Phylogenetic Identification, Diversity, and Richness of Aspergillus from Homes in Havana, Cuba


 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Isolates
2.2. Sampling Procedure
2.3. Monosporic Culture Procedure
2.4. Macro- and Micromorphology
2.5. Obtaining DNA
2.6. Phylogenetic Analysis
2.7. Richness, Abundance, and Diversity of Species
3. Results
3.1. Phenotypic Identification
3.2. Phylogenetic Analysis
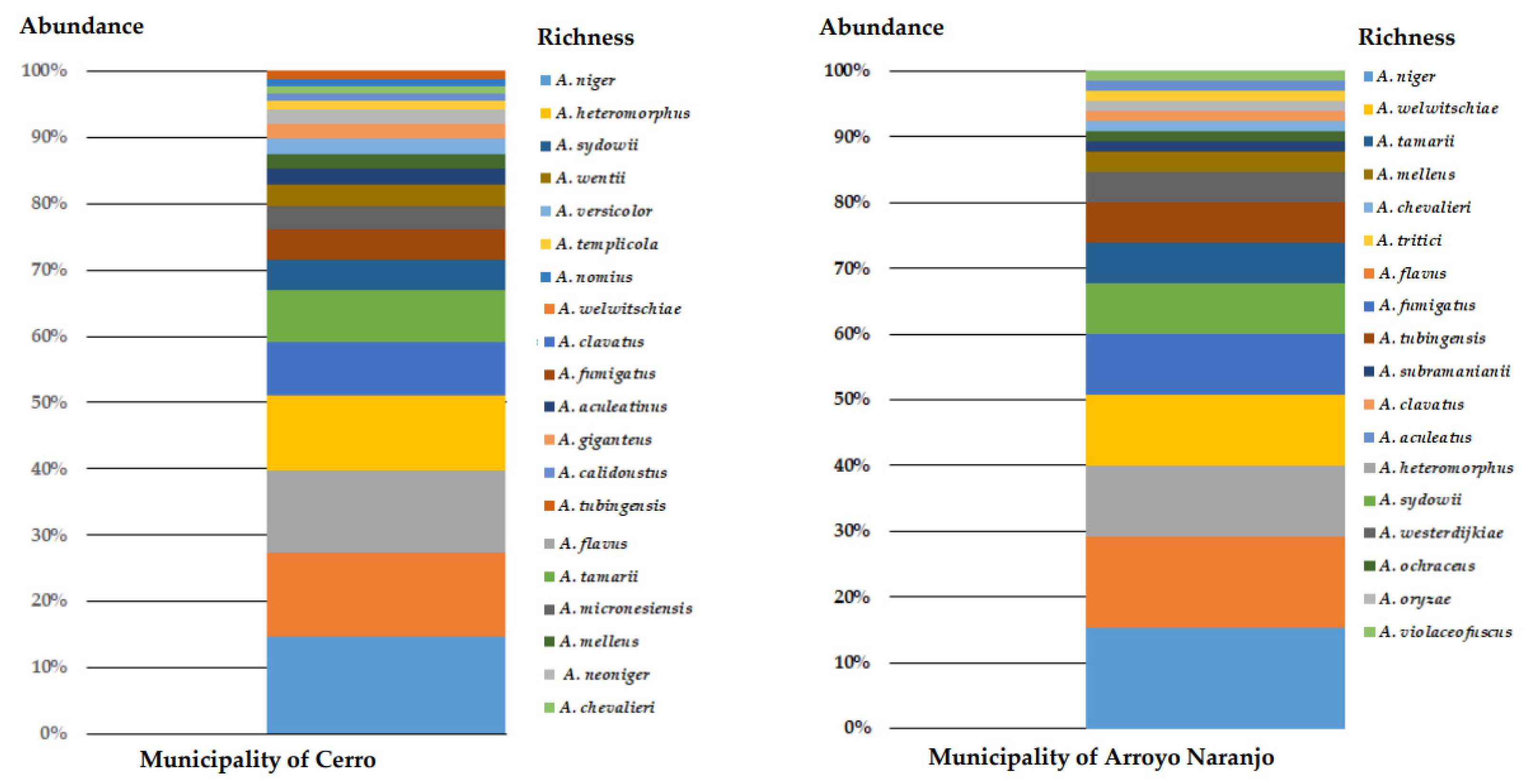
3.3. Richness, Abundance, and Diversity of Species
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Visagie, C.M.; Hirooka, Y.; Tanney, J.B.; Whitfield, E.; Mwange, K.; Meijer, M.; Amend, A.S.; Seifert, K.A.; Samson, R.A. Aspergillus, Penicillium and Talaromyces isolated from house dust samples collected around the world. Stud. Mycol. 2014, 78, 63–139. [Google Scholar] [CrossRef] [Green Version]
- Rahmawati, R.; Sembiring, L.; Zakaria, L.; Rahayu, E.S. The diversity of indoor airborne molds growing in the university libraries in Indonesia. Biodiversitas 2018, 19, 194–220. [Google Scholar] [CrossRef]
- Muhsin, T.M.; Adlan, M.M. Seasonal distribution pattern of outdoor airborne fungi in Basrah city, southern Iraq. J. Basrah Res. Sci. 2012, 38, 90–98. [Google Scholar]
- Lang-Yona, N.; Dannemiller, K.; Yamamoto, N.; Burshtein, N.; Peccia, J.; Yarden, O.; Rudich, Y. Annual distribution of allergenic fungal spores in atmospheric particulate matter in the Eastern Mediterranean; a comparative study between ergosterol and quantitative PCR analysis. Atmos. Chem. Phys. 2012, 12, 2681–2690. [Google Scholar] [CrossRef] [Green Version]
- Ahluwalia, S.K.; Matsui, E.C. Indoor environmental interventions for furry pet allergens, pest allergens, and mold: Looking to the future. J. Allergy Clin. Immunol. Pract. 2018, 6, 9–19. [Google Scholar] [CrossRef]
- Hohl, T.M.; Feldmesser, M. Aspergillus fumigatus: Principles of pathogenesis and host defense. Eukaryot. Cell 2007, 6, 1953–1963. [Google Scholar] [CrossRef] [Green Version]
- van de Veerdonk, F.L.; Gresnigt, M.S.; Romani, L.; Netea, M.G.; Latgé, J.P. Aspergillus fumigatus morphology and dynamic host interactions. Nat. Rev. Microbiol. 2017, 15, 661–674. [Google Scholar] [CrossRef]
- Gautier, M.; Normand, A.C.; Ranque, S. Previously unknown species of Aspergillus. Clin. Microbiol. Infect. 2016, 22, 662–669. [Google Scholar] [CrossRef] [Green Version]
- Vidal-Acuña, M.R.; Ruiz-Pérez de Pipaón, M.; Torres-Sánchez, M.J.; Aznar, J. Identification of clinical isolates of Aspergillus, including cryptic species, by matrix assisted laser desorption ionization time-of-flight mass spectrometry (MALDI-TOF MS). Med. Mycol. 2017, 56, 838–846. [Google Scholar] [CrossRef]
- Caillaud, D.; Leynaert, B.; Keirsbulck, M.; Nadif, R. Indoor mould exposure, asthma and rhinitis: Findings from systematic reviews and recent longitudinal studies. Eur. Respir. Rev. 2018, 27, 170137. [Google Scholar] [CrossRef] [Green Version]
- Awad, A.H.A.; Saeed, Y.; Shakour, A.A.; Abdellatif, N.M.; Ibrahim, Y.H.; Elghanam, M.; Elwakeel, F. Indoor air fungal pollution of a historical museum, Egypt: A case study. Aerobiologia 2020, 36, 197–209. [Google Scholar] [CrossRef]
- Grinn-Gofron, A. Airborne Aspergillus and Penicillium in the atmosphere of Szczecin, (Poland) (2004–2009). Aerobiologia 2011, 27, 67–76. [Google Scholar] [CrossRef] [Green Version]
- Almaguer, M.; Rojas-Flores, T.I. Airborne culturable fungi of the atmosphere of Havana, Cuba. NACC (Bioloxia) 2013, 20, 35–45. [Google Scholar]
- Diba, K.; Jangi, F.; Makhdoomi, K.; Moshiri, N.; Mansouri, F. Aspergillus diversity in the environments of nosocomial infection cases at a university hospital. J. Med. Life 2019, 12, 128–132. [Google Scholar] [CrossRef]
- Rudramurthy, S.M.; Paul, R.A.; Chakrabarti, A.; Mouton, J.W.; Meis, J.F. Invasive aspergillosis by Aspergillus flavus: Epidemiology, diagnosis, antifungal resistance, and management. J. Fungi (Basel) 2019, 5, 55. [Google Scholar] [CrossRef] [Green Version]
- Samson, R.A.; Varga, J. Molecular Systematics and its Teleomorphs. Aspergillus: Molecular biology and genomics. In Aspergillus: Molecular Biology and Genomics; Machida, M., Gomi, K., Eds.; Caister Academic Press: Poole, UK, 2010; p. 19. [Google Scholar]
- Campana, R.; Sabatini, L.; Frangipani, E. Moulds on cementitious building materials-problems, prevention and future perspectives. Appl. Microbiol. Biotechnol. 2020, 104, 509–514. [Google Scholar] [CrossRef]
- Houbraken, J.; de Vries, R.P.; Samson, R.A. Modern taxonomy of biotechnologically important Aspergillus and Penicillium species. Adv. Appl. Microbiol. 2014, 86, 199–249. [Google Scholar] [CrossRef]
- Samson, R.A.; Visagie, C.M.; Houbraken, J.; Hong, S.B.; Hubka, V.; Klaassen, C.H.W.; Perrone, G.; Seifert, K.A.; Susca, A.; Tanney, J.B.; et al. Phylogeny, identification and nomenclature of the genus Aspergillus. Stud. Mycol. 2014, 78, 141–173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wijayawardene, N.N.; Hyde, K.D.; Al-Ani, L.K.T.; Tedersoo, L.; Haelewaters, D.; Rajeshkumar, K.C.; Zhao, R.L.; Aptroot, A.; Leontyev, D.V.; Saxena, R.K.; et al. Outline of Fungi and fungus-like taxa. Mycosphere 2020, 11, 1060–1456. [Google Scholar] [CrossRef]
- Rojas, T.I.; Martinez, E.; Gomez, Y.; Alvarado, Y. Airborne spores of Aspergillus species in cultural institutions at Havana University. Grana 2002, 41, 190–193. [Google Scholar] [CrossRef]
- Júnior, D.P.L.; Pereira, R.S.; de Almeida, W.S.; Simões, S.D.A.A.; Yamamoto, A.C.A.; de Souza, J.V.R.; Martins, E.R.; dos Santos, F.A.L.; Hahn, R.C. Indoor Air Mycological Survey and Occupational Exposure in Libraries in Mato Grosso-Central Region-Brazil. Adv. Microbiol. 2018, 8, 324–353. [Google Scholar] [CrossRef] [Green Version]
- Stanley, H.O.; Onwuna, B.; Ugboma, C.J. Microbial Assessment of Indoor Air Quality of Ventilation Systems. Asian J. Adv. Res. Rep. 2019, 3, 1–7. [Google Scholar] [CrossRef]
- Beltrán Rodríguez, N.; Juan-Galán, J.L.; Fernández Andreu, C.M.; Yera, D.M.; Barrios Pita, M.; Perurena Lancha, M.R.; Velar Martínez, R.E.; Illnait Zaragozí, M.T.; Martínez Machín, G.F. Chronic pulmonary aspergillosis in patients with underlying respiratory disorders in Cuba-a pilot study. J. Fungi (Bassel) 2019, 5, 18. [Google Scholar]
- Herrera Barrios, O.; Peneque Rodríuez, I.; Rodríguez Canosa, J.; Jiménez Martínez, M.; Otero Heredia, L.; Ruiz Torres, N.; Guedes Vidal, L.; Martínez Ayala, Y.; Suárez Navaroo, N.; Linares Luna, L. Sensibilidad cutánea a hongos ambientales y estudio de la micobiota nasal de pacientes con alergias respiratoria. Investig. Medicoquir. 2019, 11, 1. [Google Scholar]
- Álvarez, M.; Castro, R.L.; Leyva, Y.; López, B.; Rodríguez, J.; Rojas, T.I.; Piera, O.M.; Labrada, A. Sensibilización a hongos anemófilos en trabajadores(as) del Archivo y Biblioteca de la Universidad de La Habana. Arch. Hosp. Univ. General Calixto García 2020, 8, 159–172. [Google Scholar]
- Irinyi, L.; Serena, C.; Garcia-Hermoso, D.; Arabatzis, M.; Desnos-Ollivier, M.; Vu, D.; Cardinali, G.; Arthur, I.; Normand, A.C.; Giraldo, A.; et al. International Society of Human and Animal Mycology (ISHAM)-ITS reference DNA barcoding database—The quality controlled standard tool for routine identification of human and animal pathogenic fungi. Med. Mycol. 2015, 53, 313–337. [Google Scholar] [CrossRef]
- Klich, M.; Pitt, J.I. A Laboratory Guide to the Common Aspergillus Species and Their Teleomorphs; Commonwealth Scientific and Industrial Research Organization, Division of Food Processing: North Ryde, NSW, Australia, 1988; p. 116.
- Glass, N.L.; Donaldson, G.C. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes. Appl. Environ. Microbiol. 1995, 61, 1323–1330. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.J.; Whelen, S.; Hall, B.D. Phylogenetic relationships among ascomycetes: Evidence from an RNA polymerase II subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Adams, R.I.; Miletto, M.; Taylor, J.W.; Bruns, T.D. The diversity and distribution of fungi on residential surfaces. PLoS ONE 2013, 8, e78866. [Google Scholar] [CrossRef] [PubMed]
- Kumari, P.; Woo, C.; Yamamoto, N.; Choi, H.L. Variations in abundance, diversity and community composition of airborne fungi in swine houses across seasons. Sci. Rep. 2016, 6, 37929. [Google Scholar] [CrossRef] [PubMed]
- De la Vega Pazitková, T.; Pérez Martínez, V.; Castillo Iriarte, L. Comportamiento clínico epidemiológico del asma bronquial en menores de 15 años. Bronchial Asthma Clinical and epidemiological pattern in children younger 15 years. Rev. Cubana Med. Gen. Integr. 2013, 29, 253–266. [Google Scholar]
- Venero Fernández, S.J.; Varona Pérez, P.; Fabret Ortiz, D.; Suárez Medina, R.; Bonet Gorbea, M.; Molina Esquivel, E.; Romero Placeres, M. Asma bronquial y rinitis en escolares de Ciudad de la Habana (2001 a 2002). Rev. Cubana Hig. Epidemiol. 2009, 47. Available online: https://www.redalyc.org/articulo.oa?id=223217513005 (accessed on 4 January 2021).
- Aira, M.J.; Rojas, T.I.; Jato, V. Fungi associated with three houses in Havana (Cuba). Grana 2002, 41, 114–118. [Google Scholar] [CrossRef]
- Rojas, T.I.; Martínez, E.; Aira, M.J.; Almaguer, M. Aeromicota de ambientes internos: Comparación de métodos de muestreo. Bol. Micol. 2008, 23, 67–73. [Google Scholar] [CrossRef] [Green Version]
- Rojas, T.I.; Aira, M.J. Fungal biodiversity in indoor environments in Havana, Cuba. Aerobiologia 2012, 28, 67–374. [Google Scholar] [CrossRef]
- Brito Devoto, T.B.; Alava, K.S.H.; Pola, S.J.; Pereda, R.; Rubeglio, E.; Finquelievich, J.L.; Cuestas, M.L. Molecular epidemiology of Aspergillus species and other moulds in respiratory samples from Argentinean patients with cystic fibrosis. Med. Mycol. 2020, 58, 867–873. [Google Scholar] [CrossRef]
- Richardson, M.; Bowyer, P.; Sabino, R. The human lung and Aspergillus: You are what you breathe in? Med. Mycol. 2019, 57, S145–S154. [Google Scholar] [CrossRef] [Green Version]
- Hashimoto, A.; Hagiwara, D.; Watanabe, A.; Yahiro, M.; Yikelamu, A.; Yaguchi, T.; Kamei, K. Drug sensitivity and resistance mechanism in Aspergillus section Nigri strains from Japan. Antimicrob. Agents Chemother. 2017, 61, e02583-16. [Google Scholar] [CrossRef] [Green Version]
- Krijgsheld, P.; Altelaar, A.M.; Post, H.; Ringrose, J.H.; Müller, W.H.; Heck, A.J.; Wösten, H.A. Spatially resolving the secretome within the mycelium of the cell factory Aspergillus niger. J. Proteome Res. 2012, 11, 2807–2818. [Google Scholar] [CrossRef] [PubMed]
- Cárdenas, M.X.; Cortes, J.A.; Parra, C.M. Presencia de Aspergillus spp. en áreas de riesgo en pacientes trasplantados en un hospital universitario. Rev. Iberoam. Micol. 2008, 25, 232–236. [Google Scholar] [CrossRef]
- Vermeulen, E.; Maertens, J.; Meersseman, P.; Saegeman, V.; Dupont, L.; Lagrou, K. Invasive Aspergillus niger complex infections in a Belgian tertiary care hospital. Clin. Microbiol. Infect. 2014, 20, O333–O335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atchade, E.; Jean-Baptiste, S.; Houzé, S.; Chabut, C.; Massias, L.; Castier, Y.; Brugière, O.; Mal, H.; Montravers, P. Fatal invasive aspergillosis caused by Aspergillus niger after bilateral lung transplantation. Med. Mycol. Case Rep. 2017, 17, 4–7. [Google Scholar] [CrossRef] [PubMed]
- Pantelides, I.S.; Aristeidou, E.; Lazari, M.; Tsolakidou, M.D.; Tsaltas, D.; Christofidou, M.; Kafouris, D.; Christou, E.; Ioannou, N. Biodiversity and ochratoxin A profile of Aspergillus section Nigri populations isolated from wine grapes in Cyprus vineyards. Food Microbiol. 2017, 67, 106–115. [Google Scholar] [CrossRef]
- Frías-De-León, M.G.; Rosas-de-Paz, E.; Arenas, R.; Atoche, C.; Duarte-Escalante, E.; Molina-de-Soschin, D.; Acosta-Altamirano, G.; Reyes-Montes, M.R. Identification of Aspergillus tubingensis in a primary skin infection. J. Mycol. Med. 2018, 28, 274–278. [Google Scholar] [CrossRef]
- Rodrigues, P.; Venâncio, A.; Kozakiewicz, Z.; Lima, N. A polyphasic approach to the identification of aflatoxigenic and non-aflatoxigenic strains of Aspergillus section Flavi isolated from Portuguese almonds. Int. J. Food Microbiol. 2009, 129, 187–193. [Google Scholar] [CrossRef] [Green Version]
- Frisvad, J.C.; Hubka, V.; Ezekiel, C.N.; Hong, S.B.; Nováková, A.; Chen, A.J.; Arzanlou, M.; Larsen, T.O.; Sklenář, F.; Mahakarnchanakul, W.; et al. Taxonomy of Aspergillus section Flavi and their production of aflatoxins, ochratoxins and other mycotoxins. Stud. Mycol. 2019, 93, 1–63. [Google Scholar] [CrossRef]
- Sabino, R.; Burco, J.; Valente, J.; Veríssimo, C.; Clemons, K.V.; Stevens, D.A.; Tell, L.A. Molecular identification of clinical and environmental avian Aspergillus isolates. Arch. Microbiol. 2019, 201, 253–257. [Google Scholar] [CrossRef]
- Visagie, C.M.; Yilmaz, N.; Renaud, J.B.; Sumarah, M.W.; Hubka, V.; Frisvad, J.C.; Chen, A.J.; Meijer, M.; Seifert, K.A. A survey of xerophilic Aspergillus from indoor environment, including descriptions of two new section Aspergillus species producing eurotium-like sexual states. MycoKeys 2017, 19, 1–30. [Google Scholar] [CrossRef] [Green Version]
- Seyedmousavi, S.; Lionakis, M.S.; Parta, M.; Peterson, S.W.; Kwon-Chung, K.J. Emerging Aspergillus species almost exclusively associated with primary immunodeficiencies. Open Forum Infect. Dis. 2018, 5, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahata, Y.; Hiruma, M.; Sugita, T.; Muto, M. A case of onychomycosis due to Aspergillus sydowii diagnosed using DNA sequence analysis. Mycoses 2008, 51, 170–173. [Google Scholar] [CrossRef] [PubMed]
- Charles, M.V.P.; Noyal, M.J.; Easow, J.M. Invasive pulmonary aspergillosis caused by Aspergillus versicolor in a patient on mechanical ventilation. Australas. Med. J. 2011, 4, 632–634. [Google Scholar] [CrossRef] [PubMed]
- Varga, J.; Due, M.; Frisvad, J.C.; Samson, R.A. Taxonomic revision of Aspergillus section Clavati based on molecular, morphological and physiological data. Stud. Mycol. 2007, 59, 89–106. [Google Scholar] [CrossRef] [PubMed]
- Houbraken, J.; Due, M.; Varga, J.; Meijer, M.; Frisvad, J.C.; Samson, R.A. Polyphasic taxonomy of Aspergillus section Usti. Stud. Mycol. 2007, 59, 107–128. [Google Scholar] [CrossRef]
- Pelaéz, T.; Álvarez-Pérez, S.; Mellado, E.; Serrano, D.; Valerio, M.; Blanco, J.L.; Garcia, M.E.; Muñoz, P.; Cuenca-Estrella, M.; Bouza, E. Invasive aspergillosis caused by cryptic Aspergillus species: A report of two consecutive episodes in a patient with leukaemia. J. Med. Microbiol. 2013, 62, 474–478. [Google Scholar] [CrossRef] [Green Version]
- Siqueira, J.P.Z.; Sutton, D.; Gené, J.; García, D.; Wiederhold, N.; Guarro, J. Species of Aspergillus section Aspergillus from clinical samples in the United States. Med. Mycol. 2017, 56, 541–550. [Google Scholar] [CrossRef]
- Oliveira, M.; Caramalho, R. Aspergillus fumigatus: A mere bioaerosol or a powerful biohazard? NACC (Bioloxia) 2014, 21, 57–64. [Google Scholar]
- Ghazaei, C. Molecular Insights into Pathogenesis and Infection with Aspergillus fumigatus. Malays J. Med. Sci. 2017, 24, 10–20. [Google Scholar] [CrossRef]
- Hubka, V.; Kubatova, A.; Mallatova, N.; Sedlacek, P.; Melichar, J.; Skorepova, M.; Mencl, K.; Lyskova, P.; Sramkova, B.; Chudickova, M.; et al. Rare and New Etiological Agents Revealed Among 178 Clinical Aspergillus strains obtained from Czech patients and characterized by molecular sequencing. Med. Mycol. 2012, 50, 601–610. [Google Scholar] [CrossRef] [Green Version]
- Duarte-Escalante, E.; Frías-De-León, M.G.; Martínez-Herrera, E.; Acosta-Altamirano, G.; Rosas-de-Paz, E.; Reséndiz-Sánchez, J.; Refojo, N.; Reyes-Montes, M.D.R. Identification of CSP types and genotypic variability of clinical and environmental isolates of Aspergillus fumigatus from different geographic origins. Microorganisms 2020, 8, 688. [Google Scholar] [CrossRef] [PubMed]
- Montenegro, G.; Puch, S.S.; Jewtuchowicz, V.M.; Pinoni, M.V.; Relloso, S.; Temporitti, E.; Iovannitti, C.A.; Mujica, M.T. Phenotypic and genotypic characterization of Aspergillus lentulus and Aspergillus fumigatus isolates in a patient with probable invasive aspergillosis. J. Med. Microbiol. 2009, 58, 391–395. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Municipality | Origin | Isolate |
|---|---|---|
| Cerro | Outdoor air | CCMFBH-835, CCMFBH-836, CCMFBH-837, CCMFBH-838, CCMFBH-841, CCMFBH-842, CCMFBH-843, CCMFBH-844, CCMFBH-845, CCMFBH-846, CCMFBH-847, CCMFBH-852, CCMFBH-853, CCMFBH-854, CCMFBH-855, CCMFBH-856, CCMFBH-857, CCMFBH-858, CCMFBH-860, CCMFBH-863, CCMFBH-864, CCMFBH-868, CCMFBH-869 |
| Arroyo Naranjo | Outdoor air | CCMFBH-878, CCMFBH-879, CCMFBH-886, CCMFBH-887, CCMFBH-891, CCMFBH-892, CCMFBH-898, CCMFBH-899 |
| Cerro | Indoor air | CCMFBH-839, CCMFBH-848, CCMFBH-849, CCMFBH-850, CCMFBH-859, CCMFBH-861, CCMFBH-862, CCMFBH-866, CCMFBH-867, CCMFBH-870, CCMFBH-871, CCMFBH-872, CCMFBH-873, CCMFBH-874, CCMFBH-899, CCMFBH-900, CCMFBH-901, CCMFBH-902, CCMFBH-903, CCMFBH-904, CCMFBH-905, CCMFBH-906, CCMFBH-907, CCMFBH-908, CCMFBH-909, CCMFBH-911, CCMFBH-912, CCMFBH-913, CCMFBH-914, CCMFBH-919, CCMFBH-920, CCMFBH-921, CCMFBH-922, CCMFBH-923, CCMFBH-924 CCMFBH-925, CCMFBH-926, CCMFBH-940, CCMFBH-941, CCMFBH-942, CCMFBH-944, CCMFBH-945, CCMFBH-946, CCMFBH-947, CCMFBH-948, CCMFBH-949, CCMFBH-950, CCMFBH-951, CCMFBH-952 |
| Arroyo Naranjo | Indoor air | CCMFBH-875, CCMFBH-876, CCMFBH-880, CCMFBH-881, CCMFBH-883, CCMFBH-884, CCMFBH-885, CCMFBH-888, CCMFBH-889, CCMFBH-890, CCMFBH-893, CCMFBH-895, CCMFBH-896, CCMFBH-916, CCMFBH-917, CCMFBH-918, CCMFBH-927, CCMFBH-928, CCMFBH-929, CCMFBH-930, CCMFBH-932, CCMFBH-933, CCMFBH-934, CCMFBH-935, CCMFBH-936, CCMFBH-937, CCMFBH-938, CCMFBH-939, CCMFBH-953, CCMFBH-954, CCMFBH-955, CCMFBH-996, CCMFBH-997, CCMFBH-1000, CCMFBH-1001 |
| Cerro | Dust | CCMFBH-965, CCMFBH-966, CCMFBH-967, CCMFBH-968, CCMFBH-969, CCMFBH-970, CCMFBH-971, CCMFBH-972, CCMFBH-973, CCMFBH-974, CCMFBH-975, CCMFBH-976, CCMFBH-977, CCMFBH-978 |
| Arroyo Naranjo | Dust | CCMFBH-979, CCMFBH-980, CCMFBH-981, CCMFBH-982, CCMFBH-983, CCMFBH-984, CCMFBH-985, CCMFBH-986, CCMFBH-987, CCMFBH-988, CCMFBH-989, CCMFBH-990, CCMFBH-992, CCMFBH-993, CCMFBH-994, CCMFBH-995 |
| Cerro | Walls | CCMFBH-956, CCMFBH-958 |
| Arroyo Naranjo | Walls | CCMFBH-959, CCMFBH-960, CCMFBH-961, CCMFBH-962, CCMFBH-963, CCMFBH-964 |
| Section | Macro- and Micromorphological Characteristics |
|---|---|
| Versicolores | Colony on CYA, 25 °C, 7 days: Variable coloration, greenish, turquoise, yellowish, and radially sulcate; velutinous texture; tawny olive to orange cinnamon reverse, vinaceous or brown or scarlet; moderate to abundant exudate, clear to yellowish to reddish-brown, reddish-brown soluble pigment. Biseriate conidial heads, radiate, globose to subglobose conidia, rough to prickly. |
| Usti | Colony on CYA, 25 °C, 7 days: Blond/greyish-yellow; floccose texture; yellow reverse with a beige center. Biseriate, predominantly pyriform vesicles, globose conidia, very rough ornamentation, with an inner and outer visible wall. |
| Flavi | Colony on CYA, 25 °C, 7 days: Green olive, brown olive, greenish-brown, brownish yellow; woolly or floccose texture; colorless, light brown or orange reverse. Presence of brown sclerotia in some isolates. Biseriate and uniseriate conidial heads, radiated to columnar; rough, colorless stipe, globose to ellipsoidal conidia, rough in some isolates. |
| Aspergillus | Colony on CYA, 25 °C, 7 days: Sulfur yellow mycelium; floccose texture; luteous to ochreous reverse. Uniseriate conidial heads, Ascomata Eurotium-like, cleistothecial, superficial, yellow, globose to subglobose. Ascospores hyaline, globose to subglobose in surface view. |
| Cremei | Colony on CYA, 25 °C, 7 days: Greyish-yellow, smooth texture; brownish-cream reverse. Biseriate, radiated conidial heads separated into columns, smooth and colorless wall stipe, globose or subglobose conidia, smooth to rough. |
| Fumigati | Colony on CYA, 25 °C, 7 days: Greyish-turquoise; velutinous texture; creamy reverse. Uniseriate, predominantly pyriform vesicles, grey to blue-green conidia. |
| Clavati | Colony on CYA, 25 °C, 7 days: Blue-green or pale blue-green; velvety texture; uncolored or dull tan reverse. Presence of exudates in some isolates but no soluble pigments. Uniseriate conidiophores with blue-green conidia, hyaline conidiophore stipes, and clavate aspergilla. |
| Nigri | Colony on CYA, 25 °C, 7 days: Brown to black; floccose texture, sulcate; beige to cream-yellow, brown, dark grey, uncolored to yellow reverse. Uniseriate or biseriate, spherical vesicles, smooth stipes, rough dark globose, finely rough, or finely spiny conidia. |
| Candidi | Colony on CYA, 25 °C, 7 days: Light cream; submerged texture; light brown reverse. Biseriate conidial heads, radiate, thick-walled conidiophores, septate, elongated vesicles, small conidia globose to subglobose, slightly roughened. |
| Flavipedes | Colony on CYA, 25 °C, 7 days: Yellowish white; floccose surface texture; brown to dark brown reverse. Biseriate conidiophores, radiate, elongated or subglobose vesicles, globose to subglobose conidia, smooth to finely roughened. |
| Circumdati | Colony on CYA, 25 °C, 7 days: Yellow to ochre colonies, cream, or pale yellow; smooth texture; yellow, orange, or cream reverse. Presence of pink to purple-brown sclerotia in some isolates. Predominantly biseriate, rough stems, smooth or finely rough globose conidia. |
| Municipality | Simpson Index | Shannon Index |
|---|---|---|
| Cerro | 0.0864 | −1.6688 |
| Arroyo Naranjo | 0.0930 | −1.1638 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sánchez Espinosa, K.C.; Almaguer Chávez, M.; Duarte-Escalante, E.; Rojas Flores, T.I.; Frías-De-León, M.G.; Reyes-Montes, M.d.R. Phylogenetic Identification, Diversity, and Richness of Aspergillus from Homes in Havana, Cuba. Microorganisms 2021, 9, 115. https://doi.org/10.3390/microorganisms9010115
Sánchez Espinosa KC, Almaguer Chávez M, Duarte-Escalante E, Rojas Flores TI, Frías-De-León MG, Reyes-Montes MdR. Phylogenetic Identification, Diversity, and Richness of Aspergillus from Homes in Havana, Cuba. Microorganisms. 2021; 9(1):115. https://doi.org/10.3390/microorganisms9010115
Chicago/Turabian StyleSánchez Espinosa, Kenia C., Michel Almaguer Chávez, Esperanza Duarte-Escalante, Teresa Irene Rojas Flores, María Guadalupe Frías-De-León, and María del Rocío Reyes-Montes. 2021. "Phylogenetic Identification, Diversity, and Richness of Aspergillus from Homes in Havana, Cuba" Microorganisms 9, no. 1: 115. https://doi.org/10.3390/microorganisms9010115