbye, K.; Naesje, T.F.; Bernatchez, L.; Himberg, M.; Hindar, K. Morphological divergence and origin of sympatric populations of European whitefish (Coregonus lavaretus (L.) in Lake Femud, Norway. J. Evol. Biol. 2005, 18, 683–702. [Google Scholar] [CrossRef]
- Mina, M.V. Fish Microevolution: Evolutionary Aspects of Phenetic Diversity; Nauka: Moscow, Russia, 1986; 193p. [Google Scholar]
- Dgebuadze, Y.Y. Ecological Patterns of Fish Growth Variability; Nauka: Moscow, Russia, 2001; 276p. [Google Scholar]
- Pauwels, M.; Frérot, H.; Souleman, D.; Vandenbulcke, F. Using biomarkers in an evolutionary context: Lessons from the analysis of biological responses of oligochaete annelids to metal exposure. Environ. Pollut. 2013, 179, 343–350. [Google Scholar] [CrossRef]
- Bickham, J.W. The four cornerstones of evolutionary toxicology. Ecotoxicology 2011, 20, 497–502. [Google Scholar] [CrossRef]
- Coutellec, M.A.; Barata, C. An introduction to evolutionary processes in ecotoxicology. Ecotoxicology 2011, 20, 493–496. [Google Scholar] [CrossRef] [PubMed]
- Shugart, L.R.; Theodorakis, C.W.; Bickham, J.W. Evolutionary toxicology. In Molecular Approaches in Natural Resource Conservation and Management; Cambridge University Press: New York, NY, USA, 2010; Volume 14, pp. 320–336. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Research Period | The Ratio of the Number of Fish Species in Catches, Specimens: | ||||||
|---|---|---|---|---|---|---|---|
| Trout | Whitefish | Vendace | Grayling | Pike | Perch | Burbot | |
| August 1990 | 6 | 150 | - | 1 | 51 | 45 | 1 |
| September 1991 | - | 371 | - | - | 28 | 40 | - |
| June–September 1992 | - | 225 | - | - | - | - | - |
| June–September 1998 | - | 373 | - | - | 2 | 5 | 7 |
| August 2004 | - | 468 | 37 | 1 | 12 | 54 | - |
| August 2005 | 1 | 23 | - | - | - | 17 | - |
| August 2007 | 2 | 177 | - | - | 1 | 57 | - |
| September 2009 | - | 113 | 1 | - | - | 1 | 6 |
| July–August 2012 | 5 | 363 | 23 | - | 12 | 33 | 7 |
| July–September 2013 | - | 90 | 5 | - | 7 | 13 | - |
| September 2015 | 2 | 201 | 34 | - | 2 | 45 | 2 |
| Layers of Sediments, cm | Ni | Cu | Zn | Co | Cd | Pb | As | Hg | Cd |
|---|---|---|---|---|---|---|---|---|---|
| 0–1 | 4032 | 1343 | 240 | 184.1 | 3.14 | 36.1 | 43.1 | 0.417 | |
| 22–23 | 32 | 40 | 80 | 15.9 | 0.10 | 6.6 | 2.62 | 0.049 | |
| Cf | 125.7 | 33.5 | 3.0 | 11.6 | 32.1 | 5.5 | 16.4 | 8.5 | 236.2 |
| Indicators | Research Periods | ||
|---|---|---|---|
| 1996–1998 | 2007–2011 | 2012–2015 | |
| Phytoplankton | |||
| Dominant taxa | Melosira varians Pandorina morum Asterionella formosa Diatoma tenuis Dinobryon sociale Eudorina sp. | Asterionella formosa Fragilaria tenera var. nanana Staurosira construens Diatoma tenuis Microcystis pulverea f. delicatissima Dinobryon bavaricum | Asterionella formosa Pseudosphaerocystis lacustris Mucidosphaerium pulchellum Fragilaria capucina Pseudanabaena sp. |
| B, g/m3 | 1.06 (1.06–2.05) | 1.95 (1.33–2.65) | 2.30 (1.23–10.68) |
| Chl «a», mg/m3 | 4.23 (2.71–5.95) | 4.51 (0.35–4.04) | 5.06 (0.46–28.34) |
| S | 1.92 (1.02–1.44) | 1.27 (1.17–1.52) | 1.42 (1.30–1.88) |
| K | III | II | II |
| T | β-mesotrophic | ||
| Zooplankton | |||
| Dominant taxa | Kellicottia longispina, Keratella cochlearis, Keratella quadrata, Notholca sp., Polyarthra sp., Bosmina obtusirostris, Daphnia cristata | Kellicottia longispina, Keratella cochlearis, Polyarthra sp., Bosmina obtusirostris | Keratella cochlearis, Notholca sp., Polyarthra sp. |
| N, thousand. Ind./m3 | 44.7–62.0 | 80.0–147.0 | 272.1–1254.3 |
| B, g/m3 | 0.2 (0.18–0.25) | 0.15 (0.02–0.68) | 2.3 (1.3–3.5) |
| H’(N) bit/ind. | 2.0–2.6 | 1.7–2.0 | 1.1–2.0 |
| B3/B2 | 0.1–0.5 | 0.01–0.5 | 0.01–0.9 |
| W = B/N, mg | 0.003–0.006 | 0.001–0.005 | 0.001–0.005 |
| T | α-oligotrophic | transitional from α- to β-oligotrophic | transitional from α- to β-mesotrophic |
| Indicators | Research Period | |
|---|---|---|
| 1990–1992 * | 2009–2013 | |
| Dominantspecies | Cricotopus ** Stictochironomus, Orthocladius, Tanytarsini, Pisidium sp. Limnephilidae, Polycentropodidae Chironomus sp., Sergentia sp., Tubifex tubifex | Cricotopus silvestris gr. Procladius choreus gr. Tubifex tubifex Sergentia coracina Chironomus sp. Prodiamesa olivacea |
| N, ind./m2 | - *** 2267 | 1680 506 |
| B, g/m2 | - 12.1 | 7.5 2.1 |
| Total number of taxa | 56 | 28 |
| T | eutrophic | oligotrophic |
| Fish Species | Research Period | ||
|---|---|---|---|
| 1990–1998 | 2004–2009 | 2013–2015 | |
| Trout | 0.5 | 0.3 | 0.8 |
| Whitefish | 85.7 | 80.5 | 77.4 |
| Vendace | 0.0 | 3.9 | 7.3 |
| Grayling | 0.1 | 0.1 | 0.0 |
| Pike | 6.2 | 1.3 | 2.5 |
| Perch | 6.9 | 13.3 | 10.8 |
| Burbot | 0.6 | 0.6 | 1.2 |
| Fish Species | Length as a Whole for the Sampling, mm | Weight as a Whole for the Sampling, g | Length of Sexually Mature Fish, mm | Weight of Sexually Mature Fish, g |
|---|---|---|---|---|
| 1990–1998 | ||||
| Sparsely rakered whitefish | 195 ± 3.7 88–465 (318) | 117 ± 9.8 4–1300 (318) | 206 ± 7.7 90–465(42) | 175 ± 22.2 4–1300 (42) |
| Densely rakered whitefish | 139 ± 3.3 85–300 (211) | 34 ± 3.4 4–278 (211) | 146 ± 5.2 93–280 (36) | 46 ± 5.9 5–261 (36) |
| Vendace | - | - | - | - |
| Pike | 384 ± 7.1 255–540 (81) | 487 ± 26.9 110–1205 (81) | 427 ± 15.6 310–540 (18) | 620 ± 63.1 207–1205 (18) |
| Perch | - | 98 ± 9.2 6–385 (90) | - | - |
| 2004–2008 | ||||
| Sparsely rakered whitefish | 160 ± 1.8 83–265 (421) | 49 ± 1.8 6–254 (421) | 161 ± 2.1 87–260 (228) | 48 ± 2.2 6–210 (228) |
| Densely rakered whitefish | 134 ± 2.4 75–293 (359) | 33 ± 2.1 2–340 (359) | 133 ± 3.1 87–293 (218) | 35 ± 2.9 4–340 (218) |
| Vendace | 110 ± 1.6 100–130 (38) | 11 ± 0.5 6–19 (38) | 109 ± 1.7 100–130 (34) | 11 ± 0.5 6–19 (34) |
| Pike | 326 ± 18.3 255–499 (13) | 373 ± 71.5 164–1079 (13) | 499 (1) | 890 (1) |
| Perch | 186 ± 4.2 110–295 (129) | 122 ± 10.7 17–604 (129) | 252 ± 5.2 128–295 (33) | 296 ± 17.9 28–604 (33) |
| 2012–2015 | ||||
| Sparsely rakered whitefish | 185 ± 3.6 77–407 (233) | 94 ± 8.2 4–981 (233) | 188 ± 2.62 116–407 (89) | 95 ± 13.33 14–843 (89) |
| Densely rakered whitefish | 160 ± 2.58 75–312 (416) | 58 ± 3.0 4–365 (416) | 160 ± 5.0 80–312 (144) | 64 ± 6.53 4–365 (144) |
| Vendace | 104 ± 1.1 87–141 (62) | 9 ± 0.4 5–24 (62) | 104 ± 1.0 96–123 (37) | 9 ± 0.5 6–19 (37) |
| Pike | 408 ± 15.7 260–547 (21) | 637 ± 78.5 150–1385 (21) | 449 ± 24.3 400–516 (4) | 788 ± 158.5 471–1220 (4) |
| Perch | 155 ± 4.9 45–290 (91) | 77 ± 10.7 1–792 (91) | 169 ± 7.8 119–290 (28) | 87 ± 17.2 23–440 (28) |
| Fish Species, Study Period | Age | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 1+ | 2+ | 3+ | 4+ | 5+ | 6+ | 7+ | 8+ | 9+ | 10+ | |
| FL | ||||||||||
| Sparsely rakered whitefish, 1990–1998 | 136 ± 18.9 103–220 (6) | 171 ± 7.0 97–235 (23) | 195 ± 5.5 124–250 (35) | 231 ± 6.4 173–325 (30) | 259 ± 9.6 206–330 (16) | 321 ± 7.3 289–343 (7) | 388 ± 9.5 360–400 (4) | 400 (1) | 285 (1) | 465 (1) |
| Sparsely rakered whitefish, 2012–2015 | 103 ± 7.0 77–137 (7) | 148 ± 4.8 84–215 (43) | 156 ± 3.8 112–237 (52) | 188 ± 4.8 124–270 (53) | 231 ± 9.2 160–334 (35) | 220 ± 12.1 162–407 (22) | 221 ± 14.8 190–292 (6) | 421 (1) | 214 ± 11.5 202–225 (2) | 362 (1) |
| T | 1.77 | 2.73 | 5.93 | 5.37 | 1.81 | 4.59 | 8.33 | - | - | - |
| Densely rakered whitefish, 1990–1998 | 117 ± 3.9 102–158 (16) | 159 ± 7.3 110–220 (19) | 207 ± 8.0 165–270 (13) | 266 ± 5.2 255–275 (4) | 202 ± 8.0 194–210 (2) | - | 300 (1) | - | - | - |
| Densely rakered whitefish, 2012–2015 | 97 ± 5.5 91–102 (2) | 126 ± 2.6 80–253 (139) | 156 ± 3.4 97–247 (88) | 184 ± 4.9 107–247 (72) | 225 ± 3.9 131–310 (58) | 261 ± 9.2 214–312 (12) | 259 ± 16.0 243–275 (2) | - | 295 (1) | - |
| T | 1.75 | 4.38 | 5.45 | 3.95 | 1.09 | - | - | - | - | - |
| W | ||||||||||
| Sparsely rakered whitefish, 1990–1998 | 33 ± 16.5 8–113 (6) | 58 ± 7.4 7–142 (23) | 89 ± 8.3 17–189 (35) | 153 ± 18.5 24–465 (30) | 233 ± 31.5 106–475 (16) | 468 ± 33.6 284–536 (7) | 853 ± 71.8 658–1000 (4) | 1037 (1) | 277 (1) | 1300 (1) |
| Sparsely rakered whitefish, 2012–2015 | 10 ± 2.4 4–24 (7) | 36 ± 4.0 5–115 (43) | 41 ± 3.7 13–135 (52) | 81 ± 8.1 16–300 (53) | 173 ± 23.9 40–488 (35) | 274 ± 52.3 86–1295 (22) | 133± 34.6 80–302 (6) | 981 (1) | 118 ± 19.5 98–137 (2) | 440 (1) |
| T | 1.47 | 2.92 | 5.84 | 4.12 | 1.47 | 2.03 | 10.09 | - | - | - |
| Densely rakered whitefish, 1990–1998 | 14 ± 2.1 6–36 (16) | 43 ± 6.7 8–118 (16) | 99 ± 13.9 38–226 (13) | 224 ± 20.1 182–261 (4) | 69 ± 31.1 47–91 (2) | - | 278 (1) | - | - | - |
| Densely rakered whitefish, 2012–2015 | 8 ± 0.5 7–8 (2) | 22.3 ± 1.8 4–176 (139) | 43 ± 3.0 7–189 (88) | 74 ± 5.2 11–187 (72) | 134 ± 7.5 24–365 (58) | 213 ± 23.9 117–349 (12) | 212 ± 52.0 160–264 (2) | - | 290 (1) | - |
| T | 1.11 | 3.71 | 5.91 | 6.67 | 1.60 | - | ||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zubova, E.M.; Kashulin, N.A.; Dauvalter, V.A.; Denisov, D.B.; Valkova, S.A.; Vandysh, O.I.; Slukovskii, Z.I.; Terentyev, P.M.; Cherepanov, A.A. Long-Term Environmental Monitoring in an Arctic Lake Polluted by Metals under Climate Change. Environments 2020, 7, 34. https://doi.org/10.3390/environments7050034
Zubova EM, Kashulin NA, Dauvalter VA, Denisov DB, Valkova SA, Vandysh OI, Slukovskii ZI, Terentyev PM, Cherepanov AA. Long-Term Environmental Monitoring in an Arctic Lake Polluted by Metals under Climate Change. Environments. 2020; 7(5):34. https://doi.org/10.3390/environments7050034
Chicago/Turabian StyleZubova, Elena M., Nikolay A. Kashulin, Vladimir A. Dauvalter, Dmitry B. Denisov, Svetlana A. Valkova, Oksana I. Vandysh, Zakhar I. Slukovskii, Peter M. Terentyev, and Alexander A. Cherepanov. 2020. "Long-Term Environmental Monitoring in an Arctic Lake Polluted by Metals under Climate Change" Environments 7, no. 5: 34. https://doi.org/10.3390/environments7050034