Emergence of a Novel Ehrlichia minasensis Strain, Harboring the Major Immunogenic Glycoprotein trp36 with Unique Tandem Repeat and C-Terminal Region Sequences, in Haemaphysalis hystricis Ticks Removed from Free-Ranging Sheep in Hainan Province, China


Abstract
:1. Introduction
2. Materials and Methods
2.1. Tick Collection and DNA Extraction
2.2. PCR Amplification and DNA Sequencing of the dsb, 16S rRNA, groEL, and trp36 Genes of E. minasensis
2.3. DNA Sequence Analysis and Phylogenetic Analysis
3. Results and Discussion
3.1. Identification of Tick Species
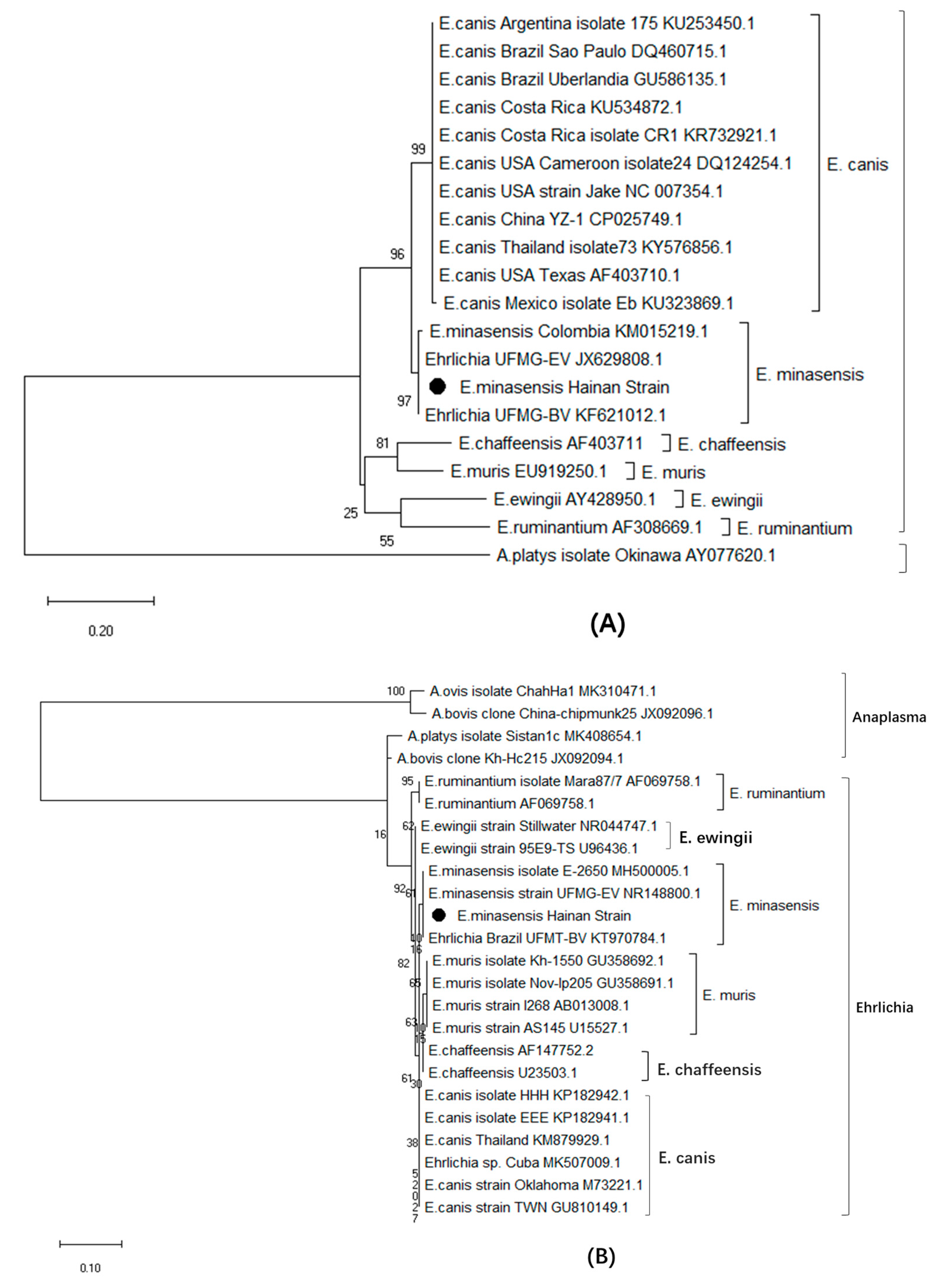
3.2. Sequence Analysis of the dsb, 16S rRNA and groEL Genes of E. minasensis
3.3. Sequence Analysis of the trp36 Gene
3.4. Sequence Analysis of the N-Terminal Region and the Upstream Tandem Repeat Region of the trp36 Gene
3.5. Sequence Analysis of the Tandem Repeat Region and C-Terminal Region of the trp36 Gene
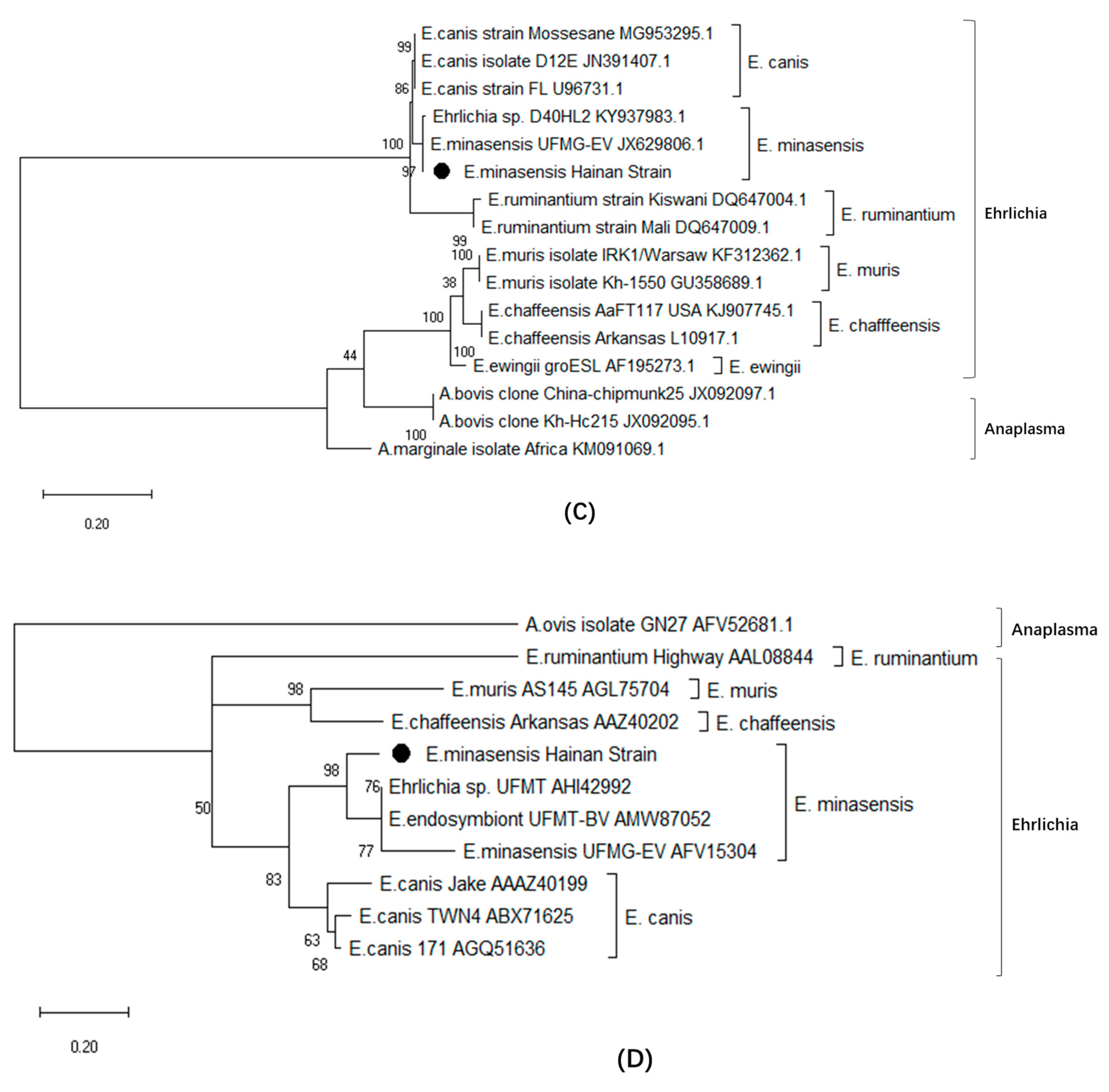
3.6. Phylogenetic Relationship Analysis Based on trp36
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Saito, T.B.; Walker, D.H. Ehrlichioses: An important one health opportunity. Vet. Sci. 2016, 3, 20. [Google Scholar] [CrossRef] [PubMed]
- Rar, V.; Golovljova, I. Anaplasma, ehrlichia, and “candidatus neoehrlichia” bacteria: Pathogenicity, biodiversity, and molecular genetic characteristics, a review. Infect. Genet. Evol. 2011, 11, 1842–1861. [Google Scholar] [CrossRef] [PubMed]
- Cabezas-Cruz, A.; Vancova, M.; Zweygarth, E.; Ribeiro, M.F.; Grubhoffer, L.; Passos, L.M. Ultrastructure of Ehrlichia mineirensis, a new member of the Ehrlichia genus. Vet. Microbiol. 2013, 167, 455–458. [Google Scholar] [CrossRef] [PubMed]
- Cabezas-Cruz, A.; Valdes, J.J.; de la Fuente, J. The glycoprotein trp36 of Ehrlichia sp. Ufmg-ev and related cattle pathogen Ehrlichia sp. Ufmt-bv evolved from a highly variable clade of E. canis under adaptive diversifying selection. Parasit. Vectors 2014, 7, 584. [Google Scholar] [CrossRef] [PubMed]
- Aguiar, D.M.; Ziliani, T.F.; Zhang, X.; Melo, A.L.; Braga, I.A.; Witter, R.; Freitas, L.C.; Rondelli, A.L.; Luis, M.A.; Sorte, E.C.; et al. A novel ehrlichia genotype strain distinguished by the trp36 gene naturally infects cattle in brazil and causes clinical manifestations associated with Ehrlichiosis. Ticks Tick Borne Dis. 2014, 5, 537–544. [Google Scholar] [CrossRef] [PubMed]
- Cruz, A.C.; Zweygarth, E.; Ribeiro, M.F.; da Silveira, J.A.; de la Fuente, J.; Grubhoffer, L.; Valdes, J.J.; Passos, L.M. New species of Ehrlichia isolated from Rhipicephalus (Boophilus) microplus shows an ortholog of the E. canis major immunogenic glycoprotein gp36 with a new sequence of tandem repeats. Parasite Vector 2012, 5, 291. [Google Scholar] [CrossRef] [PubMed]
- Gajadhar, A.A.; Lobanov, V.; Scandrett, W.B.; Campbell, J.; Al-Adhami, B. A novel Ehrlichia genotype detected in naturally infected cattle in north america. Vet. Parasitol. 2010, 173, 324–329. [Google Scholar] [CrossRef] [PubMed]
- Hailemariam, Z.; Krucken, J.; Baumann, M.; Ahmed, J.S.; Clausen, P.H.; Nijhof, A.M. Molecular detection of tick-borne pathogens in cattle from southwestern ethiopia. PLoS ONE 2017, 12, e0188248. [Google Scholar] [CrossRef] [PubMed]
- Cicculli, V.; Masse, S.; Capai, L.; de Lamballerie, X.; Charrel, R.; Falchi, A. First detection of Ehrlichia minasensis in Hyalomma marginatum ticks collected from cattle in Corsica, France. Vet. Med. Sci. 2019, 5, 243–248. [Google Scholar] [CrossRef] [PubMed]
- Thomson, K.; Yaaran, T.; Belshaw, A.; Curson, L.; Tisi, L.; Maurice, S.; Kiddle, G. A new taqman method for the reliable diagnosis of Ehrlichia spp. In canine whole blood. Parasite Vector 2018, 11. [Google Scholar] [CrossRef]
- Rehman, A.; Conraths, F.J.; Sauter-Louis, C.; Krucken, J.; Nijhof, A.M. Epidemiology of tick-borne pathogens in the semi-arid and the arid agro-ecological zones of Punjab province, Pakistan. Transbound. Emerg. Dis. 2019, 66, 526–536. [Google Scholar] [CrossRef] [PubMed]
- Iweriebor, B.C.; Mmbaga, E.J.; Adegborioye, A.; Igwaran, A.; Obi, L.C.; Okoh, A.I. Genetic profiling for Anaplasma and Ehrlichia species in ticks collected in the Eastern Cape Province of South Africa. BMC Microbiol. 2017, 17, 45. [Google Scholar] [CrossRef] [PubMed]
- Cabezas-Cruz, A.; Zweygarth, E.; Aguiar, D.M. Ehrlichia minasensis, an old demon with a new name. Ticks Tick Borne Dis. 2019, 10, 828–829. [Google Scholar] [CrossRef] [PubMed]
- Parola, P.; Cornet, J.P.; Sanogo, Y.O.; Miller, R.S.; Thien, H.V.; Gonzalez, J.P.; Raoult, D.; Telford, I.S.; Wongsrichanalai, C. Detection of Ehrlichia spp., Anaplasma spp., Rickettsia spp., and other eubacteria in ticks from the thai-myanmar border and vietnam. J. Clin. Microbiol. 2003, 41, 1600–1608. [Google Scholar] [CrossRef] [PubMed]
- Khatri-Chhetri, R.; Wang, H.C.; Chen, C.C.; Shih, H.C.; Liao, H.C.; Sun, C.M.; Khatri-Chhetri, N.; Wu, H.Y.; Pei, K.J. Surveillance of ticks and associated pathogens in free-ranging Formosan pangolins (Manis pentadactyla pentadactyla). Ticks Tick Borne Dis. 2016, 7, 1238–1244. [Google Scholar] [CrossRef]
- Nicholson, M.C.; Mather, T.N. Methods for evaluating lyme disease risks using geographic information systems and geospatial analysis. J. Med. Entomol. 1996, 33, 711–720. [Google Scholar] [CrossRef]
- Black, W.C.; Piesman, J. Phylogeny of hard- and soft-tick taxa (acari: Ixodida) based on mitochondrial 16s rdna sequences. Proc. Natl. Acad. Sci. USA 1994, 91, 10034–10038. [Google Scholar] [CrossRef]
- Fang, Y.; Shi, W.Q.; Zhang, Y. Molecular phylogeny of Anopheles hyrcanus group (diptera: Culicidae) based on mtdna coi. Infect. Dis. Poverty 2017, 6, 61. [Google Scholar] [CrossRef]
- Cicuttin, G.L.; De Salvo, M.N.; Gury Dohmen, F.E. Molecular characterization of Ehrlichia canis infecting dogs, buenos aires. Ticks Tick Borne Dis. 2016, 7, 954–957. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. Mega x: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Geiger, J.; Morton, B.A.; Vasconcelos, E.J.R.; Tngrian, M.; Kachani, M.; Barron, E.A.; Gavidia, C.M.; Gilman, R.H.; Angulo, N.P.; Lerner, R.; et al. Molecular characterization of tandem repeat protein 36 gene of Ehrlichia canis detected in naturally infected dogs from Peru. Am. J. Trop. Med. Hyg. 2018, 99, 297–302. [Google Scholar] [CrossRef] [PubMed]
- Doyle, C.K.; Nethery, K.A.; Popov, V.L.; McBride, J.W. Differentially expressed and secreted major immunoreactive protein orthologs of Ehrlichia canis and E. Chaffeensis elicit early antibody responses to epitopes on glycosylated tandem repeats. Infect. Immun 2006, 74, 711–720. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, I.T.S.; Melo, A.L.T.; Freitas, L.C.; Vercoza, R.V.; Alves, A.S.; Costa, J.S.; Chitarra, C.S.; Nakazato, L.; Dutra, V.; Pacheco, R.C.; et al. Minimum infection rate of Ehrlichia minasensis in Rhipicephalus microplus and Amblyomma sculptum ticks in brazil. Ticks Tick Borne Dis. 2016, 7, 849–852. [Google Scholar] [CrossRef] [PubMed]
- Machado, R.Z.; Andre, M.R.; Werther, K.; de Sousa, E.; Gavioli, F.A.; Alves Junior, J.R. Migratory and carnivorous birds in brazil: Reservoirs for Anaplasma and Ehrlichia species? Vector Borne Zoonotic Dis. 2012, 12, 705–708. [Google Scholar] [CrossRef] [PubMed]
- Bjoersdorff, A.; Bergstrom, S.; Massung, R.F.; Haemig, P.D.; Olsen, B. Ehrlichia-infected ticks on migrating birds. Emerg. Infect. Dis. 2001, 7, 877–879. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Species | Target | Primer Name | Sequence | PCR Condition | Length | References |
|---|---|---|---|---|---|---|
| ticks | 16S rDNA | 16S+1 16S-1 | CCGGTCTGAACTCAGATCAAG CTGCTCAATGATTTTTTAAATTGCTGTGG | 95 °C 5 min, 35 × (95 °C 30 s, 57 °C 30 s, 72 °C 40 s), 72 °C 10 min | 460 bp | [17] |
| cox1 | LCO1490 HCO2198 | GGTCAACAAATCATAAAGATATTGG TAAACTTCAGGGTGACCAAAAAATCA | 95 °C 5 min, 35 × (95 °C 30 s, 57 °C 30 s, 72 °C 40 s), 72 °C 10 min | 650 bp | [18] | |
| Ehrlichia minasensis | dsb | dsb-330 dsb-728 | GATGATGTCTGAAGATATGAAACAAAT CTGCTCGTCTATTTTACTTCTTAAAGT | 94 °C 5 min, 35 × (94 °C 30 s, 50.5 °C 60 s, 72 °C 60 s), 72 °C 10 min | 400 bp | [19] |
| 16S rRNA | Ehr-16S-D Ehr-16S-R | GGTACCYACAGAAGAAGTCC TAGCACTCATCGTTTACAGC | 94 °C 5 min, 35 × (94 °C 30 s, 54 °C 60 s, 72 °C 60 s), 72 °C 10 min | 345 bp | [9] | |
| groEL | Ehr-groel-F Ehr-groel-R | GTTGAAAARACTGATGGTATGCA ACACGRTCTTTACGYTCYTTAAC | 94 °C 5 min, 35 × (94 °C 30 s, 55 °C 60 s, 72 °C 60 s), 72 °C 10 min | 590 bp | [9] | |
| trp36 | TRP36-F2 TRP36-R1 | TTTAAAACAAAATTAACACACTA AAGATTAACTTAATACTCAATATTACT | 94 °C 5 min, 35 × (94 °C 30 s, 46 °C 60 s, 72 °C 60 s), 72 °C 10 min | 800–1000 bp | [17] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, J.; Liu, X.; Mu, J.; Yu, X.; Fei, Y.; Chang, J.; Bi, Y.; Zhou, Y.; Ding, Z.; Yin, R. Emergence of a Novel Ehrlichia minasensis Strain, Harboring the Major Immunogenic Glycoprotein trp36 with Unique Tandem Repeat and C-Terminal Region Sequences, in Haemaphysalis hystricis Ticks Removed from Free-Ranging Sheep in Hainan Province, China. Microorganisms 2019, 7, 369. https://doi.org/10.3390/microorganisms7090369
Li J, Liu X, Mu J, Yu X, Fei Y, Chang J, Bi Y, Zhou Y, Ding Z, Yin R. Emergence of a Novel Ehrlichia minasensis Strain, Harboring the Major Immunogenic Glycoprotein trp36 with Unique Tandem Repeat and C-Terminal Region Sequences, in Haemaphysalis hystricis Ticks Removed from Free-Ranging Sheep in Hainan Province, China. Microorganisms. 2019; 7(9):369. https://doi.org/10.3390/microorganisms7090369
Chicago/Turabian StyleLi, Junjiao, Xinxin Liu, Jiaqi Mu, Xibing Yu, Yidong Fei, Jin Chang, Yuhai Bi, Yulong Zhou, Zhuang Ding, and Renfu Yin. 2019. "Emergence of a Novel Ehrlichia minasensis Strain, Harboring the Major Immunogenic Glycoprotein trp36 with Unique Tandem Repeat and C-Terminal Region Sequences, in Haemaphysalis hystricis Ticks Removed from Free-Ranging Sheep in Hainan Province, China" Microorganisms 7, no. 9: 369. https://doi.org/10.3390/microorganisms7090369