Relevant Aspects of Clostridium estertheticum as a Specific Spoilage Organism of Vacuum-Packed Meat


Institute for Food Safety and Hygiene, Vetsuisse Faculty, University of Zurich, Winterthurerstrasse 272, 8057 Zurich, Switzerland
*
Author to whom correspondence should be addressed.
Microorganisms 2019, 7(5), 142; https://doi.org/10.3390/microorganisms7050142
Submission received: 22 April 2019
/
Revised: 12 May 2019
/
Accepted: 16 May 2019
/
Published: 20 May 2019
(This article belongs to the Special Issue Bioprotection in Meat and Meat Products)
Abstract
:Clostridium estertheticum is a psychrotolerant, gram-positive, motile, anaerobic, spore-forming, rod-shaped bacteria that causes blown pack spoilage (BPS). Spoilage occurs in vacuum-packed meat without temperature abuse. Having been reported in the last 30 years in several countries, BPS by Cl. estertheticum is a major issue around the world and presents a huge economic impact on the meat industry. Despite being an important spoilage microorganism, studies on Cl. estertheticum are challenged by numerous aspects. These include, lack or poor growth in laboratory media, long culturing periods, and unpredictable isolation on the media. These factors hamper the detection of Cl. estertheticum before occurrence of BPS, which further undermines efforts to prevent the occurrence of BPS. Nevertheless, considerable developments have taken place with regard to culture-independent methods. Although information on Cl. estertheticum is available, it is limited and remains highly fragmented. Therefore, this review collates the available information and discusses relevant aspects of Cl. estertheticum as a specific spoilage organism of BPS in vacuum-packed meat.
1. Introduction
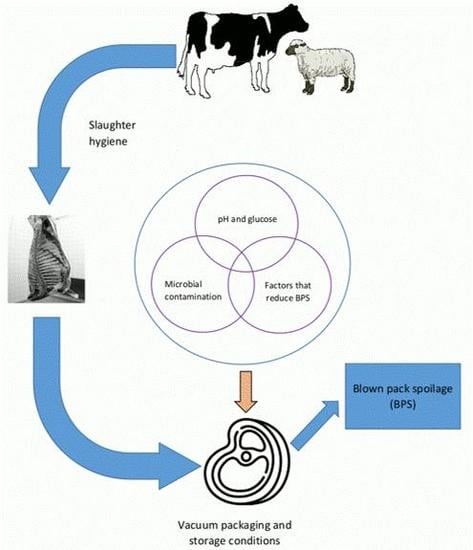
Meat is a highly perishable food commodity hence prone to microbial or chemical spoilage. Meat spoilage refers to alteration of color and production of off-odors, slime, and exudates that lead to unacceptable sensorial and organoleptic properties [1]. Spoilage of meat can have a significant effect on the global food supply. In Europe and Northern America, approximately 21% of food losses are from meat and meat products [2]. On the other hand, meat spoilage accounts for up to 40% of the production losses incurred by the meat processors and retailers [3]. These factors result in huge financial losses to the meat industry and are also a big issue in view of sustainability.
Meat spoilage results from a combination of microbial and chemical activities. Both are considered important, although microbial activities are a major cause of spoilage especially for raw meats [4,5]. Meat is generally considered sterile before slaughter, but the environment in which slaughter processes take place are not sterile, and therefore a degree of microbial contamination can occur [6], resulting in microbial spoilage. Post-slaughter microbial quality of meat is primarily determined by meat type, processing, distribution, and storage conditions [7]. Contaminated slaughter equipment, personnel, and environmental factors, such as water, air, and soil, can cross contaminate meat with spoilage-related bacteria species [8,9]. Upon storage, various intrinsic and extrinsic factors influence the process of microbial meat spoilage. These include oxygen demand, pH, temperature, and competing organisms [10]. The diversity of these ecophysiological factors affect the microbial growth dynamics, including succession of microorganisms and the microbiota composition and ultimately the type and rate of meat spoilage.
The ecosystem of meat offers easily available substrates among them glycogen and amino acids. The nutrients provide an enabling environment for diverse microbial growth and metabolism that results in spoilage [11]. Despite this, only a fraction of the initial microbial population on meat, which are referred to as ‘specific spoilage organisms’ (SSO) can develop during storage and cause spoilage [12,13]. In meat, the SSO metabolize available substrates, with subsequent changes in meat texture and production of volatile organic compounds responsible for off-odors [14]. SSO can also cause accumulation of purge especially in vacuum-packing [15].
Vacuum-packing is used by meat processors to control meat spoilage during storage. Its ability to prevent the growth of some food-borne pathogens and spoilage bacteria commonly present on meat makes it a widely used meat packing method [16]. In the vacuum-packing system, an anaerobic environment is created by removing oxygen followed by immediate sealing [17]. It is generally recognized that meat packing systems are important extrinsic factors that determines SSO in stored meats [18]. Effectively, exhaustion of oxygen alters the gaseous composition within the packs creating hurdles to the aerobic bacteria while enabling the growth of facultative and strict anaerobes [19].
In addition to creating anaerobic conditions, vacuum-packing maintains the pH of meat between 5.0 to 6.0 under long storage [20]. Lactic acid content in meat, which results from metabolism of glycogen by some anaerobic SSO [21], may also affect the growth of other SSO in vacuum packed meat by not only lowering the pH of meat but also favoring microorganisms that can metabolize it [22]. Cold storage of vacuum-packed meat further selects for psychrophilic or psychrotolerant SSO [12]. The main SSO in vacuum-packed meat include Streptococcus spp., Brochothrix spp., Psychrobacter spp., and Acinetobacter spp. [8,23,24]. A range of psychrotolerant and psychrophilic Clostridia have also been identified as SSO of chilled, vacuum-packed red meat [25].
Among the Clostridia, Clostridium estertheticum, Cl. algidicarnis, Cl. frigidicarnis, Cl. gasigenes, Cl. Frigoris, and Cl. bowmanii are particularly involved in spoilage of chilled vacuum-packed meat and meat products [26,27]. Spoilage of vacuum-packed meat by Cl. algidicarnis, Cl. frigoris, Cl. bowmanii, and Cl. frigidicarmis occurs without gas production while spoilage by Cl. estertheticum and Cl. gasigenes is characterized by gas production [25]. The spoilage by Cl. estertheticum and Cl. gasigenes is commonly referred to as ‘blown pack spoilage’ (BPS), and Cl. estertheticum is regarded as the main cause of BPS [28], making it the main SSO of BPS.
There are two recognized subspecies of Cl. estertheticum, Cl. estertheticum subp. estertheticum and Cl. estertheticum subp. laramiense [29,30], both of which are linked to BPS [31]. The first reports of BPS by Cl. estertheticum were made in 1989 of vacuum-packed raw beef in the UK and USA [32,33]. Later, BPS was reported in New Zealand [34] and Ireland [35], making it a global phenomenon with an impact on the meat industry. The present study reviews the characteristics of Cl. estertheticum as an SSO of BPS in refrigerated vacuum-packed meat.
2. Taxonomic Classification of Clostridium estertheticum
The Clostridium genus is a large, diverse group consisting of Gram-positive, spore-forming, obligate anaerobic firmicutes [36]. The genus was created in 1880 with the type species Cl. butyricum [37]. Currently, the genus consists of over 230 recognized species and subspecies [38]. This is attributed to the wide range of phenotypes displayed by the different species within the genus, which include synthesis of quinone and cytochromes, varying glycine and cysteine (GC) content, and wide growth temperature and acidity ranges [39]. The application of molecular methods, which include DNA–rRNA pairing and 16S rRNA cataloguing studies, have revealed the phylogenetic diversity and resulted in reclassification of some species within the genus [39,40]. Based on the phylogenetic analysis, the genus is currently divided into Cluster I and Cluster II with less than 80 species, among them Cl. estertheticum, falling into the Cluster I and are referred to as Clostridium sensu stricto [29,30].
Despite having been first identified and reported in the same year, different names were assigned to the species Cl. estertheticum [32,33], possibly because they were isolated in different countries. The organism identified in spoilt vacuum-packed meat in UK, was named Cl. estertheticum due to its ability to form esters [41]. On the other hand, the organism identified in spoilt vacuum-packed meat in the USA was named Cl. laramie in reference to the City of Laramie, Wyoming [42]. Collins [41] characterized the organism through 16S ribosomal RNA (rRNA) while Kalchayanand et al. [42] characterized it through phenotypic tests and its GC content. The taxonomy of the two species was later resolved when DNA–DNA hybridization experiments revealed that they shared 79% DNA–DNA similarity and 16s RNA analysis revealed they formed a tight cluster, indicating a close relationship at the species level [43]. Therefore, Spring [43] proposed that the two species be merged into one species, named Cl. estertheticum, which was divided into two subspecies, Cl. estertheticum subsp. estertheticum and Cl. estertheticum subsp. laramiense.
Sequencing of Cl. estertheticum DSM 8809 strain, revealed that it harbors a marginally higher GC content than Cl. botulinum and Cl. perfringens [44]. On the other hand, Cl. estertheticum subsp. estertheticum and Cl. estertheticum subsp. laramiense do not cluster together in a phylogenetic tree despite their 16s RNA sequence being similar [30,43]. In particular, Cl. estertheticum subsp. laramiense clusters with Cl. frigioris, while Cl. estertheticum subsp. estertheticum clusters with Cl. lacusfryxellense in the range of 98.7–99.6% [30,43]. These differences have raised questions of classifying the two subspecies as a single species under the current classification system [45]. With respect to phenotypic characteristics, the two subspecies were initially reported to have differences in hemolytic activity, growth conditions, spore position and fermentation products [41,43]. These differences formed the basis for characterizing the two subspecies of Cl. estertheticum. It was later reported that the two organisms did not display these differences [45]. This exemplifies the challenges faced when using phenotypic traits within the Clostridium genus for taxonomic classification.
3. Isolation and Conventional Culturing of Clostridium estertheticum
Even though Cl. estertheticum was first reported in 1989, its isolation was hampered by its failure to grow in laboratory media available at the time, which included thioglycollate agar, brain heart infusion agar, lactose egg-yolk-milk agar, and trypticase peptone glucose yeast extract agar [32,33]. Similar results were reported 20 years later by Byrne et al. [35], whereby their efforts to isolate Cl. estertheticum from vacuum-packed purge in BPS samples using Colombia blood agar, tryptose sulphite cycloserine, Shahidi-Ferguson perfringens agar, reinforced clostridial agar, and brain-heart infusion agar resulted only in growth of other facultative anaerobic non-spore-forming bacteria other than Cl. estertheticum. Nevertheless, pure cultures of Cl. estertheticum could initially be grown on Reinforced Clostridium Medium (RCM) [41]. Later, Broda et al. [46] developed a pre-reduced RCM-based protocol that involved treatment of samples with ethanol and heat to recover Clostridia spores after inactivating other microorganisms and applied it to isolate Clostridium spp. from vacuum-packed purge samples. This method was used by Boerema et al. [47] to isolate Cl. estertheticum from slaughterhouse processing environments, which could then be identified on colony-based morphologies on agar supplemented with blood. The method was later compared against a method that included an enrichment step using pre-reduced peptone yeast extract glucose starch (PYGS) medium (Table 1) in isolation of Cl. estertheticum from abattoir samples and it was shown that the enrichment step enhanced the successful isolation of the organism [48].
While unsuccessful culturing of Cl. estertheticum has been attributed to the production of butanol within isolation media that eventually kills the organism’s cells [49], there are currently no dedicated media to overcome this challenge. Furthermore, there are no differential media for its identification due to variable phenotypic characteristics between the two subspecies [41,43,45,48] and among psychrophilic Clostridia [50]. For these reasons, Cl. estertheticum is not detectable through most culturing methods that are presently available [51]. Therefore, culture-based methods for Cl. estertheticum have, over the years, been based on non-specific media including PYGS medium and Columbia blood agar (CBA) supplemented with 5% defibrinated horse blood [52]. On CBA, Cl. estertheticum forms colonies that are round with often coarsely granulated margins, smooth, slightly raised, cream-white to greyish and semitransparent to opaque and can either be or not be β-hemolytic [48]. The current conventional based culture processes are time consuming because Cl. estertheticum’s optimal temperature for growth is low, hence production of a workable culture is slow, often taking up to three months [44]. Even with a working culture, lack of a comprehensive list of bacterial species in currently available commercial kits, which allow for phenotypic differentiation of the species, may further hamper the correct identification of Cl. estertheticum [53].
4. Molecular and Non-Molecular Based Identification of Clostridium estertheticum
Despite the different challenges of culture-dependent method, molecular methods, such as polymerase chain reaction (PCR), have proven reliable for detection of Cl. estertheticum. The success of some the methods is dependent on an enrichment step before molecular analysis [28,54]. Collins [41] described the first molecular based detection method for Cl. estertheticum, which was based on the 16S rRNA gene that allowed the differentiation of Cl. estertheticum from Cl. acetobutylicum, Cl. aurantibutyricum and Cl. tetanomorphum. Thereafter, two ribosomal DNA (rDNA) based methods were developed to detect Cl. estertheticum in broth, meat or meat purge [55]. Broda et al. [34] developed a 16S rDNA gene-based restriction fragment length polymorphism (RFLP) analysis for Cl. estertheticum differentiation from Cl. botulinum, Cl. algidicarnis, Cl. putrefaciens, Cl. Vincentii, and Cl. fimetarium. In an additional study, Broda et al. [56] used an 16S-23S rDNA internal transcribed spacer analysis to detect Cl. estertheticum in the meat processing environment. Broda et al. [57] then described the first set of primers, 16SEF and 16SER, which allowed the differentiation of Cl. estertheticum from other closely related Clostridia and microorganisms found in meat without RFLP analysis. The protocol was also suitable for the detection of Cl. estertheticum in commercial blown packs with a detection limit of 100 Cl. estertheticum cells per gram. This method was thereafter used and validated in different further studies [47,58,59].
In an effort to reduce detection time for Cl. estertheticum, Brightwell and Clemens [28] developed and validated a real-time PCR (RT-PCR) assay that was applicable in a variety of matrices including soil, hides, feces and meat (Table 2). Bonke et al. [50] compared this RT-PCR assay to the conventional PCR method described by Broda et al. [57], and showed that the RT-PCR assay was more sensitive. Based on the RT-PCR developed by Brightwell and Clemens [28], Reid et al. [60] developed and validated a further RT-PCR method for the simultaneous detection of low concentrations Cl. estertheticum, Cl. gasigenes and Cl. ruminantium in meat juice and wet or dry swab samples. The RT-PCR could detect five spores per milliliter without the need of an enrichment step, hence also speeding up the time taken to identify Cl. estertheticum considerably. Recently, Dorn-In et al. [25] developed a multiplex quantitative-PCR (q-PCR) (Table 2) for the detection of Cl. estertheticum as well as Cl. frigoriphilum, Cl. bowmanii or Cl. tagluense and showed that the q-PCR could be applied directly with DNA extracts of meat juice from BPS samples.
Cl. estertheticum subsp. estertheticum, and Cl. estertheticum subsp. laramiense could be differentiated by SmaI digestion of their DNA followed by pulsed-field gel electrophoresis (PFGE) analysis [45]. The PFGE analysis could differentiate between the two subspecies because of their distinctly different PFGE patterns, with a Dice similarity coefficient of 90%. Most recently, Amplified rDNA Restriction Analysis (ARDRA), which is a modified RFLP method, was used to differentiate Cl. estertheticum from other spoilage-associated species of Clostridia as well as other pyschrotolerant Clostridium species associated with meat production [61]. Most recently, Matrix Assisted Laser Desorption Ionization-Time of Flight Mass Spectrometry (MALDI-TOF MS) was used to identify Cl. estertheticum isolated from sheep and cattle carcasses at different slaughtering stages, and the results verified using 16S rDNA gene sequencing [53].
5. Growth and Metabolism of Clostridium estertheticum
Cl. estertheticum is an obligate anaerobe bacterium, hence it is sensitive to oxygen when in vegetative state [62]. Being a psychrotolerant Clostridia, Cl. estertheticum grows in a temperature range between −2 and 22 °C [63,64], and does not grow at 25 °C or above [41]. The optimum growth temperature ranges between 6–15 °C [43,45]. The type of substrate may also influence the growth temperatures given that Cl. estertheticum grew in meat juice at 20 °C but not in PYGS broth [63]. Cl. estertheticum grows at pH values in the range of 5.5 to 7.5 with maximum growth occurring between 5.8 and 6.8 [63]. In a growth assay in meat juice, Cl. estertheticum utilized both glucose and glycogen for growth, but exhaustion of glucose resulted in cessation of growth with a simultaneous utilization of lactate and production of CO2 and H2, but without growth probably due to low levels of acetate [65]. Cl. estertheticum produced butyrate, acetate, and formate from glucose and 1-butanol, ethanol, butyrate, and formate from lactate [45]. Cl. estertheticum also utilized amino acids that do not contain Sulphur for growth hence do not produce hydrogen sulfide [45]. Meat with a high pH and glucose concentration is the most conducive food matrix for the growth of Cl. estertheticum [66].
6. Clostridium estertheticum as a Causative Agent of Blown Pack Spoilage
It has been previously reported that BPS during refrigeration can also be caused by gas-producing Enterobacteriaceae, including Hafnia spp., Enterobacter spp., Serratia spp., Rahnella spp., and Ewingella spp. [67]. Nevertheless, these species do not grow below 4 °C [68]. Even though different SSO, including the Enterobacteriaceae and lactic acid bacteria, can cause BPS at refrigeration temperatures of 4 °C to 15 °C [69], the spoilage below 2 °C would be a major characteristic for BPS by Cl. estertheticum [46].
A four-year survey of BPS in Ireland found that the prevalence of Cl. estertheticum was higher than that of other Clostridia [70]. In another study, beef and lamb samples from Europe, North and South America and Oceania were investigated and it was also found that Cl. estertheticum was the most prevalent Clostridia [50]. A comparison among 11 Clostridium species found that all species were able to grow on vacuum-packed meat but only Cl. estertheticum and Cl. frigioris caused BPS [71]. Similar results were obtained by Silva et al. [72]. These reports emphasize that Cl. estertheticum is by far the most common psychrotolerant Clostridia associated with BPS of vacuum-packed raw meats, hence an SSO of refrigerated vacuum-packed meat below 2 °C.
Typically, a pack suffering from BPS caused by Cl. estertheticum contains copious quantities of drip and gas, whereby the latter leads to gross pack distension [64]. Upon opening of dissented packs, BPS may be characterized by either a highly unpleasant odor followed by a very strong fruity and dairy odors [33,63]. The three odors cannot be used conclusively to characterize BPS by Cl. estertheticum given that, despite its inability to produce hydrogen sulfide [45], an unpleasant odor was perceived from a naturally spoilt pack [32], but absent in another study [63]. Spoiled meat in BPS packs is nonetheless discolored and excessively tender [33].
BPS is not regarded as a safety hazard, but meat spoiled in this way has no commercial value and causes significant financial losses to the meat industry [70]. Losses that are specifically caused by Cl. estertheticum are likely to occur in the summer, which is the season with the highest prevalence of Cl. estertheticum [70,73]. Although the safety risks associated with Cl. estertheticum are regarded to be low, genome analysis revealed that Cl. estertheticum harbors multiple genes potentially related to antibiotic, biocide and metal resistance, along with several predicted virulence factor genes [44].
7. Factors Affecting Blown Pack Spoilage by Clostridium estertheticum
Being anaerobic, BPS by Cl. estertheticum is primarily dependent on the presence of spores on meat prior to packing and their ability to germinate within vacuum packs. As low as one spore of Cl. estertheticum is sufficient to cause BPS [64]. It was suggested that 100 spores per cm2 of Cl. estertheticum is a critical number for vacuum-packed meat [74]. The rate of spore germination, hence the ratio of spores and vegetative cells during the ongoing process of BPS, can also influence the occurrence of BPS [75]. In particular, both neutral pH and lactate increase the rate of spore germination [62]. On the other hand, the maximum numbers of Cl. estertheticum in the spoilage flora of vacuum-packed meat was shown to depend on the amount of glucose available for its growth [65]. Therefore, the amount of glucose in meat after slaughter is a key factor because it is the first substrate preferably utilized by most bacteria growing in raw meat during refrigerated storage [14]. Moreover, the ability of Cl. estertheticum to compete with microflora that utilize glucose, such as Leuconostoc mesenteroides in vacuum-packed meat can influence the occurrence of BPS [76].
Storage temperature has an influence on the occurrence of BPS. In order to minimize meat spoilage by anaerobic bacteria, a storage temperature of −1.5 °C has been recommended, and in vacuum-packed meat, temperature above 0 °C is considered abusive [77]. The shelf life for vacuum-packed meat stored at −1.5 °C ranges between 60 to 70 days [78]. Unfortunately, BPS by Cl. estertheticum can occur in the absence of temperature abuse or packaging failure [59]. As low as 10 spores per cm2 of Cl. estertheticum can reduce the shelf life of vacuum-packed meat to 44 days at −1.5 °C [64]. At 2 °C and 15 °C, packs inoculated with Cl. estertheticum presented the first signs of BPS after 15 and 4 days of storage, respectively [72]. While storage of meat at −1.5 °C evidently slows the rate of BPS, the time it takes for the meat to reach −1.5 °C after chilling had no effect on the occurrence of BPS [75]. Another temperature related factor is the application of heat shrink. This involves dipping vacuum packs of meats in water at 85–90 °C, for 2–3 s immediately after pack sealing to improve the pack integrity [79]. This practice has been shown to accelerate the onset of BPS by Cl. estertheticum whereby gas was produced in heat-treated packs much earlier than pack without heat treatment [80].
Occurrence of BPS can also be influenced by post-slaughter deboning of carcasses. Deboning can either be hot-boning, which involves deboning before chilling or cold-boning, which involves deboning after chilling [81]. Hot-boning has numerous economic and technological benefits that include less chill and drip loss, cooler space, electricity, and capital investment and results in high quality meat in terms of high pH, water-holding capacity, and emulsifying capacity [82,83]. A comparison of the two deboning techniques showed that hot-boning resulted in earlier detection of BPS caused by Cl. estertheticum than in cold-boning [60].
8. Intervention and Inactivation Strategies to Reduce Blown Pack Spoilage by Clostridium estertheticum
The process of BPS occurs suddenly and is only detectable when packs are dissented indicating that contamination with Cl. estertheticum occurred pre-packing. Cl. estertheticum spores have been found on carcasses including primal cuts [53,70]. However, meat model experiments show that meat without artificially inoculated spores of Cl. estertheticum did not show signs of BPS during the entire duration of the trial 12–15 weeks [51,58,79]. Evidently, contamination of meat with Cl. estertheticum spores before packaging plays an important role in occurrence of BPS. These spores are usually transferred onto dressed carcasses during slaughter and processing [47,56]. The control of Cl. estertheticum must therefore be eliminated from the meat-processing environment, including stockyard pens, slaughter floor and soil, by means of extensive cleaning and sanitizing [59]. BPS in a meat plant can also be controlled by developing technologies to remove, kill, or inactivate spores of Cl. estertheticum present on dressed carcasses [58]. Given that Cl. estertheticum spores are resistant to heat, freezing, many chemicals and harsh environments [49], highly innovative and effective strategies would be required.
A comparison of three inactivation methods for Cl. estertheticum spores found that use of heat alone or ultrasound followed by heat treatment resulted in incomplete spore inactivation while peroxyacetic acid (POAA) sanitizer used with or without heat resulted in at least 4 log CFU per milliliter Cl. estertheticum spores at ambient temperature [49]. A similar effect of POAA on Cl. estertheticum was observed whereby BPS was delayed with a higher impact observed at −1.5 °C than 0 °C and 2 °C [58]. A POAA spray treatment on stainless steel coupons for 1 min resulted in a 2.3 log CFU per coupon reduction of Cl. estertheticum spores [84]. In the same experiment, a hydrogen peroxide vapor treatment for 150 min effectively inactivated Cl. estertheticum spores on the stainless-steel coupons [84]. On other hand, a hydrogen peroxide (15%) gel treatment for ten minutes on raw and pre-washed fleece resulted in a significant reduction of Cl. estertheticum spores [84]. Incubation of spores in meat juice at 80 °C for 10 min resulted in a 0.7 log CFU reduction in spore numbers, due to a likely loss of heat resistance of germinating spores [62].
In an in vitro experiment, Cl. estertheticum was inhibited by cultures of Lactococcus lactis, Lactobacillus sakei, Lactococcus garvieae and Leuconostoc carnosum but not by their cell-free fractions [73]. Similar results were reproduced in a meat model system, whereby BPS was delayed in meat inoculation with both Cl. estertheticum and L. lactis compared to inoculation with Cl. estertheticum only [51]. In an experiment to compare commercially available antimicrobials, Auranta FV (AFV; composed of bioflavonoids, citric, malic, lactic, and caprylic acids), Inbac-MDA (IMDA; composed of sodium diacetate, malic acid, mono and diglycerides of fatty acids, salt and excipients), and sodium octanoate (SO) incorporated in active packaging systems, found that AFV and SO prevented BPS within 42 days [85]. Times and temperature combinations of 240 s at 100 °C were also shown to inactivate Cl. estertheticum spores [49].
While the application of heat shrink after vacuum-packing may activate spores, the step is an invaluable processing step in meat processing. Initially, Bell et al. [80] recommended the application of other techniques besides heat shrink to achieve similar benefits. Later, several authors studied the practice in relation to BPS and recommended best practices for its application. Moschonas et al. [79] recommended to perform it at 50 °C for 15 s since this procedure reduced BPS compared to higher temperatures. On the other hand, Silva et al. [72] showed that a combination of high vacuum pressure (9 mbar) and shrinking temperature (87 °C) can retard BPS.
9. Conclusions
Blown pack spoilage is still a big challenge for the meat industry. Future studies improving the culture conditions of Cl. estertheticum and exploring the genetic characteristics of Cl. estertheticum might provide further insights into the mechanisms that can be explored to control the growth Cl. estertheticum in vacuum-packs, and hence reduce the economic burden associated with BPS.
Author Contributions
Conceptualization, R.S.; methodology, J.W. and R.S.; formal analysis, J.W.; resources, J.W. and R.S.; data curation, J.W.; writing-original draft preparation, J.W.; writing-review and editing, R.S.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Hilgarth, M.; Behr, J.; Vogel, R.F. Monitoring of spoilage-associated microbiota on modified atmosphere packaged beef and differentiation of psychrophilic and psychrotrophic strains. J. Appl. Microbiol. 2018, 124, 740–753. [Google Scholar] [CrossRef] [PubMed]
- Höll, L.; Behr, J.; Vogel, R.F. Identification and growth dynamics of meat spoilage microorganisms in modified atmosphere packaged poultry meat by MALDI-TOF MS. Food Microbiol. 2016, 60, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Lahmar, A.; Morcuende, D.; Andrade, M.-J.; Chekir-Ghedira, L.; Estévez, M. Prolonging shelf life of lamb cutlets packed under high-oxygen modified atmosphere by spraying essential oils from North-African plants. Meat Sci. 2018, 139, 56–64. [Google Scholar] [CrossRef] [PubMed]
- Mikš-Krajnik, M.; Yoon, Y.-J.; Ukuku, D.O.; Yuk, H.-G. Volatile chemical spoilage indexes of raw atlantic salmon (Salmo salar) stored under aerobic condition in relation to microbiological and sensory shelf lives. Food Microbiol. 2016, 53, 182–191. [Google Scholar] [CrossRef]
- Cheng, W.; Sun, D.-W.; Pu, H.; Wei, Q. Chemical spoilage extent traceability of two kinds of processed pork meats using one multispectral system developed by hyperspectral imaging combined with effective variable selection methods. Food Chem. 2017, 221, 1989–1996. [Google Scholar] [CrossRef]
- Holman, B.W.B.; Kerry, J.P.; Hopkins, D.L. Meat packaging solutions to current industry challenges: A review. Meat Sci. 2018, 144, 159–168. [Google Scholar] [CrossRef]
- Nychas, G.-J.E.; Skandamis, P.N.; Tassou, C.C.; Koutsoumanis, K.P. Meat spoilage during distribution. Meat Sci. 2008, 78, 77–89. [Google Scholar] [CrossRef]
- Stellato, G.; La Storia, A.; De Filippis, F.; Borriello, G.; Villani, F.; Ercolini, D. Overlap of spoilage-associated microbiota between meat and the meat processing environment in small-scale and large-scale retail distributions. Appl. Environ. Microbiol. 2016, 82, 4045–4054. [Google Scholar] [CrossRef]
- Wambui, J.; Lamuka, P.; Karuri, E.; Matofari, J.; Njage, P.M.K. Microbial contamination level profiles attributed to contamination of beef carcasses, personnel, and equipment: Case of small and medium enterprise slaughterhouses. J. Food Prot. 2018, 81, 684–691. [Google Scholar] [CrossRef]
- Fletcher, B.; Mullane, K.; Platts, P.; Todd, E.; Power, A.; Roberts, J.; Chapman, J.; Cozzolino, D.; Chandra, S. Advances in meat spoilage detection: A short focus on rapid methods and technologies. CyTA J. Food 2018, 16, 1037–1044. [Google Scholar] [CrossRef]
- Hilgarth, M.; Nani, M.; Vogel, R.F. Assertiveness of meat-borne Lactococcus piscium strains and their potential for competitive exclusion of spoilage bacteria in situ and in vitro. J. Appl. Microbiol. 2018, 124, 1243–1253. [Google Scholar] [CrossRef]
- Gram, L.; Ravn, L.; Rasch, M.; Bruhn, J.B.; Christensen, A.B.; Givskov, M. Food spoilage-interactions between food spoilage bacteria. Int. J. Food Microbiol. 2002, 78, 79–97. [Google Scholar] [CrossRef]
- Andreevskaya, M.; Jääskeläinen, E.; Johansson, P.; Ylinen, A.; Paulin, L.; Björkroth, J.; Auvinen, P. Food spoilage-associated Leuconostoc, Lactococcus, and Lactobacillus species display different survival strategies in response to competition. Appl. Environ. Microbiol. 2018, 84, e00554-18. [Google Scholar] [CrossRef] [PubMed]
- Casaburi, A.; Piombino, P.; Nychas, G.-J.; Villani, F.; Ercolini, D. Bacterial populations and the volatilome associated to meat spoilage. Food Microbiol. 2015, 45, 83–102. [Google Scholar] [CrossRef] [PubMed]
- Van Rooyen, L.A.; Allen, P.; Kelly-Rees, C.; O’Connor, D.I. The Effects of varying gas concentrations and exposure times on colour stability and shelf-life of vacuum packaged beef steaks subjected to carbon monoxide pretreatment. Food Packag. Shelf Life 2018, 18, 230–237. [Google Scholar] [CrossRef]
- De Filippis, F.; La Storia, A.; Villani, F.; Ercolini, D. Strain-level diversity analysis of Pseudomonas fragi after in situ pangenome reconstruction shows distinctive spoilage-associated metabolic traits clearly selected by different storage conditions. Appl. Environ. Microbiol. 2019, 85, e02212-18. [Google Scholar] [CrossRef] [PubMed]
- Lavieri, N.; Williams, S.K. Effects of packaging systems and fat concentrations on microbiology, sensory and physical properties of ground beef stored at 4 ± 1 °C for 25 days. Meat Sci. 2014, 97, 534–541. [Google Scholar] [CrossRef] [PubMed]
- Mansur, A.R.; Song, E.-J.; Cho, Y.-S.; Nam, Y.-D.; Choi, Y.-S.; Kim, D.-O.; Seo, D.-H.; Nam, T.G. Comparative evaluation of spoilage-related bacterial diversity and metabolite profiles in chilled beef stored under air and vacuum packaging. Food Microbiol. 2019, 77, 166–172. [Google Scholar] [CrossRef]
- Doulgeraki, A.I.; Ercolini, D.; Villani, F.; Nychas, G.-J.E. Spoilage microbiota associated to the storage of raw meat in different conditions. Int. J. Food Microbiol. 2012, 157, 130–141. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zhang, Y.; Yang, X.; Hopkins, D.L.; Zhu, L.; Dong, P.; Liang, R.; Luo, X. Shelf-life and microbial community dynamics of super-chilled beef imported from Australia to China. Food Res. Int. 2019, 120, 784–792. [Google Scholar] [CrossRef]
- Pothakos, V.; Devlieghere, F.; Villani, F.; Björkroth, J.; Ercolini, D. Lactic Acid Bacteria and their controversial role in fresh meat spoilage. Meat Sci. 2015, 109, 66–74. [Google Scholar] [CrossRef]
- Zhang, P.; Badoni, M.; Gänzle, M.; Yang, X. Growth of Carnobacterium spp. isolated from chilled vacuum-packaged meat under relevant acidic conditions. Int. J. Food Microbiol. 2018, 286, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Chaillou, S.; Chaulot-Talmon, A.; Caekebeke, H.; Cardinal, M.; Christieans, S.; Denis, C.; Hélène Desmonts, M.; Dousset, X.; Feurer, C.; Hamon, E.; et al. Origin and ecological selection of core and food-specific bacterial communities associated with meat and seafood spoilage. ISME J. 2015, 9, 1105–1118. [Google Scholar] [CrossRef]
- Hultman, J.; Rahkila, R.; Ali, J.; Rousu, J.; Björkroth, K.J. Meat processing plant microbiome and contamination patterns of cold-tolerant bacteria causing food safety and spoilage risks in the manufacture of vacuum-packaged cooked sausages. Appl. Environ. Microbiol. 2015, 81, 7088–7097. [Google Scholar] [CrossRef] [PubMed]
- Dorn-In, S.; Schwaiger, K.; Springer, C.; Barta, L.; Ulrich, S.; Gareis, M. Development of a Multiplex QPCR for the species identification of Clostridium estertheticum, C. frigoriphilum, C. bowmanii and C. tagluense-like from blown pack spoilage (BPS) meats and from wild boars. Int. J. Food Microbiol. 2018, 286, 162–169. [Google Scholar] [CrossRef] [PubMed]
- André, S.; Vallaeys, T.; Planchon, S. Spore-forming bacteria responsible for food spoilage. Res. Microbiol. 2017, 168, 379–387. [Google Scholar] [CrossRef]
- Zhang, Y.; Wei, J.; Yuan, Y.; Yue, T. Diversity and characterization of spoilage-associated psychrotrophs in food in cold chain. Int. J. Food Microbiol. 2019, 290, 86–95. [Google Scholar] [CrossRef] [PubMed]
- Brightwell, G.; Clemens, R. Development and validation of a Real-Time PCR assay specific for Clostridium estertheticum and C. estertheticum-like psychrotolerant bacteria. Meat Sci. 2012, 92, 697–703. [Google Scholar] [CrossRef] [PubMed]
- Collins, M.D.; Lawson, P.A.; Willems, A.; Cordoba, J.J.; Fernandez-Garayzabal, J.; Garcia, P.; Cai, J.; Hippe, H.; Farrow, J.A.E. The phylogeny of the genus Clostridium: Proposal of five new genera and eleven new species combinations. Int. J. Syst. Bacteriol. 1994, 44, 812–826. [Google Scholar] [CrossRef] [PubMed]
- Vos, P.; Garrity, G.M.; Jones, D.; Krieg, N.R.; Ludwig, W.; Rainey, F.A.; Schleifer, K.-H. Bergey’s Manual of Systematic Bacteriology, Volume 3: The Firmicutes; Springer: Berlin/Heidelberg, Germany, 2012; p. 1450. [Google Scholar]
- Moschonas, G.; Bolton, D.J.; Sheridan, J.J.; McDowell, D.A. The effect of storage temperature and inoculum level on the time of onset of “blown pack” spoilage. J. Appl. Microbiol. 2010, 108, 532–539. [Google Scholar] [CrossRef]
- Dainty, R.H.; Edwards, R.A.; Hibbard, C.M. Spoilage of vacuum-packed beef by Aclostridium sp. J. Sci. Food Agric. 1989, 49, 473–486. [Google Scholar] [CrossRef]
- Kalchayanand, N.; Ray, B.; Field, R.A.; Johnson, M.C. Spoilage of vacuum-packaged refrigerated beef by Clostridium. J. Food Prot. 1989, 52, 424–426. [Google Scholar] [CrossRef] [PubMed]
- Broda, D.M.; Musgrave, D.R.; Bell, R.G. Use of Restriction Fragment Length Polymorphism analysis to differentiate strains of psychrophilic and psychrotrophic Clostridia associated with “blown pack” spoilage of vacuum-packed meats. J. Appl. Microbiol. 2000, 88, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Byrne, B.; Monaghan, A.M.; Lyng, J.G.; Sheridan, J.J.; Bolton, D.J. A case of “blown pack” meat linked to Clostridium estertheticum in Ireland. J. Food Saf. 2009, 29, 629–635. [Google Scholar] [CrossRef]
- Joseph, R.C.; Kim, N.M.; Sandoval, N.R. Recent developments of the synthetic biology toolkit for Clostridium. Front. Microbiol. 2018, 9, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Prazmowski, A. Untersuchung über die Entwickelungsgeschichte und Fermentwirking einiger Bakterien-arten. Inaugural Dissertation, Hugo Voigt, Leipzig, Germany, 1880. [Google Scholar]
- Alou, M.T.; Ndongo, S.; Frégère, L.; Labas, N.; Andrieu, C.; Richez, M.; Couderc, C.; Baudoin, J.P.; Abrahão, J.; Brah, S.; et al. Taxonogenomic description of four new Clostridium species isolated from human gut: ‘Clostridium amazonitimonense’, ‘Clostridium merdae’, ‘Clostridium massilidielmoense’ and ‘Clostridium nigeriense’. New Microbes New Infect. 2018, 21, 128–139. [Google Scholar] [CrossRef]
- Lawson, P.A.; Rainey, F.A. Proposal to restrict the Genus Clostridium prazmowski to Clostridium butyricum and related species. Int. J. Syst. Evol. Microbiol. 2016, 66, 1009–1016. [Google Scholar] [CrossRef]
- Oren, A.; Rupnik, M. Clostridium difficile and Clostridioides difficile: Two validly published and correct names. Anaerobe 2018, 52, 125–126. [Google Scholar] [CrossRef]
- Collins, M.D. Taxonomic studies on a psychrophilic Clostridium from vacuum-packed beef: Description of Clostridium estertheticum sp. nov. FEMS Microbiol. Lett. 1992, 96, 235–239. [Google Scholar] [CrossRef]
- Kalchayanand, N.; Ray, B.; Field, R.A. Characteristics of psychrotrophic Clostridium laramie causing spoilage of vacuum-packaged refrigerated fresh and roasted beef. J. Food Prot. 1993, 56, 13–17. [Google Scholar] [CrossRef]
- Spring, S. Characterization of novel psychrophilic Clostridia from an antarctic microbial mat: Description of Clostridium frigoris sp. nov., Clostridium lacusfryxellense sp. nov., Clostridium bowmanii sp. nov. and Clostridium psychrophilum sp. nov. and reclassification of Clostridium laramiense as Clostridium estertheticum subsp. laramiense subsp. nov. Int. J. Syst. Evol. Microbiol. 2003, 53, 1019–1029. [Google Scholar]
- Yu, Z.; Gunn, L.; Brennan, E.; Reid, R.; Wall, P.G.; Gaora, P.; Hurley, D.; Bolton, D.; Fanning, S. Complete genome sequence of Clostridium estertheticum DSM 8809, a microbe identified in spoiled vacuum-packed beef. Front. Microbiol. 2016, 7, 1–10. [Google Scholar] [CrossRef]
- Yang, X.; Gill, C.O.; Balamurugan, S. Products of glucose and lactate fermentation, and utilization of amino acids by Clostridium estertheticum ssp. laramiense and estertheticum growing in meat juice medium. J. Food Prot. 2016, 73, 1348–1352. [Google Scholar]
- Broda, D.M.; Delacy, K.M.; Bell, R.G.; Braggins, T.J.; Cook, R.L. Psychrotrophic Clostridium spp. associated with ‘blown pack’ spoilage of chilled vacuum-packed red meats and dog rolls in gas-impermeable plastic casings. Int. J. Food Microbiol. 1996, 29, 335–352. [Google Scholar] [CrossRef]
- Boerema, J.A.; Broda, D.M.; Bell, R.G. Abattoir sources of psychrophilic Clostridia causing blown pack spoilage of vacuum-packed chilled meats determined by culture-based and molecular detection procedures. Lett. Appl. Microbiol. 2003, 36, 406–411. [Google Scholar] [CrossRef]
- Moschonas, G.; Bolton, D.J.; Sheridan, J.J.; McDowell, D.A. Isolation and sources of blown pack spoilage Clostridia in beef abattoirs. J. Appl. Microbiol. 2009, 107, 616–624. [Google Scholar] [CrossRef]
- Broda, D.M. The Effect of peroxyacetic acid-based sanitizer, heat and ultrasonic waves on the survival of Clostridium estertheticum spores in vitro. Lett. Appl. Microbiol. 2007, 45, 336–341. [Google Scholar] [CrossRef]
- Bonke, R.; Drees, N.; Gareis, M. Detection of psychrophilic and psychrotolerant Clostridium spp. in chilled fresh vacuum-packed meat using different PCR methods. FEMS Microbiol. Lett. 2015, 363, fnv218. [Google Scholar] [CrossRef]
- Jones, R.J.; Zagorec, M.; Brightwell, G.; Tagg, J.R. Inhibition by Lactobacillus sakei of other species in the flora of vacuum packaged raw meats during prolonged storage. Food Microbiol. 2009, 26, 876–881. [Google Scholar] [CrossRef]
- Moschonas, G.; Bolton, D.J.; McDowell, D.A.; Sheridan, J.J. Diversity of culturable psychrophilic and psychrotrophic anaerobic bacteria isolated from beef abattoirs and their environments. Appl. Environ. Microbiol. 2011, 77, 4280–4284. [Google Scholar] [CrossRef]
- Bakhtiary, F.; Sayevand, H.R.; Remely, M.; Hippe, B.; Indra, A.; Hosseini, H.; Haslberger, A.G. Identification of Clostridium spp. derived from a sheep and cattle slaughterhouse by Matrix-Assisted Laser Desorption and Ionization-Time of Flight Mass Spectrometry (MALDI-TOF MS) and 16S RDNA sequencing. J. Food Sci. Technol. 2018, 55, 3232–3240. [Google Scholar] [CrossRef]
- Yang, X.; Gill, C.O.; Balamurugan, S. Enumeration of Clostridium estertheticum spores in samples from meat plant conveyors and silage stacks by conventional and Real-Time PCR procedures. Internet J. Food Saf. 2010, 12, 115–121. [Google Scholar]
- Helps, C.R.; Harbour, D.A.; Corry, J.E.L. PCR-based 16S ribosomal DNA detection technique for Clostridium estertheticum causing spoilage in vacuum-packed chill-stored beef. Int. J. Food Microbiol. 1999, 52, 57–65. [Google Scholar] [CrossRef]
- Broda, D.M.; Bell, R.G.; Boerema, J.A.; Musgrave, D.R. The abattoir source of culturable psychrophilic Clostridium spp. causing “blown pack” spoilage of vacuum-packed chilled venison. J. Appl. Microbiol. 2002, 93, 817–824. [Google Scholar] [CrossRef]
- Broda, D.M.; Boerema, J.A.; Bell, R.G. PCR Detection of psychrophilic Clostridium spp. causing “blown pack” spoilage of vacuum-packed chilled meats. J. Appl. Microbiol. 2003, 94, 515–522. [Google Scholar] [CrossRef]
- Boerema, J.A.; Broda, D.M.; Penney, N.; Brightwell, G. Influence of peroxyacetic acid–based carcass rinse on the onset of “blown pack” spoilage in artificially inoculated vacuum-packed chilled beef. J. Food Prot. 2007, 70, 1434–1439. [Google Scholar] [CrossRef]
- Brightwell, G.; Broda, D.M.; Boerema, J.A. Sources of psychrophilic and psychrotolerant Clostridia causing spoilage of vacuum-packed chilled meats, as determined by PCR amplification procedure. J. Appl. Microbiol. 2009, 107, 178–186. [Google Scholar]
- Reid, R.; Fanning, S.; Whyte, P.; Kerry, J.; Bolton, D. Comparison of hot versus cold boning of beef carcasses on bacterial growth and the risk of blown pack spoilage. Meat Sci. 2017, 125, 46–52. [Google Scholar] [CrossRef]
- Brightwell, G.; Horváth, K.M. Molecular discrimination of New Zealand sourced meat spoilage associated psychrotolerant Clostridium species by ARDRA and its comparison with 16s RNA gene sequencing. Meat Sci. 2018, 138, 23–27. [Google Scholar] [CrossRef]
- Rajagopal, S.; McMullen, L.M.; Gill, C.O.; Yang, X. Characterization of germination of spores of Clostridium estertheticum, the primary causative agent of blown pack spoilage of vacuum packaged beef. Food Res. Int. 2016, 87, 109–114. [Google Scholar] [CrossRef]
- Yang, X.; Gill, C.O.; Balamurugan, S. Effects of temperature and pH on the growth of bacteria isolated from blown packs of vacuum-packaged beef. J. Food Prot. 2009, 72, 2380–2385. [Google Scholar] [CrossRef]
- Clemens, R.M.; Adam, K.H.; Brightwell, G. Contamination levels of Clostridium estertheticum spores that result in gaseous spoilage of vacuum-packaged chilled beef and lamb meat. Lett. Appl. Microbiol. 2010, 50, 591–596. [Google Scholar] [CrossRef]
- Yang, X.; Balamurugan, S.; Gill, C.O. Substrate utilization by Clostridium estertheticum cultivated in meat juice medium. Int. J. Food Microbiol. 2009, 128, 501–505. [Google Scholar] [CrossRef]
- Yang, X.; Wang, H.; Badoni, M. Effects of meat pH and the initial numbers of spores of Clostridium estertheticum on the development of blown pack spoilage of vacuum-packaged beef. Int. J. Food Sci. Technol. 2014, 49, 1619–1625. [Google Scholar] [CrossRef]
- Brightwell, G.; Clemens, R.; Urlich, S.; Boerema, J. Possible involvement of psychrotolerant Enterobacteriaceae in blown pack spoilage of vacuum-packaged raw meats. Int. J. Food Microbiol. 2007, 119, 334–339. [Google Scholar] [CrossRef] [PubMed]
- Kang, D.-H.; Arthur, T.M.; Siragusa, G.R. Gas formation in ground beef chubs due to Hafnia alvei is reduced by multiple applications of antimicrobial interventions to artificially inoculated beef trim stock. J. Food Prot. 2002, 65, 1651–1655. [Google Scholar] [CrossRef]
- Chaves, R.D.; Silva, A.R.; Sant’Ana, A.S.; Campana, F.B.; Massaguer, P.R. Gas-producing and spoilage potential of Enterobacteriaceae and Lactic Acid Bacteria isolated from chilled vacuum-packaged beef. Int. J. Food Sci. Technol. 2012, 47, 1750–1756. [Google Scholar] [CrossRef]
- Bolton, D.J.; Carroll, J.; Walsh, D. A four-year survey of blown pack spoilage Clostridium estertheticum and Clostridium gasigenes on beef primal cuts. Lett. Appl. Microbiol. 2015, 61, 153–157. [Google Scholar] [CrossRef]
- Yang, X.; Youssef, M.K.; Gill, C.O.; Badoni, M.; López-Campos, Ó. Effects of meat pH on growth of 11 species of psychrotolerant Clostridia on vacuum packaged beef and blown pack spoilage of the product. Food Microbiol. 2014, 39, 13–18. [Google Scholar] [CrossRef]
- Silva, A.R.; Tahara, A.C.C.; Chaves, R.D.; Sant’Ana, A.S.; Faria, J.d.A.F.; Massaguer, P.R. Influence of different shrinking temperatures and vacuum conditions on the ability of psychrotrophic Clostridium to cause “blown pack” spoilage in chilled vacuum-packaged beef. Meat Sci. 2012, 92, 498–505. [Google Scholar] [CrossRef]
- Jones, R.J.; Hussein, H.M.; Zagorec, M.; Brightwell, G.; Tagg, J.R. Isolation of Lactic Acid Bacteria with inhibitory activity against pathogens and spoilage organisms associated with fresh meat. Food Microbiol. 2008, 25, 228–234. [Google Scholar] [CrossRef] [PubMed]
- Silva, A.R.; Carvalho, J.; Massaguer, P.R. “Blown pack” probabilistic modeling for C. algidicarnis and C. estertheticum under the effects of storage temperature, vacuum level and package shrink temperature. Procedia Food Sci. 2016, 7, 59–62. [Google Scholar] [CrossRef]
- Reid, R.; Fanning, S.; Whyte, P.; Kerry, J.; Bolton, D. An investigation of the effect of rapid slurry chilling on blown pack spoilage of vacuum-packaged beef primals. Lett. Appl. Microbiol. 2017, 64, 177–181. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Balamurugan, S.; Gill, C.O. Effects on the development of blown pack spoilage of the initial numbers of Clostridium estertheticum spores and Leuconostoc mesenteroides on vacuum packed beef. Meat Sci. 2011, 88, 361–367. [Google Scholar] [CrossRef]
- Mills, J.; Donnison, A.; Brightwell, G. Factors affecting microbial spoilage and shelf-life of chilled vacuum-packed lamb transported to distant markets: A review. Meat Sci. 2014, 98, 71–80. [Google Scholar] [CrossRef]
- James, S.J.; James, C. Meat Refrigeration; CRC Press: Boca Raton, FL, USA, 2002; p. 360. [Google Scholar]
- Moschonas, G.; Bolton, D.J.; Sheridan, J.J.; McDowell, D.A. The effect of heat shrink treatment and storage temperature on the time of onset of “blown pack” spoilage. Meat Sci. 2011, 87, 115–118. [Google Scholar] [CrossRef]
- Bell, R.G.; Moorhead, S.M.; Broda, D.M. Influence of heat shrink treatments on the onset of Clostridial “blown pack” spoilage of vacuum-packed chilled meat. Food Res. Int. 2001, 34, 271–275. [Google Scholar] [CrossRef]
- Sumner, J.; Jenson, I.; Ross, T. Using predictive microbiology to benefit the Australian meat industry. In Case Studies in Food Safety and Authenticity; Elsevier: Amsterdam, The Netherlands, 2012; pp. 276–283. [Google Scholar]
- Claus, J.R.; Sørheim, O. Preserving pre-rigor meat functionality for beef patty production. Meat Sci. 2006, 73, 287–294. [Google Scholar] [CrossRef]
- Sukumaran, A.T.; Holtcamp, A.J.; Campbell, Y.L.; Burnett, D.; Schilling, M.W.; Dinh, T.T.N. Technological characteristics of pre- and post-rigor deboned beef mixtures from Holstein steers and quality attributes of cooked beef sausage. Meat Sci. 2018, 145, 71–78. [Google Scholar] [CrossRef]
- Mills, J.; Horváth, K.M.; Brightwell, G. Antimicrobial effect of different peroxyacetic acid and hydrogen peroxide formats against spores of Clostridium estertheticum. Meat Sci. 2018, 143, 69–73. [Google Scholar] [CrossRef]
- Reid, R.; Bolton, D.; Tiuftin, A.; Kerry, J.; Fanning, S.; Whyte, P. Controlling blown pack spoilage using anti-microbial packaging. Foods 2017, 6, 67. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
Table 1.
Composition of peptone yeast extract glucose starch.
| Substance | g/L or mL/L |
|---|---|
| Proteose Peptone | 5 |
| Tryptone | 5 |
| Yeast extract | 10 |
| Meat extract Powder | 10 |
| Glucose | 2 |
| Soluble starch | 1 |
| Resazurin | 0.001 |
| Cysteine HCl | 0.2 |
| Solution of Silicon Antifoaming Agent 20% | 0.25 |
| Salts Solution A | 40 |
| Salts Solution B | 40 |
| Salts Solution A | |
| CaCl2·2H2O | 0.265 |
| MgSO4·7H2O | 0.48 |
| NaCl | 2 |
| Salts Solution B | |
| KH2PO4 | 1 |
| K2HPO4·3H2O | 1.3 |
| NaHCO3 | 10 |
Table 2.
Primers and probes used to detect Cl. estertheticum.
| Assay | Primer and Probe | Sequence | Reference | |
|---|---|---|---|---|
| PCR | Primer | 16SEF | 5′-TCG GAA TTT CAC TTT GAG-3′ | [57] |
| 16SER | 5′-AAG GAC TTC ACT CAT CTC TG-3′ | |||
| RT-PCR | Primer | TMF | 5′-CGG CGG ACG GGT GAG TAA C-3′ | [28] |
| TMR | 5′-CGG GTC CAT CTC AAA GTG RAA CT-3′ | |||
| Probe | 5′-FAM-CGT GGG TAA CCT GCC TCA AAG AGG GG-TAMRA-3′ | |||
| qPCR | Primer | TMF | 5′-CGGCGGACGGGTGAGTAAC-3′ | [25] |
| Cl642-R | 5′-CCTCTCCTGCACTCTAGA-3′ | |||
| Probe | Cest | 5′-HEX-CAAAGGAATTTTTCGGAATTTCACTTTGAG-BHQ1-3′ | ||
PCR: Polymerase chain reaction; RT-PCR: Real time PCR; and qPCR; Quantitative PCR.
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Wambui, J.; Stephan, R. Relevant Aspects of Clostridium estertheticum as a Specific Spoilage Organism of Vacuum-Packed Meat. Microorganisms 2019, 7, 142. https://doi.org/10.3390/microorganisms7050142
AMA Style
Wambui J, Stephan R. Relevant Aspects of Clostridium estertheticum as a Specific Spoilage Organism of Vacuum-Packed Meat. Microorganisms. 2019; 7(5):142. https://doi.org/10.3390/microorganisms7050142
Chicago/Turabian StyleWambui, Joseph, and Roger Stephan. 2019. "Relevant Aspects of Clostridium estertheticum as a Specific Spoilage Organism of Vacuum-Packed Meat" Microorganisms 7, no. 5: 142. https://doi.org/10.3390/microorganisms7050142
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.