Frequency of Diarrheagenic Virulence Genes and Characteristics in Escherichia coli Isolates from Pigs with Diarrhea in China

Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Isolates
2.2. Detection of Virulence Genes Using Real-time PCR
2.3. Strain Selection Strategy for Genotypic and Antimicrobial Analysis
2.4. Multilocus Sequence Typing
2.5. Pulsed-Field Gel Electrophoresis
2.6. Antimicrobial Susceptibility Testing
2.7. Hemolytic Activity Determination
2.8. Adhesion Assay
2.9. Cell Death Assay
2.10. Statistical Analysis
3. Results
3.1. Prevalence of Virulence Genes in E. coli Isolates from Pigs with Diarrhea
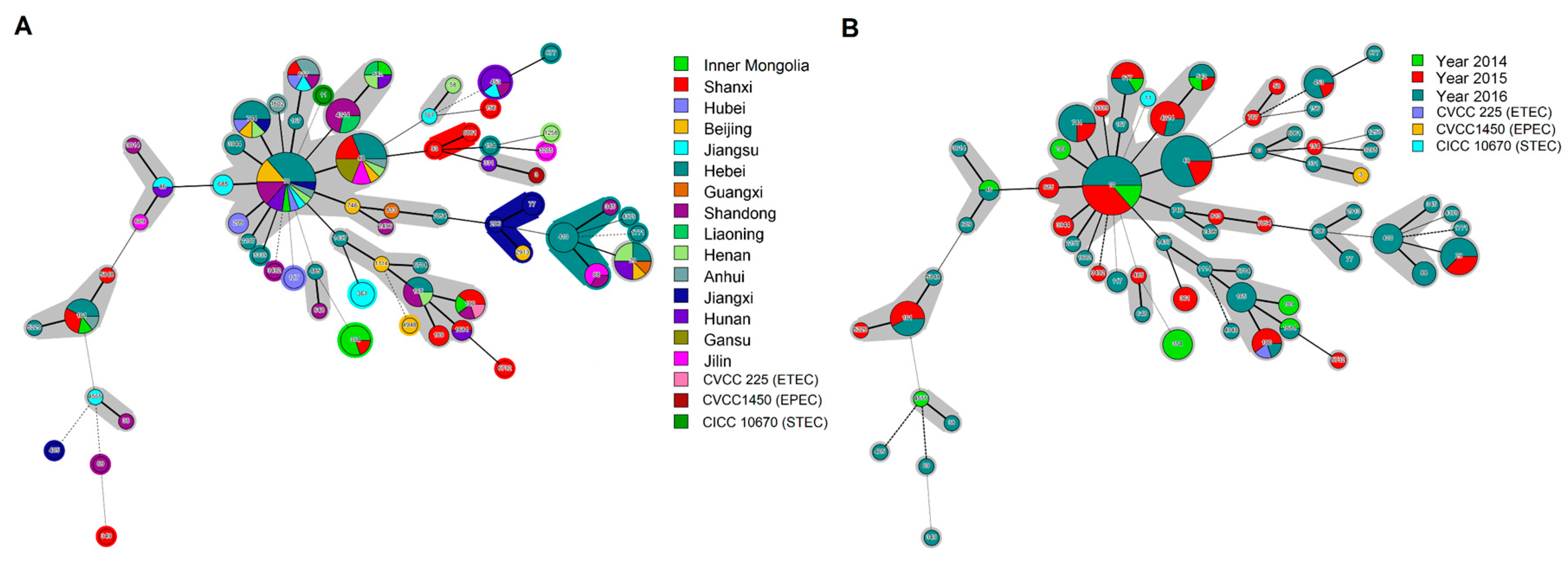
3.2. Phylogenetic Analysis of Diarrheagenic E. coli Using MLST
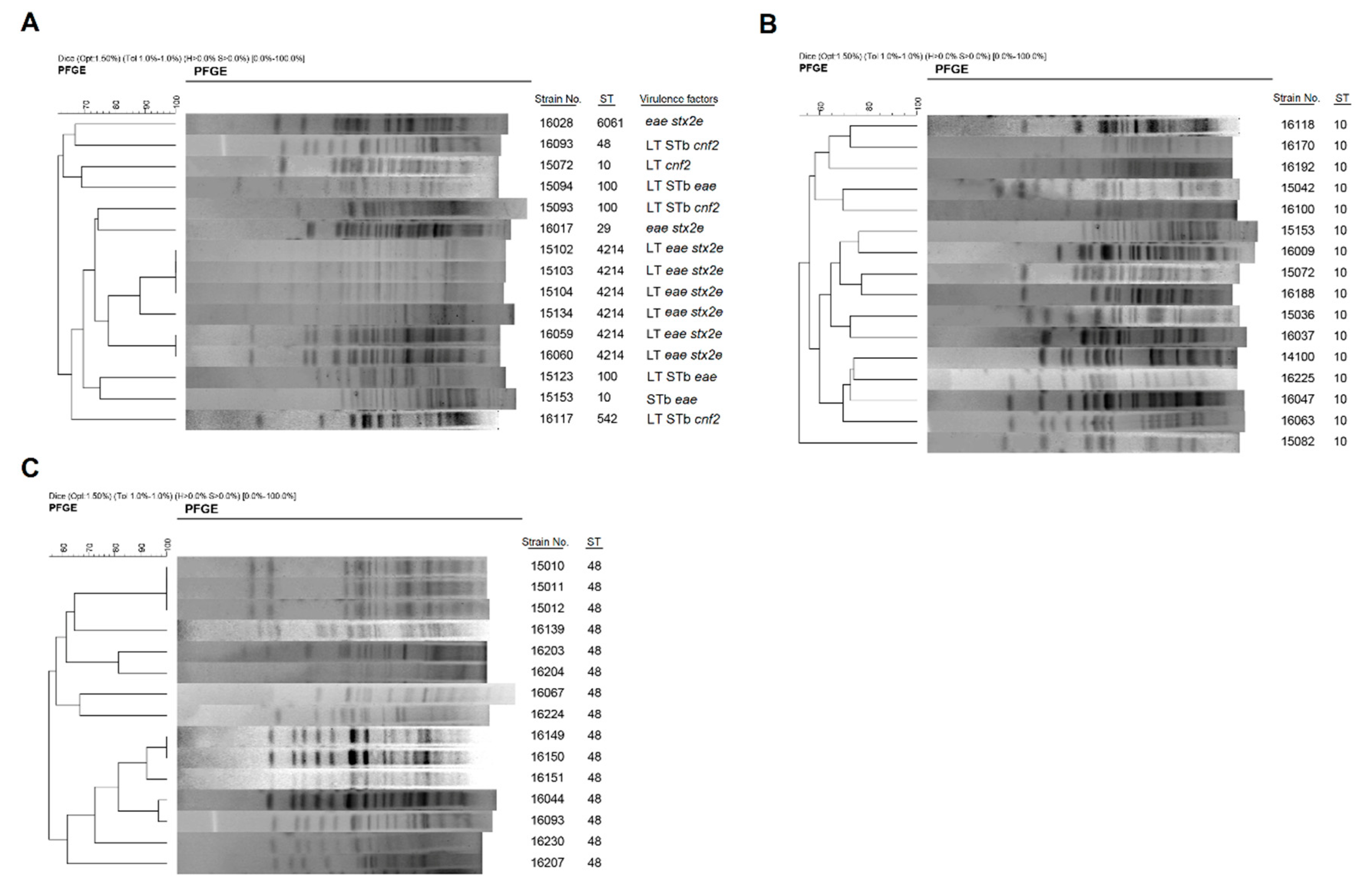
3.3. Phylogenetic Analysis of Diarrheagenic E. coli Using PFGE
3.4. Antimicrobial Susceptibility
3.5. Adherence and Cytotoxicity of Intestinal Pathogenic E. coli Hybrids to Intestinal Epithelial cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Fairbrother, J.M.; Nadeau, E.; Gyles, C.L. Escherichia coli in postweaning diarrhea in pigs: An update on bacterial types, pathogenesis, and prevention strategies. Anim. Health Res. Rev. 2005, 6, 17–39. [Google Scholar] [CrossRef] [PubMed]
- Croxen, M.A.; Law, R.J.; Scholz, R.; Keeney, K.M.; Wlodarska, M.; Finlay, B.B. Recent advances in understanding enteric pathogenic Escherichia coli. Clin. Microbiol. Rev. 2013, 26, 822–880. [Google Scholar] [CrossRef]
- Clermont, O.; Olier, M.; Hoede, C.; Diancourt, L.; Brisse, S.; Keroudean, M.; Glodt, J.; Picard, B.; Oswald, E.; Denamur, E. Animal and human pathogenic Escherichia coli strains share common genetic backgrounds. Infect. Genet. Evol. 2011, 11, 654–662. [Google Scholar] [CrossRef] [PubMed]
- Frydendahl, K. Prevalence of serogroups and virulence genes in Escherichia coli associated with postweaning diarrhoea and edema disease in pigs and a comparison of diagnostic approaches. Vet. Microbiol. 2002, 85, 169–182. [Google Scholar] [CrossRef]
- Luppi, A. Swine enteric colibacillosis: Diagnosis, therapy and antimicrobial resistance. Porcine Health Manag. 2017, 3, 16. [Google Scholar] [CrossRef]
- Tseng, M.; Fratamico, P.M.; Bagi, L.; Delannoy, S.; Fach, P.; Manning, S.D.; Funk, J.A. Diverse virulence gene content of Shiga toxin-producing Escherichia coli from finishing swine. Appl. Environ. Microbiol. 2014, 80, 6395–6402. [Google Scholar] [CrossRef]
- Nyholm, O.; Heinikainen, S.; Pelkonen, S.; Hallanvuo, S.; Haukka, K.; Siitonen, A. Hybrids of Shigatoxigenic and Enterotoxigenic Escherichia coli (STEC/ETEC) Among Human and Animal Isolates in Finland. Zoonoses Public Health 2015, 62, 518–524. [Google Scholar] [CrossRef]
- Zhou, D.; Zhu, Y.H.; Zhang, W.; Wang, M.L.; Fan, W.Y.; Song, D.; Yang, G.Y.; Jensen, B.B.; Wang, J.F. Oral administration of a select mixture of Bacillus probiotics generates Tr1 cells in weaned F4ab/acR− pigs challenged with an F4+ ETEC/VTEC/EPEC strain. Vet. Res. 2015, 46, 95. [Google Scholar] [CrossRef]
- Chandran, A.; Mazumder, A. Prevalence of diarrhea-associated virulence genes and genetic diversity in Escherichia coli isolates from fecal material of various animal hosts. Appl. Environ. Microbiol. 2013, 79, 7371–7380. [Google Scholar] [CrossRef]
- Chandran, A.; Mazumder, A. Occurrence of diarrheagenic virulence genes and genetic diversity in Escherichia coli isolates from fecal material of various avian hosts in British Columbia, Canada. Appl. Environ. Microbiol. 2014, 80, 1933–1940. [Google Scholar] [CrossRef]
- Gresse, R.; Chaucheyras-Durand, F.; Fleury, M.A.; Van de Wiele, T.; Forano, E.; Blanquet-Diot, S. Gut microbiota dysbiosis in postweaning piglets: Understanding the keys to health. Trends Microbiol. 2017, 25, 851–873. [Google Scholar] [CrossRef] [PubMed]
- Enne, V.I.; Cassar, C.; Sprigings, K.; Woodward, M.J.; Bennett, P.M. A high prevalence of antimicrobial resistant Escherichia coli isolated from pigs and a low prevalence of antimicrobial resistant E. coli from cattle and sheep in Great Britain at slaughter. FEMS Microbiol. Lett. 2008, 278, 193–199. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Griffiths, M. PCR differentiation of Escherichia coli from other Gram-negative bacteria using primers derived from the nucleotide sequences flanking the gene encoding the universal stress protein. Lett. Appl. Microbiol. 1998, 27, 369–371. [Google Scholar] [CrossRef] [PubMed]
- Holt, J.G.; Krieg, N.R.; Sneath, P.H.A.; Staley, J.T.; Williams, S.T. Bergey’s Manual of Determinative Bacteriology, 9th ed.; Williams & Wilkins: Hagerstown, MD, USA, 1994. [Google Scholar]
- Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing; Twenty-sixth informational supplement. CLSI document, approved standard M100-S26; Clinical and laboratory standards institute: Wayne, PA, USA, 2016. [Google Scholar]
- Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Disk and Dilution Susceptibility Tests for Bacteria Isolated from Animals, 4th ed.; Clinical and laboratory standards institute: Wayne, PA, USA, 2013. [Google Scholar]
- Su, J.H.; Zhu, Y.H.; Ren, T.Y.; Guo, L.; Yang, G.Y.; Jiao, L.G.; Wang, J.F. Distribution and antimicrobial resistance of Salmonella isolated from pigs with diarrhea in China. Microorganisms 2018, 6, 117. [Google Scholar] [CrossRef] [PubMed]
- Dutta, S.; Pazhani, G.P.; Nataro, J.P.; Ramamurthy, T. Heterogenic virulence in a diarrheagenic Escherichia coli: Evidence for an EPEC expressing heat-labile toxin of ETEC. Int. J. Med. Microbiol. 2015, 305, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Qadri, F.; Svennerholm, A.M.; Faruque, A.S.; Sack, R.B. Enterotoxigenic Escherichia coli in developing countries: Epidemiology, microbiology, clinical features, treatment, and prevention. Clin. Microbiol. Rev. 2005, 18, 465–483. [Google Scholar] [CrossRef] [PubMed]
- Erume, J.; Berberov, E.M.; Kachman, S.D.; Scott, M.A.; Zhou, Y.; Francis, D.H.; Moxley, R.A. Comparison of the contributions of heat-labile enterotoxin and heat-stable enterotoxin b to the virulence of enterotoxigenic Escherichia coli in F4ac receptor-positive young pigs. Infect. Immun. 2008, 76, 3141–3149. [Google Scholar] [CrossRef]
- Rao, M.R.; Abu-Elyazeed, R.; Savarino, S.J.; Naficy, A.B.; Wierzba, T.F.; Abdel-Messih, I.; Shaheen, H.; Frenck, R.W., Jr.; Svennerholm, A.M.; Clemens, J.D. High disease burden of diarrhea due to enterotoxigenic Escherichia coli among rural Egyptian infants and young children. J. Clin. Microbiol. 2003, 41, 4862–4864. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Yuan, C.; Meng, X.; Du, Y.; Gao, R.; Tang, J.; Shi, D. Frequency of virulence factors in Escherichia coli isolated from suckling pigs with diarrhoea in China. Vet. J. 2014, 199, 286–289. [Google Scholar] [CrossRef]
- Chen, X.; Gao, S.; Jiao, X.; Liu, X.F. Prevalence of serogroups and virulence factors of Escherichia coli strains isolated from pigs with postweaning diarrhoea in eastern China. Vet. Microbiol. 2004, 103, 13–20. [Google Scholar] [CrossRef]
- Post, K.W.; Bosworth, B.T.; Knoth, J.L. Frequency of virulence factors in Escherichia coli isolated from pigs with postweaning diarrhea and edema disease in North Carolina. Swine Health Prod. 2000, 8, 119–120. [Google Scholar]
- Osek, J. Prevalence of virulence factors of Escherichia coli strains isolated from diarrheic and healthy piglets after weaning. Vet. Microbiol. 1999, 68, 209–217. [Google Scholar] [CrossRef]
- Cepeda-Molero, M.; Berger, C.N.; Walsham, A.D.S.; Ellis, S.J.; Wemyss-Holden, S.; Schuller, S.; Frankel, G.; Fernandez, L.A. Attaching and effacing (A/E) lesion formation by enteropathogenic E. coli on human intestinal mucosa is dependent on non-LEE effectors. PLoS Pathog. 2017, 13, e1006706. [Google Scholar] [CrossRef] [PubMed]
- Fratamico, P.M.; Bagi, L.K.; Bush, E.J.; Solow, B.T. Prevalence and characterization of shiga toxin-producing Escherichia coli in swine feces recovered in the National Animal Health Monitoring System’s Swine 2000 study. Appl. Environ. Microbiol. 2004, 70, 7173–7178. [Google Scholar] [CrossRef] [PubMed]
- Cheng, D.; Sun, H.; Xu, J.; Gao, S. PCR detection of virulence factor genes in Escherichia coli isolates from weaned piglets with edema disease and/or diarrhea in China. Vet. Microbiol. 2006, 115, 320–328. [Google Scholar] [CrossRef]
- Schuller, S.; Frankel, G.; Phillips, A.D. Interaction of Shiga toxin from Escherichia coli with human intestinal epithelial cell lines and explants: Stx2 induces epithelial damage in organ culture. Cell Microbiol. 2004, 6, 289–301. [Google Scholar] [CrossRef] [PubMed]
- Sonntag, A.K.; Bielaszewska, M.; Mellmann, A.; Dierksen, N.; Schierack, P.; Wieler, L.H.; Schmidt, M.A.; Karch, H. Shiga toxin 2e-producing Escherichia coli isolates from humans and pigs differ in their virulence profiles and interactions with intestinal epithelial cells. Appl. Environ. Microbiol. 2005, 71, 8855–8863. [Google Scholar] [CrossRef] [PubMed]
- Kaper, J.B.; Nataro, J.P.; Mobley, H.L. Pathogenic Escherichia coli. Nat. Rev. Microbiol. 2004, 2, 123–140. [Google Scholar] [CrossRef] [PubMed]
- Gomes, T.A.; Elias, W.P.; Scaletsky, I.C.; Guth, B.E.; Rodrigues, J.F.; Piazza, R.M.; Ferreira, L.C.; Martinez, M.B. Diarrheagenic Escherichia coli. Braz. J. Microbiol. 2016, 47, 3–30. [Google Scholar] [CrossRef]
- Nyholm, O.; Halkilahti, J.; Wiklund, G.; Okeke, U.; Paulin, L.; Auvinen, P.; Haukka, K.; Siitonen, A. Comparative genomics and characterization of hybrid shigatoxigenic and enterotoxigenic Escherichia coli (STEC/ETEC) strains. PLoS ONE 2015, 10, e0135936. [Google Scholar] [CrossRef]
- Martinez, J.L.; Baquero, F. Interactions among strategies associated with bacterial infection: Pathogenicity, epidemicity, and antibiotic resistance. Clin. Microbiol. Rev. 2002, 15, 647–679. [Google Scholar] [CrossRef] [PubMed]
- Roderova, M.; Halova, D.; Papousek, I.; Dolejska, M.; Masarikova, M.; Hanulik, V.; Pudova, V.; Broz, P.; Htoutou-Sedlakova, M.; Sauer, P.; et al. Characteristics of quinolone resistance in Escherichia coli isolates from humans, animals, and the environment in the Czech Republic. Front. Microbiol. 2016, 7, 2147. [Google Scholar] [CrossRef]
- Wang, Y.; Tian, G.B.; Zhang, R.; Shen, Y.; Tyrrell, J.M.; Huang, X.; Zhou, H.; Lei, L.; Li, H.Y.; Doi, Y.; et al. Prevalence, risk factors, outcomes, and molecular epidemiology of mcr-1-positive Enterobacteriaceae in patients and healthy adults from China: An epidemiological and clinical study. Lancet Infect. Dis. 2017, 17, 390–399. [Google Scholar] [CrossRef]
- Xu, Y.; Bai, X.; Jin, Y.; Hu, B.; Wang, H.; Sun, H.; Fan, R.; Fu, S.; Xiong, Y. High prevalence of virulence genes in specific genotypes of atypical enteropathogenic Escherichia coli. Front. Cell Infect. Microbiol. 2017, 7, 109. [Google Scholar] [CrossRef] [PubMed]
- Kusumoto, M.; Hikoda, Y.; Fujii, Y.; Murata, M.; Miyoshi, H.; Ogura, Y.; Gotoh, Y.; Iwata, T.; Hayashi, T.; Akiba, M. Emergence of a multidrug-resistant Shiga Toxin-producing enterotoxigenic Escherichia coli lineage in diseased swine in Japan. J. Clin. Microbiol. 2016, 54, 1074–1081. [Google Scholar] [CrossRef] [PubMed]
- Cespedes, S.; Saitz, W.; Del Canto, F.; De la Fuente, M.; Quera, R.; Hermoso, M.; Munoz, R.; Ginard, D.; Khorrami, S.; Giron, J.; et al. Genetic diversity and virulence determinants of Escherichia coli strains isolated from patients with Crohn’s disease in Spain and Chile. Front. Microbiol. 2017, 8, 639. [Google Scholar] [CrossRef]
- Herrero-Fresno, A.; Ahmed, S.; Hansen, M.H.; Denwood, M.; Zachariasen, C.; Olsen, J.E. Genotype variation and genetic relationship among Escherichia coli from nursery pigs located in different pens in the same farm. BMC Microbiol. 2017, 17, 5. [Google Scholar] [CrossRef] [PubMed]
- Meng, Q.; Bai, X.; Zhao, A.; Lan, R.; Du, H.; Wang, T.; Shi, C.; Yuan, X.; Bai, X.; Ji, S.; et al. Characterization of Shiga toxin-producing Escherichia coli isolated from healthy pigs in China. BMC Microbiol. 2014, 14, 5. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.G.; Johnson, T.A.; Su, J.Q.; Qiao, M.; Guo, G.X.; Stedtfeld, R.D.; Hashsham, S.A.; Tiedje, J.M. Diverse and abundant antibiotic resistance genes in Chinese swine farms. Proc. Natl. Acad. Sci. USA 2013, 110, 3435–3440. [Google Scholar] [CrossRef]
- Fijan, S.; Sulc, D.; Steyer, A. Study of the in vitro antagonistic activity of various single-strain and multi-strain probiotics against Escherichia coli. Int. J. Environ. Res. Public Health 2018, 15, 1539. [Google Scholar] [CrossRef]
- Rund, S.A.; Rohde, H.; Sonnenborn, U.; Oelschlaeger, T.A. Antagonistic effects of probiotic Nissle 1917 on EHEC strains of serotype O104:H4 and O157:H7. Int. J. Med. Microbiol. 2013, 303, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.Y.; Zhu, Y.H.; Zhang, W.; Zhou, D.; Zhai, C.C.; Wang, J.F. Influence of orally fed a select mixture of Bacillus probiotics on intestinal T-cell migration in weaned MUC4 resistant pigs following Escherichia coli challenge. Vet. Res. 2016, 47, 71. [Google Scholar] [CrossRef] [PubMed]
- Robins-Browne, R.M.; Holt, K.E.; Ingle, D.J.; Hocking, D.M.; Yang, J.; Tauschek, M. Are Escherichia coli pathotypes still relevant in the era of whole-genome sequencing? Front. Cell Infect. Microbiol. 2016, 6, 141. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sources (Number of Isolates) | Farm location (Number of Isolates) | Month | Year | Total Number of Isolates |
|---|---|---|---|---|
| Small intestine (93) | Shanxi (26), Hubei (24), Beijing (21), Liaoning (11), Hebei (11) | February to December | 2014 | 93 |
| Small intestine (139) | Hebei (37), Guangxi (21), Jilin (19), Shanxi (17), Liaoning (13), Jiangsu (13), Shandong (12), Henan (7) | January to December | 2015 | 139 |
| Small intestine (121), feces (102) | Hebei (64), Henan (37), Beijing (20), Shandong (20), Hunan (17), Jiangxi (14), Hubei (13), Shanxi (12), Jilin (11), Anhui (6), Gansu (6), Inner Mongolia (3) | March to December | 2016 | 223 |
| Gene Product a | Primer | Product Size (bp) | Accession Number | |
|---|---|---|---|---|
| Direction b | Sequence (5′→3′) | |||
| uspA | F | CCGATACGCTGCCAATCAGT | 884 | CP006636.1 |
| R | ACGCAGACCGTAGGCCAGAT | |||
| LT | F | TTCCCACCGGATCACCAA | 62 | KF733767.1 |
| R | CAACCTTGTGGTGCATGATGA | |||
| STa | F | CAACTGAATCACTTGACTCTT | 158 | CP025841.1 |
| R | TTAATAACATCCAGCACAGG | |||
| STb | F | ATGTAAATACCTACAACGGGTGAT | 360 | M35729 |
| R | TATTTGGGCGCCAAAGCATGCTCC | |||
| eae | F | CATTGATCAGGATTTTTCTGGTGATA | 102 | Z11541 |
| R | CTCATGCGGAAATAGCCGTTA | |||
| stx2 | F | CCACATCGGTGTCTGTTATTAACC | 93 | X07865 |
| R | GGTCAAAACGCGCCTGATAG | |||
| stx2e | F | ATACGATGACGCCGGAAGAC | 291 | U72191.1 |
| R | TCAGAAACGCTGCTGCTGTA | |||
| cnf2 | F | GCGGAAATTTGAGCGGTTGT | 165 | U01097.1 |
| R | CGCGCGGCATTGGATTATTT | |||
| Tir | F | GTTGGCTTTGACACCGGAAC | 379 | AF022236 |
| R | TACACCAGCACCAATTCCCC | |||
| espA | F | TCAGAATCGCAGCCTGAAAA | 60 | AF022236 |
| R | CGAAGGATGAGGTGGTTAAGCT | |||
| escV | F | ATTCTGGCTCTCTTCTTCTTTATGGCTG | 544 | AF022236 |
| R | CGTCCCCTTTTACAAACTTCATCGC | |||
| F4 | F | GAATCTGTCCGAGAATATCA | 499 | EU570252.1 |
| R | GTTGGTACAGGTCTTAATGG | |||
| F18 | F | TGGCACTGTAGGAGATACCATTCAGC | 337 | JX987521.1 |
| R | GGTTTGACCACCTTTCAGTTGAGCAG | |||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, G.-Y.; Guo, L.; Su, J.-H.; Zhu, Y.-H.; Jiao, L.-G.; Wang, J.-F. Frequency of Diarrheagenic Virulence Genes and Characteristics in Escherichia coli Isolates from Pigs with Diarrhea in China. Microorganisms 2019, 7, 308. https://doi.org/10.3390/microorganisms7090308
Yang G-Y, Guo L, Su J-H, Zhu Y-H, Jiao L-G, Wang J-F. Frequency of Diarrheagenic Virulence Genes and Characteristics in Escherichia coli Isolates from Pigs with Diarrhea in China. Microorganisms. 2019; 7(9):308. https://doi.org/10.3390/microorganisms7090308
Chicago/Turabian StyleYang, Gui-Yan, Liang Guo, Jin-Hui Su, Yao-Hong Zhu, Lian-Guo Jiao, and Jiu-Feng Wang. 2019. "Frequency of Diarrheagenic Virulence Genes and Characteristics in Escherichia coli Isolates from Pigs with Diarrhea in China" Microorganisms 7, no. 9: 308. https://doi.org/10.3390/microorganisms7090308
APA StyleYang, G.-Y., Guo, L., Su, J.-H., Zhu, Y.-H., Jiao, L.-G., & Wang, J.-F. (2019). Frequency of Diarrheagenic Virulence Genes and Characteristics in Escherichia coli Isolates from Pigs with Diarrhea in China. Microorganisms, 7(9), 308. https://doi.org/10.3390/microorganisms7090308