Arbuscular Mycorrhizal Fungal Communities of Native Plant Species under High Petroleum Hydrocarbon Contamination Highlights Rhizophagus as a Key Tolerant Genus



Abstract
:
1. Introduction
2. Materials and Methods
2.1. Site of Study and Sampling
2.2. DNA Extraction
2.3. PCR, Cloning and Sequencing
2.4. Soil Contaminant Analyses
2.5. Bioinformatics and Statistical Analyses
3. Results
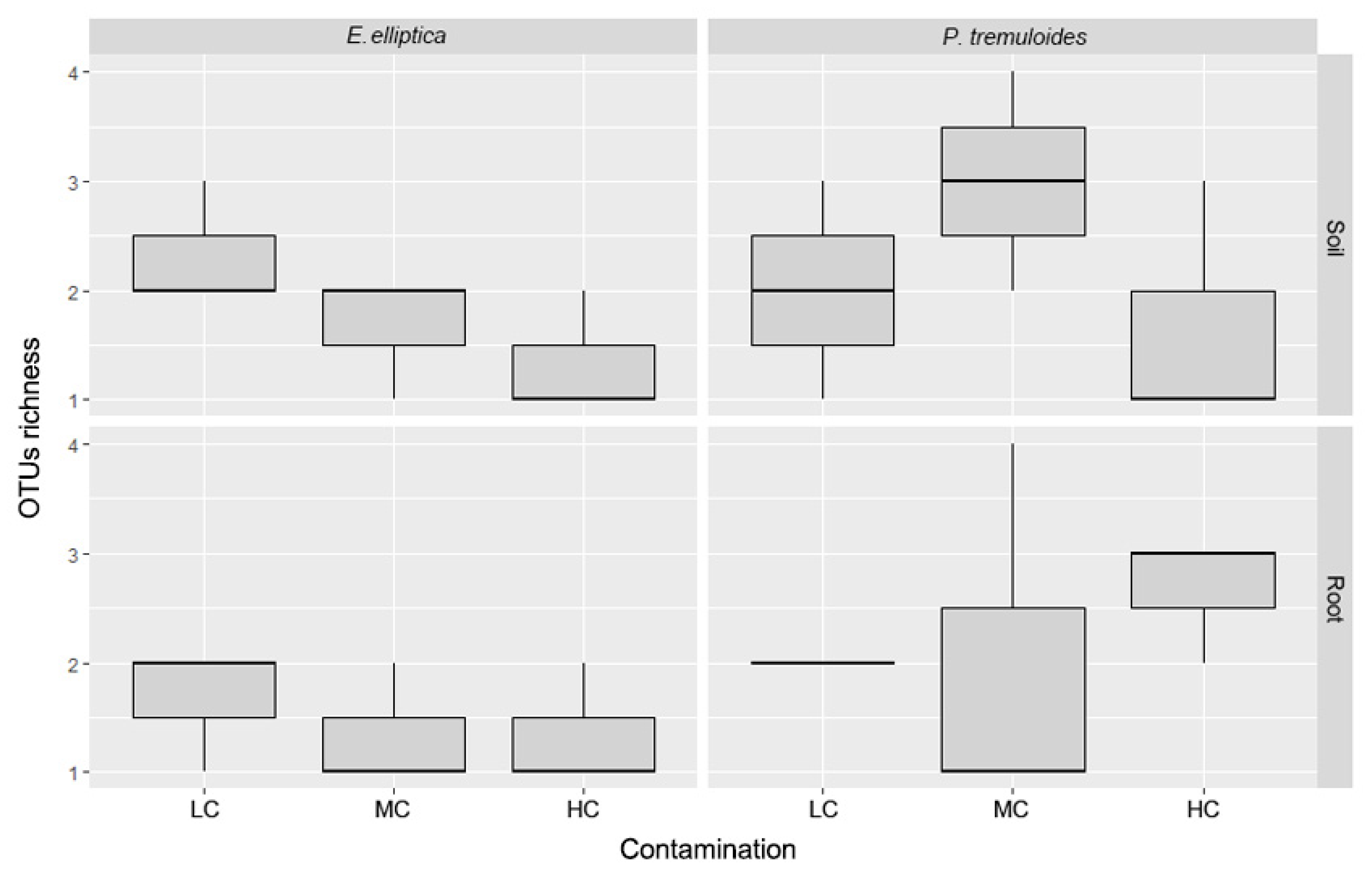
3.1. AMF Molecular Identity and Diversity
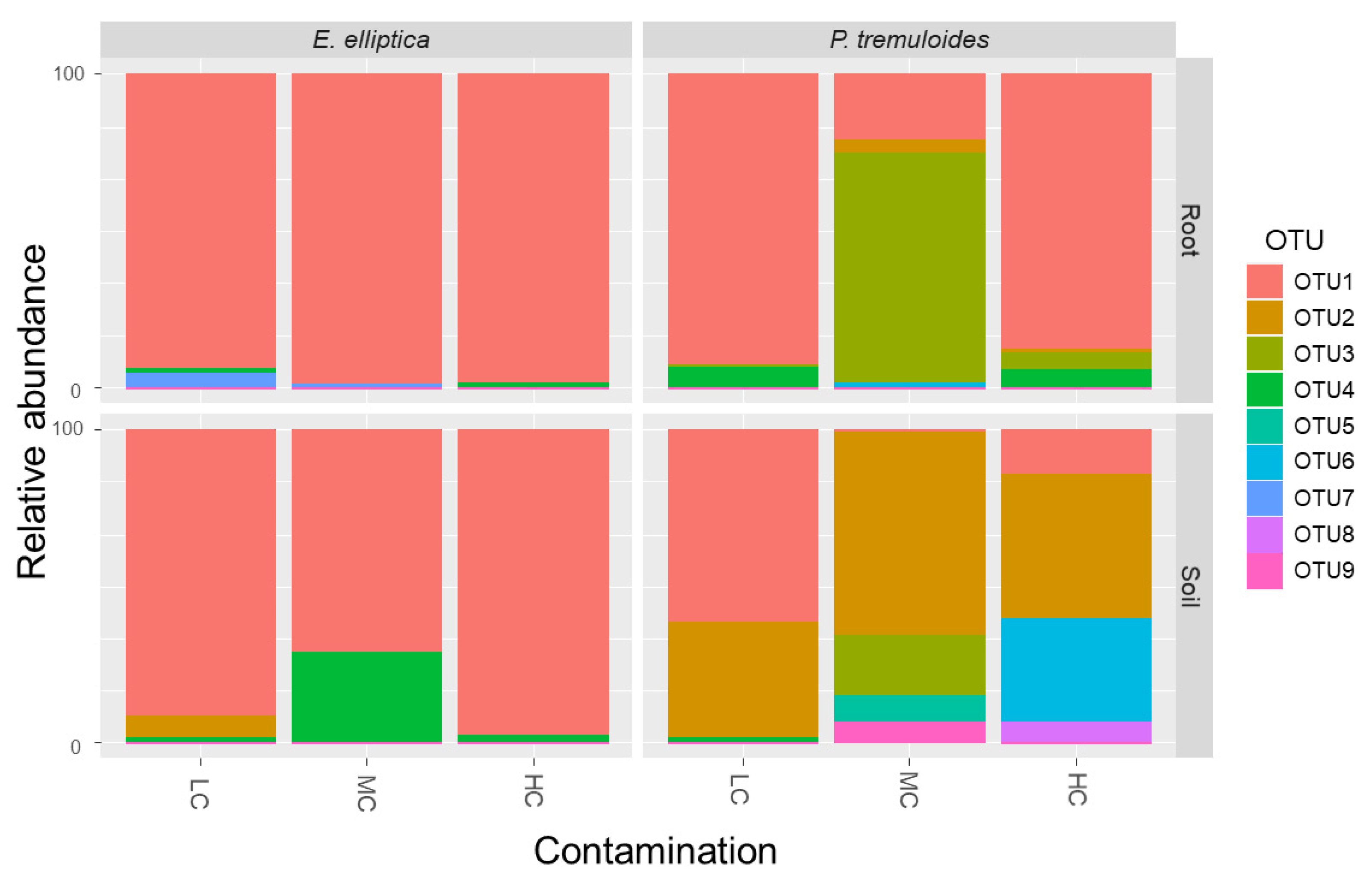
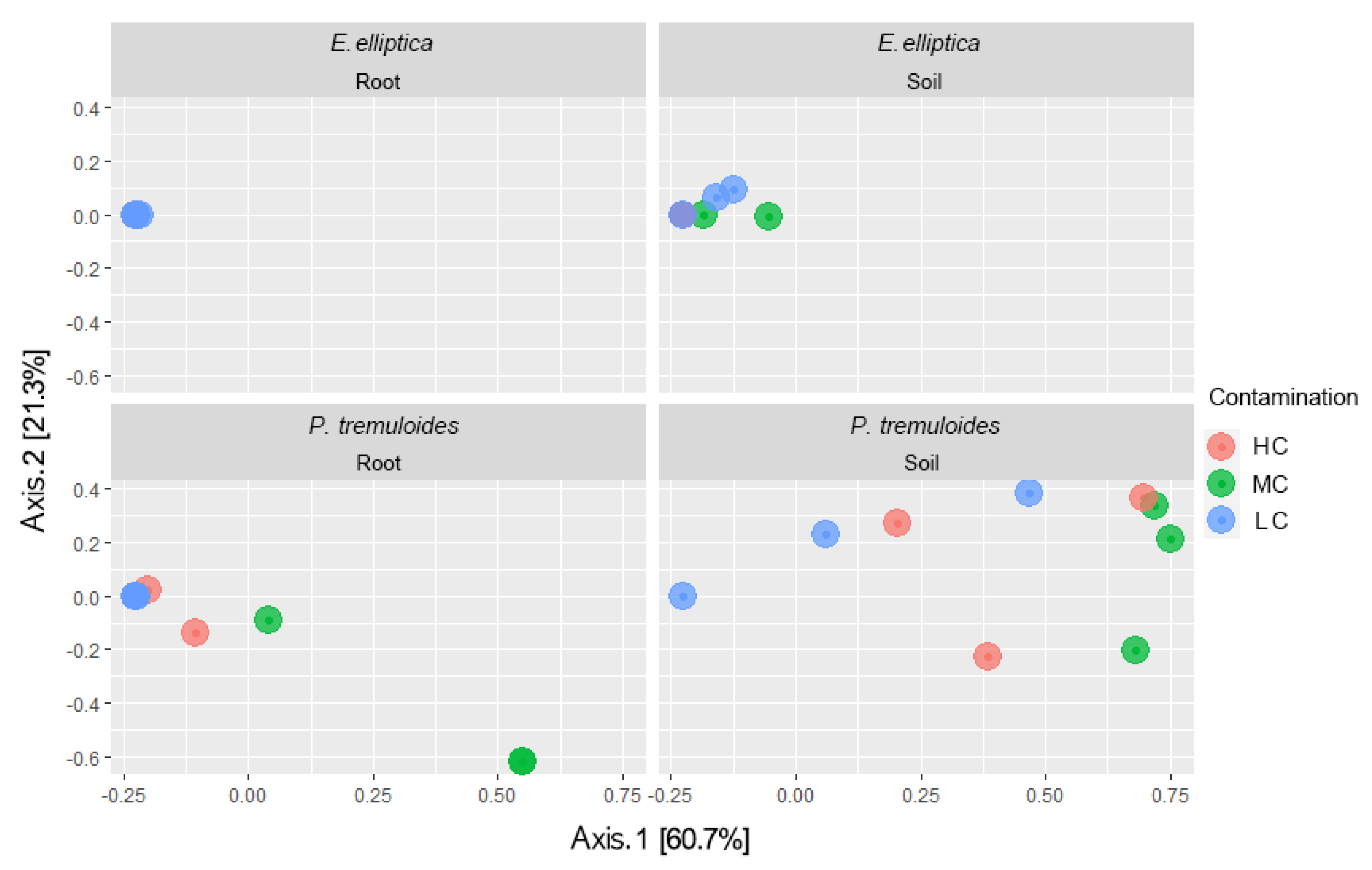
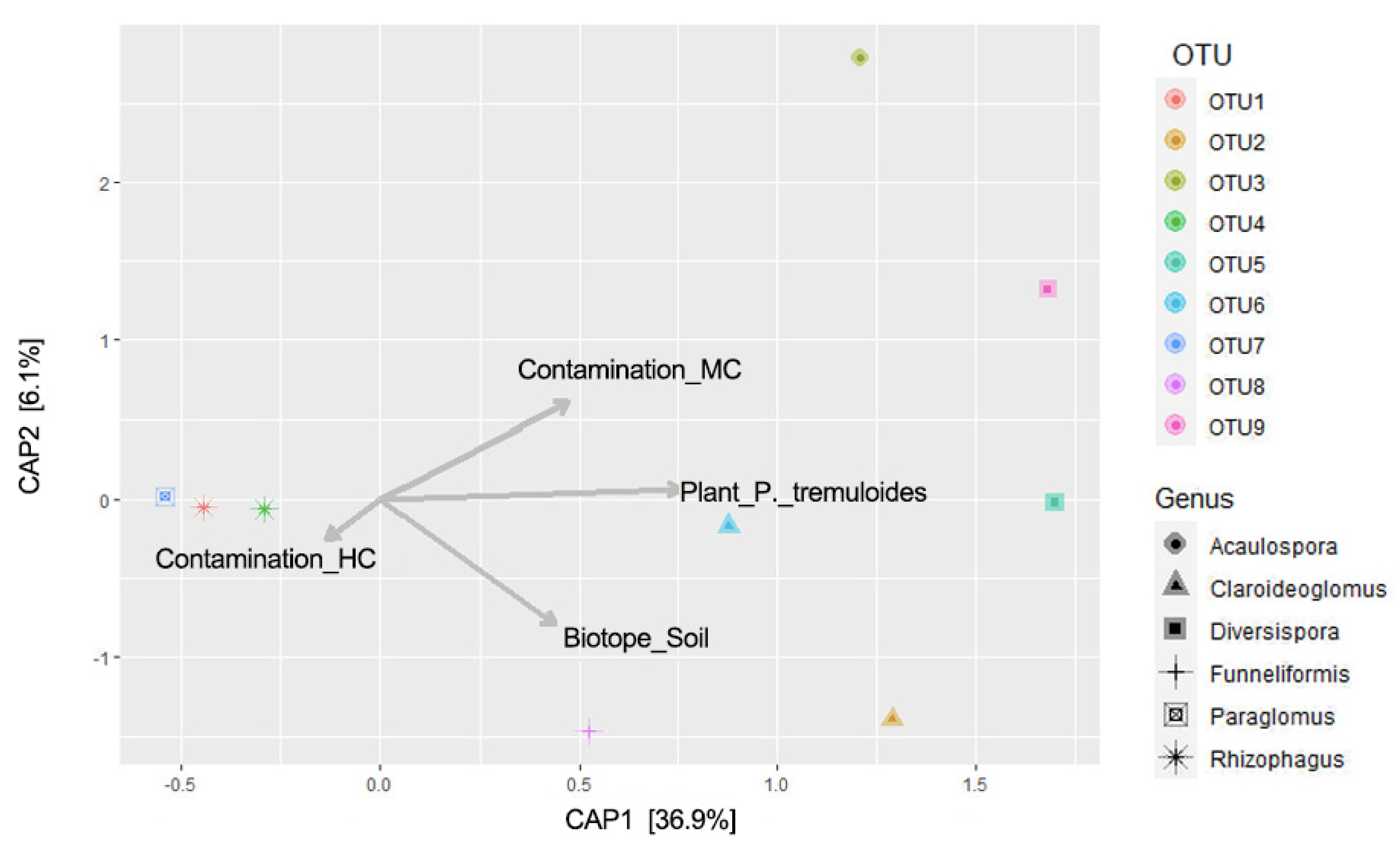
3.2. AMF Community Structure
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Alkan, N.; Gadkar, V.; Yarden, O.; Kapulnik, Y. Analysis of quantitative interactions between two species of arbuscular mycorrhizal fungi, Glomus mosseae and G. intraradices, by real-time PCR. Appl. Environ. Microbiol. 2006, 72, 4192–4199. [Google Scholar] [CrossRef] [Green Version]
- Brundrett, M. Mycorrhizas in Natural Ecosystems. Adv. Ecol. Res. 1991, 21, 171–313. [Google Scholar]
- Smith, S.; Read, D. Mycorrhizal Symbiosis, 3rd ed.; Academic Press: London, UK, 2008. [Google Scholar]
- Roy-Bolduc, A.; Hijri, M. The Use of Mycorrhizae to Enhance Phosphorus Uptake: A Way Out the Phosphorus Crisis. J. Biofertil. Biopestic. 2011, 2, 104. [Google Scholar] [CrossRef] [Green Version]
- Hijri, M. Analysis of a large dataset of mycorrhiza inoculation field trials on potato shows highly significant increases in yield. Mycorrhiza 2016, 26, 209–214. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.E.; Barker, S.J. Plant phosphate transporter genes help harness the nutritional benefits of arbuscular, mycorrhizal symbiosis. Trends Plant Sci. 2002, 7, 189–190. [Google Scholar] [CrossRef]
- Davies, F.T.; Puryear, J.D.; Newton, R.J.; Egilla, J.N.; Grossi, J.A.S. Mycorrhizal fungi increase chromium uptake by sunflower plants: Influence on tissue mineral concentration, growth, and gas exchange. J. Plant Nutr. 2002, 25, 2389–2407. [Google Scholar] [CrossRef] [Green Version]
- Hassan, S.E.; Hijri, M.; St-Arnaud, M. Effect of arbuscular mycorrhizal fungi on trace metal uptake by sunflower plants grown on cadmium contaminated soil. New Biotechnol. 2013, 30, 780–787. [Google Scholar] [CrossRef]
- Ismail, Y.; Hijri, M. Arbuscular mycorrhisation with Glomus irregulare induces expression of potato PR homologues genes in response to infection by Fusarium sambucinum. Funct. Plant Biol. 2012, 39, 236. [Google Scholar] [CrossRef]
- Ismail, Y.; McCormick, S.; Hijri, M. The arbuscular mycorrhizal fungus, Glomus irregulare, controls the mycotoxin production of Fusarium sambucinum in the pathogenesis of potato. FEMS Microbiol. Lett. 2013, 348, 46–51. [Google Scholar] [CrossRef] [Green Version]
- Miransari, M. Hyperaccumulators, arbuscular mycorrhizal fungi and stress of heavy metals. Biotechnol. Adv. 2011, 29, 645–653. [Google Scholar] [CrossRef]
- Porcel, R.; Aroca, R.; Ruiz-Lozano, J.M. Salinity stress alleviation using arbuscular mycorrhizal fungi. A review. Agron. Sustain. Dev. 2012, 32, 181–200. [Google Scholar] [CrossRef] [Green Version]
- St-Arnaud, M.; Vujanovic, V. Effect of the arbuscular mycorrhizal symbiosis on plant diseases and pests. In Mycorrhizae in Crop Production; Hamel, C., Plenchette, C., Eds.; Haworth Food & Agricultural Products Press: Binghamton, NY, USA, 2007. [Google Scholar]
- Yang, Y.R.; Han, X.Z.; Liang, Y.; Ghosh, A.; Chen, J.; Tang, M. The Combined Effects of Arbuscular Mycorrhizal Fungi (AMF) and Lead (Pb) Stress on Pb Accumulation, Plant Growth Parameters, Photosynthesis, and Antioxidant Enzymes in Robinia pseudoacacia L. PLoS ONE 2015, 10, e0145726. [Google Scholar] [CrossRef] [PubMed]
- Debiane, D.; Garçon, G.; Verdin, A.; Fontaine, J.; Durand, R.; Shirali, P.; Grandmougin-Ferjani, A.; Lounès-Hadj Sahraoui, A. Mycorrhization alleviates benzo[a]pyrene-induced oxidative stress in an in vitro chicory root model. Phytochemistry 2009, 70, 1421–1427. [Google Scholar] [CrossRef]
- Lenoir, I.; Fontaine, J.; Tisserant, B.; Laruelle, F.; Lounes-Hadj Sahraoui, A. Beneficial contribution of the arbuscular mycorrhizal fungus, Rhizophagus irregularis, in the protection of Medicago truncatula roots against benzo[a]pyrene toxicity. Mycorrhiza 2017, 27, 465–476. [Google Scholar] [CrossRef]
- Rivera-Becerril, F.; Calantzis, C.; Turnau, K.; Caussanel, J.-P.; Belimov, A.A.; Gianinazzi, S.; Strasser, R.J.; Gianinazzi-Pearson, V. Cadmium accumulation and buffering of cadmium-induced stress by arbuscular mycorrhiza in three Pisum sativum L. genotypes. J. Exp. Bot. 2002, 53, 1177–1185. [Google Scholar] [CrossRef] [Green Version]
- Iffis, B.; St-Arnaud, M.; Hijri, M. Petroleum hydrocarbon contamination, plant identity and arbuscular mycorrhizal fungal (AMF) community determine assemblages of the AMF spore-associated microbes. Environ. Microbiol. 2016, 18, 2689–2704. [Google Scholar] [CrossRef]
- Iffis, B.; St-Arnaud, M.; Hijri, M. Petroleum Contamination and Plant Identity Influence Soil and Root Microbial Communities While AMF Spores Retrieved from the Same Plants Possess Markedly Different Communities. Front. Plant Sci. 2017, 8, 1381. [Google Scholar] [CrossRef]
- Eom, A.H.; Hartnett, D.C.; Wilson, G.W.T. Host plant species effects on arbuscular mycorrhizal fungal communities in tallgrass prairie. Oecologia 2000, 122, 435–444. [Google Scholar] [CrossRef]
- Guo, X.H.; Gong, J. Differential effects of abiotic factors and host plant traits on diversity and community composition of root-colonizing arbuscular mycorrhizal fungi in a salt-stressed ecosystem. Mycorrhiza 2014, 24, 79–94. [Google Scholar] [CrossRef]
- Torrecillas, E.; Alguacil, M.M.; Roldan, A. Host Preferences of Arbuscular Mycorrhizal Fungi Colonizing Annual Herbaceous Plant Species in Semiarid Mediterranean Prairies. Appl. Environ. Microbiol. 2012, 78, 6180–6186. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.S.; Zang, Y.Y.; Yuan, Y.G.; Tang, J.J.; Chen, X. Selectivity by host plants affects the distribution of arbuscular mycorrhizal fungi: Evidence from ITS rDNA sequence metadata. BMC Evol. Biol. 2012, 12, 50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hassan, S.E.D.; Bell, T.H.; Stefani, F.O.P.; Denis, D.; Hijri, M.; St-Arnaud, M. Contrasting the Community Structure of Arbuscular Mycorrhizal Fungi from Hydrocarbon-Contaminated and Uncontaminated Soils following Willow (Salix spp. L.) Planting. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [Green Version]
- Hassan, S.E.D.; Boon, E.; St-Arnaud, M.; Hijri, M. Molecular biodiversity of arbuscular mycorrhizal fungi in trace metal-polluted soils. Mol. Ecol. 2011, 20, 3469–3483. [Google Scholar] [CrossRef] [PubMed]
- de la Providencia, I.E.; Stefani, F.O.P.; Labridy, M.; St-Arnaud, M.; Hijri, M. Arbuscular mycorrhizal fungal diversity associated with Eleocharis obtusa and Panicum capillare growing in an extreme petroleum hydrocarbon-polluted sedimentation basin. FEMS Microbiol. Lett. 2015, 362, fnv081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, E.C.H.; Morin, E.; Beaudet, D.; Noel, J.; Yildirir, G.; Ndikumana, S.; Charron, P.; St-Onge, C.; Giorgi, J.; Kruger, M.; et al. High intraspecific genome diversity in the model arbuscular mycorrhizal symbiont Rhizophagus irregularis. New Phytol. 2018, 220, 1161–1171. [Google Scholar] [CrossRef] [Green Version]
- Morin, E.; Miyauchi, S.; San Clemente, H.; Chen, E.C.H.; Pelin, A.; de la Providencia, I.; Ndikumana, S.; Beaudet, D.; Hainaut, M.; Drula, E.; et al. Comparative genomics of Rhizophagus irregularis, R. cerebriforme, R. diaphanus and Gigaspora rosea highlights specific genetic features in Glomeromycotina. New Phytol. 2019, 222, 1584–1598. [Google Scholar] [CrossRef]
- Bourdel, G.; Roy-Bolduc, A.; St-Arnaud, M.; Hijri, M. Concentration of Petroleum-Hydrocarbon Contamination Shapes Fungal Endophytic Community Structure in Plant Roots. Front. Microbiol. 2016, 7, 685. [Google Scholar] [CrossRef]
- Dagher, D.J.; de la Providencia, I.E.; Pitre, F.E.; St-Arnaud, M.; Hijri, M. Arbuscular Mycorrhizal Fungal Assemblages Significantly Shifted upon Bacterial Inoculation in Non-Contaminated and Petroleum-Contaminated Environments. Microorganisms 2020, 8, 602. [Google Scholar] [CrossRef] [Green Version]
- Desjardins, D.; Nissim, W.G.; Pitre, F.E.; Naud, A.; Labrecque, M. Distribution patterns of spontaneous vegetation and pollution at a former decantation basin in southern Quebec, Canada. Ecol. Eng. 2014, 64, 385–390. [Google Scholar] [CrossRef]
- Lee, J.; Lee, S.; Young, J.P. Improved PCR primers for the detection and identification of arbuscular mycorrhizal fungi. FEMS Microbiol. Ecol. 2008, 65, 339–349. [Google Scholar] [CrossRef] [Green Version]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-Source, Platform-Independent, Community-Supported Software for Describing and Comparing Microbial Communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Opik, M.; Vanatoa, A.; Vanatoa, E.; Moora, M.; Davison, J.; Kalwij, J.M.; Reier, U.; Zobel, M. The online database MaarjAM reveals global and ecosystemic distribution patterns in arbuscular mycorrhizal fungi (Glomeromycota). New Phytol. 2010, 188, 223–241. [Google Scholar] [CrossRef]
- Kruger, M.; Kruger, C.; Walker, C.; Stockinger, H.; Schussler, A. Phylogenetic reference data for systematics and phylotaxonomy of arbuscular mycorrhizal fungi from phylum to species level. New Phytol. 2012, 193, 970–984. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [Green Version]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [Green Version]
- Del Val, C.; Barea, J.M.; Azcon-Aguilar, C. Diversity of arbuscular mycorrhizal fungus populations in heavy-metal-contaminated soils. Appl. Environ. Microbiol. 1999, 65, 718–723. [Google Scholar] [CrossRef] [Green Version]
- Pawlowska, T.E.; Chaney, R.L.; Chin, M.; Charvat, I. Effects of metal phytoextraction practices on the indigenous community of arbuscular mycorrhizal fungi at a metal-contaminated landfill. Appl. Environ. Microbiol. 2000, 66, 2526–2530. [Google Scholar] [CrossRef] [Green Version]
- Geras’kin, S.; Evseeva, T.; Oudalova, A. Effects of long-term chronic exposure to radionuclides in plant populations. J. Environ. Radioact. 2013, 121, 22–32. [Google Scholar] [CrossRef]
- Heckenroth, A.; Rabier, J.; Dutoit, T.; Torre, F.; Prudent, P.; Laffont-Schwob, I. Selection of native plants with phytoremediation potential for highly contaminated Mediterranean soil restoration: Tools for a non-destructive and integrative approach. J. Environ. Manag. 2016, 183, 850–863. [Google Scholar] [CrossRef]
- Woch, M.W.; Stefanowicz, A.M.; Stanek, M. Waste heaps left by historical Zn-Pb ore mining are hotspots of species diversity of beech forest understory vegetation. Sci. Total Environ. 2017, 599–600, 32–41. [Google Scholar] [CrossRef]
- Sanchez-Castro, I.; Gianinazzi-Pearson, V.; Cleyet-Marel, J.C.; Baudoin, E.; van Tuinen, D. Glomeromycota communities survive extreme levels of metal toxicity in an orphan mining site. Sci. Total Environ. 2017, 598, 121–128. [Google Scholar] [CrossRef]
- Lee, S.J.; Morse, D.; Hijri, M. Holobiont chronobiology: Mycorrhiza may be a key to linking aboveground and underground rhythms. Mycorrhiza 2019, 29, 403–412. [Google Scholar] [CrossRef]
- Lioussanne, L.; Perrault, F.; Jolicoeur, M.; St-Arnaud, M. The bacterial community of tomato rhizosphere is modified by inoculation with arbuscular mycorrhizal fungi but unaffected by soil enrichment with mycorrhizal root exudates or inoculation with Phytophthora nicotianae. Soil Biol. Biochem. 2010, 42, 473–483. [Google Scholar] [CrossRef]
- Nazir, R.; Warmink, J.; Boersma, H.; van Elsas, J. Mechanisms that promote bacterial fitness in fungal-affected soil microhabitats. FEMS Microbiol. Ecol. 2010, 71, 169–185. [Google Scholar] [CrossRef] [Green Version]
- Taktek, S.; St-Arnaud, M.; Piche, Y.; Fortin, J.A.; Antoun, H. Igneous phosphate rock solubilization by biofilm-forming mycorrhizobacteria and hyphobacteria associated with Rhizoglomus irregulare DAOM 197198. Mycorrhiza 2017, 27, 13–22. [Google Scholar] [CrossRef] [Green Version]
- Correia, M.; Heleno, R.; da Silva, L.P.; Costa, J.M.; Rodriguez-Echeverria, S. First evidence for the joint dispersal of mycorrhizal fungi and plant diaspores by birds. New Phytol. 2019, 222, 1054–1060. [Google Scholar] [CrossRef]
- Egan, C.; Li, D.; Klironomos, J. Detection of arbuscular mycorrhizal fungal spores in the air across different biomes and ecoregions. Fungal Ecol. 2014, 12, 26–31. [Google Scholar] [CrossRef]
- Su, Y.Y.; Sun, X.; Guo, L.D. Seasonality and Host Preference of Arbuscular Mycorrhizal Fungi of Five Plant Species in the Inner Mongolia Steppe, China. Braz. J. Microbiol. 2011, 42, 57–65. [Google Scholar] [CrossRef] [Green Version]
- Giovannetti, M. Seasonal variations of vesicular-arbuscular mycorrhizas and endogonaceous spores in a maritime sand dune. Trans. Br. Mycol. Soc. 1985, 84, 679–684. [Google Scholar] [CrossRef]
- Velazquez, M.S.; Cabello, M.N.; Barrera, M. Composition and structure of arbuscular-mycorrhizal communities in El Palmar National Park, Argentina. Mycologia 2013, 105, 509–520. [Google Scholar] [CrossRef]
- Dagher, J.D.; Pitre, E.P.; Hijri, M. Ectomycorrhizal fungal inoculation of Sphaerosporella brunnea significantly increased stem biomass of Salix miyabeana and decreased lead, tin, and zinc, soil concentrations during the phytoremediation of a trace elements-contaminated industrial landfill. J. Fungi 2020. submitted. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 1 Contamination | Plant Species | Biotope | 2 Shannon | 2 Simpson | 2 InvSimpson | |||
|---|---|---|---|---|---|---|---|---|
| Mean | S.D. | Mean | S.D. | Mean | S.D. | |||
| LC | E. elliptica | Root | 0.198941 | 0.199154 | 0.110179 | 0.118500 | 1.137693 | 0.156639 |
| LC | E. elliptica | Soil | 0.292518 | 0.186952 | 0.151000 | 0.105701 | 1.19018 | 0.148864 |
| LC | P. tremuloides | Root | 0.237981 | 0.124852 | 0.124935 | 0.080647 | 1.149346 | 0.107140 |
| LC | P. tremuloides | Soil | 0.441605 | 0.409238 | 0.279218 | 0.256173 | 1.505256 | 0.506857 |
| MC | E. elliptica | Root | 0.051359 | 0.088956 | 0.022959 | 0.039766 | 1.024658 | 0.042708 |
| MC | E. elliptica | Soil | 0.431257 | 0.375213 | 0.303265 | 0.264944 | 1.561379 | 0.500268 |
| MC | P. tremuloides | Root | 0.336737 | 0.583245 | 0.180979 | 0.313464 | 1.39596 | 0.685822 |
| MC | P. tremuloides | Soil | 0.731617 | 0.509229 | 0.402308 | 0.271181 | 2.054042 | 1.260088 |
| HC | E. elliptica | Root | 0.055981 | 0.096963 | 0.0256 | 0.044341 | 1.02773 | 0.048029 |
| HC | E. elliptica | Soil | 0.085773 | 0.148563 | 0.044218 | 0.076587 | 1.05098 | 0.088301 |
| HC | P. tremuloides | Root | 0.402262 | 0.225463 | 0.209017 | 0.138528 | 1.293265 | 0.248802 |
| HC | P. tremuloides | Soil | 0.349495 | 0.605343 | 0.212032 | 0.367249 | 1.582656 | 1.009189 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, S.-J.; Kong, M.; St-Arnaud, M.; Hijri, M. Arbuscular Mycorrhizal Fungal Communities of Native Plant Species under High Petroleum Hydrocarbon Contamination Highlights Rhizophagus as a Key Tolerant Genus. Microorganisms 2020, 8, 872. https://doi.org/10.3390/microorganisms8060872
Lee S-J, Kong M, St-Arnaud M, Hijri M. Arbuscular Mycorrhizal Fungal Communities of Native Plant Species under High Petroleum Hydrocarbon Contamination Highlights Rhizophagus as a Key Tolerant Genus. Microorganisms. 2020; 8(6):872. https://doi.org/10.3390/microorganisms8060872
Chicago/Turabian StyleLee, Soon-Jae, Mengxuan Kong, Marc St-Arnaud, and Mohamed Hijri. 2020. "Arbuscular Mycorrhizal Fungal Communities of Native Plant Species under High Petroleum Hydrocarbon Contamination Highlights Rhizophagus as a Key Tolerant Genus" Microorganisms 8, no. 6: 872. https://doi.org/10.3390/microorganisms8060872