Molecular Mechanisms Involved in the Multicellular Growth of Ustilaginomycetes

Abstract
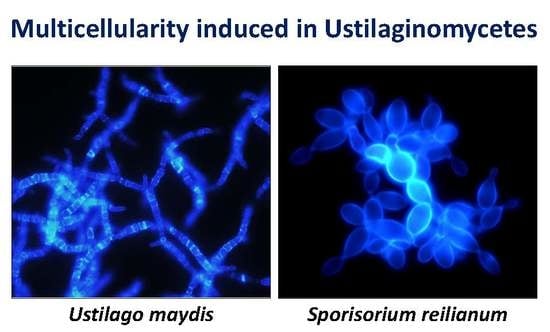
:
1. Introduction
2. Signaling Pathways Involved in the Ustilaginomycetes Multicellular Growth
2.1. MAPK Pathways
2.2. cAMP/PKA Pathway
3. The Role of Polyamines in the Multicellular Growth of Ustilaginomycetes
4. Epigenetic Regulation of Multicellular Growth in Ustilaginomycetes
5. The Cell Wall in the Multicellular Growth of Ustilaginomycetes
6. Polarized Growth during the Multicellularity of Ustilaginomycetes
7. Other Cellular Processes Involved in the Ustilaginomycetes Multicellular Growth
8. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Maynard-Smith, J.; Szathmary, E. Major Transitions in Evolution; Spektrum: New York, NY, USA, 1995. [Google Scholar]
- Bonner, J.T. The origins of multicellularity. Integr. Biol. 1998, 1, 27–36. [Google Scholar] [CrossRef]
- Pfeiffer, T.; Bonhoeffer, S. An evolutionary scenario for the transition to undifferentiated multicellularity. Proc. Natl. Acad. Sci. USA 2003, 100, 1095–1098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rokas, A. The origins of multicellularity and the early history of the genetic toolkit for animal development. Annu. Rev. Genet. 2008, 42, 235–251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rivera-Yoshida, N.; Arias Del Angel, J.A.; Benítez, M. Microbial multicellular development: Mechanical forces in action. Curr. Opin. Genet. Dev. 2018, 51, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Kessin, R.H.; Gundersen, G.G.; Zaydfudim, V.; Grimson, M. How cellular slime molds evade nematodes. Proc. Natl. Acad. Sci. USA 1996, 93, 4857–4861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boraas, M.; Seale, D.; Boxhorn, J. Phagotrophy by a flagellate selects for colonial prey: A posible origin of multicellularity. Evol. Ecol. 1998, 12, 153–164. [Google Scholar] [CrossRef]
- Smukalla, S.; Caldara, M.; Pochet, N.; Beauvais, A.; Guadagnini, S.; Yan, C.; Vinces, M.D.; Jansen, A.; Prevost, M.C.; Latgé, J.P.; et al. FLO1 is a variable green beard gene that drives biofilm-like cooperation in budding yeast. Cell 2008, 135, 726–737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knoll, A. The multiple origins of complex multicellularity. Annu. Rev. Earth Planet. Sci. 2011, 39, 217–239. [Google Scholar] [CrossRef] [Green Version]
- Koschwanez, J.H.; Foster, K.R.; Murray, A. Sucrose utilization in budding yeast as a model for the origin of undifferentiated multicellularity. PLoS Biol. 2011, 9, e1001122. [Google Scholar] [CrossRef]
- Oud, B.; Guadalupe-Medina, V.; Nijkamp, J.F.; de Ridder, D.; Pronk, J.T.; van Maris, A.J.; Daran, J.M. Genome duplication and mutations in ACE2 cause multicellular, fast-sedimenting phenotypes in evolved Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 2013, 110, E4223–E4231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arias Del Angel, J.A.; Escalante, A.; Martínez-Castilla, L.; Benítez, M. An evo-devo perspective on multicelular development of Myxobacteria. J. Exp. Zool. Part. B Mol. Dev. Evol. 2017, 328, 165–178. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Soto, D.; Velez-Haro, J.M.; León-Ramírez, C.G.; Galán-Vásquez, E.; Chávez-Munguía, B.; Ruiz-Herrera, J. Multicellular growth of the Basidiomycota phytopathogen fungus Sporisorium reilianum induced by acid conditions. Folia Microbiol. (Praha) 2020, 65, 511–521. [Google Scholar] [CrossRef] [PubMed]
- Rensing, S.A. (Why) Does evolution favour embryiogenesis? Trends Plant. Sci. 2016, 21, 562–573. [Google Scholar] [CrossRef]
- Pentz, J.T.; Marquez-Zacarías, P.; Yunker, P.J.; Libby, E.; Ratcliff, W.C. Ecological advantages and evolutionary limitations of aggregative multicellular development. bioRxiv 2018, 255307. [Google Scholar] [CrossRef] [Green Version]
- Kiss, E.; Hegedüs, B.; Virágh, M.; Varga, T.; Merényi, Z.; Kószó, T.; Bálint, B.; Prasanna, A.N.; Krizsán, K.; Kocsubé, S.; et al. Comparative genomics reveals the origin of fungal hyphae and multicellularity. Nat. Commun. 2019, 10, 4080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagy, L.G.; Kovács, G.M.; Krizsan, K. Complex multicellularity in fungi: Evolutionary convergence, single origin, or both? Biol Rev. Camb Philos Soc. 2018, 93, 1778–1794. [Google Scholar] [CrossRef]
- Harris, S.D. Cell polarity in filamentous fungi: Shaping the mold. Intl. Rev. Cytol. 2006, 251, 41–77. [Google Scholar]
- Fischer, M.S.; Glass, N.L. Communicate and fuse: How filamentous fungi establish and maintain an interconnected mycelial network. Front. Microbiol. 2019, 10, 619. [Google Scholar] [CrossRef]
- Goncalves, A.P.; Heller, J.; Span, E.A.; Rosenfield, G.; Do, H.P.; Palma-Guerrero, J.; Requena, N.; Marletta, M.A.; Glass, N.L. Allorecognition upon fungal cell-cell contact determines social cooperation and impacts the acquisition of multicellularity. Curr. Biol. 2019, 29, 3006–3017. [Google Scholar] [CrossRef]
- Kües, U. Life history and developmental processes in the basidiomycete Coprinus cinereus. Microbiol. Mol. Biol. Rev. 2000, 64, 316–353. [Google Scholar] [CrossRef] [Green Version]
- Gancedo, J.M. Control of pseudohyphae formation in Saccharomyces cerevisiae. FEMS Microbiol. Rev. 2001, 25, 107–123. [Google Scholar] [CrossRef]
- Ruiz-Herrera, J.; Sentandreu, R. Different effectors of dimorphism in Yarrowia lipolytica. Arch. Microbiol. 2002, 178, 477–483. [Google Scholar] [CrossRef] [PubMed]
- Kelly, M.T.; MacCallum, D.M.; Clancy, S.D.; Odds, F.C.; Brown, A.J.; Butler, G. The Candida albicans CaACE2 gene affects morphogenesis, adherence and virulence. Mol. Microbiol. 2004, 53, 969–983. [Google Scholar] [CrossRef] [PubMed]
- Ratcliff, W.C.; Frankhauser, J.D.; Rogers, D.W.; Greig, D.; Travisano, M. Origins of multicellular evolvability in snowflake yeast. Nat. Commun 2015, 6, 6102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, M.-Q.; Zhao, R.-L.; Hyde, K.D.; Begerow, D.; Kemler, M.; Yurkov, A.; McKenzie, E.H.C.; Raspé, O.; Kakishima, M.; Sánchez-Ramírez, S.; et al. Notes, outline and divergence times of Basidiomycota. Fungal Divers. 2019, 99, 105–367. [Google Scholar] [CrossRef] [Green Version]
- Snetselaar, K.M.; Mims, C.W. Light and electron microscopy of Ustilago maydis hyphae in maize. Mycol. Res. 1994, 98, 347–355. [Google Scholar] [CrossRef]
- Martinez-Espinoza, A.D.; Gerhardt, S.A.; Sherwood, J.E. Morphological and mutational analysis of mating in Ustilago hordei. Exp. Mycol. 1993, 17, 200–214. [Google Scholar] [CrossRef]
- Martinez, C.; Roux, C.; Dargent, R. Biotrophic development of Sporisorium reilianum f. sp. zeae in vegetative shoot apex of maize. Phytopathology 1999, 89, 247–253. [Google Scholar] [CrossRef] [Green Version]
- Martinez, C.; Roux, C.; Jauneau, A.; Dargent, R. The biological cycle of Sporisorium reilianum f. sp. zeae: An overview using microscopy. Mycologia 2002, 94, 505–514. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Soto, D.; Velez-Haro, J.M.; León-Ramírez, C.G.; Ruiz-Medrano, R.; Xoconostle-Cázares, B.; Ruiz-Herrera, J. The cereal phytopathogen Sporisorium reilianum is able to infect the non-natural host Arabidopsis thaliana. Eur. J. Plant. Pathol. 2019, 153, 417–427. [Google Scholar] [CrossRef]
- Brefort, T.; Doehlemann, G.; Mendoza-Mendoza, A.; Reissmann, S.; Djamei, A.; Kahmann, R. Ustilago maydis as a pathogen. Annu. Rev. Phytopathol. 2009, 47, 423–445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhaskaran, S.; Smith, R.H. Carbohydrates, invertase activity, growth and dimorphism in Sporisorium reilianum. Mycopathologia 1993, 122, 35–41. [Google Scholar] [CrossRef]
- Banuett, F.; Herskowitz, I. Morphological transitions in the life cycle of Ustilago maydis and their genetic control by the a and b loci. Exp. Mycol. 1994, 18, 247–266. [Google Scholar] [CrossRef]
- Ruiz-Herrera, J.; León-Ramírez, C.G.; Guevara-Olvera, L.; Cárabez-Trejo, A. Yeast-mycelial dimorphism of haploid and diploid strains of Ustilago maydis. Microbiology 1995, 141, 695–703. [Google Scholar] [CrossRef] [Green Version]
- Klose, J.; de Sá, M.M.; Kronstad, J.W. Lipid-induced filamentous growth in Ustilago maydis. Mol. Microbiol. 2004, 52, 823–835. [Google Scholar] [CrossRef] [PubMed]
- Zapata-Morín, P.A.; Fuetes-Dávila, G.; Adame-Rodríguez, J.M.; Aréchiga-Carvajal, E.T. Effect of pH and carbon source on the vegetative growth of Ustilago cynodontis (Pass.) Henn. in a solid and liquid culture médium. Rev. Mex. Fitopatol. 2010, 28, 159–161. [Google Scholar]
- Poloni, A.; Schirawski, J. Host specificity in Sporisorium reilianum is determined by distinct mechanisms in maize and sorghum. Mol. Plant. Pathol. 2016, 17, 741–754. [Google Scholar] [CrossRef]
- Sabbagh, S.K.; Diagne-Léye, G.; Naudan, M.; Roux, C.P. Solopathogenic strain formation strongly differs among Ustilaginaceae species. FEMS Microbiol. Lett. 2010, 305, 121–127. [Google Scholar] [CrossRef] [Green Version]
- Cabrera-Ponce, J.L.; León-Ramírez, C.G.; Verver-Vargas, A.; Palma-Tirado, L.; Ruiz-Herrera, J. Metamorphosis of the Basidiomycota Ustilago maydis: Transformation of yeast-like cells into basidiocarps. Fungal Genet. Biol. 2012, 49, 765–771. [Google Scholar] [CrossRef]
- León-Ramírez, C.G.; Cabrera-Ponce, J.L.; Martínez-Soto, D.; Sánchez-Arreguin, A.; Aréchiga-Carvajal, E.T.; Ruiz-Herrera, J. Transcriptomic analysis of basidiocarp development in Ustilago maydis (DC) Cda. Fungal Genet. Biol. 2017, 101, 34–45. [Google Scholar] [CrossRef]
- Gold, S.; Duncan, G.; Barrett, K.; Kronstad, J. cAMP regulates morphogenesis in the fungal pathogen Ustilago maydis. Genes Dev. 1994, 8, 2805–2816. [Google Scholar] [CrossRef] [Green Version]
- Dürrenberger, F.; Wong, K.; Kronstad, J.W. Identification of a cAMP-dependent protein kinase catalytic subunit required for virulence and morphogenesis in Ustilago maydis. Proc. Natl. Acad. Sci. USA 1998, 95, 5684–5689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez-Espinoza, A.D.; Ruiz-Herrera, J.; León-Ramírez, C.G.; Gold, S.E. MAP kinase and cAMP signaling pathways modulate the pH-induced yeast-to-mycelium dimorphic transition in the corn smut fungus Ustilago maydis. Curr. Microbiol. 2004, 49, 274–281. [Google Scholar] [CrossRef]
- Martínez-Soto, D.; Ruiz-Herrera, J. Functional analysis of the MAPK pathways in fungi. Rev. Iberoam. Micol. 2017, 34, 192–202. [Google Scholar] [CrossRef]
- Guevara-Olvera, L.; Xoconostle-Cázares, B.; Ruiz-Herrera, J. Cloning and disruption of the ornithine decarboxylase gene of Ustilago maydis: Evidence for a role of polyamines in its dimorphic transition. Microbiology 1997, 143, 2237–2245. [Google Scholar] [CrossRef] [Green Version]
- Valdés-Santiago, L.; Ruiz-Herrera, J. Stress and polyamine metabolism in fungi. Front. Chem. 2014, 10, 42. [Google Scholar] [CrossRef] [Green Version]
- González-Prieto, J.M.; Rosas-Quijano, R.; Domínguez, A.; Ruiz-Herrera, J. The UmGcn5 gene encoding histone acetyltransferase from Ustilago maydis is involved in dimorphism and virulence. Fungal Genet. Biol. 2014, 71, 86–95. [Google Scholar] [CrossRef]
- Martínez-Soto, D.; González-Prieto, J.M.; Ruiz-Herrera, J. Transcriptomic analysis of the GCN5 gene reveals mechanisms of the epigenetic regulation of virulence and morphogenesis in Ustilago maydis. FEMS Yeast Res. 2015, 15, fov055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elías-Villalobos, A.; Fernández-Álvarez, A.; Moreno-Sánchez, I.; Helmlinger, D.; Ibeas, J.I. The Hos2 histone deacetylase controls Ustilago maydis virulence through direct regulation of mating-type genes. PLoS Pathog. 2015, 11, e1005134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elías-Villalobos, A.; Dominique-Helmlinger, D.; Ibeas, J.I. Histone deacetylases: Revealing the molecular base of dimorphism in pathogenic fungi. Microb. Cell 2015, 2, 491–493. [Google Scholar] [CrossRef] [Green Version]
- Robledo-Briones, M.; Ruiz Herrera, J. Transcriptional regulation of the genes encoding chitin and β-1, 3-glucan synthases from Ustilago maydis. Curr. Microbiol. 2012, 65, 85–90. [Google Scholar] [CrossRef]
- Robledo-Briones, M.; Ruiz Herrera, J. Regulation of genes involved in cell wall synthesis and structure during Ustilago maydis dimorphism. FEMS Yeast Res. 2013, 13, 74–84. [Google Scholar] [CrossRef] [Green Version]
- Langner, T.; Öztürk, M.; Hartmann, S.; Cord-Landwehr, S.; Moerschbacher, B.; Walton, J.D.; Göhre, V. Chitinases Are essential for cell separation in Ustilago maydis. Eukaryot. Cell 2015, 14, 846–857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klosterman, S.J.; Perlin, M.H.; Garcia-Pedrajas, M.; Covert, S.F.; Gold, S.E. Genetics of morphoenesis and pathogenic development of Ustilago maydis. Adv. Genet. 2007, 57, 1–47. [Google Scholar] [CrossRef] [PubMed]
- Vollmeister, E.; Schipper, K.; Baumann, S.; Haag, C.; Pohlmann, T.; Janpeter Stock, J.; Feldbrügge, M. Fungal development of the plant pathogen Ustilago maydis. FEMS Microbiol Rev. 2012, 36, 59–77. [Google Scholar] [CrossRef] [Green Version]
- Valdés-Santiago, L.; Cervantes-Chávez, J.A.; Ruiz-Herrera, J. Ustilago maydis spermidine synthase is encoded by a chimeric gene, required for morphogenesis, and indispensable for survival in the host. FEMS Yeast Res. 2009, 9, 923–935. [Google Scholar] [CrossRef] [Green Version]
- Valdés-Santiago, L.; Guzmán-de-Peña, D.; Ruiz-Herrera, J. Life without putrescine: Disruption of the gene-encoding polyamine oxidase in Ustilago maydis odc mutants. FEMS Yeast Res. 2010, 10, 928–940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valdés-Santiago, L.; Cervantes-Chávez, J.A.; Winkler, R.; León-Ramírez, C.G.; Ruiz-Herrera, J. Phenotypic comparison of Samdc and Spe mutants reveals complex relationships of polyamine metabolism in Ustilago Maydis. Microbiology 2012, 158, 674–684. [Google Scholar] [CrossRef]
- Aréchiga-Carvajal, E.T.; Ruiz-Herrera, J. The RIM101/pacC homologue from the Basidiomycete Ustilago maydis is functional in multiple pH-sensitive phenomena. Eukaryot Cell. 2005, 4, 999–1008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cervantes-Chávez, J.A.; Ortiz-Castellanos, L.; Tejeda-Sartorius, M.; Gold, S.; Ruiz-Herrera, J. Functional analysis of the pH responsive pathway Pal/Rim in the phytopathogenic Basidiomycete Ustilago maydis. Fungal Genet. Biol. 2010, 47, 446–457. [Google Scholar] [CrossRef] [PubMed]
- Cervantes-Montelongo, J.A.; Ruiz-Herrera, J. Identification of a novel member of the pH responsive pathway Pal/Rim in Ustilago maydis. J. Basic Microbiol. 2019, 59, 14–23. [Google Scholar] [CrossRef] [Green Version]
- Méndez-Morán, L.; Reynaga-Peña, C.G.; Springer, P.S.; Ruiz-Herrera, J. Ustilago maydis Infection of the nonnatural host Arabidopsis thaliana. Phytopatholoy 2005, 95, 480–488. [Google Scholar] [CrossRef]
- Martínez-Soto, D.; Robledo-Briones, A.M.; Estrada-Luna, A.A.; Ruiz-Herrera, J. Transcriptomic analysis of Ustilago maydis infecting Arabidopsis reveals important aspects of the fungus pathogenic mechanisms. Plant. Signal. Behav. 2013, 8, e25059. [Google Scholar] [CrossRef] [Green Version]
- Peñalva, M.A.; Arst, H.N., Jr. Recent advances in the characterization of ambient pH regulation of gene expression in filamentous fungi and yeasts. Annu. Rev. Microbiol. 2004, 58, 425–451. [Google Scholar] [CrossRef]
- Kämper, J.; Kahmann, R.; Bölker, M.; Ma, L.-J.; Brefort, T.; Saville, B.J.; Banuett, F.; Kronstad, J.W.; Gold, S.E.; Müller, O.; et al. Insights from the genome of the biotrophic fungal plant pathogen Ustilago maydis. Nature 2006, 444, 97–101. [Google Scholar] [CrossRef]
- Laurie, J.D.; Ali, S.; Linning, R.; Mannhaupt, G.; Wong, P.; Güldener, U.; Münsterkötter, M.; Moore, R.; Kahmann, R.; Bakkeren, G.; et al. Genome comparison of barley and maize smut fungi reveals targeted loss of RNA silencing components and species-specific presence of transposable elements. Plant. Cell. 2012, 24, 1733–1745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rabe, F.; Bosch, J.; Stirnberg, A.; Guse, T.; Bauer, L.; Seitner, D.; Rabanal, F.A.; Czedik-Eysenberg, A.; Uhse, S.; Bindics, J.; et al. A complete toolset for the study of Ustilago bromivora and Brachypodium sp. as a fungal-temperate grass pathosystem. Elife 2016, 5, e20522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schirawski, J.; Mannhaupt, G.; Münch, K.; Brefort, T.; Schipper, K.; Doehlemann, G.; Di Stasio, M.; Rössel, N.; Mendoza-Mendoza, A.; Pester, D.; et al. Pathogenicity determinants in smut fungi revealed by genome comparison. Science 2010, 330, 1546–1548. [Google Scholar] [CrossRef]
- Solano-González, S.; Darby, A.C.; Cossar, D.; Caddick, M.X. High-quality draft genome sequence and annotation of the Basidiomycete yeast Sporisorium graminicola CBS10092, a producer of mannosylerythritol lipids. Microbiol Resour Announc. 2019, 8, e00479-19. [Google Scholar] [CrossRef] [Green Version]
- Kijpornyongpan, T.; Mondo, S.J.; Barry, K.; Sandor, L.; Lee, J.; Lipzen, A.; Pangilinan, J.; LaButti, K.; Hainaut, M.; Henrissat, B.; et al. Broad Genomic Sampling Reveals a Smut Pathogenic Ancestry of the Fungal Clade Ustilaginomycotina. Mol. Biol. Evol. 2018, 35, 1840–1854. [Google Scholar] [CrossRef] [Green Version]
- Banuett, F. Signalling in the yeasts: An informational cascade with links to thefilamentous fungi. Microbiol. Mol. Biol. Rev. 1998, 62, 249–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dohlman, H.G. Thematic minireview series: Complexities of cellular signaling revealed by simple model organisms. J. Biol. Chem. 2016, 291, 7786–7787. [Google Scholar] [CrossRef] [Green Version]
- Correia, I.; Román, E.; Prieto, D.; Eisman, B.; Pla, J. Complementary roles of the cek1 and cek2 map kinases in Candida albicans cell-wall biogénesis. Future Microbiol. 2016, 11, 51–67. [Google Scholar] [CrossRef]
- Davidson, R.C.; Nichols, C.B.; Cox, G.M.; Perfect, J.R.; Heitman, J. A MAP kinase cascade composed of cell type specific and nonspecific elements controls mating and differentiation of the fungal pathogen Cryptococcus neoformans. Mol. Microbiol. 2003, 49, 469–485. [Google Scholar] [CrossRef]
- Leberer, E.; Harcus, D.; Broadbent, I.D.; Clark, K.L.; Dignard, D.; Ziegelbauer, K.; Schmidt, A.; Gow, N.A.; Brown, A.J.; Thomas, D.Y. Signal transduction through homologs of the Ste20p and Ste7p protein kinases can trigger hyphal formation in the pathogenic fungus Candida albicans. Proc. Natl. Acad. Sci. USA 1996, 93, 13217–13222. [Google Scholar] [CrossRef] [Green Version]
- Pan, X.; Harashima, T.; Heitman, J. Signal transduction cascades regulating pseudohyphal differentiation of Saccharomyces cerevisiae. Curr. Opin. Microbiol. 2000, 3, 567–572. [Google Scholar] [CrossRef]
- Biswas, S.; Van Dijck, P.; Datta, A. Environmental sensing and signal transduction pathways regulating morphopathogenic determinants of Candida albicans. Microbiol. Mol. Biol. Rev. 2007, 71, 348–376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klosterman, S.J.; Martinez-Espinoza, A.D.; Andrews, D.L.; Seay, J.R.; Gold, S.E. UBC2, an ortholog of the yeast Ste50p adaptor, possesses a Basidiomycete-specific carboxy terminal extension essential for pathogenicity independent of pheromone response. Mol. Plant. Microbe Interact. 2008, 21, 110–121. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Soto, D.; Ruiz-Herrera, J. Regulation of the expression of the whole genome of Ustilago maydis by a MAPK pathway. Arch. Microbiol. 2015, 197, 575–588. [Google Scholar] [CrossRef] [PubMed]
- Banuett, F.; Herskowitz, I. Identification of fuz7, a Ustilago maydis MEK/MAPKK homolog required for a-locus-dependent and -independent steps in the fungal life cycle. Genes Devev. 1994b, 8, 1367–1378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayorga, M.E.; Gold, S.E. Characterization and molecular genetic complementation of mutants affecting dimorphism in the fungus Ustilago maydis. Fungal Genet. Biol. 1998, 24, 364–376. [Google Scholar] [CrossRef] [PubMed]
- Mayorga, M.E.; Gold, S.E. A MAP kinase encoded by the ubc3 gene of Ustilago maydis is required for filamentous growth and full virulence. Mol. Microbiol. 1999, 34, 485–497. [Google Scholar] [CrossRef] [PubMed]
- Andrews, D.L.; Egan, J.D.; Mayorga, M.E.; Gold, S.E. The Ustilago maydis ubc4 and ubc5 genes encode members of a MAP Kinase cascade required for filamentous growth. Mol. Plant. Microbe Interact. 2000, 13, 781–786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Müller, P.; Aichinger, C.; Feldbrügge, M.; Kahmann, R. The MAP Kinase kpp2 regulates mating and pathogenic development in Ustilago maydis. Mol. Microbiol. 1999, 34, 1007–1017. [Google Scholar] [CrossRef] [PubMed]
- Müller, P.; Weinzierl, G.; Brachmann, A.; Feldbrügge, M.; Kahmann, R. Mating and pathogenic development of the smut fungus Ustilago maydis are regulated by one mitogen-activated protein kinase cascade. Eukaryot. Cell 2003, 2, 1187–1199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, Y.Z.; Zhang, B.; Chang, C.; Wang, Y.; Lu, S.; Sun, S.; Zhang, X.; Chen, B.; Jiang, Z. The MAP Kinase SsKpp2 is required for mating/filamentation in Sporisorium scitamineum. Front. Microbiol. 2018, 9, 2555. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Ge, Q.; Cao, Q.; Cui, H.; Hu, P.; Yu, X.; Ye, Z. Cloning and characterization of two mapk genes Uekpp2 and Uekpp6 in Ustilago esculenta. Curr. Microbiol. 2018, 75, 1016–1024. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Fink, G.R. Feedback control of morphogenesis in fungi by aromatic alcohols. Genes Dev. 2006, 20, 1150–1161. [Google Scholar] [CrossRef] [Green Version]
- Wongsuk, T.; Pumeesat, P.; Luplertlop, N. Fungal quorum sensingmolecules: Role in fungal morphogenesis and pathogenicity. J. Basic. Microbiol. 2016, 56, 440–447. [Google Scholar] [CrossRef]
- Mayorga, M.E.; Gold, S.E. The ubc2 gene of Ustilago maydis encodes a putative novel adaptor protein required for filamentous growth, pheromone response and virulence. Mol. Microbiol. 2001, 41, 1365–1379. [Google Scholar] [CrossRef]
- Müller, P.; Katzenberger, J.D.; Loubradou, G.; Kahmann, R. Guanyl nucleotide exchange factor Sql2 and Ras2 regulate filamentous growth in Ustilago maydis. Eukaryot. Cell 2003, 2, 609–617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, N.; Kronstad, J.W. Ras2 controls morphogenesis, pheromone response, and pathogenicity in the fungal pathogen Ustilago maydis. Eukaryot. Cell 2002, 1, 954–966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mösch, H.U.; Roberts, R.L.; Fink, G.R. Ras2 signals via the cdc42/ste20/mitogen-activated protein kinase module to induce filamentous growth in Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 1996, 93, 5352–5356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garrido, E.; Pérez-Martín, J. The crk1 gene encodes an Ime2-related protein that is required for morphogenesis in the plant pathogen Ustilago maydis. Mol. Microbiol. 2003, 47, 729–743. [Google Scholar] [CrossRef]
- Cheng, C.K.; Au, C.H.; Wilke, S.K.; Stajich, J.E.; Miriam, E.; Zolan, M.E.; Pukkila, P.J.; Kwan, H.S. 5′-Serial analysis of gene expression studies reveal a transcriptomic switch during fruiting body development in Coprinopsis Cinerea. BMC Genomics 2013, 14, 195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilman, A.G. G proteins and dual control of adenylate cyclase. Cell 1984, 36, 577–579. [Google Scholar] [CrossRef]
- Taylor, S.S.; Juechler, J.A.; Yonemoto, W. cAMP-dependent protein kinase: Framework for a diverse family of regulatory enzymes. Annu. Rev. Biochem. 1990, 59, 971–1005. [Google Scholar] [CrossRef]
- Levitzki, A.; Bar-Sinai, A. The regulation of Adenylyl cyclase by receptor-perated G proteins. Pharmacol. Ther. 1991, 50, 271–283. [Google Scholar] [CrossRef]
- Thevelein, J.M.; Cauwenberg, L.; Colombo, S.; De Winde, J.H.; Donation, M.; Dumortier, F.; Kraakman, L.; Lemaire, K.; Ma, P.; Nauwelaers, D.; et al. Nutrient-induced signal transduction through the protein kinase a pathway and its role in the control of metabolism, stress resistance, and growth in yeast. Enzyme Microb. Technol. 2000, 26, 819–825. [Google Scholar] [CrossRef]
- Caza, M.; Kronstad, J.W. The cAMP/protein kinase a pathway regulates virulence and adaptation to host conditions in Cryptococcus neoformans. Front. Cell Infect. Microbiol. 2019, 9, 212. [Google Scholar] [CrossRef]
- Sabie, F.T.; Gadd, G.M. Effect of Nucleosides and Nucleotides and the relationship between cellular adenosine 3′:5′-cyclic monophosphate (Cyclic AMP) and germ tube formation in Candida albicans. Mycopathologia 1992, 119, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Lichter, A.; Mills, D. Fil1, a G-protein alpha-subunit that acts upstream of camp and is essential for dimorphic switching in haploid cells of Ustilago hordei. Mol. Gen. Genet. 1997, 256, 26–35. [Google Scholar] [CrossRef]
- Chang, C.; Enping Cai, E.; Deng, Y.Z.; Mei, D.; Qiu, S.; Chen, B.; Zhang, L.H.; Jiang, Z. cAMP/PKA signalling pathway regulates redox homeostasis essential for Sporisorium scitamineum mating/filamentation and virulence. Environ. Microbiol. 2019, 21, 959–971. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Deng, Y.Z.; Cui, G.; Huang, C.; Zhang, B.; Chang, C.; Jiang, Z.; Zhang, L.H. The AGC Kinase SsAgc1 regulates Sporisorium scitamineum mating/filamentation and pathogenicity. mSphere 2019, 4, e00259-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dürrenberger, F.; Laidlaw, R.D.; Kronstad, J.W. The hgl1 gene is required for dimorphism and teliospore formation in the fungal pathogen Ustilago maydis. Mol. Microbiol. 2001, 41, 337–348. [Google Scholar] [CrossRef]
- Loubradou, G.; Brachmann, A.; Feldbrugge, M.; Kahmann, R. A homologue of the transcriptional repressor Ssn6p antagonizes cAMP signalling in Ustilago maydis. Mol. Microbiol. 2001, 40, 719–730. [Google Scholar] [CrossRef]
- Schwalb, M.N. Regulation of fruiting. In Genetics and Morphogenesis in the Basidiomicetes; Schwalb, M.N., Miles, P.G., Eds.; Academic Press: New York, NY, USA, 1978; pp. 135–165. [Google Scholar]
- Swamy, S.; Uno, I.; Ishikawa, T. Regulation of cyclic AMP-dependent phosphorylation of cellular proteins by the incompatibility factors in Coprinus cinereus. J. Gen. Appl. Microbiol. 1985, 31, 339–346. [Google Scholar] [CrossRef] [Green Version]
- Swamy, S.; Uno, I.; Ishikawa, T. Regulation of cyclic AMP metabolism by the incompatibility factors in Coprinus cinereus. J. Gen. Microbiol. 1985, 131, 3211–3217. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.; Lian, L.; Guo, L.; Xie, B.; Wang, W.; Chen, B.; van Peer, A.F.; Li, S.; Wu, T.; Xie, B. The accordant trend of both parameters (rgs expression and cAMP content) follows the pattern of development of fruiting body in Volvariella volvácea. Curr. Microbiol. 2015, 71, 579–784. [Google Scholar] [CrossRef]
- Tabor, C.W.; Tabor, H. Polyamines. Annu. Rev. Biochem. 1984, 53, 749–790. [Google Scholar] [CrossRef]
- D’Agostino, L.; Di Pietro, M.; Di Luccia, A. Nuclear aggregates of polyamines. IUBMB Life 2006, 58, 75–82. [Google Scholar] [CrossRef] [Green Version]
- Igarashi, K.; Kashiwagi, K. Modulation of cellular function by polyamines. Int. J. Biochem. Cell B. 2010, 42, 39–51. [Google Scholar] [CrossRef] [PubMed]
- Rajam, M.V.; Galston, A.W. The effects of some polyamine biosynthetic inhibitors on growth and morphology of phytopathogenic fungi. Plant. Cell Physiol. 1985, 26, 683–692. [Google Scholar] [CrossRef]
- Ruiz-Herrera, J. Polyamines, DNA methylation, and fungal differentiation. Crit. Rev. Microbiol. 1994, 20, 143–150. [Google Scholar] [CrossRef]
- Khurana, N.; Saxena, R.K.; Gupta, R.; Rajam, M.V. Polyamines as modulators of microcycle conidiation in Aspergillus flavus. Microbiology 1996, 142, 517–523. [Google Scholar] [CrossRef] [Green Version]
- Mueller, E.; Bailey, A.; Corran, A.; Michael, A.J.; Bowyer, P. Ornithine decarboxylase knockout in Tapesia yallundae abolishes infection plaque formation in vitro but does not reduce virulence toward wheat. Mol. Plant. Microbe Interact. 2001, 14, 1303–1311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pal, M.; Janda, T. Role of polyamine metabolism in plant pathogen interactions. J. Plant. Sci. Phytopathol. 2017, 1, 95–100. [Google Scholar] [CrossRef] [Green Version]
- Nickerson, K.W.; Dunkle, L.D.; Van Etten, J.L. Absence of spermine in filamentous fungi. J. Bacteriol. 1977, 129, 173–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- León-Ramírez, C.G.; Valdés-Santiago, L.; Campos-Góngora, E.; Ortiz-Castellanos, L.; Aréchiga-Carvajal, E.T.; Ruiz-Herrera, J. A molecular probe for Basidiomycota: The spermidine synthase-saccharopine dehydrogenase chimeric gene. FEMS Microbiol. Lett. 2010, 312, 77–83. [Google Scholar] [CrossRef] [Green Version]
- Rocha, R.O.; Wilson, R.A. Essential, deadly, enigmatic: Polyamine metabolism and roles in fungal cells. Fungal. Biol. Rev. 2019, 33, 47–57. [Google Scholar] [CrossRef]
- Martínez-Soto, D.; Ruiz-Herrera, J. Transcriptomic analysis of the dimorphic transition of Ustilago maydis induced in vitro by a change in pH. Fungal Genet. Biol. 2013, 58–59, 116–125. [Google Scholar] [CrossRef]
- Kou, M.H.; Allis, C.D. Roles of histone acetyltransferases and deacetylases in gene regulation. BioEssays 1998, 20, 615–626. [Google Scholar] [CrossRef]
- Marmorstein, R.; Roth, S.Y. Histone acetyltransferases: Function, structure, and catalysis. Curr. Opin. Genet. Dev. 2001, 11, 155–161. [Google Scholar] [CrossRef]
- Legube, G.; Trouche, D. Regulating histone acetyltransferases and deacetylases. EMBO Rep. 2003, 4, 944–947. [Google Scholar] [CrossRef] [Green Version]
- Thiagalingam, S.; Cheng, K.H.; Lee, H.J.; Mineva, N.; Thiagalingam, A.; Ponte, J.F. Histone deacetylases: Unique players in shaping the epigenetic histone code. Ann. N. Y. Acad. Sci. 2003, 983, 84–100. [Google Scholar] [CrossRef]
- Lee, K.K.; Workman, J.L. Histone acetyltransferase complexes: One size doesn’t fit all. Nat. Rev. Mol. Cell Biol. 2007, 8, 284–295. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Yang, S.; Zhao, M.; Lou, M.; Yu, C.M.; Chen, C.Y.; Tai, R.; Wu, K. Transcriptional repression by histone deacetylases in plants. Mol. Plant. 2014, 7, 764–772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reyna-López, G.E.; Simpson, J.; Ruiz-Herrera, J. Differences in DNA methylation patterns are detectable during the dimorphic transition of fungi by amplification of restriction polymorphisms. Mol. Gen. Genet. 1997, 253, 703–710. [Google Scholar] [CrossRef]
- Reyna-López, G.E.; Ruiz-Herrera, J. Specificity of DNA methylation changes during fungal dimorphism and its relationship to polyamines. Curr. Microbiol. 2004, 48, 118–123. [Google Scholar] [CrossRef]
- Kusano, T.; Berberich, T.; Tateda, C.; Takahashi, Y. Polyamines: Essential factors for growth and survival. Planta 2008, 228, 367–381. [Google Scholar] [CrossRef]
- He, C.; Zhang, Z.; Li, B.; Tian, S. The pattern and function of DNA methylation in fungal plant pathogens. Microorganisms 2020, 8, 227. [Google Scholar] [CrossRef] [Green Version]
- Ruiz-Herrera, J.; Pérez-Rodríguez, F.; Velez-Haro, J. The signaling mechanisms involved in the dimorphic phenomenon of the Basidiomycota fungus Ustilago maydis. Int. Microbiol. 2020, 23, 121–126. [Google Scholar] [CrossRef]
- Ruiz-Herrera, J. Fungal Cell Wall Structure, Synthesis, and Assembly, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2012; p. 168. [Google Scholar]
- Ruiz-Herrera, J.; Elorza, M.V.; Álvarez, P.E.; Sentandreu, R. Biosynthesis of the fungal cell wall. In Pathogenic Fungi, Structural, Biology and Taxonomy; San-Blas, G., Calderone, R.A., Eds.; Caister Academic Press: Wymondham, UK, 2004; pp. 41–99. [Google Scholar]
- Sentandreu, R.; Elorza, M.V.; Valentín, E.; Ruiz-Herrera, J. The structure and composition of fungal cell wall. In Pathogenic Fungi, Structural, Biology and Taxonomy; San-Blas, G., Calderone, R.A., Eds.; Caister Academic Press: Wymondham, UK, 2004; pp. 3–27. [Google Scholar]
- Cabib, E.; Sburlati, A.; Bowers, B.; Silverman, S.J. Chitin synthase 1, an auxiliary enzyme for chitin synthesis in Saccharomyces cerevisiae. J. Cell Biol. 1989, 108, 1665–1672. [Google Scholar] [CrossRef]
- Silverman, S.J.; Sburlati, A.; Slater, M.L.; Cabib, E. Chitin synthase 2 is essential for septum formation and cell division in Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 1988, 85, 4735–4739. [Google Scholar] [CrossRef] [Green Version]
- Shaw, J.A.; Mol, P.C.; Bowers, B.; Silverman, S.J.; Valdivieso, M.H.; Durán, A.; Cabib, E. The function of chitin synthases 2 and 3 in the Saccharomyces cerevisiae cell cycle. J. Cell Biol. 1991, 114, 111–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yarden, O.; Yanofsky, C. chitin synthase 1 plays a major role in cell wall biogenesis in Neurospora crassa. Genes Dev. 1991, 5, 2420–2430. [Google Scholar] [CrossRef]
- Borgia, P.-T.; Iartchouk, N.; Riggle, P.J.; Winter, K.R.; Koltin, Y.; Bulawa, C.E. The chsB gene of Aspergillus nidulans is necessary for normal hyphal growth and development. Fungal Genet. Biol. 1996, 20, 193–203. [Google Scholar] [CrossRef]
- Garcerá-Teruel, A.; Xoconoxtle-Cázares, B.; Rosas-Quijano, R.; Ortíz, L.; León-Ramírez, C.; Specht, C.A.; Sentandreu, R.; Ruiz Herrera, J. Loss of virulence in Ustilago maydis by Umchs6 gene disruption. Res. Microbiol. 2004, 155, 87–97. [Google Scholar] [CrossRef]
- Weber, I.; Assmann, D.; Thines, E.; Steinberg, G. Polar localizing class V myosin chitin synthases are essential during early plant infection in the plant pathogenic fungus Ustilago maydis. Plant. Cell 2006, 18, 225–242. [Google Scholar] [CrossRef] [Green Version]
- López-Matas, M.A.; Eslava, A.P.; Díaz-Minguez, J.M. Mcchs1, a member of a chitin synthase gene family in Mucor circinelloides, is differentially expressed during dimorphism. Curr. Microbiol. 2000, 40, 169–175. [Google Scholar] [CrossRef]
- Niño-Vega, G.A.; Munro, C.A.; San-Blas, G.; Gooday, G.W.; Gow, N.A. Differential expression of chitin synthase genes during temperature-induced dimorphic transitions in Paracoccidioides Brasiliensis. Med. Mycol. 2000, 38, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Christodoulidou, A.; Bouriotis, V.; Thireos, G. Two sporulation-specific chitin deacetylase-encoding genes are required for the ascospore wall rigidity of Saccharomyces cerevisiae. J. Biol. Chem. 1996, 271, 31420–31425. [Google Scholar] [CrossRef] [Green Version]
- Geoghegan, I.A.; Gurr, S.J. Investigating chitin deacetylation and chitosan hydrolysis during vegetative growth in Magnaporthe oryzae. Cell Microbiol. 2017, 19, e12743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruiz-Herrera, J.; Robledo-Briones, M.; Martínez-Soto, D. Experimental pathosystems as a tool for the identification of virulence factors in pathogenic fungi. In Biotechnology: Beyond Borders; Deshpande, M.V., Ruiz-Herrera, J., Eds.; CSIR-National Chemical Laboratory: Pune, India, 2013; pp. 30–38. [Google Scholar]
- Tentler, S.; Palas, J.; Enderlin, C.; Campbell, J.; Taft, C.; Miller, T.K.; Wood, R.L.; Selitrennikoff, C.P. Inhibition of Neurospora crassa growth by a glucan synthase-1 antisense construct. Curr. Microbiol. 1997, 34, 303–308. [Google Scholar] [CrossRef] [PubMed]
- Verdín, J.; Bartnicki-Garcia, S.; Riquelme, M. Functional stratification of the Spitzenkörper of Neurospora crassa. Mol. Microbiol. 2009, 74, 1044–1053. [Google Scholar] [CrossRef]
- Kuranda, M.J.; Robbins, P.W. Chitinase is required for cell separation during growth of Saccharomyces cerevisiae. J. Biol. Chem. 1991, 266, 19758–19767. [Google Scholar]
- Colman-Lerner, A.; Chin, T.E.; Brent, R. Yeast Cbk1 and Mob2 activate daughter-specific genetic programs to induce asymmetric cell fates. Cell 2001, 107, 739–750. [Google Scholar] [CrossRef] [Green Version]
- Dünkler, A.; Walther, A.; Specht, C.A.; Wendland, J. Candida albicans Cht3 encodes the functional homolog of the cts1 chitinase of Saccharomyces cerevisiae. Fungal Genet. Biol. 2005, 42, 935–947. [Google Scholar] [CrossRef]
- Tzelepis, G.D.; Melin, P.; Jensen, D.F.; Stenlid, J.; Magnus Karlsson, M. Functional analysis of glycoside hydrolase family 18 and 20 genes in Neurospora crassa. Fungal Genet. Biol. 2012, 49, 717–730. [Google Scholar] [CrossRef]
- Baladrón, V.; Ufano, S.; Dueñas, E.; Martín-Cuadrado, A.B.; del Rey, F.; Vázquez de Aldana, C.R. Eng1p, an endo-1,3-beta-glucanase localized at the daughter side of the septum, is involved in cell separation in Saccharomyces cerevisiae. Eukaryot. Cell 2002, 1, 774–786. [Google Scholar] [CrossRef] [Green Version]
- Riquelme, M.; Aguirre, J.; Bartnicki-García, S.; Braus, G.H.; Feldbrügge, M.; Fleig, U.; Hansberg, W.; Herrera-Estrella, A.; Kämper, J.; Kück, U.; et al. Fungal morphogenesis, from the polarized growth of hyphae to complex reproduction and infection structures. Microbiol. Mol. Biol. Rev. 2018, 82, e00068-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banuett, F.; Herskowitz, I. Bud morphogenesis and the actin and microtubule cytoskeletons during budding in the corn smut fungus, Ustilago maydis. Fungal Genet. Biol. 2002, 37, 149–170. [Google Scholar] [CrossRef]
- Valinluck, M.; Ahlgren, S.; Sawada, M.; Locken, K.; Banuett, F. Role of the nuclear migration protein Lis1 in cell morphogenesis in Ustilago maydis. Mycologia 2010, 102, 493–512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valinluck, M.; Woraratanadharm, T.; Lu, C.Y.; Quintanilla, R.H., Jr.; Bannuett, F. The cell end marker Tea4 regulates morphogenesis and pathogenicity in the basidiomycete fungus Ustilago maydis. Fungal Genet. Biol. 2014, 66, 54–68. [Google Scholar] [CrossRef] [Green Version]
- Chew, E.; Aweiss, Y.; Lu, C.Y.; Banuett, F. Fuz1, a MYND domain protein, is required for cell morphogenesis in Ustilago maydis. Mycologia 2008, 100, 31–46. [Google Scholar] [CrossRef]
- Schuchardt, I.; Assmann, D.; Thines, E.; Schuberth, C.; Steinberg, G. Myosin-V, Kinesin-1, and Kinesin-3 cooperate in hyphal growth of the fungus Ustilago maydis. Mol. Biol. Cell. 2005, 16, 5191–5201. [Google Scholar] [CrossRef] [Green Version]
- Weber, I.; Gruber, C.; Steinberg, G. A class V myosin required for mating, hyphal growth, and pathogenicity in the dimorphic plant pathogen Ustilago maydis. Plant. Cell 2003, 15, 2826–2842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woraratanadharm, T.; Kmosek, S.; Banuett, F. UmTea1, a Kelch and BAR domain-containing protein, acts at the cell cortex to regulate cell morphogenesis in the dimorphic fungus Ustilago maydis. Fungal Genet. Biol. 2018, 121, 10–28. [Google Scholar] [CrossRef] [PubMed]
- Wösten, H.A.; Bohlmann, R.; Eckerskorn, C.; Lottspeich, F.; Bölker, M.; Kahmann, R. A novel class of small amphipathic peptides affect aerial hyphal growth and surface hydrophobicity in Ustilago maydis. EMBO J. 1996, 15, 4272–4281. [Google Scholar] [CrossRef]
- Teertstra, W.R.; Deelstra, H.J.; Vranes, M.; Bohlmann, R.; Kahmann, R.; Kämper, J.; Wösten, H.A.B. Repellents have functionally replaced hydrophobins in mediating attachment to a hydrophobic surface and in formation of hydrophobic aerial hyphae in Ustilago maydis. Microbiology 2006, 152, 3607–3612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soberanes-Gutiérrez, C.V.; Juárez-Montiel, M.; Olguín-Rodríguez, O.; Hernández-Rodríguez, C.; Ruiz-Herrera, J.; Villa-Tanaca, L. The Pep4 gene encoding proteinase A is involved in dimorphism and pathogenesis of Ustilago maydis. Mol. Plant. Pathol. 2015, 16, 837–846. [Google Scholar] [CrossRef]
- Pham, C.D.; Yu, Z.; Sandrock, B.; Bölker, M.; Gold, S.E.; Perlin, M.H. Ustilago maydis Rho1 and 14-3-3 homologues participate in pathways controlling cell separation and cell polarity. Eukaryot. Cell 2009, 8, 977–989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frieser, S.H.; Hlubek, A.; Sandrock, B.; Bölker, M. Cla4 kinase triggers destruction of the Rac1-GEF Cdc24 during polarized growth in Ustilago maydis. Mol. Biol. Cell 2011, 22, 3253–3262. [Google Scholar] [CrossRef] [PubMed]
- König, J.; Baumann, S.; Koepke, J.; Pohlmann, T.; Zarnack, K.; Feldbrügge, M. The fungal RNA-binding protein Rrm4 mediates long-distance transport of ubi1 and rho3 mRNAs. EMBO J. 2009, 28, 1855–1866. [Google Scholar] [CrossRef] [Green Version]
- Smith, D.G.; Garcia-Pedrajas, M.D.; Gold, S.E.; Perlin, M.H. Isolation and characterization from pathogenic fungi of genes encoding ammonium permeases and their roles in dimorphism. Mol. Microbiol. 2003, 50, 259–275. [Google Scholar] [CrossRef]
- Zhong, Y.; Yan, M.; Jiang, J.; Zhang, Z.; Huang, J.; Zhang, L.; Deng, Y.; Xiaofan Zhou, X.; He, F. Mycophenolic acid as a promising fungal dimorphism inhibitor to control sugar cane disease caused by Sporisorium scitamineum. J. Agric. Food Chem. 2019, 67, 112–119. [Google Scholar] [CrossRef]
- Zhang, B.; Cui, G.B.; Chang, C.Q.; Wang, Y.X.; Zhang, H.Y.; Chen, B.S.; Deng, Y.Z.; Jiang, Z.D. The autophagy gene ATG8 affects morphogenesis and oxidative stress tolerance in Sporisorium scitamineum. J. Integr. Agric. 2019, 18, 1024–1034. [Google Scholar] [CrossRef]
- Zhang, Y.; Wu, M.; Ge, Q.; Yang, M.; Xia, W.; Cui, H.; Yu, X.; Zhang, S.; Ye, Z. Cloning and disruption of the UeArginase in Ustilago esculenta: Evidence for a role of arginine in its dimorphic transition. BMC Microbiol. 2019, 19, 208. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pathway | Gene | Ustilago maydisa | Ustilago hordeia,b | Ustilago bromivorac | Sporisorium reilianuma,b | Sporisorium graminicolaa | Testicularia cyperia,b,d |
|---|---|---|---|---|---|---|---|
| PKA | RAL2 | UMAG_00884 | UHOR_01334 | UBRO_01334 | sr12177 | EX895_006321 | BCV70DRAFT_173605 |
| GPA3 | UMAG_04474 | UHOR_06981 | UBRO_06981 | sr15360 | EX895_002426 | CE102777_5455 | |
| BPP1 | UMAG_00703 | UHOR_01085 | UBRO_01086 | sr11991 | EX895_000934 | CE92962_8479 | |
| PKA | UMAG_06450 | UHOR_09003 | UBRO_09003 | sr16888 | EX895_004218 | CE31316_43035 | |
| UAC1 | UMAG_05232 | UHOR_03218 | UBRO_03218 | sr13141 | EX895_004629 | BCV70DRAFT_200755 | |
| UBC1 | UMAG_00525 | UHOR_00805 | UBRO_00805 | sr10199.2 | EX895_000753 | BCV70DRAFT_164254 | |
| ADR1 | UMAG_04456 | UHOR_06957 | UBRO_06957 | sr15343 | EX895_002409 | BCV70DRAFT_103006 | |
| SQL1 | UMAG_05501 | UHOR_07888 | UBRO_07888 | sr16182 | EX895_002861 | BCV70DRAFT_212144 | |
| HGL1 | UMAG_11450 | UHOR_00981 | UBRO_00981 | sr11921 | EX895_000866 | BCV70DRAFT_202631 | |
| CRK1 | UMAG_11410 | UHOR_04041 | UBRO_04041 | sr10962 | EX895_005166 | BCV70DRAFT_157158 | |
| MAPK | RAS2 | UMAG_01643 | UHOR_01498 | UBRO_02437 | sr12711 | EX895_004437 | CE98444_23931 |
| SQL2 | UMAG_10803 | UHOR_02247 | UBRO_02247 | sr12585 | EX895_004314 | BCV70DRAFT_231562 | |
| SQL2 | UMAG_11476 | UHOR_04466 | UBRO_04466 | sr13877 | EX895_001846 | BCV70DRAFT_223969 | |
| UBC2 | UMAG_05261 | UHOR_08070 | UBRO_08070 | sr16309 | EX895_003017 | BCV70DRAFT_197656 | |
| MAPKKK | UMAG_04258 | UHOR_06391 | UBRO_06391 | sr15150 | EX895_002006 | BCV70DRAFT_154027 | |
| MAPKK | UMAG_01514 | UHOR_02245 | UBRO_02245 | sr12583 | EX895_004312 | BCV70DRAFT_211523 | |
| MAPKK | UMAG_11453 | UHOR_01494 | UBRO_00992 | sr11928 | EX895_000873 | CE125301_8671 | |
| MAPKK | UMAG_00721 | UHOR_01105 | UBRO_01105 | sr12009 | EX895_000953 | BCV70DRAFT_71012 | |
| MAPK | UMAG_03305 | UHOR_05144 | UBRO_05144 | sr14305 | EX895_005556 | CE107062_12366 | |
| MAPK | UMAG_02331 | UHOR_05144 | UBRO_05144 | sr14305 | EX895_005556 | BCV70DRAFT_107063 | |
| CRK1 | UMAG_11410 | UHOR_04041 | UBRO_04041 | sr10962 | EX895_005166 | BCV70DRAFT_157158 | |
| RIM | RIM101 | UMAG_10426 | UHOR_06410 | UBRO_06410 | sr15166 | EX895_002022 | CE711_11906 |
| RIM9 | UMAG_00581 | UHOR_00899 | UBRO_00899 | sr11859 | EX895_000793 | CE73867_14878 | |
| RIM20 | UMAG_11510 | UHOR_05885 | UBRO_05885 | sr14775 | EX895_001317 | BCV70DRAFT_202631 | |
| RIM13 | UMAG_02075 | UHOR_03447 | UBRO_03447 | sr13297 | EX895_003420 | BCV70DRAFT_198765 | |
| RIM23 | UMAG_04392 | UHOR_06863 | UBRO_06863 | sr15280 | EX895_002350 | BCV70DRAFT_161602 | |
| RHO4 | UMAG_10663 | UHOR_01182 | UBRO_05736 | sr14670 | EX895_001201 | CE23374_4310 |
| Gene | Ustilago maydisa | Ustilago hordeia,b | Ustilago bromivorac | Sporisorium reilianuma,b | Sporisorium graminicolaa | Testicularia cyperia,b,d | |
|---|---|---|---|---|---|---|---|
| Acetylornithine aminotransferase | OAT | UMAG_05671 | UHOR_07330 | UBRO_07330 | sr15996 | EX895_002715 | BCV70DRAFT_201257 |
| Ornithine decarboxylas | ODC | UMAG_01048 | UHOR_01580 | UBRO_01580 | sr12348 | EX895_006099 | BCV70DRAFT_212462 |
| Polyamine oxidase | PAO | UMAG_05850 | UHOR_08347 | UBRO_08347 | sr16473 | EX895_008244 | BCV70DRAFT_173429 |
| Chimeric spermidine synthase/saccharopine reductase | SPE | UMAG_05818 | UHOR_08297 | UBRO_08297 | sr16440 | EX895_003211 | BCV70DRAFT_199161 |
| S-adenosylmethionine decarboxylase | SAMDC | UMAG_10792 | UHOR_01520 | UBRO_01520 | sr12300 | EX895_006201 | BCV70DRAFT_205658 |
| Putative spermidine acetyltrasferase | SSAT | UMAG_00127 | UHOR_00200 | UBRO_00200 | sr11469 | EX895_000152 | BCV70DRAFT_169192 |
| Gene | Ustilago maydisa | Ustilago hordeia,b | Ustilago bromivorac | Sporisorium reilianuma,b | Sporisorium graminícolaa | Testicularia cyperia,b,d |
|---|---|---|---|---|---|---|
| GCN5—Histone acetyltransferase | UMAG_05168 | UHOR_03120 | UBRO_03120 | sr13072 | EX895_004562 | BCV70DRAFT_198404 |
| Putative histone acetylase | UMAG_10190 | UHOR_03120 | UBRO_03120 | sr13072 | EX895_004562 | BCV70DRAFT_198404 |
| CLR3—Histone deacetylase | UMAG_02102 | UHOR_03487 | UBRO_03487 | sr13325 | EX895_004958 | BCV70DRAFT_166948 |
| HOS2—Histone deacetylase | UMAG_11828 | UHOR_01015 | UBRO_01015 | sr11943 | EX895_000889 | BCV70DRAFT_70918 |
| TEA4—SH3 domain protein | UMAG_01012 | UHOR_01534 | UBRO_01534 | sr12319 | EX895_006176 | BCV70DRAFT_159133 |
| TEA1—Kelch domain protein | UMAG_15019 | UHOR_01124 | UBRO_01124 | sr12022 | EX895_000966 | BCV70DRAFT_69247 |
| REP1—Repellent | UMAG_03924 | UHOR_05948 | UBRO_05948 | sr14829 | EX895_001361 | BCV70DRAFT_80316 |
| REP4—Repellent | UMAG_04517 | UHOR_06499 | UBRO_06499 | sr15402 | EX895_002086 | BCV70DRAFT_202324 |
| HUM2—Hydrophobin 2 | UMAG_11562 | UHOR_07158 | UBRO_07158 | sr15890 | EX895_002555 | BCV70DRAFT_207191 |
| HUM3—Hydrophobin 3 | UMAG_04433 | UHOR_06926 | UBRO_06926 | sr15320 | EX895_002383 | BCV70DRAFT_189664 |
| RRM4—RNA-binding protein | UMAG_03494 | UHOR_05377 | UBRO_05377 | sr14484 | EX895_006466 | BCV70DRAFT_213009 |
| Actin-binding protein | UMAG_05340 | UHOR_08160 | UBRO_08160 | sr16324 | EX895_003081 | BCV70DRAFT_155405 |
| Actin | UMAG_11232 | UHOR_08813 | UBRO_08813 | sr11345 | EX895_003701 | BCV70DRAFT_201455 |
| Actin-interactin gprotein; actin patch component | UMAG_05949 | UHOR_08467 | UBRO_08467 | sr16579 | EX895_005119 | BCV70DRAFT_213703 |
| Actin filament organization | UMAG_04613 | UHOR_06638 | UBRO_06638 | sr15500 | EX895_002183 | BCV70DRAFT_202237 |
| LIS1—Nuclear migration | UMAG_03164 | UHOR_04938 | UBRO_04938 | sr14206 | EX895_005820 | BCV70DRAFT_200264 |
| Actin regulating Ser/Thr kinase | UMAG_03081 | UHOR_04824 | UBRO_04824 | sr14140 | EX895_002483 | - |
| Formin; actin nucleation | UMAG_12254 | UHOR_06202 | UBRO_06202 | sr15020 | EX895_001591 | BCV70DRAFT_205747 |
| Formin; actin nucleation. Functionally redundant with Bni1 | UMAG_01141 | UHOR_01723 | UBRO_01723 | sr12440 | EX895_006065 | BCV70DRAFT_12160 |
| F-actin capping protein alpha subunit | UMAG_00423 | UHOR_00664 | UBRO_00664 | sr11788 | EX895_000652 | BCV70DRAFT_196835 |
| F-actin capping protein beta subunit | UMAG_11177 | UHOR_08001 | UBRO_08001 | sr16266 | EX895_002944 | BCV70DRAFT_201290 |
| Chaperonin role for actin and tubulin | UMAG_01279 | UHOR_01918 | UBRO_01918 | sr10401 | EX895_001741 | BCV70DRAFT_197510 |
| Chaperonin role for actin and tubulin | UMAG_06235 | UHOR_08845 | UBRO_08845 | sr16767 | EX895_003720 | BCV70DRAFT_199599 |
| Chaperonin role for actin and tubulin | UMAG_06067 | UHOR_08612 | UBRO_08612 | sr16694 | EX895_003550 | BCV70DRAFT_202419 |
| Chaperonin role for actin and tubulin | UMAG_02571 | UHOR_04113 | UBRO_04113 | sr17132 | EX895_005225 | BCV70DRAFT_198632 |
| Chaperonin role for actin and tubulin | UMAG_03959 | UHOR_06000 | UBRO_06000 | sr14864 | EX895_001395 | BCV70DRAFT_69984 |
| Chaperonin role for actin and tubulin | UMAG_02350 | UHOR_03838 | UBRO_03838 | sr13553 | EX895_004884 | BCV70DRAFT_198957 |
| Chaperonin role for actin and tubulin | UMAG_00565 | UHOR_00911 | UBRO_00911 | sr11843 | EX895_000809 | BCV70DRAFT_73687 |
| Chaperonin role for actin and tubulin | UMAG_04401 | UHOR_06879 | UBRO_06879 | sr15289 | EX895_002355 | BCV70DRAFT_200184 |
| Actin binding and severing protein | UMAG_04314 | UHOR_06753 | UBRO_06753 | sr15202 | EX895_002269 | BCV70DRAFT_211263 |
| Cortical actin cytoskeleton component | UMAG_04417 | UHOR_06902 | UBRO_06902 | sr15303 | EX895_002369 | BCV70DRAFT_200171 |
| Kinesin—Related to UMAG_ Kin14 | UMAG_11986 | UHOR_06484 | UBRO_06484 | sr15392 | EX895_002076 | BCV70DRAFT_202332 |
| Kinesin—Functionally redundant with Cin8 | UMAG_10678 | UHOR_07574 | UBRO_07574 | sr15603 | EX895_000239 | BCV70DRAFT_203263 |
| Kinesin—Related motor protein (UMAG_ Kin7a and Kin7b) | UMAG_00896 | UHOR_01350 | UBRO_01350 | sr12193 | EX895_006269 | BCV70DRAFT_199338 |
| Kinesin—related motor protein / Related to UMAG_ Kin8 | UMAG_01560 | UHOR_02319 | UBRO_02319 | sr12632 | EX895_004356 | BCV70DRAFT_199992 |
| Interacts with Myo2—related to motor domain of kinesins | UMAG_04218 | UHOR_06328 | UBRO_06328 | sr15103 | EX895_001669 | - |
| Type V myosin | UMAG_04555 | UHOR_06551 | UBRO_06551 | sr15438 | EX895_002121 | BCV70DRAFT_40829 |
| MEP1—Low affinity ammonium transporter | UMAG_04523 | UHOR_06506 | UBRO_06506 | sr15408 | EX895_002092 | BCV70DRAFT_208136 |
| MEP2—High affinity ammonium transporter | UMAG_05889 | UHOR_08388 | UBRO_08388 | sr16507 | EX895_003276 | BCV70DRAFT_199125 |
| Putative arginase—Homology with G6606 of U. esculenta | UMAG_04939 | UHOR_07065 | UBRO_07065 | sr15821 | EX895_005725 | BCV70DRAFT_178133 |
| Homology with SSAGC1 (SPSC_00276) of S. scitamineum) | UMAG_11677 | UHOR_02215 | UBRO_02215 | sr12574 | EX895_004280 | BCV70DRAFT_160776 |
| CDC24—Cell division control protein | UMAG_02422 | UHOR_03963 | UBRO_03963 | sr10836 | EX895_004832 | BCV70DRAFT_192816 |
| CLA4—Serine/threonine-protein kinase | UMAG_10145 | UHOR_03893 | UBRO_03893 | sr10814.2 | EX895_004846 | BCV70DRAFT_158032 |
| RHO1—Ras-like GTP-binding protein | UMAG_01032 | UHOR_01559 | UBRO_01559 | sr12336 | EX895_006088 | BCV70DRAFT_166498 |
| RHO1—Ras-like GTP-binding protein | UMAG_05734 | UHOR_12739 | UBRO_07439 | sr16067 | EX895_002745 | CE53947_104590 |
| FUZ1—MYND domain protein | UMAG_02587 | UHOR_04136 | UBRO_04136 | sr13628 | EX895_005246 | BCV70DRAFT_63386 |
| Cell wall | Gene | Ustilago maydisa | Ustilago hordeia,b | Ustilago bromivorac | Sporisorium reilianuma,b | Sporisorium graminicolaa | Testicularia cyperia,b,d |
|---|---|---|---|---|---|---|---|
| Synthesis | 1,3-β-glucan synthase | UMAG_01639 | UHOR_02430 | UBRO_02430 | sr12707 | EX895_004433 | BCV70DRAFT_98536 |
| CHS1 | UMAG_10718 | UHOR_07282 | UBRO_07282 | sr16010 | EX895_002642 | BCV70DRAFT_198128 | |
| CHS2 | UMAG_04290 | UHOR_06435 | UBRO_06435 | sr15181 | EX895_002037 | - | |
| CHS3 | UMAG_10120 | UHOR_00740 | UBRO_00740 | sr10158 | EX895_000703 | BCV70DRAFT_196781 | |
| CHS4 | UMAG_10117 | UHOR_00723 | UBRO_00723 | sr10139 | EX895_000691 | CE2605_17934 | |
| CHS5 | UMAG_10277 | UHOR_04112 | UBRO_04112 | sr11048.2 | EX895_005224 | BCV70DRAFT_156817 | |
| CHS6 | UMAG_10367 | UHOR_04780 | UBRO_04780 | sr14109 | EX895_002515 | BCV70DRAFT_22903 | |
| CHS7 | UMAG_05480 | UHOR_07854 | UBRO_07854 | sr16158 | EX895_002841 | BCV70DRAFT_162870 | |
| CHS8 | UMAG_03204 | UHOR_04988 | UBRO_04988 | sr11106 | EX895_005864 | BCV70DRAFT_105640 | |
| CDA1d | UMAG_11922 | UHOR_04296 | UBRO_04296 | sr13741 | EX895_005345 | BCV70DRAFT_210413 | |
| CDA2d | UMAG_00126 | UHOR_00199 | UBRO_00199 | sr11468 | EX895_000151 | BCV70DRAFT_197174 | |
| CDA3d | UMAG_00638 | - | - | sr11918 | EX895_000863 | - | |
| CDA4d | UMAG_01143 | UHOR_01725 | UBRO_01725 | sr12442 | EX895_006063 | - | |
| CDA5d | UMAG_01788 | UHOR_02660 | UBRO_02660 | sr12866 | EX895_003869 | BCV70DRAFT_211320 | |
| CDA6d | UMAG_02019 | UHOR_03000 | UBRO_03000 | sr12981 | EX895_004113 | BCV70DRAFT_34433 | |
| CDA7d | UMAG_02381 | UHOR_03898 | UBRO_03898 | sr13587 | EX895_004867 | BCV70DRAFT_163992 | |
| CDA8d | UMAG_05792 | UHOR_04296 | UBRO_04296 | sr16123 | EX895_002800 | BCV70DRAFT_210413 | |
| Degradation | CTS1 | UMAG_10419 | UHOR_06394 | UBRO_06394 | sr15153 | EX895_002009 | BCV70DRAFT_196730 |
| CTS2 | UMAG_02758 | UHOR_04393 | UBRO_04393 | sr13813 | EX895_005415 | CE60997_75476 | |
| CTS3 | UMAG_06190 | UHOR_08772 | UBRO_06394 | sr11305 | EX895_003673 | BCV70DRAFT_178298 | |
| CTS4 | UMAG_00695 | UHOR_01069 | UBRO_01069 | sr11983 | EX895_000921 | BCV70DRAFT_213542 | |
| CTS5 | UMAG_05290 | UHOR_08119 | UBRO_08119 | sr10043 | EX895_003044 | BCV70DRAFT_197812 | |
| Glucanase 1 | UMAG_04368 | - | UBRO_06828 | sr15253 | - | - | |
| Glucanase 2 | UMAG_05036 | UHOR_07211 | UBRO_07211 | sr15917 | - | CE121843_61034 | |
| Glucanase 3 | UMAG_00876 | UHOR_00370 | UBRO_01319 | sr12165 | EX895_006333 | CE84129_149800 | |
| Glucanase 4 | UMAG_01898 | - | UBRO_02820 | sr10624 | EX895_003984 | - | |
| Glucanase 5 | UMAG_05223 | UHOR_03204 | UBRO_03204 | sr13131 | EX895_004619 | BCV70DRAFT_227049 | |
| Glucanase 6 | UMAG_02134 | - | UBRO_03533 | sr13360 | EX895_004991 | - | |
| Glucanase 7 | UMAG_04357 | UHOR_06809 | UBRO_06809 | sr15243 | EX895_002306 | BCV70DRAFT_189826 | |
| Glucanase 8 | UMAG_05550 | - | UBRO_06823 | sr15250 | EX895_002316 | BCV70DRAFT_200231 | |
| Glucanase 9 | UMAG_10211 | - | UBRO_08206 | sr16359 | EX895_003114 | - |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martínez-Soto, D.; Ortiz-Castellanos, L.; Robledo-Briones, M.; León-Ramírez, C.G. Molecular Mechanisms Involved in the Multicellular Growth of Ustilaginomycetes. Microorganisms 2020, 8, 1072. https://doi.org/10.3390/microorganisms8071072
Martínez-Soto D, Ortiz-Castellanos L, Robledo-Briones M, León-Ramírez CG. Molecular Mechanisms Involved in the Multicellular Growth of Ustilaginomycetes. Microorganisms. 2020; 8(7):1072. https://doi.org/10.3390/microorganisms8071072
Chicago/Turabian StyleMartínez-Soto, Domingo, Lucila Ortiz-Castellanos, Mariana Robledo-Briones, and Claudia Geraldine León-Ramírez. 2020. "Molecular Mechanisms Involved in the Multicellular Growth of Ustilaginomycetes" Microorganisms 8, no. 7: 1072. https://doi.org/10.3390/microorganisms8071072