A Polyphasic Approach Aids Early Detection of Potentially Toxigenic Aspergilli in Soil

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Soil Samples Collection and DNA Template Preparation
2.2. Visual Assessment of Isolates and Coconut Medium Test
2.3. MALDI-ToF-MS
2.4. ITS rRNA Metabarcoding
2.5. Whole Genome Sequencing
2.6. Data Analysis
3. Results
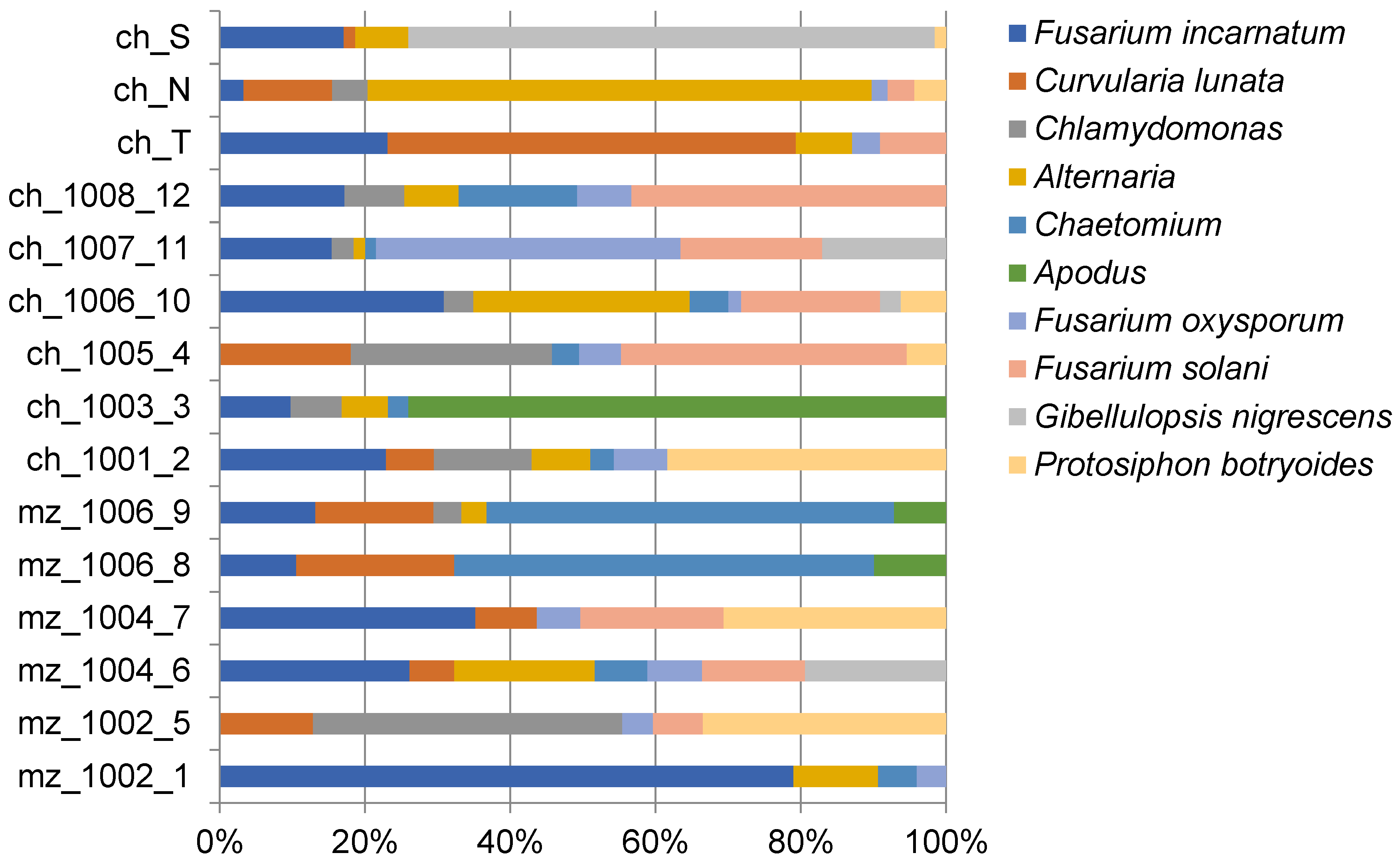
3.1. ITS rRNA Metabarcoding of Maize and Chili Soils
3.2. Trends of Aspergillus flavus (OTU62) in the Soil
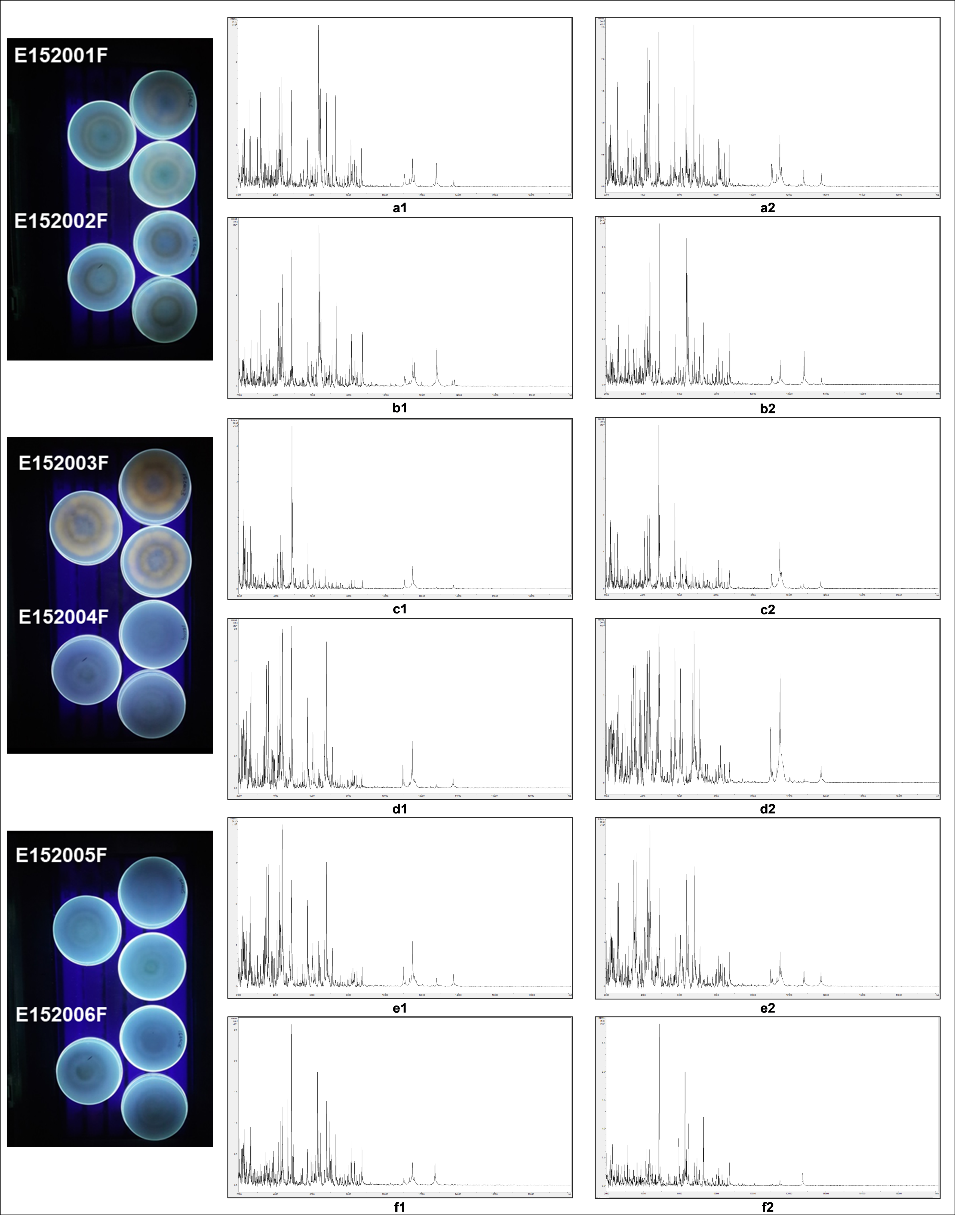
3.3. Characterisation of A. flavus Strains by Coconut Medium Test
3.4. MALDI-ToF-MS of Aspergillus flavus Strains
3.5. Whole Mitochondrial Genomes and Nuclear Ribosomal Cluster of A. flavus
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hedayati, M.T.; Pasqualotto, A.C.; Warn, P.A.; Bowyer, P.; Denning, C.W. Aspergillus flavus: Human pathogen, allergen and mycotoxin producer. Microbiology 2007, 153, 1677–1692. [Google Scholar] [CrossRef] [PubMed]
- Mori, T.; Matsumura, M.; Yamada, K.; Irie, S.; Oshimi, K.; Suda, K.; Oguri, T.; Ichinoe, M. Systemic aspergillosis caused by an aflatoxin-producing strain of Aspergillus flavus. Med. Mycol. 1998, 36, 107–112. [Google Scholar] [CrossRef] [PubMed]
- Vonberg, R.P.; Gastmeier, P. Nosocomial aspergillosis in outbreak settings. J. Hosp. Infect. 2006, 63, 246–254. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, S.; Manavathu, E.K.; Chandrasekar, P.H. Aspergillus flavus: An emerging non-fumigatus Aspergillus species of significance. Mycoses 2009, 52, 206–222. [Google Scholar] [CrossRef] [PubMed]
- Masih, A.; Singh, P.K.; Kathuria, S.; Agarwal, K.; Meis, J.F.; Chowdhary, A. Identification by Molecular Methods and Matrix-Assisted Laser Desorption Ionization–Time of Flight Mass Spectrometry and Antifungal Susceptibility Profiles of Clinically Significant Rare Aspergillus Species in a Referral Chest Hospital in Delhi, India. J. Clin. Microbiol. 2016, 54, 2354–2364. [Google Scholar] [CrossRef] [PubMed]
- Razzaghi-Abyaneh, M.; Chang, P.K.; Shams-Ghahfarokhi, M.; Rai, M. Global health issues of aflatoxins in food and agriculture: Challenges and opportunities. Front. Microbiol. 2014, 5, 420. [Google Scholar] [CrossRef] [PubMed]
- Villers, F. Aflatoxins and safe storage. Front. Microbiol. 2014, 5, 158–164. [Google Scholar] [CrossRef] [PubMed]
- Perrone, G.; Logrieco, A.F.; Gallo, A. Biodiversity of Aspergillus section Flavi in Europe in relation to the management of aflatoxin risk. Front. Microbiol. 2014, 5, 377. [Google Scholar] [CrossRef] [PubMed]
- Garber, R.K.; Cotty, P.J. Formation of Sclerotia and Aflatoxins in Developing Cotton Bolls Infected by the S Strain of Aspergillus flavus and Potential for Biocontrol with an Atoxigenic Strain. Phytopathology 1997, 87, 940–945. [Google Scholar] [CrossRef]
- Rodrigues, P.; Venâncio, A.; Kozakiewicz, Z.; Lima, N. A polyphasic approach to the identification of aflatoxigenic and non-aflatoxigenic strains of Aspergillus Section Flavi isolated from Portuguese almonds. Int. J. Food Microbiol. 2009, 129, 187–193. [Google Scholar] [CrossRef]
- Wu, F.; Stacy, S.L.; Kensler, T.W. Global Risk Assessment of Aflatoxins in Maize and Peanuts: Are Regulatory Standards Adequately Protective? Toxicol. Sci. 2013, 135, 251–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Probst, C.; Bandyopadhyay, R.; Cotty, P.J. Diversity of aflatoxin-producing fungi and their impact on food safety in sub-Saharan Africa. Int. J. Food Microbiol. 2014, 174, 113–122. [Google Scholar] [CrossRef] [PubMed]
- WHO. Food Safety Digest Aflatoxins. 2018. Available online: https://www.who.int/foodsafety/FSDigest_Aflatoxins_EN.pdf?ua=1 (accessed on 19 August 2019).
- Bowen, K.L.; Flanders, K.L.; Hagan, A.K.; Ortiz, B. Insect Damage, Aflatoxin Content, and Yield of Bt Corn in Alabama. J. Econ. Entomol. 2014, 107, 1818–1827. [Google Scholar] [CrossRef] [PubMed]
- Lewis, L.; Onsongo, M.; Njapau, H.; Schurz-Rogers, H.; Luber, G.; Kieszak, S.; Nyamongo, J.; Backer, L.; Dahiye, A.M.; Misore, A.; et al. Aflatoxin contamination of commercial maize products during an outbreak of acute aflatoxicosis in eastern and central Kenya. Environ. Health Perspect. 2005, 113, 1763–1767. [Google Scholar] [CrossRef] [PubMed]
- Dieme, R.M.A.; Faye, I.; Zoclanclounon, Y.A.B.; Fonceka, D.; Ndoye, O.; Diedhiou, P.M. Identification of Sources of Resistance for Peanut Aspergillus flavus Colonization and Aflatoxin Contamination. Int. J. Agronomy 2018. [Google Scholar] [CrossRef]
- Kumar, P.; Mahato, D.K.; Kamle, M.; Mohanta, T.K.; Kang, S.G. Aflatoxins: A Global Concern for Food Safety, Human Health and Their Management. Front. Microbiol. 2017, 7, 2170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iqbal, S.Z.; Paterson, R.R.M.; Bhatti, I.A.; Asi, M.R.; Sheikh, M.A.; Bhatti, H.N. Aflatoxin B1 in chilies from the Punjab region, Pakistan. Mycotoxin Res. 2010, 26, 205–209. [Google Scholar] [CrossRef] [Green Version]
- Udovicki, B.; Audenaert, K.; De Saeger, S.; Rajkovic, A. Overview on the mycotoxins incidence in Serbia in the period 2004–2016. Toxins 2018, 10, 279. [Google Scholar] [CrossRef]
- FAO. Crop Prospects and Food Situation. 2014. Available online: http://www.fao.org/3/a-i3899e.pdf (accessed on 27 December 2018).
- Economic Survey of Pakistan. Economic Survey of Pakistan, 2015–2016; Government of Pakistan, Finance Division Economic Adviser’s Wing: Islamabad, Pakistan, 2017.
- Cotty, P.J.; Mellon, J.E. Ecology of aflatoxin producing fungi and biocontrol of aflatoxin contamination. Mycotoxin Res. 2016, 22, 110–117. [Google Scholar] [CrossRef]
- Ehrlich, K.C. Non-aflatoxigenic Aspergillus flavus to prevent aflatoxin contamination in crops: Advantages and limitations. Front. Microbiol. 2014, 5, 50. [Google Scholar] [CrossRef]
- Yu, J. Current understanding on aflatoxin biosynthesis and future perspective in reducing aflatoxin contamination. Toxins 2012, 4, 1024–1057. [Google Scholar] [CrossRef]
- Frisvad, J.C.; Hubka, V.; Ezekiel, C.N.; Hong, S.-B.; Nováková, A.; Chen, A.J.; Arzanlou, M.; Larsen, T.O.; Sklenář, F.; Mahakarnchanakul, W.; et al. Taxonomy of Aspergillus section Flavi and their production of aflatoxins, ochratoxins and other mycotoxins. Stud. Mycol. 2019, 93, 1–63. [Google Scholar] [CrossRef]
- Tam, E.W.T.; Chen, J.H.K.; Lau, E.C.L.; Ngan, A.H.Y.; Fung, K.S.C.; Lee, K.; Lam, C.; Yuen, K.; Lau, S.K.P.; Woo, P.C.Y. Misidentification of Aspergillus nomius and Aspergillus tamarii as Aspergillus flavus: Characterization by Internal Transcribed Spacer, β-Tubulin, and Calmodulin Gene Sequencing, Metabolic Fingerprinting, and Matrix-Assisted Laser Desorption Ionization–Time of Flight Mass Spectrometry. J. Clin. Microbiol. 2014, 52, 1153–1160. [Google Scholar] [CrossRef]
- Saldan, N.C.; Almeida, R.T.R.; Avíncola, A.; Porto, C.; Galuch, M.B.; Magon, T.F.S.; Pilau, E.J.; Svidzinski, T.I.E.; Oliveira, C.C. Development of an analytical method for identification of Aspergillus flavus based on chemical markers using HPLC-MS. Food Chem. 2018, 241, 113–121. [Google Scholar] [CrossRef]
- Tran-Dinh, N.; Pitt, J.I.; Carter, D.A. Molecular genotype analysis of natural toxigenic and nontoxigenic isolates of Aspergillus flavus and A. parasiticus. Mycol. Res. 1999, 103, 1485–1490. [Google Scholar] [CrossRef]
- Montiel, D.; Dickinson, M.J.; Lee, H.A.; Dyer, P.S.; Jeenes, D.J.; Robertis, I.N.; James, S.; Fuller, L.J. Genetic differentiation of the Aspergillus section flavi complex using AFLP fingerprints. Mycol. Res. 2003, 107, 1427–1432. [Google Scholar] [CrossRef]
- Grubisha, L.C.; Cotty, P.J. Twenty-four microsatellite markers for the aflatoxin-producing fungus Aspergillus flavus. Mol. Ecol. Resour. 2009, 9, 264–267. [Google Scholar] [CrossRef]
- Baird, R.E.; Trigiano, R.N.; Windham, G.; Williams, P.; Kelley, R.; Abbas, H.K.; Moulton, J.K.; Scruggs, M.L. Comparison of aflatoxigenic and non aflatoxigenic isolates of Aspergillus flavus using DNA amplification fingerprinting techniques. Mycopathologia 2006, 161, 93–99. [Google Scholar] [CrossRef]
- Geisen, R. Multiplex polymerase chain reaction for the detection of potential aflatoxin and sterigmatocystin producing fungi. Syst. Appl. Microbiol. 1996, 19, 388–392. [Google Scholar] [CrossRef]
- Godet, M.; Munaut, F. Molecular strategy for identification in Aspergillus section Flavi. FEMS Microbiol. Lett. 2010, 304, 157–168. [Google Scholar] [CrossRef]
- Das, M.K.; Ehrlich, K.C.; Cotty, P.J. Use of pyrosequencing to quantify incidence of a specific Aspergillus flavus strain within complex fungal communities associated with commercial cotton crops. Phytopathology 2008, 98, 282–288. [Google Scholar] [CrossRef]
- Mylroie, E.; Ozkan, S.; Shivaji, R.; Windham, G.L.; Alpe, M.N.; Williams, W.P. Identification and Quantification of a Toxigenic and Non-Toxigenic Aspergillus flavus Strain in Contaminated Maize Using Quantitative Real-Time PCR. Toxins 2016, 8, 15. [Google Scholar] [CrossRef]
- Ponts, N.; Richard-Forget, F.; Zhang, H.; Barroso, G.; Zhao, C. Genome sequence of the emerging mycotoxin-producing filamentous fungus Fusarium tricinctum strain INRA104. Genome Announc. 2018, 6, e00509-18. [Google Scholar] [CrossRef]
- Orgiazzi, A.; Dunbar, M.B.; Panagos, P.; De Groot, G.A.; Lemanceau, P. Soil biodiversity and DNA barcodes: Opportunities and challenges. Soil Biol. Biochem. 2015, 80, 244–250. [Google Scholar] [CrossRef]
- Saeed, A. Types of Soil in Pakistan–soil Classification & Characteristics. Available online: https://businessfinancearticles.org/types-soil-pakistan (accessed on 24 August 2019).
- Kirk, P.M.; Cannon, P.F.; Minter, D.W.; Stalpers, J.A. (Eds.) Dictionary of the Fungi edn 10; CAB International: Wallingford, UK, 2008; p. 771. [Google Scholar]
- Fente, C.M.; Ordaz, J.J.; Vazquez, B.I.; Franco, C.M.; Cepeda, A. New additive for culture media for rapid identification of aflatoxin-producing Aspergillus strains. Appl. Environ. Microbiol. 2001, 67, 58–62. [Google Scholar] [CrossRef]
- Reeve, M.A.; Buddie, A.G.; Pollard, K.M.; Varia, S.; Seier, M.K.; Offord, L.C.; Cock, M.J.W. A highly-simplified and inexpensive MALDI-TOF mass spectrometry sample-preparation method with broad applicability to microorganisms, plants, and insects. J. Biol. Methods 2018, 5, e103. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, P.; Balint, M.; Greshake, B.; Bandow, C.; Rombke, J.; Schmitt, I. Illumina metabarcoding of a soil fungal community. Soil Biol. Biochem. 2013, 65, 128–132. [Google Scholar] [CrossRef]
- White, T.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M., Gelfand, D., Shinsky, J., White, T., Eds.; Academic Press: Cambridge, MA, USA, 1990; pp. 315–322. [Google Scholar]
- Ryan, M.J.; Kasulyte-Creasey, D.; Kermode, A.; San, S.P.; Buddie, A.G. Controlled Rate Cooling of Fungi Using a Stirling Cycle Freezer. Cryoletters 2014, 35, 63–69. [Google Scholar]
- Bengtsson-Palme, J.; Ryberg, M.; Hartmann, M.; Branco, S.; Wang, Z.; Godhe, A.; Wit, P.; Sánchez-García, M.; Ebersberger, I.; Sousa, F.; et al. Improved software detection and extraction of ITS1 and ITS2 from ribosomal ITS sequences of fungi and other eukaryotes for analysis of environmental sequencing data. Methods Ecol. Evol. 2013, 4, 914–919. [Google Scholar] [CrossRef]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A New Genome Assembly Algorithm and Its Applications to Single-Cell Sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [Green Version]
- Tillich, M.; Lehwark, P.; Pellizzer, T.; Ulbricht-Jones, E.S.; Fischer, A.; Bock, R.; Greiner, S. GeSeq–versatile and accurate annotation of organelle genomes. Nucleic Acid Res. 2017, 45, W6–W11. [Google Scholar] [CrossRef]
- Lohse, M.; Drechsel, O.; Bock, R. OrganellarGenomeDRAW (OGDRAW): A tool for the easy generation of high-quality custom graphical maps of plastid and mitochondrial genomes. Curr. Genet. 2007, 52, 267–274. [Google Scholar] [CrossRef]
- Fredricks, D.N.; Smith, C.; Meier, A. Comparison of Six DNA Extraction Methods for Recovery of Fungal DNA as Assessed by Quantitative PCR. J. Clin. Microbiol. 2005, 43, 5122–5128. [Google Scholar] [CrossRef]
- Rittenoura, W.R.; Parkb, J.; Cox-Ganserb, J.M.; Beezholda, D.H.; Green, B.J. Comparison of DNA extraction methodologies used for assessing fungal diversity via ITS sequencing. J. Environ. Monit. 2012, 14, 766–774. [Google Scholar] [CrossRef]
- Korani, W.; Chu, Y.; Holbrook, C.C.; Ozias-Akins, P. Insight into Genes Regulating Postharvest Aflatoxin Contamination of Tsetraploid Peanut from Transcriptional Profiling. Genetics 2018, 209, 143–156. [Google Scholar] [CrossRef]
- Peterson, S.W. Phylogenetic analysis of Aspergillus species using DNA sequences from four loci. Mycologia 2008, 100, 205–226. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reference | Soil Type | Location | Date of Collection | Geospatial Coordinates |
|---|---|---|---|---|
| mz_1002_1 | Silt-loam | Okara | 29/12/2017 | 30.801380° N, 73.448334° E |
| mz_1002_5 | Silt-loam | Okara | 29/12/2017 | 30.801380° N, 73.448334° E |
| mz_1004_6 | Silt-loam | Kasur | 29/12/2017 | 31.11866° N, 74.4502487° E |
| mz_1004_7 | Silt-loam | Kasur | 29/12/2017 | 31.11866° N, 74.4502487° E |
| mz_1006_8 | Sandy-loam | Chiniot | 30/12/2017 | 31.72° N, 72.97889° E |
| mz_1006_9 | Sandy-loam | Chiniot | 30/12/2017 | 31.72° N, 72.97889° E |
| ch_1001_2 | Silt-loam | Okara | 29/12/2017 | 30.801380° N, 73.448334° E |
| ch_1003_3 | Silt-loam | Kasur | 29/12/2017 | 31.11866° N, 74.4502487° E |
| ch_1005_4 | Sandy-loam | Chiniot | 30/12/2017 | 31.72° N, 72.97889° E |
| ch_1006_10 | Silty/clay-loam | Mirpurkhas | 14/01/2018 | 25.5251° N, 69.0159° E |
| ch_1007_11 | Loamy-soil | Tando Allah Yar | 14/01/2018 | 25.46263° N, 68.71923° E |
| ch_1008_12 | Loamy-soil | Matiari | 15/01/2018 | 25.59609° N, 68.44666° E |
| ch_T | Sandy to clay loam | Sheikhupura | 01/11/2017 | 31.7167° N, 73.9850° E |
| ch_N | Sandy to clay loam | Sheikhupura | 01/11/2017 | 31.7167° N, 73.9850° E |
| ch_S | Sandy to clay loam | Sheikhupura | 01/11/2017 | 31.7167° N, 73.9850° E |
| Sample ID | Number of Reads | Number of OTU |
|---|---|---|
| mz_1002_1 | 153,684 | 39 |
| mz_1002_5 | 173,931 | 42 |
| mz_1004_6 | 161,120 | 45 |
| mz_1004_7 | 107,104 | 33 |
| mz_1006_8 | 234,939 | 34 |
| mz_1006_9 | 267,858 | 41 |
| ch_1001_2 | 136,776 | 44 |
| ch_1003_3 | 158,472 | 33 |
| ch_1005_4 | 108,841 | 53 |
| ch_1006_10 | 141,503 | 37 |
| ch_1007_11 | 205,729 | 29 |
| ch_1008_12 | 35,556 | 47 |
| ch_T | 231,006 | 37 |
| ch_N | 208,840 | 29 |
| ch_S | 200,165 | 17 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cafà, G.; Caggiano, B.; Reeve, M.A.; Bhatti, H.; Honey, S.F.; Bajwa, B.; Buddie, A.G. A Polyphasic Approach Aids Early Detection of Potentially Toxigenic Aspergilli in Soil. Microorganisms 2019, 7, 300. https://doi.org/10.3390/microorganisms7090300
Cafà G, Caggiano B, Reeve MA, Bhatti H, Honey SF, Bajwa B, Buddie AG. A Polyphasic Approach Aids Early Detection of Potentially Toxigenic Aspergilli in Soil. Microorganisms. 2019; 7(9):300. https://doi.org/10.3390/microorganisms7090300
Chicago/Turabian StyleCafà, Giovanni, Benedetta Caggiano, Michael A. Reeve, Hamzah Bhatti, Sabyan F. Honey, Babar Bajwa, and Alan G. Buddie. 2019. "A Polyphasic Approach Aids Early Detection of Potentially Toxigenic Aspergilli in Soil" Microorganisms 7, no. 9: 300. https://doi.org/10.3390/microorganisms7090300