The Root Endophytic Fungi Community Structure of Pennisetum sinese from Four Representative Provinces in China

Abstract

1. Introduction
2. Materials and Methods
2.1. Sampling Sites and Sample Collection
2.2. DNA Extraction and Illumina Sequencing
2.3. Data Analysis
3. Results
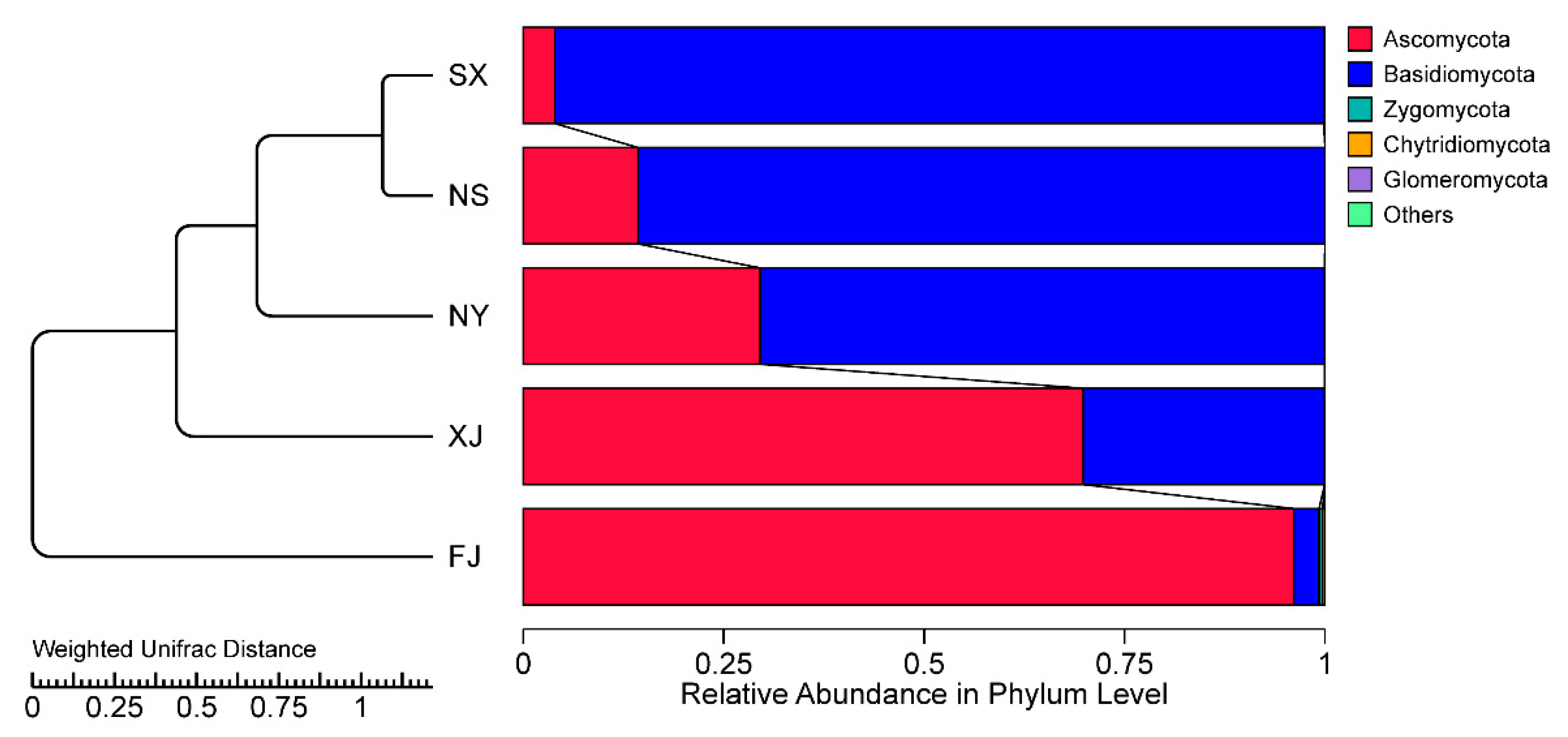
3.1. Endophytic Fungal Community in Illumina Sequencing
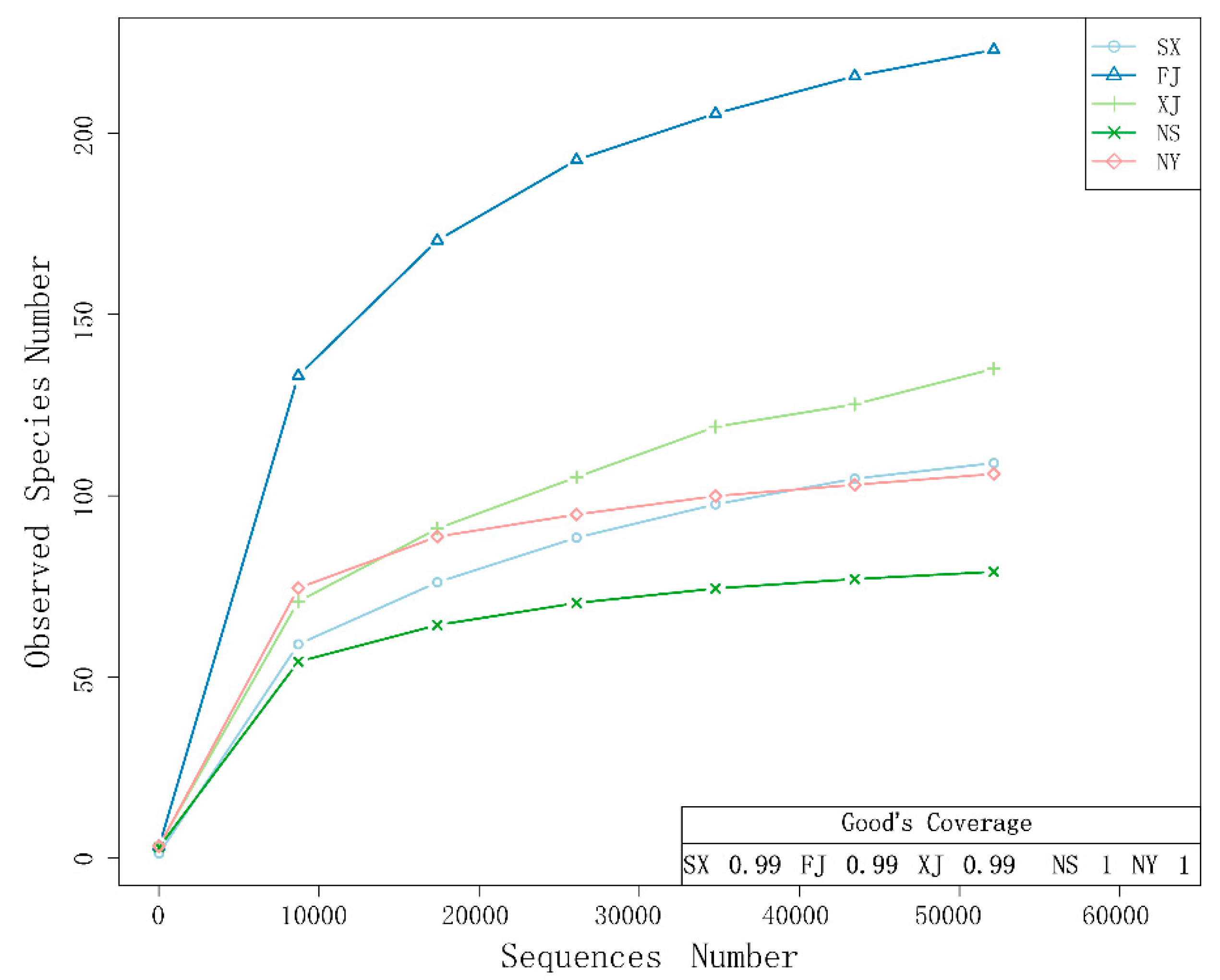
3.2. Alpha Diversity Analysis
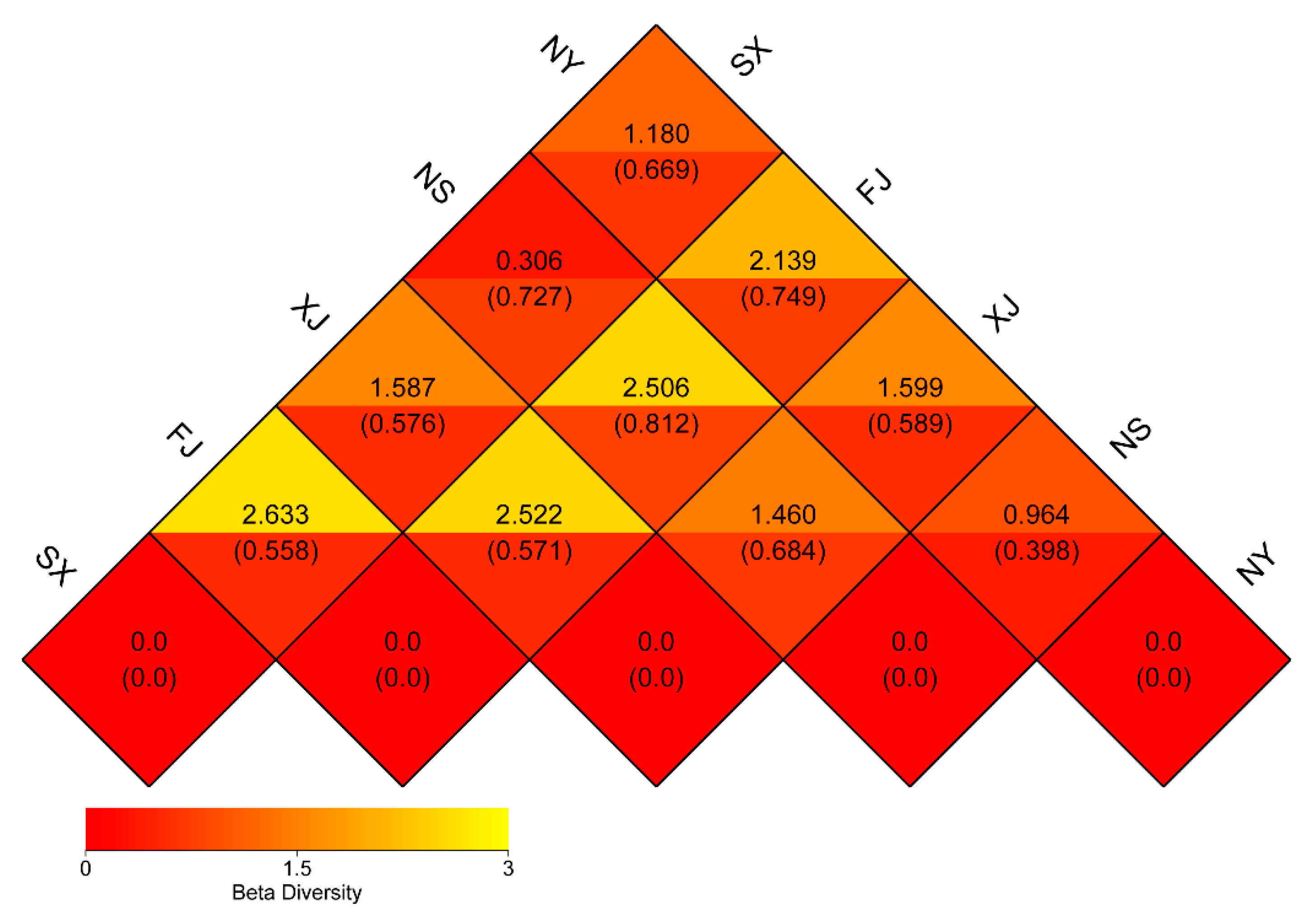
3.3. Beta Diversity Analysis
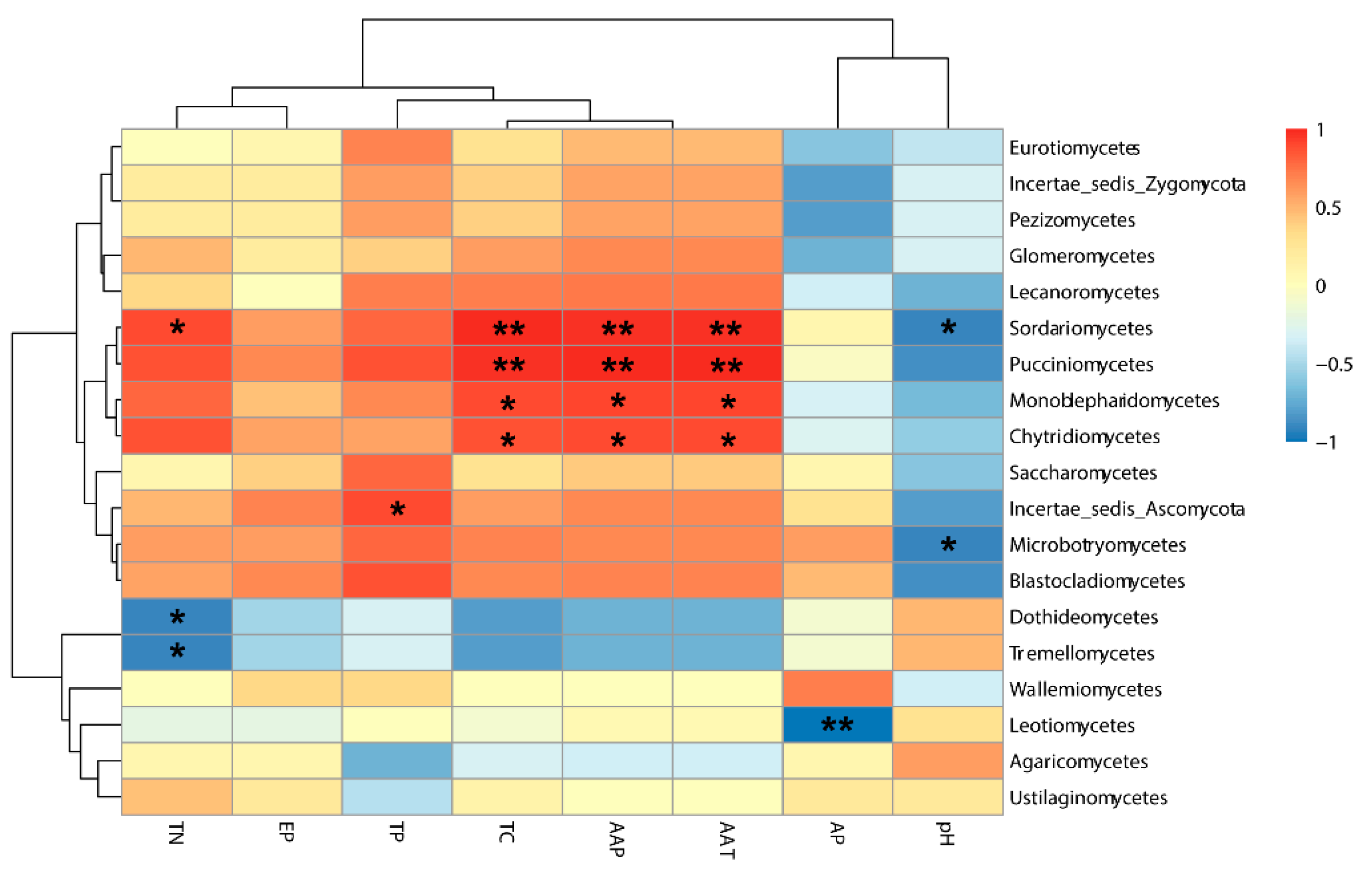
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Deng, L.; Shangguan, Z.P.; Sweeney, S. “Grain for Green” driven land use change and carbon sequestration on the Loess Plateau, China. Sci. Rep. 2014, 4, 7039. [Google Scholar] [CrossRef] [PubMed]
- Lü, Y.H.; Fu, B.J.; Feng, X.M.; Zeng, Y.; Liu, Y.; Chang, R.Y.; Sun, G.; Wu, B.F. A policy-driven large scale ecological restoration: Quantifying ecosystem services changes in the Loess Plateau of China. PLoS ONE 2012, 7, e31782. [Google Scholar] [CrossRef] [PubMed]
- Lu, Q.L.; Tang, L.R.; Wang, S.Q.; Huang, B.; Chen, Y.D.; Chen, X.R. An investigation on the characteristics of cellulose nanocrystals from Pennisetum sinese. Biomass Bioenergy 2014, 70, 267–272. [Google Scholar] [CrossRef]
- Xu, Q.L.; Hunag, Z.J.; Wang, X.M.; Cui, L.H. Pennisetum sinese Roxb and Pennisetum purpureum Schum. as vertical-flow constructed wetland vegetation for removal of N and P from domestic sewage. Ecol. Eng. 2015, 83, 120–124. [Google Scholar] [CrossRef]
- Hu, L.; Wang, R.; Liu, X.; Xu, B.; Xie, T.; Li, Y.; Wang, M.; Wang, G.; Chen, Y. Cadmium phytoextraction potential of king grass (pennisetum sinese roxb.) and responses of rhizosphere bacterial communities to a cadmium pollution gradient. Environ. Sci. Pollut. Res. 2018, 25, 21671–21681. [Google Scholar] [CrossRef] [PubMed]
- Hardoim, P.R.; Van Overbeek, L.S.; Berg, G.; Pirttilä, A.M.; Compant, S.; Campisano, A.; Döring, M.; Sessitsch, A. The hidden world within plants: Ecological and evolutionary considerations for defining functioning of microbial endophytes. Microbiol. Mol. Biol. Rev. 2015, 79, 293–320. [Google Scholar] [CrossRef] [PubMed]
- Banach, A.; Kuźniar, A.; Mencfel, R.; Wolińska, A. The Study on the Cultivable Microbiome of the Aquatic Fern Azolla Filiculoides L. as New Source of Beneficial Microorganisms. Appl. Sci. 2019, 9, 2143. [Google Scholar] [CrossRef]
- Ryan, R.P.; Germaine, K.; Franks, A.; Ryan, D.J.; Dowling, D.N. Bacterial endophytes: Recent developments and applications. FEMS Microbiol. Lett. 2008, 278, 1–9. [Google Scholar] [CrossRef]
- Taghavi, S.; Garafola, C.; Monchy, S.; Newman, L.; Hoffman, A.; Weyens, N.; Barac, T.; Vangronsveld, J.; van der Lelie, D. Genome survey and characterization of endophytic bacteria exhibiting a beneficial effect on growth and development of poplar trees. Appl. Environ. Microbiol. 2009, 75, 748–757. [Google Scholar] [CrossRef] [PubMed]
- Loaces, I.; Ferrando, L.; Scavino, A.F. Dynamics, diversity and function of endophytic siderophore-producing bacteria in rice. Microb. Ecol. 2011, 61, 606–618. [Google Scholar] [CrossRef]
- Deng, Z.S.; Zhang, B.C.; Qi, X.Y.; Sun, Z.H.; He, X.L.; Liu, Y.Z.; Li, J.; Chen, K.K.; Lin, Z.X. Root-Associated Endophytic Bacterial Community Composition of Pennisetum sinese from Four Representative Provinces in China. Microorganisms 2019, 7, 47. [Google Scholar] [CrossRef] [PubMed]
- Lin, B.S.; Song, Z.Z.; Zhang, L.L.; Fan, J.L.; Lin, Z.X. Composition diversity and differences of endophytic bacteria in root, stem and leaf at different growth stages of Pennisetum sp. J. Fujian Agric. For. Univ. 2018, 3, 2885–2893. [Google Scholar]
- USDA. Soil Survey Laboratory Methods Manual; Version 3.0; Soil Survey Investigations Report No. 42; United States Department of Agriculture, Natural Resources Conservation Service, National Soil Survey Centre: Lincoln, NE, USA, 1996.
- Magoč, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef] [PubMed]
- Bokulich, N.A.; Subramanian, S.; Faith, J.J.; Gevers, D.; Gordon, J.I.; Knight, R.; Mills, D.A.; Caporaso, J.G. Quality-filtering vastly improves diversity estimates from Illumina amplicon sequencing. Nat. Methods 2013, 10, 57. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Pena, A.G.; Goodrich, J.K.; Gordon, J.I. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996. [Google Scholar] [CrossRef]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013. [Google Scholar]
- da Rocha, P.; Paula, V.M.B.; Estevinho, L.M.F.; dos Santos, E.L.; de Picoli Souza, K. Antioxidant and antibacterial activity of endophytic fungi extracts from Schinus teribinthifolius Raddi. Free Radic. Biol. Med. 2018, 128, S116. [Google Scholar] [CrossRef]
- Sabra, M.; Aboulnasr, A.; Franken, P.; Perreca, E.; Wright, L.P.; Camehl, I. Beneficial Root Endophytic Fungi Increase Growth and Quality Parameters of Sweet Basil in Heavy Metal Contaminated Soil. Front. Plant. Sci. 2018, 9, 1726. [Google Scholar] [CrossRef]
- Christian, N.; Herre, E.A.; Clay, K. Foliar endophytic fungi alter patterns of nitrogen uptake and distribution in Theobroma cacao. New Phytol. 2019, 222, 1573–1583. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, R.; White Jr, J.; Arnold, A.E.; Redman, a.R.a. Fungal endophytes: Diversity and functional roles. New Phytol. 2009, 182, 314–330. [Google Scholar] [CrossRef] [PubMed]
- Toju, H.; Yamamoto, S.; Sato, H.; Tanabe, A.S.; Gilbert, G.S.; Kadowaki, K. Community composition of root-associated fungi in a Quercus-dominated temperate forest:“codominance” of mycorrhizal and root-endophytic fungi. Ecol. Evol. 2013, 3, 1281–1293. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Duan, Y.; Yang, Y. Effects of long-term fertilization on flue-cured tobacco soil nutrients and microorganisms community structure. Sci. Agric. Sin. 2014, 47, 3424–3433. [Google Scholar]
- Shen, Z.Z.; Ruan, Y.Z.; Chao, X.; Zhang, J.; Li, R.; Shen, Q.R. Rhizosphere microbial community manipulated by 2 years of consecutive biofertilizer application associated with banana Fusarium wilt disease suppression. Biol. Fertil. Soils 2015, 51, 553–562. [Google Scholar] [CrossRef]
- Maia, N.d.C.; Souza, P.N.d.C.; Godinho, B.T.V.; Moreira, S.I.; Abreu, L.M.d.; Jank, L.; Cardoso, P.G. Fungal endophytes of Panicum maximum and Pennisetum purpureum: Isolation, identification, and determination of antifungal potential. Revista Brasileira de Zootecnia 2018, 47. [Google Scholar] [CrossRef]
- Jiang, Y. Study on the classification of soil melanospora in subtropical and western sichuan plateau. Shandong Agric. Univ. 2007. [Google Scholar]
- Diouf, D.; Samba-Mbaye, R.; Lesueur, D.; Ba, A.T.; Dreyfus, B.; De Lajudie, P.; Neyra, M. Genetic diversity of Acacia seyal Del. rhizobial populations indigenous to Senegalese soils in relation to salinity and pH of the sampling sites. Microb. Ecol. 2007, 54, 553–566. [Google Scholar] [CrossRef]
- Rana, K.L.; Kour, D.; Sheikh, I.; Yadav, N.; Yadav, A.N.; Kumar, V.; Singh, B.P.; Dhaliwal, H.S.; Saxena, A.K. Biodiversity of endophytic fungi from diverse niches and their biotechnological applications. In Advances in Endophytic Fungal Research; Springer: Berlin, Germany, 2019; pp. 105–144. [Google Scholar]
- Gomes, T.; Pereira, J.A.; Benhadi, J.; Lino-Neto, T.; Baptista, P. Endophytic and epiphytic phyllosphere fungal communities are shaped by different environmental factors in a Mediterranean ecosystem. Microb. Ecol. 2018, 76, 668–679. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sampling Sites | SX | FJ | XJ | NS | NY |
|---|---|---|---|---|---|
| Yanan, Shaanxi Province | Fuzhou, Fujian Province | Changji, Xinjiang Uygur Autonomous Prefecture | Bayannaoer, Inner Mongolia (Sandy Land) | Bayannaoer, Inner Mongolia (Saline Alkali Land) | |
| Total carbon (%) | 2.47 | 4 | 1.44 | 0.77 | 0.23 |
| Total nitrogen (%) | 0.62 | 0.53 | 0.24 | 0.12 | 0.04 |
| Total phosphorus (mg/kg) | 1784 | 2065 | 2036 | 1465 | 1718 |
| Effective phosphorus (mg/kg) | 12.6 | 10 | 10.8 | 3 | 3.5 |
| Quick acting potassium (mg/kg) | 132.9 | 112.8 | 324.7 | 154.1 | 99.5 |
| pH | 8.35 | 6.98 | 8.27 | 8.9 | 9.38 |
| Sample Name | Raw Tags | Clean Tags | Effective Tags | Base (nt) | Average Length (nt) | Q20 | GC% | Effective% |
|---|---|---|---|---|---|---|---|---|
| SX | 79,608 | 77,989 | 77,902 | 20,915,412 | 268 | 99.27 | 56.08 | 97.02 |
| FJ | 79,018 | 78,476 | 78,175 | 16,975,477 | 217 | 99.38 | 49.92 | 97.33 |
| XJ | 52,987 | 52,439 | 52,308 | 12,849,443 | 246 | 99.18 | 45.34 | 97.65 |
| NS | 94,083 | 90,362 | 90,280 | 23,771,714 | 263 | 99.34 | 55.38 | 95.27 |
| NY | 79,742 | 76,343 | 76,210 | 18,856,088 | 247 | 99.36 | 51.65 | 94.37 |
| Sample Name | Observed_Species | Shannon | Simpson | Chao1 | ACE | PD_Whole_Tree |
|---|---|---|---|---|---|---|
| SX | 109 | 0.56 | 0.12 | 130.37 | 136.71 | 29.92 |
| FJ | 223 | 2.12 | 0.60 | 237.88 | 246.86 | 57.80 |
| XJ | 135 | 2.205 | 0.71 | 238.50 | 179.16 | 32.84 |
| NS | 79 | 1.19 | 0.30 | 83.58 | 89.13 | 15.89 |
| NY | 106 | 2.43 | 0.69 | 110.13 | 113.46 | 21.33 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deng, Z.-S.; Liu, X.-D.; Zhang, B.-C.; Jiao, S.; Qi, X.-Y.; Sun, Z.-H.; He, X.-L.; Liu, Y.-Z.; Li, J.; Chen, K.-K.; et al. The Root Endophytic Fungi Community Structure of Pennisetum sinese from Four Representative Provinces in China. Microorganisms 2019, 7, 332. https://doi.org/10.3390/microorganisms7090332
Deng Z-S, Liu X-D, Zhang B-C, Jiao S, Qi X-Y, Sun Z-H, He X-L, Liu Y-Z, Li J, Chen K-K, et al. The Root Endophytic Fungi Community Structure of Pennisetum sinese from Four Representative Provinces in China. Microorganisms. 2019; 7(9):332. https://doi.org/10.3390/microorganisms7090332
Chicago/Turabian StyleDeng, Zhen-Shan, Xiao-Dong Liu, Bao-Cheng Zhang, Shuo Jiao, Xiang-Ying Qi, Zhi-Hong Sun, Xiao-Long He, Yu-Zhen Liu, Jing Li, Kai-Kai Chen, and et al. 2019. "The Root Endophytic Fungi Community Structure of Pennisetum sinese from Four Representative Provinces in China" Microorganisms 7, no. 9: 332. https://doi.org/10.3390/microorganisms7090332
APA StyleDeng, Z.-S., Liu, X.-D., Zhang, B.-C., Jiao, S., Qi, X.-Y., Sun, Z.-H., He, X.-L., Liu, Y.-Z., Li, J., Chen, K.-K., Lin, Z.-X., & Jiang, Y.-Y. (2019). The Root Endophytic Fungi Community Structure of Pennisetum sinese from Four Representative Provinces in China. Microorganisms, 7(9), 332. https://doi.org/10.3390/microorganisms7090332