Mode of Action of Dietary Dexamethasone May Not Be Dependent Upon Microbial Mechanisms in Broilers

Abstract
1. Introduction
2. Materials and Methods
2.1. Serum FITC-d Recovery
2.2. Bacterial Translocation and Recovery
2.3. Experimental Animals
2.3.1. Experiment 1: Effect of Dietary BMD50 Supplementation on Dexamethasone-Mediated Changes in Intestinal Permeability
2.3.2. Experiment 2: Effect of Dietary Baytril Supplementation on Dexamethasone-Mediated Intestinal Permeability and Enteric Bacterial Translocation
2.3.3. Experiment 3: Effect of Dietary Dexamethasone on Intestinal Permeability and Translocation of Culturable, Differential, Enteric Bacteria
2.4. Statistical Analysis
3. Results
3.1. Body Weight
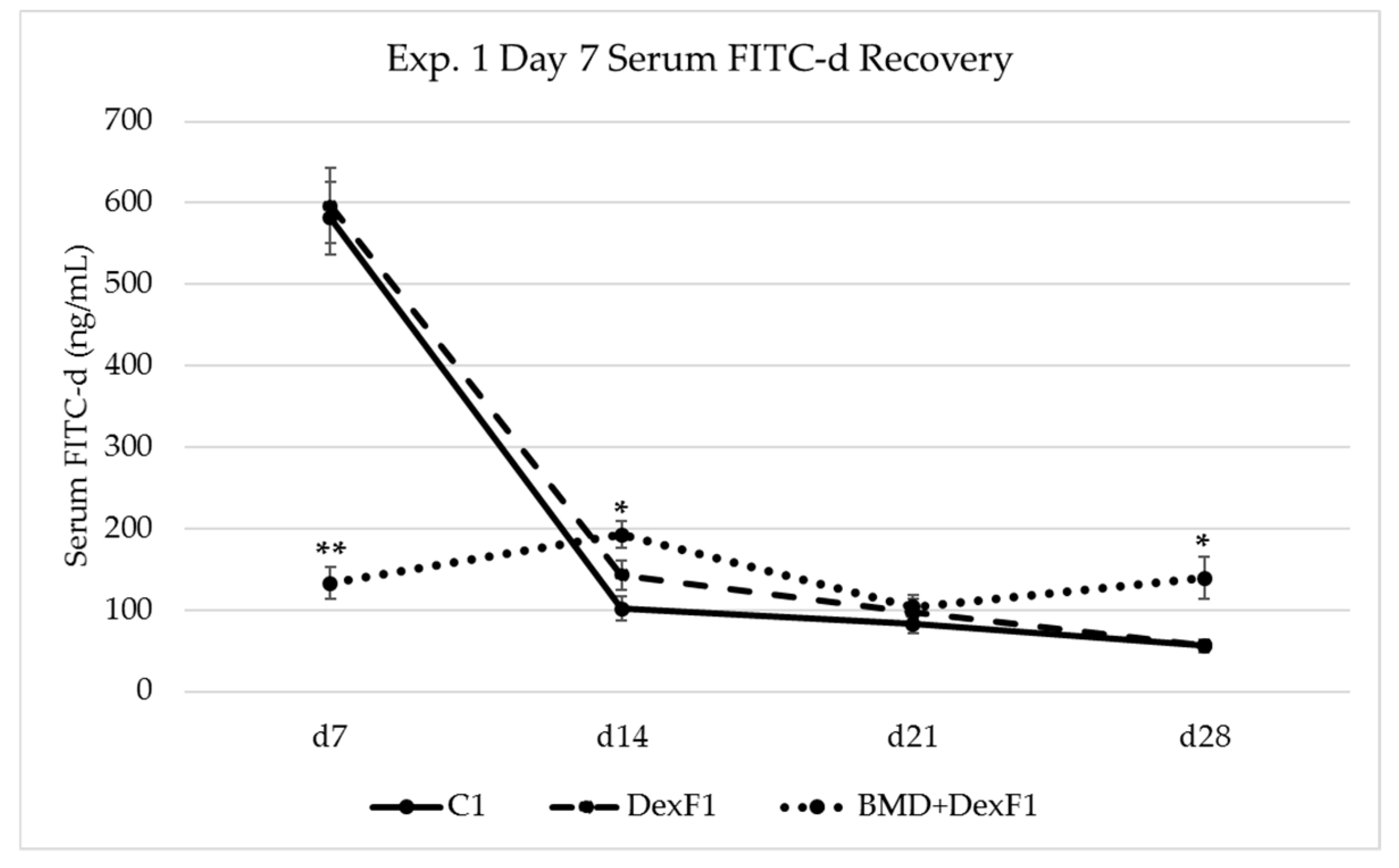
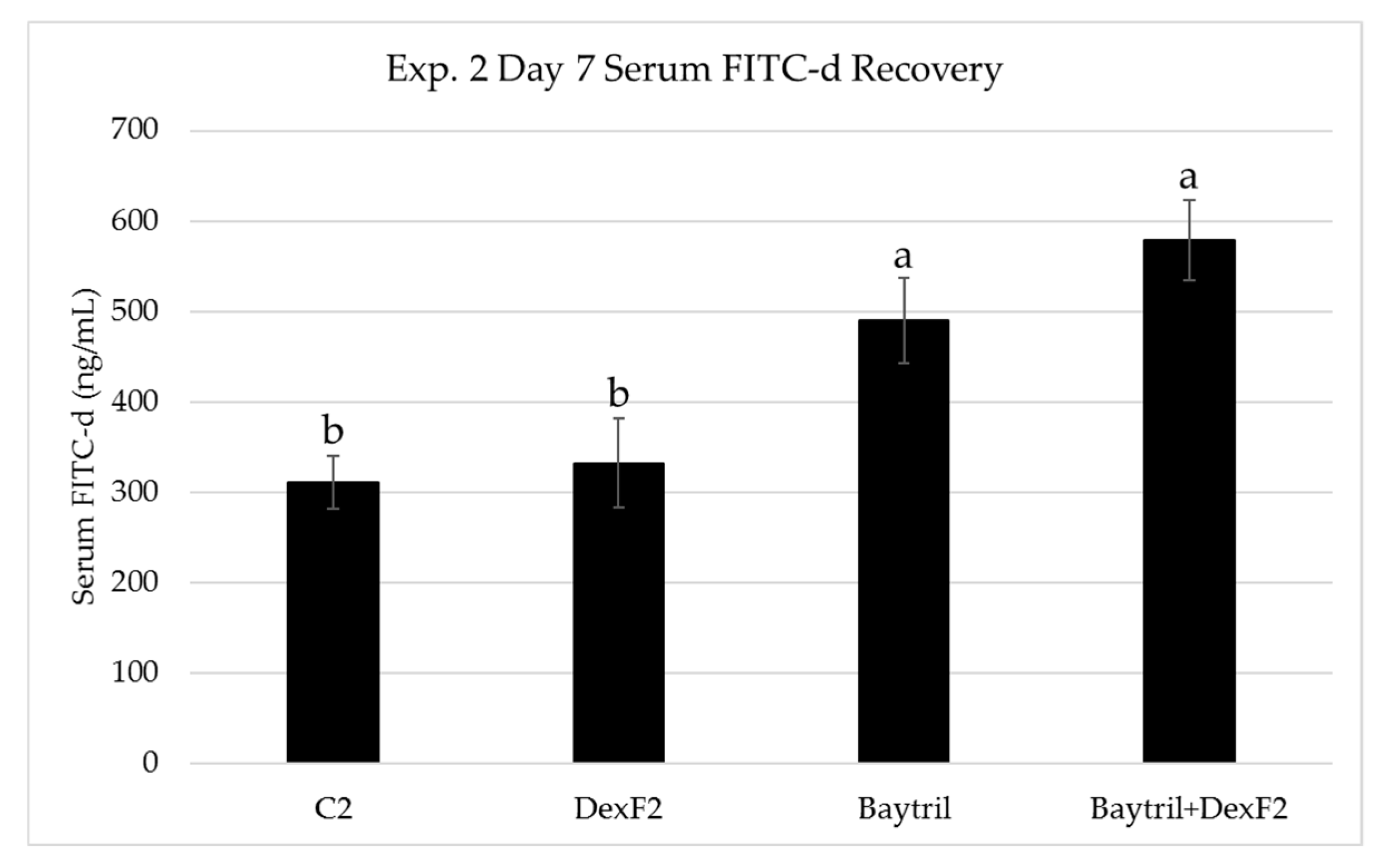
3.2. Serum FITC-d Recovery
3.3. Bacterial Translocation and Recovery
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lopetuso, L.R.; Scaldaferri, F.; Bruno, G.; Petito, V.; Franceschi, F.; Gasbarrini, A. The therapeutic management of gut barrier leaking: The emerging role for mucosal barrier protectors. Eur. Rev. Med. Pharmacol. Sci. 2015, 9, 1068–1076. [Google Scholar]
- Söderholm, J.D.; Perdue, M.H., II. Stress and intestinal barrier function. Am. J. Physiol.-Gastrointest. Liver Physiol. 2001, 280, G7–G13. [Google Scholar] [CrossRef] [PubMed]
- Groschwitz, K.R.; Hogan, S.P. Intestinal barrier function: Molecular regulation and disease pathogenesis. J. Allergy Clin. Immunol. 2009, 124, 3–20. [Google Scholar] [CrossRef] [PubMed]
- Awad, W.; Hess, C.; Hess, M. Enteric pathogens and their toxin-induced disruption of the intestinal barrier through alteration of tight junctions in chickens. Toxins 2017, 9, 60. [Google Scholar] [CrossRef] [PubMed]
- Kelly, J.R.; Kennedy, P.J.; Cryan, J.F.; Dinan, T.G.; Clarke, G.; Hyland, N.P. Breaking down the barriers: The gut microbiome, intestinal permeability and stress-related psychiatric disorders. Front. Cell. Neurosci. 2015, 9, 392. [Google Scholar] [CrossRef]
- Rodiño-Janeiro, B.K.; Alonso-Cotoner, C.; Pigrau, M.; Lobo, B.; Vicario, M.; Santos, J. Role of Corticotropin-releasing Factor in Gastrointestinal Permeability. J. Neurogastroenterol. Motil. 2015, 21, 33–50. [Google Scholar] [CrossRef]
- Siegel, H.S. Stress, strains and resistance. Br. Poult. Sci. 1995, 36, 3–22. [Google Scholar] [CrossRef]
- Siegel, H.S. Physiological Stress in Birds. BioScience 1980, 30, 529–534. [Google Scholar] [CrossRef]
- Eid, D.Y.; Ebeid, T.; Younis, H. Vitamin E supplementation reduces dexamethasone-induced oxidative stress in chicken semen. Br. Poult. Sci. 2006, 47, 350–356. [Google Scholar] [CrossRef]
- Freeman, B.M. Stress and the Domestic Fowl: a Physiological Appraisal. Worlds Poult. Sci. J. 1971, 27, 263–275. [Google Scholar] [CrossRef]
- Klasing, K.C.; Laurin, D.E.; Peng, R.K.; Fry, M.D. Immunologically mediated growth depression in chicks: Influence of feed intake, corticosterone and interleukin-1. J. Nutr. 1987, 117, 1629–1637. [Google Scholar] [CrossRef] [PubMed]
- Yegani, M.; Korver, D.R. Factors affecting intestinal health in poultry. Poult. Sci. 2008, 87, 2052–2063. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Song, Z.; Kerr, K.A.; Moeser, A.J. Chronic social stress in pigs impairs intestinal barrier and nutrient transporter function, and alters neuro-immune mediator and receptor expression. PLoS ONE 2017, 12, e0171617. [Google Scholar] [CrossRef] [PubMed]
- Bartov, I.; Jensen, L.S.; Veltmann, J.R. Effect of Corticosterone and Prolactin on Fattening in Broiler Chicks. Poult. Sci. 1980, 59, 1328–1334. [Google Scholar] [CrossRef] [PubMed]
- Kannan, G.; Heath, J.L.; Wabeck, C.J.; Souza, M.C.; Howe, J.C.; Mench, J.A. Effects of crating and transport on stress and meat quality characteristics in broilers. Poult. Sci. 1997, 76, 523–529. [Google Scholar] [CrossRef]
- Wideman, R.F.; Pevzner, I. Dexamethasone triggers lameness associated with necrosis of the proximal tibial head and proximal femoral head in broilers. Poult. Sci. 2012, 91, 2464–2474. [Google Scholar] [CrossRef]
- Tache, Y.; Perdue, M.H. Role of peripheral CRF signalling pathways in stress-related alterations of gut motility and mucosal function. Neurogastroenterol. Motil. 2004, 16, 137–142. [Google Scholar] [CrossRef]
- Spitz, J.; Hecht, G.; Taveras, M.; Aoys, E.; Alverdy, J. The effect of dexamethasone administration on rat intestinal permeability: The role of bacterial adherence. Gastroenterology 1994, 106, 35–41. [Google Scholar] [CrossRef]
- Huff, G.R.; Huff, W.E.; Balog, J.M.; Rath, N.C. The effects of dexamethasone immunosuppression on turkey osteomyelitis complex in an experimental Escherichia coli respiratory infection. Poult. Sci. 1998, 77, 654–661. [Google Scholar] [CrossRef]
- Zong, Y.; Zhu, S.; Zhang, S.; Zheng, G.; Wiley, J.W.; Hong, S. Chronic stress and intestinal permeability: Lubiprostone regulates glucocorticoid receptor-mediated changes in colon epithelial tight junction proteins, barrier function, and visceral pain in the rodent and human. Neurogastroenterol. Motil. 2019, 31, e13477. [Google Scholar] [CrossRef]
- Li, Y.; Cai, H.Y.; Liu, G.H.; Dong, X.L.; Chang, W.H.; Zhang, S.; Zheng, A.J.; Chen, G.L. Effects of stress simulated by dexamethasone on jejunal glucose transport in broilers. Poult. Sci. 2009, 88, 330–337. [Google Scholar] [CrossRef] [PubMed]
- Vicuña, E.A.; Kuttappan, V.A.; Galarza-Seeber, R.; Latorre, J.D.; Faulkner, O.B.; Hargis, B.M.; Tellez, G.; Bielke, L.R. Effect of dexamethasone in feed on intestinal permeability, differential white blood cell counts, and immune organs in broiler chicks. Poult. Sci. 2015, 94, 2075–2080. [Google Scholar] [CrossRef]
- Söderholm, J.D.; Yang, P.; Ceponis, P.; Vohra, A.; Riddell, R.; Sherman, P.M.; Perdue, M.H. Chronic stress induces mast cell–dependent bacterial adherence and initiates mucosal inflammation in rat intestine. Gastroenterology 2002, 123, 1099–1108. [Google Scholar] [CrossRef]
- Scanes, C.G. Biology of stress in poultry with emphasis on glucocorticoids and the heterophil to lymphocyte ratio. Poult. Sci. 2016, 95, 2208–2215. [Google Scholar] [CrossRef]
- Santos, J.; Benjamin, M.; Yang, P.-C.; Prior, T.; Perdue, M.H. Chronic stress impairs rat growth and jejunal epithelial barrier function: role of mast cells. Am. J. Physiol.-Gastrointest. Liver Physiol. 2000, 278, G847–G854. [Google Scholar] [CrossRef]
- Alverdy, J.; Aoys, E. The effect of glucocorticoid administration on bacterial translocation. Evidence for an acquired mucosal immunodeficient state. Ann. Surg. 1991, 214, 719–723. [Google Scholar] [CrossRef]
- Bailey, M.T.; Dowd, S.E.; Parry, N.M.A.; Galley, J.D.; Schauer, D.B.; Lyte, M. Stressor Exposure Disrupts Commensal Microbial Populations in the Intestines and Leads to Increased Colonization by Citrobacter rodentium. Infect. Immun. 2010, 78, 1509–1519. [Google Scholar] [CrossRef]
- Bailey, M.T.; Engler, H.; Sheridan, J.F. Stress induces the translocation of cutaneous and gastrointestinal microflora to secondary lymphoid organs of C57BL/6 mice. J. Neuroimmunol. 2006, 171, 29–37. [Google Scholar] [CrossRef]
- Knowles, S.R.; Nelson, E.A.; Palombo, E.A. Investigating the role of perceived stress on bacterial flora activity and salivary cortisol secretion: A possible mechanism underlying susceptibility to illness. Biol. Psychol. 2008, 77, 132–137. [Google Scholar] [CrossRef]
- Everest, P. Stress and bacteria: microbial endocrinology. Gut 2007, 56, 1037–1038. [Google Scholar] [CrossRef]
- Ünsal, H.; Balkaya, M.; Ünsal, C.; Bıyık, H.; Başbülbül, G.; Poyrazoğlu, E. The Short-Term Effects of Different Doses of Dexamethasone on the Numbers of some Bacteria in the Ileum. Dig. Dis. Sci. 2008, 53, 1842–1845. [Google Scholar] [CrossRef]
- Proctor, A.; Phillips, G. Frontiers | Differential Effects of Bacitracin Methylene Disalicylate (BMD) on the Distal Colon and Cecal Microbiota of Young Broiler Chickens | Veterinary Science. Front. Vet. Sci. 2019, 6, 114. [Google Scholar] [CrossRef]
- Kuttappan, V.A.; Vicuña, E.A.; Latorre, J.D.; Wolfenden, A.D.; Téllez, G.I.; Hargis, B.M.; Bielke, L.R. Evaluation of gastrointestinal leakage in multiple enteric inflammation models in chickens. Front. Vet. Sci. 2015, 2, 66. [Google Scholar] [CrossRef]
- Vicuña, E.A.; Kuttappan, V.A.; Tellez, G.; Hernandez-Velasco, X.; Seeber-Galarza, R.; Latorre, J.D.; Faulkner, O.B.; Wolfenden, A.D.; Hargis, B.M.; Bielke, L.R. Dose titration of FITC-D for optimal measurement of enteric inflammation in broiler chicks. Poult. Sci. 2015, 94, 1353–1359. [Google Scholar] [CrossRef]
- National Research Council (U.S.). Nutrient Requirements of Poultry, 9th revised ed.; National Academies Press: Washington, DC, USA, 1994. [Google Scholar]
- Ghosh, D.; Vogt, A. Outliers: An Evaluation of Methodologies. In Proceedings of the Joint Statistical Meetings, San Diego, CA, USA, 28 July–2 August 2012; pp. 3455–3460. [Google Scholar]
- Tsigos, C.; Kyrou, I.; Kassi, E.; Chrousos, G.P. Stress, Endocrine Physiology and Pathophysiology. Available online: https://www.endotext.org/ (accessed on 1 July 2019).
- Rose, M.E. Immunity to coccidiosis: Effect of betamethasone treatment of fowls on Eimeria mivati infection. Parasitology 1970, 60, 137–146. [Google Scholar] [CrossRef]
- Schaffner, A. Therapeutic concentrations of glucocorticoids suppress the antimicrobial activity of human macrophages without impairing their responsiveness to gamma interferon. J. Clin. Investig. 1985, 76, 1755–1764. [Google Scholar] [CrossRef]
- Gomes, A.V.S.; Quinteiro-Filho, W.M.; Ribeiro, A.; Ferraz-de-Paula, V.; Pinheiro, M.L.; Baskeville, E.; Akamine, A.T.; Astolfi-Ferreira, C.S.; Ferreira, A.J.P.; Palermo-Neto, J. Overcrowding stress decreases macrophage activity and increases Salmonella enteritidis invasion in broiler chickens. Avian Pathol. 2014, 43, 82–90. [Google Scholar] [CrossRef]
- Teitelbaum, A.A.; Gareau, M.G.; Jury, J.; Yang, P.C.; Perdue, M.H. Chronic peripheral administration of corticotropin-releasing factor causes colonic barrier dysfunction similar to psychological stress. Am. J. Physiol.-Gastrointest. Liver Physiol. 2008, 295, G452–G459. [Google Scholar] [CrossRef]
- Karalis, K.; Sano, H.; Redwine, J.; Listwak, S.; Wilder, R.L.; Chrousos, G.P. Autocrine or Paracrine Inflammatory Actions of Corticotropin-Releasing Hormone in Vivo. Sci. Wash. 1991, 254, 421–423. [Google Scholar] [CrossRef]
- Elenkov, I.J.; Chrousos, G.P. Stress system--organization, physiology and immunoregulation. Neuroimmunomodulation 2006, 13, 257–267. [Google Scholar] [CrossRef]
- Chrousos, G.P. The Stress Response and Immune Function: Clinical Implications: The 1999 Novera H. Spector Lecture. Ann. N. Y. Acad. Sci. 2000, 917, 38–67. [Google Scholar] [CrossRef]
- Isobe, T.; Lillehoj, H.S. Dexamethasone suppresses T cell-mediated immunity and enhances disease susceptibility to Eimeria mivati infection. Vet. Immunol. Immunopathol. 1993, 39, 431–446. [Google Scholar] [CrossRef]
- Lv, Z.-P.; Peng, Y.-Z.; Zhang, B.-B.; Fan, H.; Liu, D.; Guo, Y.-M. Glucose and lipid metabolism disorders in the chickens with dexamethasone-induced oxidative stress. J. Anim. Physiol. Anim. Nutr. 2018, 102, e706–e717. [Google Scholar] [CrossRef]
- Wang, X.J.; Wei, D.L.; Song, Z.G.; Jiao, H.C.; Lin, H. Effects of Fatty Acid Treatments on the Dexamethasone-Induced Intramuscular Lipid Accumulation in Chickens. PLoS ONE 2012, 7, e36663. [Google Scholar] [CrossRef]
- Ilan, Y. Leaky gut and the liver: A role for bacterial translocation in nonalcoholic steatohepatitis. World J. Gastroenterol. 2012, 18, 2609–2618. [Google Scholar] [CrossRef]
- Baxter, M.F.A.; Merino-Guzman, R.; Latorre, J.D.; Mahaffey, B.D.; Yang, Y.; Teague, K.D.; Graham, L.E.; Wolfenden, A.D.; Hernandez-Velasco, X.; Bielke, L.R.; et al. Optimizing fluorescein isothiocyanate dextran measurement as a biomarker in a 24-h feed restriction model to induce gut permeability in broiler chickens. Front. Vet. Sci. 2017, 4, 56. [Google Scholar] [CrossRef]
- Tellez, G.; Latorre, J.D.; Kuttappan, V.A.; Kogut, M.H.; Wolfenden, A.; Hernandez-Velasco, X.; Hargis, B.M.; Bottje, W.G.; Bielke, L.R.; Faulkner, O.B. Utilization of rye as energy source affects bacterial translocation, intestinal viscosity, microbiota composition, and bone mineralization in broiler chickens. Front. Genet. 2014, 5, 339. [Google Scholar] [CrossRef]
- Hamal, K.R.; Burgess, S.C.; Pevzner, I.Y.; Erf, G.F. Maternal Antibody Transfer from Dams to Their Egg Yolks, Egg Whites, and Chicks in Meat Lines of Chickens. Poult. Sci. 2006, 85, 1364–1372. [Google Scholar] [CrossRef]
- Morales-Barrera, E.; Calhoun, N.; Lobato-Tapia, J.L.; Lucca, V.; Prado-Rebolledo, O.; Hernandez-Velasco, X.; Merino-Guzman, R.; Petrone-García, V.M.; Latorre, J.D.; Mahaffey, B.D.; et al. Risks Involved in the Use of Enrofloxacin for Salmonella enteritidis or Salmonella heidelberg in Commercial Poultry. Front. Vet. Sci. 2016, 3, 72. [Google Scholar] [CrossRef]
- Van Ampting, M.T.J.; Schonewille, A.J.; Vink, C.; Brummer, R.J.M.; van der Meer, R.; Bovee-Oudenhoven, I.M.J. Damage to the Intestinal Epithelial Barrier by Antibiotic Pretreatment of Salmonella-Infected Rats Is Lessened by Dietary Calcium or Tannic Acid. J. Nutr. 2010, 140, 2167–2172. [Google Scholar] [CrossRef]
- Lyte, M.; Vulchanova, L.; Brown, D.R. Stress at the intestinal surface: catecholamines and mucosa–bacteria interactions. Cell Tissue Res. 2011, 343, 23–32. [Google Scholar] [CrossRef]
- Williams, P.H.; Rabsch, W.; Methner, U.; Voigt, W.; Tschäpe, H.; Reissbrodt, R. Catecholate receptor proteins in Salmonella enterica: Role in virulence and implications for vaccine development. Vaccine 2006, 24, 3840–3844. [Google Scholar] [CrossRef]
- Coulanges, V.; Andre, P.; Ziegler, O.; Buchheit, L.; Vidon, D.J. Utilization of iron-catecholamine complexes involving ferric reductase activity in Listeria monocytogenes. Infect. Immun. 1997, 65, 2778–2785. [Google Scholar]



{kind=link}
{kind=link}
| BWG (g) | d0–7 | d7–14 | d14–21 | d21–28 | d14–28 |
|---|---|---|---|---|---|
| C1 | 135.17 ± 1.76 ab | 295.07 ± 3.48 a | 446.16 ± 5.92 a | 645.24 ± 7.89 a | 1089.20 ± 12.71 a |
| DexF1 | 139.14 ± 1.56 a | 178.36 ± 1.71 b | 337.05 ± 3.39 c | 589.32 ± 5.57 b | 926.52 ± 8.37 b |
| BMD+DexF1 | 133.57 ± 1.37 b | 180.87 ± 1.62 b | 351.58 ± 3.62 b | 564.75 ± 7.24 c | 916.26 ± 9.75 b |
| Bacterial Recovery % Positive (Log10 CFU/g of Tissue) | |||
|---|---|---|---|
| Total Aerobic Bacteria (Liver) | Lactose Positive (Ceca) | Lactose Negative (Ceca) | |
| C2 | 33% (0.73 ± 0.32) | 92% (7.07 ± 0.26) a | 92% (7.22 ± 0.19) a |
| DexF2 | 8% (0.14 ± 0.14) | 83% (7.58 ± 0.21) a | 83% (5.00 ± 1.10) a |
| Baytril | 17% (0.33 ± 0.23) | 0% (0.00 ± 0.00) b | 0% (0.00 ± 0.00) b |
| Baytril+DexF2 | 25% (0.47 ± 0.25) | 0% (0.00 ± 0.00) b | 0% (0.00 ± 0.00) b |
| P-value | 0.13 | <0.001 | <0.001 |
| Bacterial Recovery % Positive (Log10 CFU/g of Tissue) | ||||||||
|---|---|---|---|---|---|---|---|---|
| Total Aerobic Bacteria | Lactose positive | Lactose negative | ||||||
| Liver | Ceca | Liver | Ileum | Ceca | Liver | Ileum | Ceca | |
| C3 | 76% (2.27 ± 0.31) | 8.38 ± 0.11 | 16% (0.80 ± 0.30) | 48% (1.54 ± 0.33) | 100% (6.26 ± 0.18) | 16% (0.46 ± 0.21) a | 32% (1.04 ± 0.31) | 56% (3.27 ± 0.60) |
| DexF3 | 68% (2.04 ± 0.30) | 8.20 ± 0.26 | 12% (0.40 ± 0.22) | 40% (1.96 ± 0.44) | 96% (6.44 ± 0.33) | 0% (0.00 ± 0.00) b | 13% (0.35 ± 0.19) | 48% (3.03 ± 0.63) |
| P-value | 0.53 | -† | 0.68 | 0.57 | 0.31 | 0.04 | 0.09 | 0.57 |
| Differential Bacterial Recovery % Positive (Log10 CFU/g) | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| E. coli | Enterococcus/Klebsiella1 | Pseudomonas/Staphylococcus1 | |||||||
| Liver | Ileum | Ceca | Liver | Ileum | Ceca | Liver | Ileum | Ceca | |
| C3 | 24% (0.87 ± 0.32) | 60% (4.56 ± 0.78) a | 100% (7.21 ± 0.20) | 44% (2.10 ± 0.40) | 8.23 ± 0.15 | 8.90 ± 0.15 | 8% (0.44 ± 0.25) | 0% (0.00 ± 0.00) | 28% (1.79 ± 0.60) |
| DexF3 | 12% (0.65 ± 0.27) | 32% (2.29 ± 0.70) b | 96% (6.90 ± 0.38) | 40% (1.87 ± 0.36) | 7.86 ± 0.24 | 8.80 ± 0.24 | 8% (0.23 ± 0.16) | 4% (0.16 ± 0.16) | 20% (1.85 ± 0.61) |
| P-value | 0.27 | 0.05 | 0.31 | 0.77 | -† | -† | 1.00 | 0.31 | 0.51 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Duff, A.F.; Baxter, M.F.A.; Graham, B.D.; Hargis, B.M.; Bielke, L.R. Mode of Action of Dietary Dexamethasone May Not Be Dependent Upon Microbial Mechanisms in Broilers. Microorganisms 2019, 7, 346. https://doi.org/10.3390/microorganisms7090346
Duff AF, Baxter MFA, Graham BD, Hargis BM, Bielke LR. Mode of Action of Dietary Dexamethasone May Not Be Dependent Upon Microbial Mechanisms in Broilers. Microorganisms. 2019; 7(9):346. https://doi.org/10.3390/microorganisms7090346
Chicago/Turabian StyleDuff, Audrey F., Mikayla F. A. Baxter, B. Danielle Graham, Billy M. Hargis, and Lisa R. Bielke. 2019. "Mode of Action of Dietary Dexamethasone May Not Be Dependent Upon Microbial Mechanisms in Broilers" Microorganisms 7, no. 9: 346. https://doi.org/10.3390/microorganisms7090346
APA StyleDuff, A. F., Baxter, M. F. A., Graham, B. D., Hargis, B. M., & Bielke, L. R. (2019). Mode of Action of Dietary Dexamethasone May Not Be Dependent Upon Microbial Mechanisms in Broilers. Microorganisms, 7(9), 346. https://doi.org/10.3390/microorganisms7090346