Pseudomonas aeruginosa Keratitis: Protease IV and PASP as Corneal Virulence Mediators


Abstract
:1. Introduction: Pseudomonas aeruginosa Keratitis and the Role of the Secreted Proteases
2. Physical, Enzymatic, and Virulence Properties of Protease IV
2.1. Protease Discovery and Gene Identification
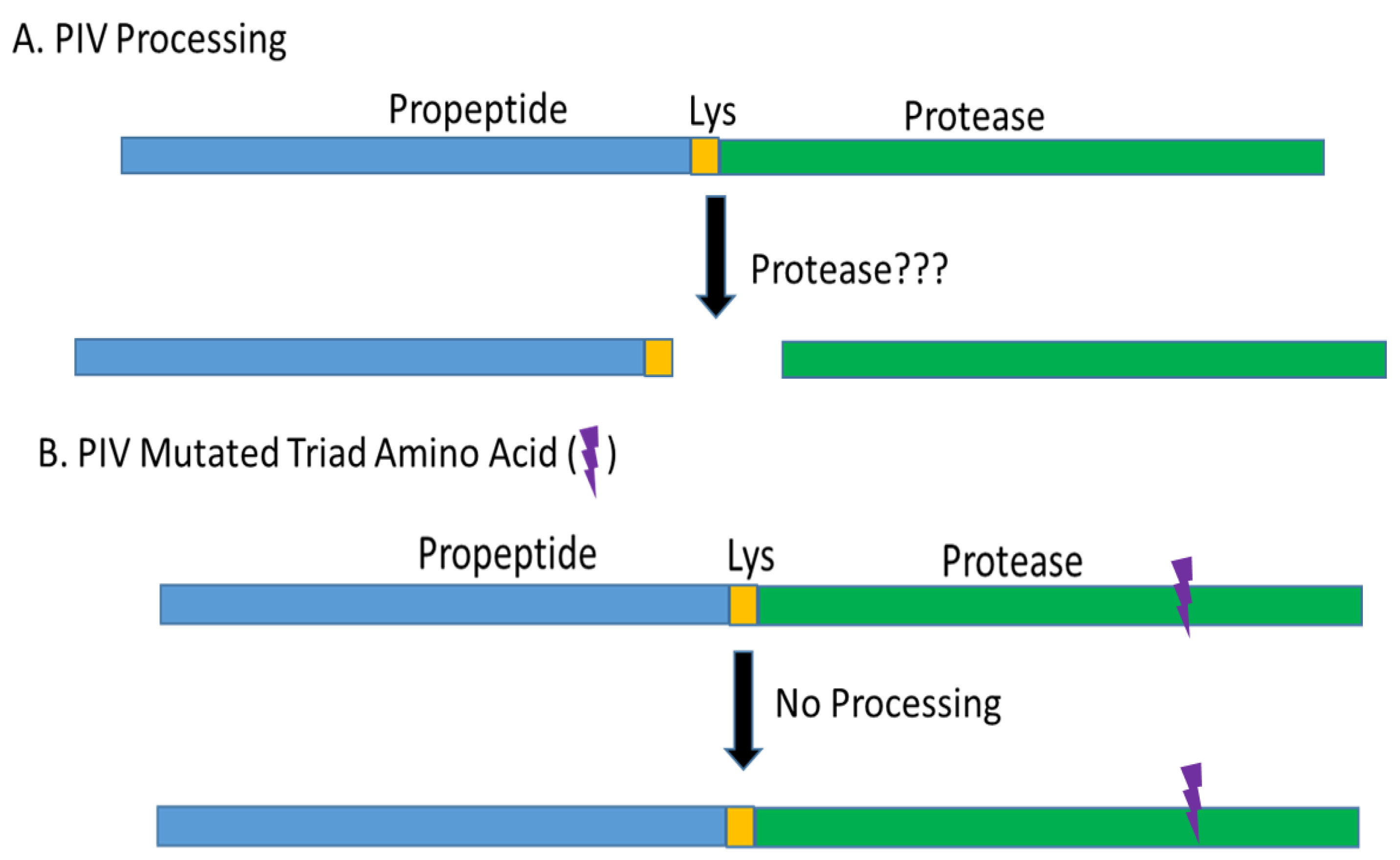
2.2. PIV Protein Processing
2.3. Enzymatic Properties of PIV
2.4. Regulation of Piv Expression
3. Virulence of PIV
3.1. Biological Activity of PIV
3.2. Infection of Plants
3.3. Infection of Insects
3.4. Infection of Mammalian Corneas
3.5. Protease IV in the Respiratory Tract
3.5.1. Surfactant Proteins
3.5.2. Rat Lung Infection
3.5.3. Human Cytokine IL-22
3.5.4. Augmenting Pneumococcal Lung and Systemic Infection
4. PASP: A Second Ocular Virulence Factor
5. Conclusions
Funding
Conflicts of Interest
References
- Vincent, J.-L. Nosocomial infections in adult intensive-care units. Lancet 2003, 361, 2068–2077. [Google Scholar] [CrossRef]
- Smolin, G.; Okumoto, M.; Wilson, F.M. The effect of tobramycin on gentamicin-resistant strains in Pseudomonas keratitis. Am. J. Ophthalmol. 1974, 77, 583–588. [Google Scholar] [CrossRef]
- Konda, N.; Motukupally, S.R.; Garg, P.; Sharma, S.; Ali, M.H.; Willcox, M.D.P. Microbial analyses of contact lens-associated microbial keratitis. Optom. Vis. Sci. 2014, 91, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Marquart, M.E.; O’Callaghan, R.J. Infectious keratitis: Secreted bacterial proteins that mediate corneal damage. J. Ophthalmol. 2013, 2013, 369094. [Google Scholar] [CrossRef] [PubMed]
- O’Callaghan, R.J. Role of exoproteins in bacterial keratitis: The Fourth Annual Thygeson Lecture, presented at the Ocular Microbiology and Immunology Group Meeting, November 7, 1998. Cornea 1999, 18, 532–537. [Google Scholar] [CrossRef] [PubMed]
- Steuhl, K.P.; Döring, G.; Henni, A.; Thiel, H.J.; Botzenhart, K. Relevance of host-derived and bacterial factors in Pseudomonas aeruginosa corneal infections. Investig. Ophthalmol. Vis. Sci. 1987, 28, 1559–1568. [Google Scholar] [PubMed]
- Matsumoto, K.; Shams, N.B.; Hanninen, L.A.; Kenyon, K.R. Cleavage and activation of corneal matrix metalloproteases by Pseudomonas aeruginosa proteases. Investig. Ophthalmol. Vis. Sci. 1993, 34, 1945–1953. [Google Scholar] [PubMed]
- Kreger, A.S.; Gray, L.D. Purification of Pseudomonas aeruginosa proteases and microscopic characterization of pseudomonal protease-induced rabbit corneal damage. Infect. Immun. 1978, 19, 630–648. [Google Scholar] [PubMed]
- Fisher, E.; Allen, J.H. Corneal ulcers produced by cell-free extracts of Pseudomonas aeruginosa. Am. J. Ophthalmol. 1958, 46, 21–27. [Google Scholar] [CrossRef]
- Kawaharajo, K.; Abe, C.; Homma, J.Y.; Kawano, M.; Goto, E. Corneal ulcers caused by protease and elastase from Pseudomonas aeruginosa. Jpn. J. Exp. Med. 1974, 44, 435–442. [Google Scholar]
- Ohman, D.E.; Sadoff, J.C.; Iglewski, B.H. Toxin A-deficient mutants of Pseudomonas aeruginosa PA103: Isolation and characterization. Infect. Immun. 1980, 28, 899–908. [Google Scholar] [PubMed]
- Nicas, T.I.; Iglewski, B.H. The contribution of exoproducts to virulence of Pseudomonas aeruginosa. Can. J. Microbiol. 1985, 31, 387–392. [Google Scholar] [CrossRef] [PubMed]
- Schad, P.A.; Bever, R.A.; Nicas, T.I.; Leduc, F.; Hanne, L.F.; Iglewski, B.H. Cloning and characterization of elastase genes from Pseudomonas aeruginosa. J. Bacteriol. 1987, 169, 2691–2696. [Google Scholar] [CrossRef] [PubMed]
- Parmely, M.; Gale, A.; Clabaugh, M.; Horvat, R.; Zhou, W.W. Proteolytic inactivation of cytokines by Pseudomonas aeruginosa. Infect. Immun. 1990, 58, 3009–3014. [Google Scholar] [PubMed]
- Hong, Y.Q.; Ghebrehiwet, B. Effect of Pseudomonas aeruginosa elastase and alkaline protease on serum complement and isolated components C1q and C3. Clin. Immunol. Immunopathol. 1992, 62, 133–138. [Google Scholar] [CrossRef]
- Heck, L.W.; Alarcon, P.G.; Kulhavy, R.M.; Morihara, K.; Russell, M.W.; Mestecky, J.F. Degradation of IgA proteins by Pseudomonas aeruginosa elastase. J. Immunol. 1990, 144, 2253–2257. [Google Scholar] [PubMed]
- Kharazmi, A. Interactions of Pseudomonas aeruginosa proteases with the cells of the immune system. Antibiot Chemother. 1989, 42, 42–49. [Google Scholar]
- Howe, T.R.; Iglewski, B.H. Isolation and characterization of alkaline protease-deficient mutants of Pseudomonas aeruginosa in vitro and in a mouse eye model. Infect. Immun. 1984, 43, 1058–1063. [Google Scholar]
- Kernacki, K.A.; Hobden, J.A.; Hazlett, L.D.; Fridman, R.; Berk, R.S. In vivo bacterial protease production during Pseudomonas aeruginosa corneal infection. Investig. Ophthalmol. Vis. Sci. 1995, 36, 1371–1378. [Google Scholar]
- Kessler, E.; Safrin, M.; Olson, J.C.; Ohman, D.E. Secreted LasA of Pseudomonas aeruginosa is a staphylolytic protease. J. Biol. Chem. 1993, 268, 7503–7508. [Google Scholar]
- Preston, M.J.; Seed, P.C.; Toder, D.S.; Iglewski, B.H.; Ohman, D.E.; Gustin, J.K.; Goldberg, J.B.; Pier, G.B. Contribution of proteases and LasR to the virulence of Pseudomonas aeruginosa during corneal infections. Infect. Immun. 1997, 65, 3086–3090. [Google Scholar] [PubMed]
- Alionte, L.G.; Cannon, B.M.; White, C.D.; Caballero, A.R.; O’Callaghan, R.J.; Hobden, J.A. Pseudomonas aeruginosa LasA protease and corneal infections. Curr. Eye Res. 2001, 22, 266–271. [Google Scholar] [CrossRef] [PubMed]
- White, C.D.; Alionte, L.G.; Cannon, B.M.; Caballero, A.R.; O’Callaghan, R.J.; Hobden, J.A. Corneal virulence of LasA protease--deficient Pseudomonas aeruginosa PAO1. Cornea 2001, 20, 643–646. [Google Scholar] [CrossRef] [PubMed]
- Hobden, J.A. Pseudomonas aeruginosa proteases and corneal virulence. DNA Cell Biol. 2002, 21, 391–396. [Google Scholar] [CrossRef] [PubMed]
- Kessler, E.; Safrin, M. Elastinolytic and proteolytic enzymes. Methods Mol. Biol. 2014, 1149, 135–169. [Google Scholar] [PubMed]
- Cryz, S.J.; Iglewski, B.H. Production of alkaline protease by Pseudomonas aeruginosa. J. Clin. Microbiol. 1980, 12, 131–133. [Google Scholar] [PubMed]
- Elliott, B.W.; Cohen, C. Isolation and characterization of a lysine-specific protease from Pseudomonas aeruginosa. J. Biol. Chem. 1986, 261, 11259–11265. [Google Scholar] [PubMed]
- O’Callaghan, R.J.; Engel, L.S.; Hobden, J.A.; Callegan, M.C.; Green, L.C.; Hill, J.M. Pseudomonas keratitis. the role of an uncharacterized exoprotein, protease IV, in corneal virulence. Investig. Ophthalmol. Vis. Sci. 1996, 37, 534–543. [Google Scholar]
- Marquart, M.E.; Caballero, A.R.; Chomnawang, M.; Thibodeaux, B.A.; Twining, S.S.; O’Callaghan, R.J. Identification of a novel secreted protease from Pseudomonas aeruginosa that causes corneal erosions. Investig. Ophthalmol. Vis. Sci. 2005, 46, 3761–3768. [Google Scholar] [CrossRef] [PubMed]
- Tang, A.; Marquart, M.E.; Fratkin, J.D.; McCormick, C.C.; Caballero, A.R.; Gatlin, H.P.; O’Callaghan, R.J. Properties of PASP: A Pseudomonas protease capable of mediating corneal erosions. Investig. Ophthalmol. Vis. Sci. 2009, 50, 3794–3801. [Google Scholar] [CrossRef]
- Tang, A.; Caballero, A.R.; Marquart, M.E.; O’Callaghan, R.J. Pseudomonas aeruginosa small protease (PASP), a keratitis virulence factor. Investig. Ophthalmol. Vis. Sci. 2013, 54, 2821–2828. [Google Scholar] [CrossRef] [PubMed]
- Tang, A.; Caballero, A.R.; Marquart, M.E.; Bierdeman, M.A.; O’Callaghan, R.J. Mechanism of Pseudomonas aeruginosa small protease (PASP), a corneal virulence factor. Investig. Ophthalmol Vis. Sci. 2018, 59, 5993–6002. [Google Scholar] [CrossRef] [PubMed]
- Kida, Y.; Higashimoto, Y.; Inoue, H.; Shimizu, T.; Kuwano, K. A novel secreted protease from Pseudomonas aeruginosa activates NF-kappaB through protease-activated receptors. Cell. Microbiol. 2008, 10, 1491–1504. [Google Scholar] [CrossRef] [PubMed]
- Kida, Y.; Taira, J.; Yamamoto, T.; Higashimoto, Y.; Kuwano, K. EprS, an autotransporter protein of Pseudomonas aeruginosa, possessing serine protease activity induces inflammatory responses through protease-activated receptors. Cell. Microbiol. 2013, 15, 1168–1181. [Google Scholar] [CrossRef] [PubMed]
- Passmore, I.J.; Nishikawa, K.; Lilley, K.S.; Bowden, S.D.; Chung, J.C.S.; Welch, M. Mep72, a metzincin protease that is preferentially secreted by biofilms of Pseudomonas aeruginosa. J. Bacteriol. 2015, 197, 762–773. [Google Scholar] [CrossRef] [PubMed]
- Ogino, H.; Watanabe, F.; Yamada, M.; Nakagawa, S.; Hirose, T.; Noguchi, A.; Yasuda, M.; Ishikawa, H. Purification and characterization of organic solvent-stable protease from organic solvent-tolerant Pseudomonas aeruginosa PST-01. J. Biosci. Bioeng. 1999, 87, 61–68. [Google Scholar] [CrossRef]
- Toder, S.; Gambello, M. Pseudomonas aeruginosa protease IV: Cloning of a caseinase which is not transcriptionally activated by the lasR. Gen. Meet. Am. Soc. Microbiol. 1992. [Google Scholar]
- Engel, L.S.; Hill, J.M.; Caballero, A.R.; Green, L.C.; O’Callaghan, R.J. Protease IV, a unique extracellular protease and virulence factor from Pseudomonas aeruginosa. J. Biol. Chem. 1998, 273, 16792–16797. [Google Scholar] [CrossRef]
- Caballero, A.R.; Moreau, J.M.; Engel, L.S.; Marquart, M.E.; Hill, J.M.; O’Callaghan, R.J. Pseudomonas aeruginosa protease IV enzyme assays and comparison to other Pseudomonas proteases. Anal. Biochem. 2001, 290, 330–337. [Google Scholar] [CrossRef]
- Caballero, A.; Thibodeaux, B.; Marquart, M.; Traidej, M.; O’Callaghan, R. Pseudomonas keratitis: Protease IV gene conservation, distribution, and production relative to virulence and other Pseudomonas proteases. Investig. Ophthalmol. Vis. Sci. 2004, 45, 522–530. [Google Scholar] [CrossRef]
- Traidej, M.; Marquart, M.E.; Caballero, A.R.; Thibodeaux, B.A.; O’Callaghan, R.J. Identification of the active site residues of Pseudomonas aeruginosa protease IV. Importance of enzyme activity in autoprocessing and activation. J. Biol. Chem. 2003, 278, 2549–2553. [Google Scholar] [CrossRef] [PubMed]
- Traidej, M.; Caballero, A.R.; Marquart, M.E.; Thibodeaux, B.A.; O’Callaghan, R.J. Molecular analysis of Pseudomonas aeruginosa protease IV expressed in Pseudomonas putida. Investig. Ophthalmol. Vis. Sci. 2003, 44, 190–196. [Google Scholar] [CrossRef] [PubMed]
- Conibear, T.C.R.; Willcox, M.D.P.; Flanagan, J.L.; Zhu, H. Characterization of protease IV expression in Pseudomonas aeruginosa clinical isolates. J. Med. Microbiol. 2012, 61, 180–190. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Cai, M.; Wu, F.; Zhang, Y.; Xiong, Z.; Xu, P. Recombinant expression, refolding, purification and characterization of Pseudomonas aeruginosa protease IV in Escherichia coli. Protein Expr. Purif. 2016, 126, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Wilderman, P.J.; Vasil, A.I.; Johnson, Z.; Wilson, M.J.; Cunliffe, H.E.; Lamont, I.L.; Vasil, M.L. Characterization of an endoprotease (PrpL) encoded by a PvdS-regulated gene in Pseudomonas aeruginosa. Infect. Immun. 2001, 69, 5385–5394. [Google Scholar] [CrossRef] [PubMed]
- Oh, J.; Li, X.-H.; Kim, S.-K.; Lee, J.-H. Post-secretional activation of Protease IV by quorum sensing in Pseudomonas aeruginosa. Sci. Rep. 2017, 7, 4416. [Google Scholar] [CrossRef] [PubMed]
- Ishii, T.; Fukushima, J.; Fujita, S.; Shigematsu, T.; Ando, N.; Ishiwata, T.; Kurata, M.; Kawamoto, S.; Morihara, K.; Okuda, K. Elastase gene expression in non-elastase-producing Pseudomonas aeruginosa strains using novel shuttle vector systems. FEMS Microbiol. Lett. 1994, 116, 307–313. [Google Scholar] [CrossRef] [PubMed]
- Bellemare, A.; Vernoux, N.; Morisset, D.; Bourbonnais, Y. Human pre-elafin inhibits a Pseudomonas aeruginosa-secreted peptidase and prevents its proliferation in complex media. Antimicrob. Agents Chemother. 2008, 52, 483–490. [Google Scholar] [CrossRef]
- Beaufort, N.; Seweryn, P.; de Bentzmann, S.; Tang, A.; Kellermann, J.; Grebenchtchikov, N.; Schmitt, M.; Sommerhoff, C.P.; Pidard, D.; Magdolen, V. Activation of human pro-urokinase by unrelated proteases secreted by Pseudomonas aeruginosa. Biochem. J. 2010, 428, 473–482. [Google Scholar] [CrossRef]
- Malloy, J.L.; Veldhuizen, R.A.W.; Thibodeaux, B.A.; O’Callaghan, R.J.; Wright, J.R. Pseudomonas aeruginosa protease IV degrades surfactant proteins and inhibits surfactant host defense and biophysical functions. Am. J. Physiol. Lung Cell Mol. Physiol. 2005, 288, L409–L418. [Google Scholar] [CrossRef]
- Guillon, A.; Brea, D.; Morello, E.; Tang, A.; Jouan, Y.; Ramphal, R.; Korkmaz, B.; Perez-Cruz, M.; Trottein, F.; O’Callaghan, R.J.; et al. Pseudomonas aeruginosa proteolytically alters the interleukin 22-dependent lung mucosal defense. Virulence 2017, 8, 810–820. [Google Scholar] [CrossRef] [PubMed]
- West, S.E.; Sample, A.K.; Runyen-Janecky, L.J. The vfr gene product, required for Pseudomonas aeruginosa exotoxin A and protease production, belongs to the cyclic AMP receptor protein family. J. Bacteriol. 1994, 176, 7532–7542. [Google Scholar] [CrossRef] [PubMed]
- Kanack, K.J.; Runyen-Janecky, L.J.; Ferrell, E.P.; Suh, S.-J.; West, S.E.H. Characterization of DNA-binding specificity and analysis of binding sites of the Pseudomonas aeruginosa global regulator, Vfr, a homologue of the Escherichia coli cAMP receptor protein. Microbiology 2006, 152, 3485–3496. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, E.L.; Brutinel, E.D.; Klem, E.R.; Fehr, A.R.; Yahr, T.L.; Wolfgang, M.C. In vitro and in vivo characterization of the Pseudomonas aeruginosa cyclic AMP (cAMP) phosphodiesterase CpdA, required for cAMP homeostasis and virulence factor regulation. J. Bacteriol. 2010, 192, 2779–2790. [Google Scholar] [CrossRef] [PubMed]
- Stintzi, A.; Evans, K.; Meyer, J.M.; Poole, K. Quorum-sensing and siderophore biosynthesis in Pseudomonas aeruginosa: lasR/lasI mutants exhibit reduced pyoverdine biosynthesis. FEMS Microbiol. Lett. 1998, 166, 341–345. [Google Scholar] [CrossRef] [PubMed]
- Arevalo-Ferro, C.; Hentzer, M.; Reil, G.; Görg, A.; Kjelleberg, S.; Givskov, M.; Riedel, K.; Eberl, L. Identification of quorum-sensing regulated proteins in the opportunistic pathogen Pseudomonas aeruginosa by proteomics. Environ. Microbiol. 2003, 5, 1350–1369. [Google Scholar] [CrossRef]
- Nouwens, A.S.; Beatson, S.A.; Whitchurch, C.B.; Walsh, B.J.; Schweizer, H.P.; Mattick, J.S.; Cordwell, S.J. Proteome analysis of extracellular proteins regulated by the las and rhl quorum sensing systems in Pseudomonas aeruginosa PAO1. Microbiology 2003, 149, 1311–1322. [Google Scholar] [CrossRef] [PubMed]
- Lamont, I.L.; Beare, P.A.; Ochsner, U.; Vasil, A.I.; Vasil, M.L. Siderophore-mediated signaling regulates virulence factor production in Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. USA 2002, 99, 7072–7077. [Google Scholar] [CrossRef]
- Beare, P.A.; For, R.J.; Martin, L.W.; Lamont, I.L. Siderophore-mediated cell signalling in Pseudomonas aeruginosa: Divergent pathways regulate virulence factor production and siderophore receptor synthesis. Mol. Microbiol. 2003, 47, 195–207. [Google Scholar] [CrossRef]
- Ochsner, U.A.; Wilderman, P.J.; Vasil, A.I.; Vasil, M.L. GeneChip expression analysis of the iron starvation response in Pseudomonas aeruginosa: Identification of novel pyoverdine biosynthesis genes. Mol. Microbiol. 2002, 45, 1277–1287. [Google Scholar] [CrossRef]
- Britigan, B.E.; Hayek, M.B.; Doebbeling, B.N.; Fick, R.B. Transferrin and lactoferrin undergo proteolytic cleavage in the Pseudomonas aeruginosa-infected lungs of patients with cystic fibrosis. Infect. Immun. 1993, 61, 5049–5055. [Google Scholar] [PubMed]
- Kim, E.-J.; Wang, W.; Deckwer, W.-D.; Zeng, A.-P. Expression of the quorum-sensing regulatory protein LasR is strongly affected by iron and oxygen concentrations in cultures of Pseudomonas aeruginosa irrespective of cell density. Microbiology. 2005, 151, 1127–1138. [Google Scholar] [CrossRef] [PubMed]
- Imperi, F.; Tiburzi, F.; Fimia, G.M.; Visca, P. Transcriptional control of the pvdS iron starvation sigma factor gene by the master regulator of sulfur metabolism CysB in Pseudomonas aeruginosa. Environ. Microbiol. 2010, 12, 1630–1642. [Google Scholar] [PubMed]
- Marquart, M.E.; Dajcs, J.J.; Caballero, A.R.; Thibodeaux, B.A.; O’Callaghan, R.J. Calcium and magnesium enhance the production of Pseudomonas aeruginosa protease IV, a corneal virulence factor. Med. Microbiol. Immunol. 2005, 194, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Sarkisova, S.; Patrauchan, M.A.; Berglund, D.; Nivens, D.E.; Franklin, M.J. Calcium-induced virulence factors associated with the extracellular matrix of mucoid Pseudomonas aeruginosa biofilms. J Bacteriol. 2005, 187, 4327–4337. [Google Scholar] [CrossRef] [PubMed]
- Termine, E.; Michel, G.P.F. Transcriptome and secretome analyses of the adaptive response of Pseudomonas aeruginosa to suboptimal growth temperature. Int. Microbiol. 2009, 12, 7–12. [Google Scholar] [PubMed]
- Visaggio, D.; Pasqua, M.; Bonchi, C.; Kaever, V.; Visca, P.; Imperi, F. Cell aggregation promotes pyoverdine-dependent iron uptake and virulence in Pseudomonas aeruginosa. Front Microbiol. 2015, 6, 902. [Google Scholar] [CrossRef]
- Zaborin, A.; Romanowski, K.; Gerdes, S.; Holbrook, C.; Lepine, F.; Long, J.; Poroyko, V.; Diggle, S.P.; Wilke, A.; Righetti, K.; et al. Red death in Caenorhabditis elegans caused by Pseudomonas aeruginosa PAO1. Proc. Natl. Acad. Sci. USA 2009, 106, 6327–6332. [Google Scholar] [CrossRef]
- Romanowski, K.; Zaborin, A.; Fernandez, H.; Poroyko, V.; Valuckaite, V.; Gerdes, S.; Liu, D.C.; Zaborina, O.Y.; Alverdy, J.C. Prevention of siderophore- mediated gut-derived sepsis due to P. aeruginosa can be achieved without iron provision by maintaining local phosphate abundance: Role of pH. BMC Microbiol. 2011, 11, 212. [Google Scholar] [CrossRef]
- Long, J.; Zaborina, O.; Holbrook, C.; Zaborin, A.; Alverdy, J. Depletion of intestinal phosphate following surgical injury activates the virulence of P. aeruginosa causing lethal gut-derived sepsis. Surgery 2008, 144, 189–197. [Google Scholar] [CrossRef]
- Overhage, J.; Bains, M.; Brazas, M.D.; Hancock, R.E.W. Swarming of Pseudomonas aeruginosa is a complex adaptation leading to increased production of virulence factors and antibiotic resistance. J. Bacteriol. 2008, 190, 2671–2679. [Google Scholar] [CrossRef] [PubMed]
- Tan, S.Y.-Y.; Liu, Y.; Chua, S.L.; Vejborg, R.M.; Jakobsen, T.H.; Chew, S.C.; Li, Y.; Nielsen, T.E.; Tolker-Nielsen, T.; Yang, L.; et al. Comparative systems biology analysis to study the mode of action of the isothiocyanate compound iberin on Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2014, 58, 6648–6659. [Google Scholar] [CrossRef] [PubMed]
- Imperi, F.; Massai, F.; Facchini, M.; Frangipani, E.; Visaggio, D.; Leoni, L.; Bragonzi, A.; Visca, P. Repurposing the antimycotic drug flucytosine for suppression of Pseudomonas aeruginosa pathogenicity. Proc. Natl. Acad. Sci. USA 2013, 110, 7458–7463. [Google Scholar] [CrossRef] [PubMed]
- Sonawane, A.; Jyot, J.; Ramphal, R. Pseudomonas aeruginosa LecB is involved in pilus biogenesis and protease IV activity but not in adhesion to respiratory mucins. Infect. Immun. 2006, 74, 7035–7039. [Google Scholar] [CrossRef] [PubMed]
- Andrejko, M.; Cytryńska, M.; Jakubowicz, T. Apolipophorin III is a substrate for protease IV from Pseudomonas aeruginosa. FEMS Microbiol. Lett. 2005, 243, 331–337. [Google Scholar] [CrossRef] [PubMed]
- Andrejko, M.; Mizerska-Dudka, M.; Jakubowicz, T. Changes in Galleria mellonella apolipophorin III level during Pseudomonas aeruginosa infection. J. Invertebr. Pathol. 2008, 97, 14–19. [Google Scholar] [CrossRef]
- Cheng, Z.; Li, J.-F.; Niu, Y.; Zhang, X.-C.; Woody, O.Z.; Xiong, Y.; Djonović, S.; Millet, Y.; Bush, J.; McConkey, B.J.; et al. Pathogen-secreted proteases activate a novel plant immune pathway. Nature 2015, 521, 213–216. [Google Scholar] [CrossRef] [PubMed]
- Mizerska-Dudka, M.; Andrejko, M. Galleria mellonella hemocytes destruction after infection with Pseudomonas aeruginosa. J. Basic Microbiol. 2014, 54, 232–246. [Google Scholar] [CrossRef]
- Park, S.-J.; Kim, S.-K.; So, Y.-I.; Park, H.-Y.; Li, X.-H.; Yeom, D.H.; Lee, M.-N.; Lee, B.-L.; Lee, J.-H. Protease IV, a quorum sensing-dependent protease of Pseudomonas aeruginosa modulates insect innate immunity. Mol. Microbiol. 2014, 94, 1298–1314. [Google Scholar] [CrossRef]
- Bradshaw, J.L.; Caballero, A.R.; Bierdeman, M.A.; Adams, K.V.; Pipkins, H.R.; Tang, A.; O’Callaghan, R.J.; McDaniel, L.S. Pseudomonas aeruginosa protease IV exacerbates pneumococcal pneumonia and systemic disease. mSphere 2018, 3, e00212-18. [Google Scholar] [CrossRef]
- Thibodeaux, B.A.; Caballero, A.R.; Dajcs, J.J.; Marquart, M.E.; Engel, L.S.; O’Callaghan, R.J. Pseudomonas aeruginosa protease IV: A corneal virulence factor of low immunogenicity. Ocul. Immunol. Inflamm. 2005, 13, 169–182. [Google Scholar] [CrossRef] [PubMed]
- Upritchard, H.G.; Cordwell, S.J.; Lamont, I.L. Immunoproteomics to examine cystic fibrosis host interactions with extracellular Pseudomonas aeruginosa proteins. Infect. Immun. 2008, 76, 4624–4632. [Google Scholar] [CrossRef] [PubMed]
- Engel, L.S.; Hobden, J.A.; Moreau, J.M.; Callegan, M.C.; Hill, J.M.; O’Callaghan, R.J. Pseudomonas deficient in protease IV has significantly reduced corneal virulence. Investig. Ophthalmol. Vis. Sci. 1997, 38, 1535–1542. [Google Scholar] [PubMed]
- Engel, L.S.; Hill, J.M.; Moreau, J.M.; Green, L.C.; Hobden, J.A.; O’Callaghan, R.J. Pseudomonas aeruginosa protease IV produces corneal damage and contributes to bacterial virulence. Investig. Ophthalmol. Vis. Sci. 1998, 39, 662–665. [Google Scholar] [PubMed]
- Rahme, L.G.; Ausubel, F.M.; Cao, H.; Drenkard, E.; Goumnerov, B.C.; Lau, G.W.; Mahajan-Miklos, S.; Plotnikova, J.; Tan, M.W.; Tsongalis, J.; et al. Plants and animals share functionally common bacterial virulence factors. Proc. Natl. Acad. Sci. USA 2000, 97, 8815–8821. [Google Scholar] [CrossRef] [Green Version]
- Gotz, P.; Weise, C.; Kopacek, P.; Losen, S.; Wiesner, A. Isolated apolipophorin III from Galleria mellonella stimulates the immune reactions of this insect. J. Insect Physiol. 1997, 43, 383–391. [Google Scholar] [PubMed]
- Wu, Y.T.-Y.; Willcox, M.; Zhu, H.; Stapleton, F. Contact lens hygiene compliance and lens case contamination: A review. Cont. Lens Anterior Eye 2015, 38, 307–316. [Google Scholar] [CrossRef]
- Twining, S.S.; Kirschner, S.E.; Mahnke, L.A.; Frank, D.W. Effect of Pseudomonas aeruginosa elastase, alkaline protease, and exotoxin A on corneal proteinases and proteins. Investig. Ophthalmol. Vis. Sci. 1993, 34, 2699–2712. [Google Scholar]
- Estrellas, P.S.; Alionte, L.G.; Hobden, J.A. A Pseudomonas aeruginosa strain isolated from a contact lens-induced acute red eye (CLARE) is protease-deficient. Curr. Eye Res. 2000, 20, 157–165. [Google Scholar] [CrossRef]
- Choy, M.H.; Stapleton, F.; Willcox, M.D.P.; Zhu, H. Comparison of virulence factors in Pseudomonas aeruginosa strains isolated from contact lens- and non-contact lens-related keratitis. J. Med Microbiol. 2008, 57, 1539–1546. [Google Scholar] [CrossRef]
- Zhu, H.; Conibear, T.C.R.; Bandara, R.; Aliwarga, Y.; Stapleton, F.; Willcox, M.D.P. Type III secretion system-associated toxins, proteases, serotypes, and antibiotic resistance of Pseudomonas aeruginosa isolates associated with keratitis. Curr. Eye Res. 2006, 31, 297–306. [Google Scholar] [CrossRef] [PubMed]
- Madhu, S.N.; Jha, K.K.; Karthyayani, A.P.; Gajjar, D.U. Ex vivo caprine model to study virulence factors in keratitis. J. Ophthalmic. Vis. Res. 2018, 13, 383–391. [Google Scholar] [PubMed]
- Smith, L.; Rose, B.; Tingpej, P.; Zhu, H.; Conibear, T.; Manos, J.; Bye, P.; Elkins, M.; Willcox, M.; Bell, S.; et al. Protease IV production in Pseudomonas aeruginosa from the lungs of adults with cystic fibrosis. J. Med. Microbiol. 2006, 55, 1641–1644. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, F.; Masiá, M.; Rodríguez, J.C.; Mirete, C.; Soldán, B.; Padilla, S.; Hernández, I.; Royo, G.; Martin-Hidalgo, A. Community-acquired pneumonia of mixed etiology: Prevalence, clinical characteristics, and outcome. Eur. J. Clin. Microbiol. Infect. Dis. 2005, 24, 377–383. [Google Scholar] [CrossRef] [PubMed]
- Okada, F.; Ando, Y.; Matsushita, S.; Ishii, R.; Nakayama, T.; Morikawa, K.; Ono, A.; Maeda, T.; Mori, H. Thin-section CT findings of patients with acute Streptococcus pneumoniae pneumonia with and without concurrent infection. Br. J. Radiol. 2012, 85, e357–e364. [Google Scholar] [CrossRef] [PubMed]
- Scott, N.E.; Hare, N.J.; White, M.Y.; Manos, J.; Cordwell, S.J. Secretome of transmissible Pseudomonas aeruginosa AES-1R grown in a cystic fibrosis lung-like environment. J. Proteome Res. 2013, 12, 5357–5369. [Google Scholar] [CrossRef] [PubMed]
- Fothergill, J.L.; Walshaw, M.J.; Winstanley, C. Transmissible strains of Pseudomonas aeruginosa in cystic fibrosis lung infections. Eur. Respir. J. 2012, 40, 227–238. [Google Scholar] [CrossRef]
- Zegans, M.E.; Becker, H.I.; Budzik, J.; O’Toole, G. The role of bacterial biofilms in ocular infections. DNA Cell Biol. 2002, 21, 415–420. [Google Scholar] [CrossRef]


{kind=link}
| Name | Abbreviation | Type | Comment | Reference |
|---|---|---|---|---|
| Elastase A | LasA | Metalloprotease | Staphylolytic protease | [25] |
| Elastase B | LasB | Metalloprotease | Potent enzyme and virulence related | [25] |
| Alkaline Protease | AP | Metalloprotease | Broad spectrum protease and virulence related | [26] |
| Protease IV | PIV (Ps-1; PrpL) | Serine Protease | Lysine specific and important virulence factor | [27,28] |
| Pseudomonas aeruginosa Small Protease | PASP | Serine Protease | Cleaves collagen and other proteins, virulence factor | [29,30,31,32] |
| Large Protease | LepA | Serine Protease | Activates inflammation and virulence factor | [33] |
| Autotransporter Protein | EprS | Serine Protease | Activates inflammation | [34] |
| Metzincin Protease | Mep72 | Metalloprotease | Biofilm secreted protein | [35] |
| ATCC27853 Protease | / | Metalloprotease | Stable in organic solvents | [36] |
| Type of Host | Host Studied | Effect on Host | References |
|---|---|---|---|
| Plant | Arabidopsis (thale cress, a cabbage) | Disruption of normal homeostatis and induction of a defensive host response | [77] |
| Insect | Tenebrio molitor (mealworms) or Galleria mellonella (wax moth) | Bacteremia protected by PIV-mediated killing of defensive cells. Also, destruction of apolipophorin III to reduce phagocytosis of bacteria | [75,76,78,79] |
| Mammals | Rabbit corneal infection | Destruction of host defensive proteins and possibly collagens and induction of inflammation | [38,39,40,41,42,64,81,83,84] |
| Rodent respiratory tract | Wild-type P. aeruginosa in agar beads had more colony forming units that a mutant deficient in PIV. | [45] | |
| Rodent respiratory tract | Destruction of host defense proteins (surfactants A, D, and B) that opsonize the bacteria. Also, enhance bacteria (CFU) in lung more than PIV-lacking mutant | [50] | |
| Mouse lung | PIV enhanced pneumococcal lung infection and bacteremia by destroying IL-22 in the lung. | [80] | |
| Human lung | Destroy IL-22 in human lung fluids including sputum of patients with Pseudomonas pneumonia | [51] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
O’Callaghan, R.; Caballero, A.; Tang, A.; Bierdeman, M. Pseudomonas aeruginosa Keratitis: Protease IV and PASP as Corneal Virulence Mediators. Microorganisms 2019, 7, 281. https://doi.org/10.3390/microorganisms7090281
O’Callaghan R, Caballero A, Tang A, Bierdeman M. Pseudomonas aeruginosa Keratitis: Protease IV and PASP as Corneal Virulence Mediators. Microorganisms. 2019; 7(9):281. https://doi.org/10.3390/microorganisms7090281
Chicago/Turabian StyleO’Callaghan, Richard, Armando Caballero, Aihua Tang, and Michael Bierdeman. 2019. "Pseudomonas aeruginosa Keratitis: Protease IV and PASP as Corneal Virulence Mediators" Microorganisms 7, no. 9: 281. https://doi.org/10.3390/microorganisms7090281
APA StyleO’Callaghan, R., Caballero, A., Tang, A., & Bierdeman, M. (2019). Pseudomonas aeruginosa Keratitis: Protease IV and PASP as Corneal Virulence Mediators. Microorganisms, 7(9), 281. https://doi.org/10.3390/microorganisms7090281