
Staphylococcus aureus in Animals and Food: Methicillin Resistance, Prevalence and Population Structure. A Review in the African Continent


Abstract
:
1. Introduction
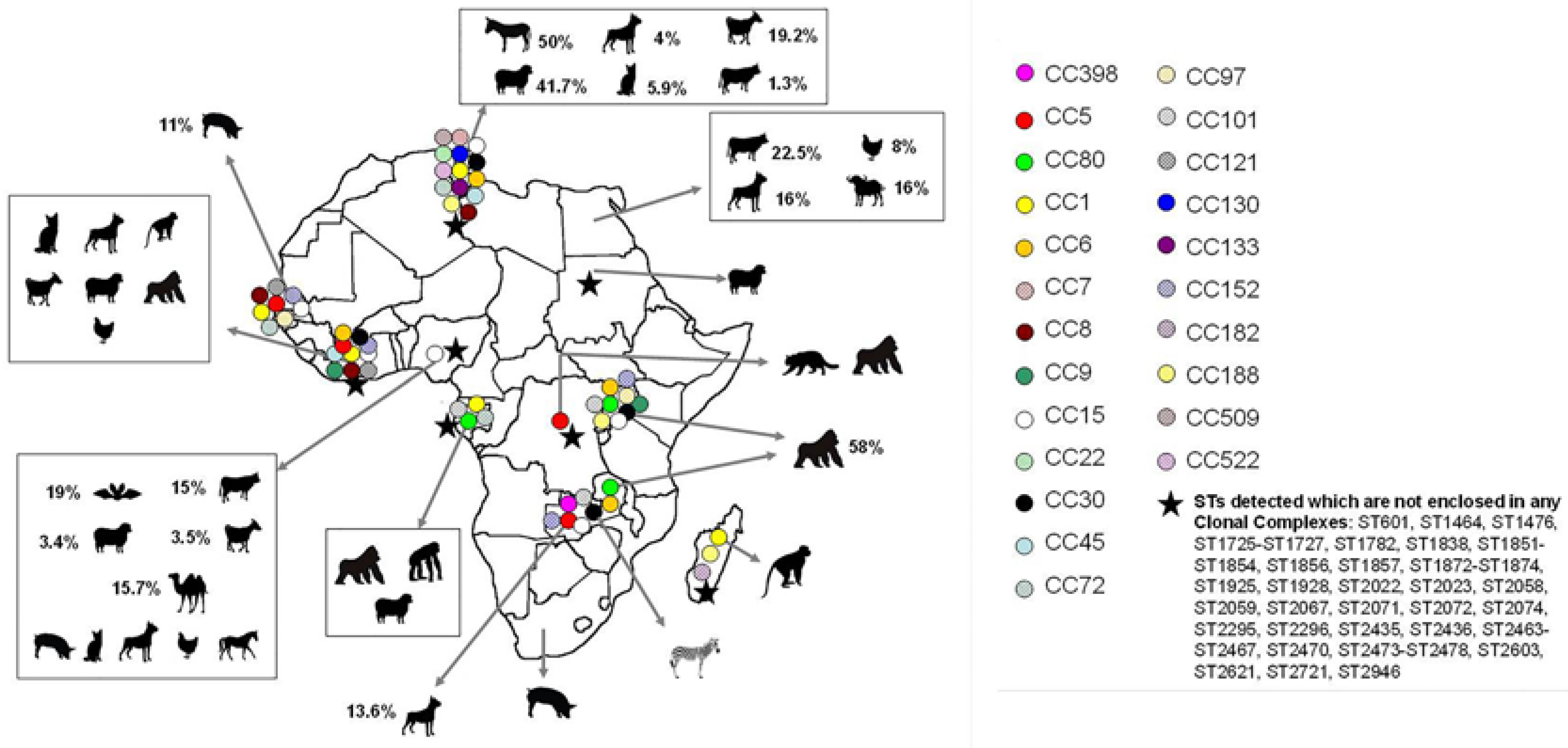
2. S. aureus in Animals in Africa
2.1. S. aureus Prevalence in Animals

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Country | Tested Animals | Animals from Which S. aureus Was Detected | Detection of MRSA | Animals from Which MRSA Was Detected | Sampling Date | Healthy/Sick | Samples | Lineages of MRSA | Lineages of MSSA b | Reference |
|---|---|---|---|---|---|---|---|---|---|---|
| Côte d’Ivoire | Domestic and wild animals | Goats, cats, dogs, sheep, poultry, primates | yes | Sheep | 2010–2013 | Healthy | Nasal and pharyngeal | CC88 | CC5, CC6, CC8, CC15, CC121, CC152, ST567, ST1472, ST2946, among others | [17] |
| Primates | Chimpanzees | no | - | 2007–2012 | Healthy | Mucosal, feces, oral, genital, fruit wedges | - | CC1, CC45, ST601, ST1928, ST2603, ST2621 | [18] | |
| Primates | Monkeys | no | - | - | Healthy | Nasal and fruit wedges | - | CC1, CC9, CC45, ST601, ST1782, ST1928, ST2023, ST2058, ST2059, ST2072, ST2603, ST2621 | [19] | |
| Democratic Republic of Congo | Domestic and wild animals | Civet, primates | no | - | 2010–2013 | Healthy | Nasal and pharyngeal | - | CC5, ST2473-ST2478, among others | [17] |
| Egypt | Dogs and cats | Dogs | yes | Dogs | - | Healthy and sick | Nasal, oral, ear, wound | HA-MRSA and CA-MRSA | - | [20] |
| Cattle, dogs, buffaloes, poultry | Cattle, dogs, buffaloes, poultry | yes | No specified | - | Sick | Milk, wounds, abscesses, internal organs, urine and nasal | ND a | - | [21] | |
| Gabon | Primates | Gorillas, chimpanzees | no | - | 2011 | Healthy and sick | Nasal, oral, vaginal, rectal | - | CC72, CC101 | [22] |
| Primates | Monkeys, gorillas, chimpanzees | no | - | - | Healthy | Nasal and fruit wedges | - | CC1, CC80, ST1851-ST1854, ST1856, ST1857, ST1872 ST1874, ST1928, ST2022, ST2023, ST2067, ST2071, ST2074 | [19] | |
| Domestic and wild animals | Sheep, primates | no | - | 2010–2013 | Healthy | Nasal and pharyngeal | - | CC101, CC80, ST1838, ST1851-ST1854, ST1857, ST1872-ST1874, ST1925, ST2022, ST2067, ST2071, ST2074, ST2295, ST2296, ST2721, among others | [17] | |
| Madagascar | Primates | Lemurs | no | - | 2007–2012 | Healthy | Mucosal, feces, oral, genital, fruit wedges | - | CC1, CC182, CC188, ST2435, ST2436 | [18] |
| Nigeria | Dogs, cats, chickens, pigs, horses, sheep, cattle, goats | Dogs, cats, chickens, pigs, horses, sheep, cattle, goats | - | - | - | Healthy and sick | Skin lesions, nasal, cloacal, milk | - | - | [23] |
| Bats | Bats | no | - | 2008–2010 | Healthy | Fecal | - | CC15, ST1725-ST1727, ST2463-ST2467, ST2470 | [24] | |
| Camels, sheep, cattle, goats | Camels, sheep, cattle, goats | yes | Camels, sheep, cattle, goats | 2012 | Healthy | Nasal and milk | ND | - | [25] | |
| Chickens | Chickens | no | - | - | Healthy | - | - | - | [26] | |
| Senegal | Pigs | Pigs | yes | Pigs | 2009–2011 | Healthy | Nasal | CC5, CC88 | CC1, CC5, CC8, CC15, CC72, CC97, CC121, CC152 | [27] |
| South Africa | Pigs, cattle, goats, chickens | Pigs | yes | Pigs | - | Healthy | Nasal, mouth wash, ear | ND | - | [28] |
| Chimpanzees | Chimpanzees | no | - | 2007, 2010, 2011 | Healthy | Nasal and oral | - | CC15, CC6, CC30, CC80, CC101 | [29] | |
| Cattle and pigs | Cattle and pigs | yes | Cattle and pigs | - | Healthy | Rump, flank, brisket, neck | ND | - | [30] | |
| Sudan | Sheep | Sheep | no | - | 2007–2008 | Sick | Abscesses | - | ST1464 | [31] |
| Sheep | Sheep | no | - | 2003–2005 | Sick | Pus samples | - | - | [32] | |
| Horse | Horse | yes | Horse | - | Sick | Lungs and peritoneum | ND | - | [33] | |
| Tunisia | Sheep | Sheep | yes | Sheep | 2010 | Healthy | Nasal | CC80 | CC8, CC130, CC522, ST1476, ST2076 | [34] |
| Donkeys | Donkeys | no | - | 2010 | Healthy | Nasal | - | CC1, CC6, CC7, CC15, CC22, CC72, CC133, CC522 | [35] | |
| Cattle, goats, dogs, cats | Cattle, goats, dogs, cats | no | - | 2010–2011 | Healthy | Nasal | - | CC6, CC15, CC30, CC45, CC130, CC133, CC188, CC522 | [36] | |
| Uganda | Chimpanzees | Chimpanzees | no | - | 2007, 2010, 2011 | Healthy | Nasal and oral | - | CC15, CC6, CC30, CC80, CC101 | [29] |
| Primates | Chimpanzees | no | - | 2007–2012 | Healthy | Mucosal, feces, oral, genital, fruit wedges | - | CC6, CC9, CC15, CC30, CC80, CC152, CC188 | [18] | |
| Zambia | Zebra | Zebra | - | - | Sick | Tissue | - | - | [37] | |
| Chimpanzees | Chimpanzees | no | - | 2007, 2010, 2011 | Healthy | Nasal and oral | - | CC15, CC6, CC30, CC80, CC101 | [29] | |
| Dogs and cats | Dogs | no | - | 2012 | Sick | Skin, ear, wound | - | CC398, CC5, CC15, CC152 | [38] |
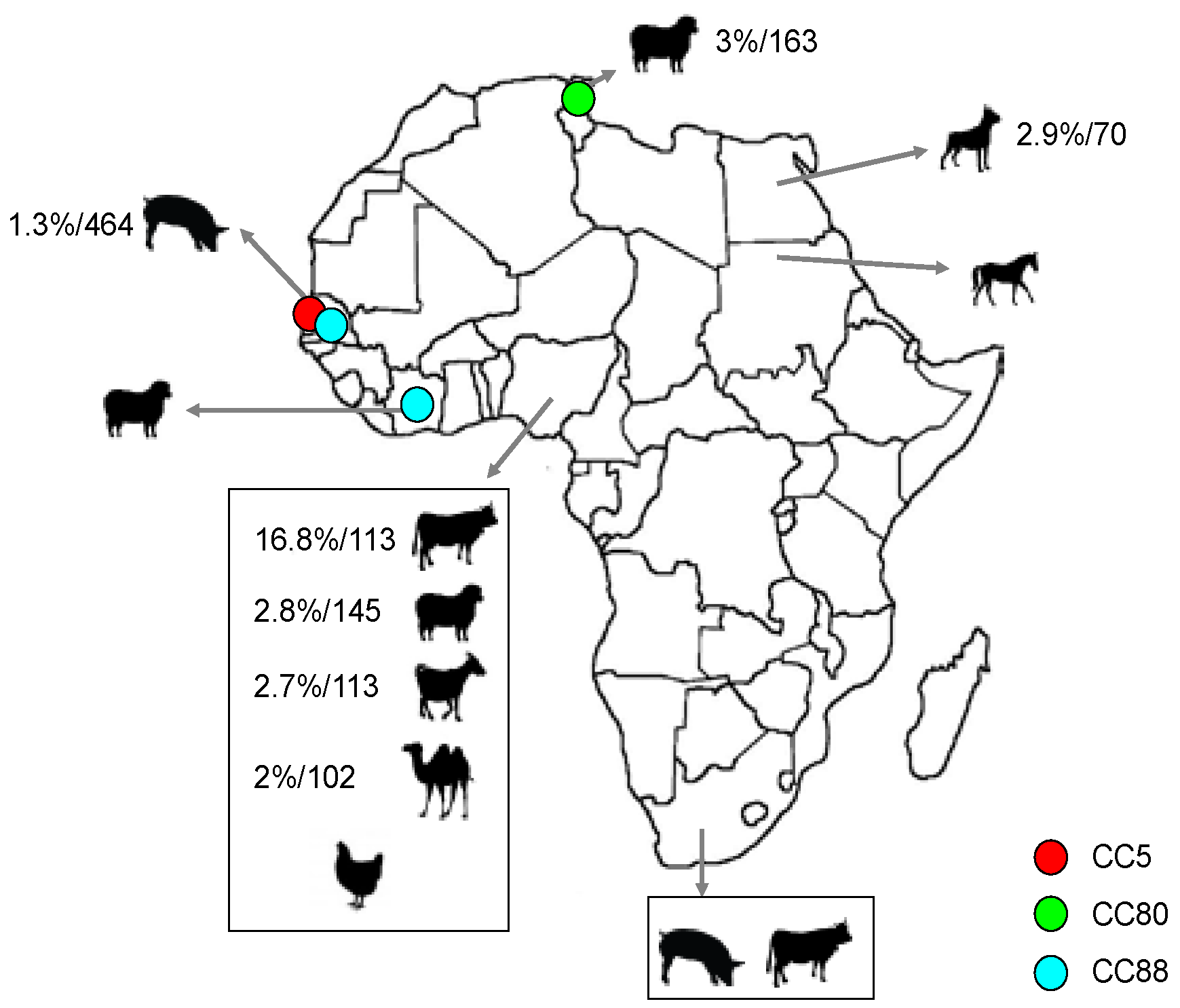
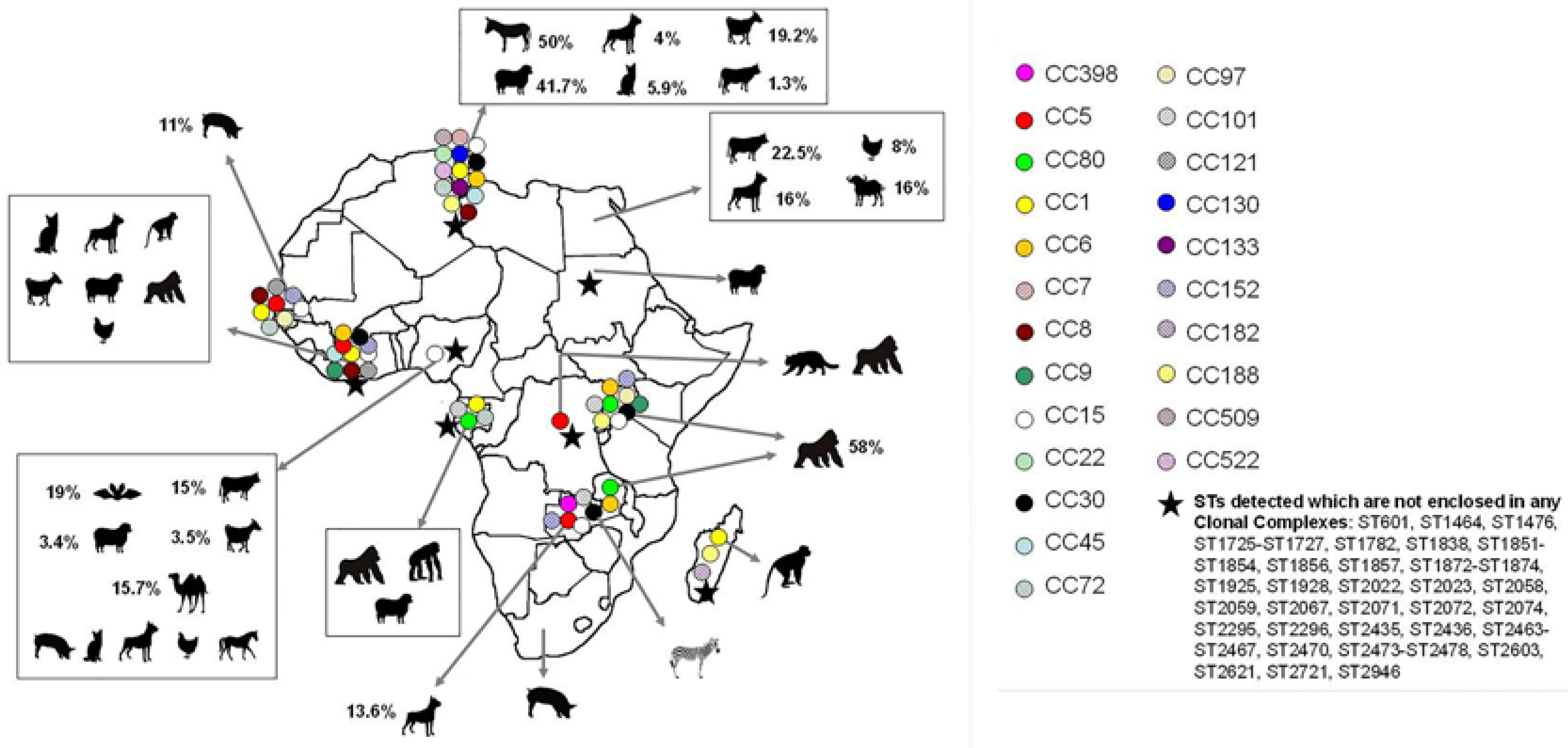
2.2. Population Structure of MSSA in Animals
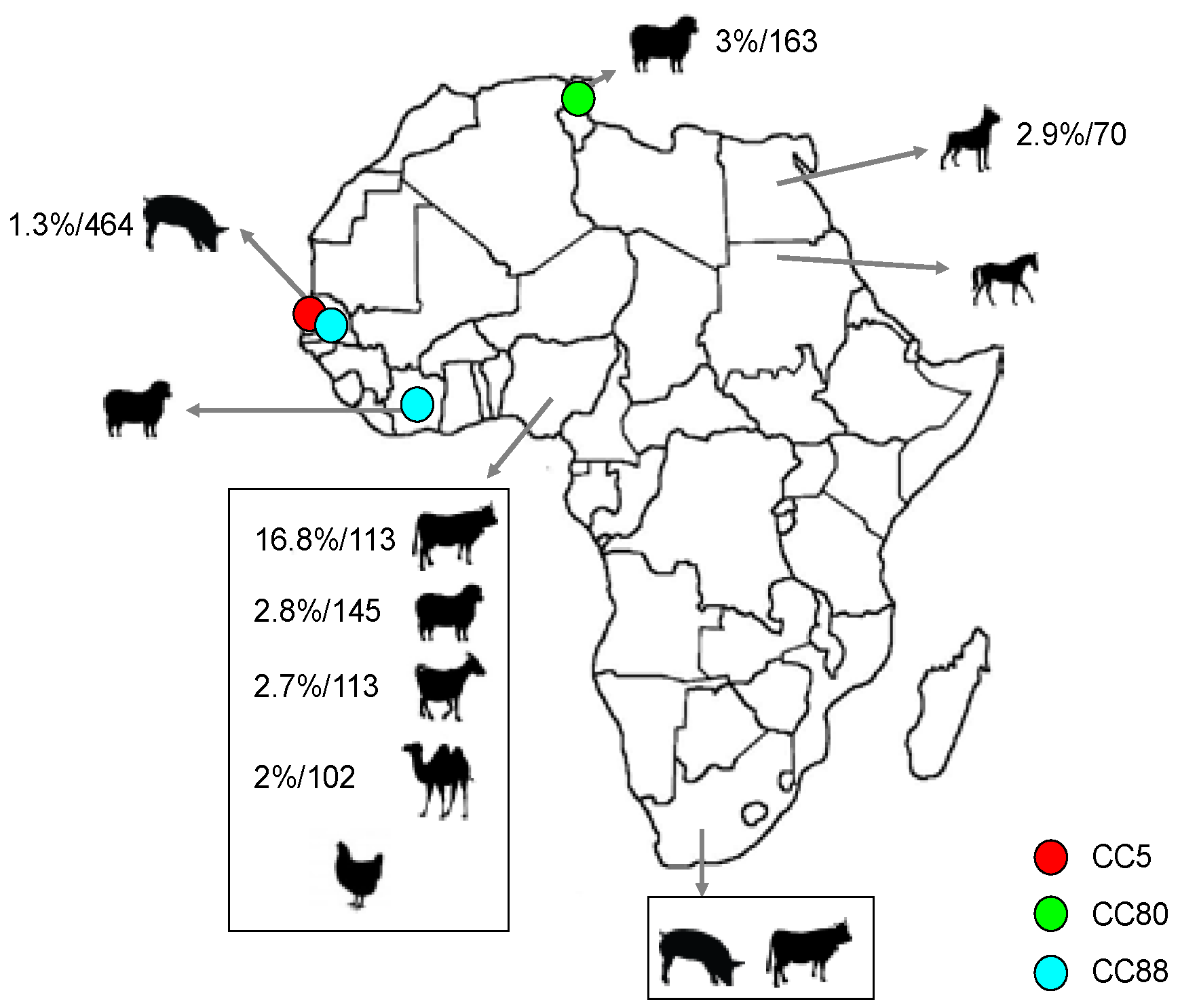
2.3. Population Structure of MRSA in Animals
2.4. S. aureus Interspecies Transmission
3. S. aureus in Food in Africa
3.1. MSSA Detection in Food Samples
3.2. MRSA Detection in Food Samples
| Country | Samples | Number of Samples Studied | Date of Sampling | Raw/Cooked | Detection of MRSA a | S. aureus Prevalence b | Reference |
|---|---|---|---|---|---|---|---|
| Cameroon | Pork | 11 | - | Cooked | ND a | 81.8% | [79] |
| Côte d’Ivoire | Beef, chickens, pork | 240 | 2010 | Cooked | Yes | 7.9% | [80] |
| Democratic Republic of Congo | Beef | - | - | Raw | ND | - | [81] |
| Egypt | Sausage, hamburger | 60 | - | Raw | ND | - | [65] |
| Liver, meat | 60 | - | Cooked | ND | - | [63] | |
| Fish (sardine, feseikh, molouha) | 60 | - | Cooked | Yes | - | [82] | |
| Milk | 150 | - | Raw | ND | 41.2% | [83] | |
| Goat (milk and meat) | 100 | - | Raw | ND | 58% milk 18% goat meat | [84] | |
| Ethiopia | Milk | - | 2011–2012 | Raw | yes | 100% | [85] |
| Meat samples | 100 | - | Raw | ND | 21.2% | [86] | |
| Gabon | Chicken | 151 | 2011–2012 | Raw | no | 3.3% | [78] |
| Kenya | Milk | - | - | Raw and cooked | no | - | [87] |
| Milk, minced meat | 96 | - | - | yes | - | [88] | |
| Milk | - | 2001–2002 | Raw | yes | - | [89] | |
| Lesotho | Cattle, pigs, sheep | 237 | - | Raw | ND | 5% | [90] |
| Libya | Chicken burger | 120 | - | Raw and cooked | ND | 29.6% raw 3.12% cooked | [91] |
| Malawi | Home cooked food | 132 | - | Cooked | ND | 61% (63% maize flour porridge, 51% fish, 75% vegetables, 69% beans, 38% others) | [92] |
| Morocco | Turkey | 96 | 2011–2012 | Raw | no | - | [93] |
| Milk, lben, jben | - | 2005–2006 | Raw and cooked | yes | - | [94] | |
| Meat and beef offal | 156 | 2002–2004 | Raw | ND | 16% | [95] | |
| Namibia | Milk | 15 | 1995–1996 | Cooked | ND | - | [96] |
| Nigeria | Ready-to-eat food | 168 | - | Raw and cooked | ND | 33.3% (57.1% salad, 19.1% meat pie, 14.3% fish roll, 9.6% doughnut) | [97] |
| Milk | 510 | 2012 | Raw | yes | 30.4% | [25] | |
| Suya, balangu, kilishi, dambunnama, raw beef | 300 | - | Raw and cooked | yes | 9.7% | [98] | |
| Chicken | 400 | - | Raw | yes | - | [26] | |
| Ready to eat food (meat, fish, vegetable) | 880 | - | Raw and cooked | ND | 62% | [99] | |
| Somalia | Milk | - | - | Raw and cooked | no | - | [87] |
| South Africa | Milk | 28 | - | Raw | yes | 100% | [100] |
| Milk | 156 | 1995–1996 | Cooked | ND | - | [96] | |
| Poultry | - | - | Raw | ND | 24.1% | [64] | |
| Street food vending (beef, chicken, salad, gravy) | 132 | - | Raw and cooked | ND | 3% | [101] | |
| Sudan | Sausage | 40 | - | Raw | ND | - | [68] |
| Milk | 320 | - | Raw | ND | 8.8% | [102] | |
| Milk | 90 | - | Raw | ND | - | [103] | |
| Tanzania | Milk | 128 | 2003 | Raw | ND | 6.3% | [104] |
| Tunisia | Chicken, horse, sheep, veal | 164 | 2010–2011 | Raw | yes | 26.2% | [46] |
| Uganda | Eggs | 171 | - | Raw | ND | 18% surface 4% inside | [105] |
| Zimbabwe | Milk | 140 | 2009–2010 | Raw and cooked | ND | - | [106] |
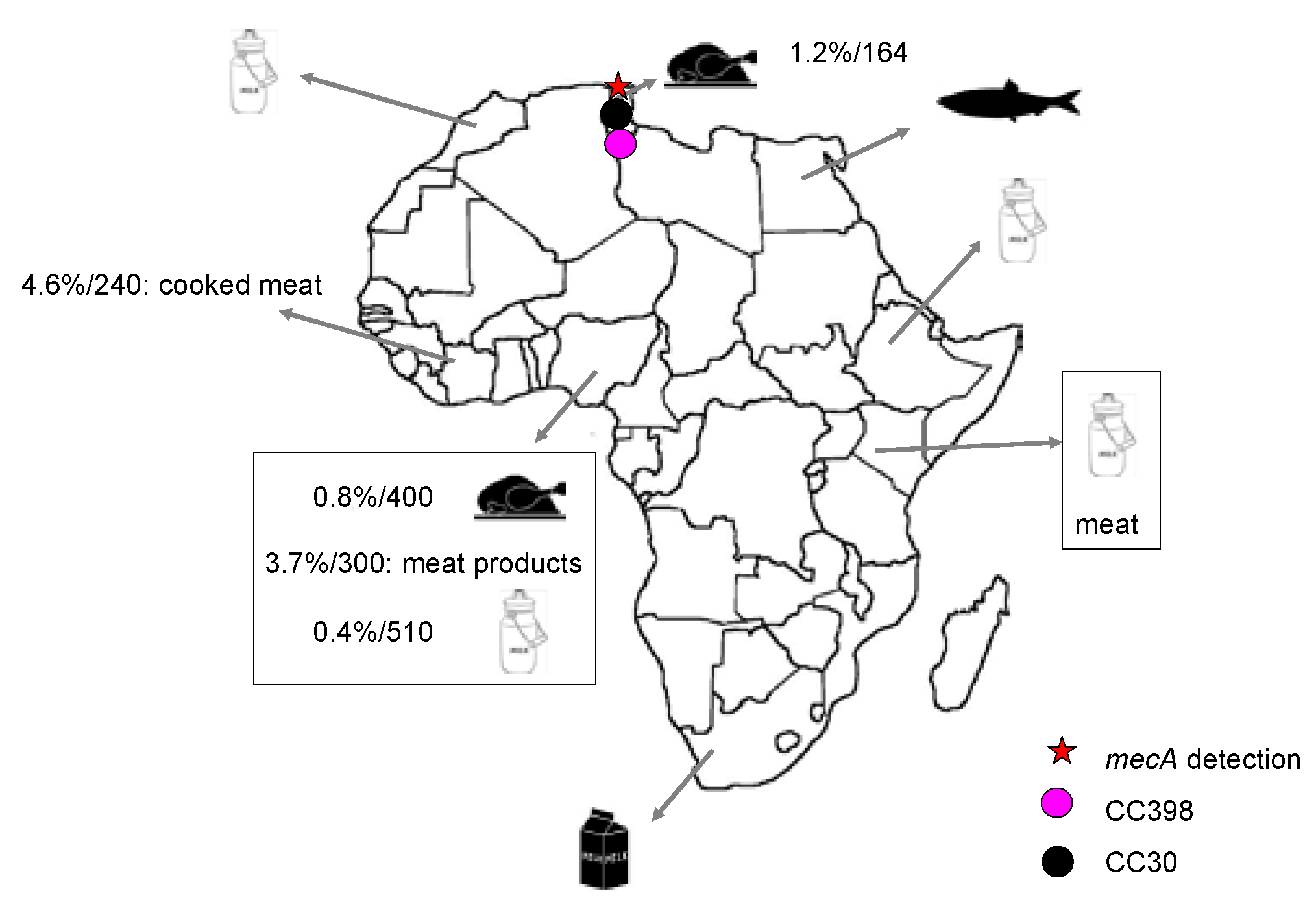
4. Other Important Characteristics of S. aureus from Animals and Food in Africa
4.1. Antimicrobial Resistance of S. aureus
4.2. Virulence Determinants
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Golding, G.R.; Bryden, L.; Levett, P.N.; McDonald, R.R.; Wong, A.; Wylie, J.; Graham, M.R.; Tyler, S.; van Domselaar, G.; Simor, A.E.; et al. Livestock-associated methicillin-resistant Staphylococcus aureus sequence type 398 in humans, Canada. Emerg. Infect. Dis. 2010, 16, 587–594. [Google Scholar] [CrossRef] [PubMed]
- Mediavilla, J.R.; Chen, L.; Uhlemann, A.C.; Hanson, B.M.; Rosenthal, M.; Stanak, K.; Koll, B.; Fries, B.C.; Armellino, D.; Schilling, M.E.; et al. Methicillin-susceptible Staphylococcus aureus ST398, New York and New Jersey, USA. Emerg. Infect. Dis. 2012, 18, 700–702. [Google Scholar] [CrossRef] [PubMed]
- Cuny, C.; Layer, F.; Köck, R.; Werner, G.; Witte, W. Methicillin susceptible Staphylococcus aureus (MSSA) of clonal complex CC398, t571 from infections in humans are still rare in Germany. PLoS ONE 2013, 8, e83165. [Google Scholar] [CrossRef] [PubMed]
- Witte, W.; Strommenger, B.; Stanek, C.; Cuny, C. Methicillin resistant Staphylococcus aureus ST398 in humans and animals, Central Europe. Emerg. Infect. Dis. 2007, 13, 255–258. [Google Scholar] [CrossRef] [PubMed]
- Gómez, P.; Lozano, C.; Camacho, M.C.; Lima-Barbero, J.F.; Hernández, J.M.; Zarazaga, M.; Höfle, U.; Torres, C. Detection of MRSA ST3061-t843-mecC and ST398-t011-mecA in white stork nestlings exposed to human residues. J. Antimicrob. Chemother. 2016, 71, 53–57. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Sanz, E.; Simón, C.; Ortega, C.; Gómez, P.; Lozano, C.; Zarazaga, M.; Torres, C. First detection of methicillin-resistant Staphylococcus aureus ST398 and Staphylococcus pseudintermedius ST68 from hospitalized equines in Spain. Zoonoses Public Health 2014, 61, 192–201. [Google Scholar] [CrossRef] [PubMed]
- Benito, D.; Lozano, C.; Rezusta, A.; Ferrer, I.; Vasquez, M.A.; Ceballos, S.; Zarazaga, M.; Revillo, M.J.; Torres, C. Characterization of tetracycline and methicillin resistant Staphylococcus aureus strains in a Spanish hospital: Is livestock-contact a risk factor in infections caused by MRSA CC398? Int. J. Med. Microbiol. 2014, 304, 1226–1232. [Google Scholar] [CrossRef] [PubMed]
- Gómez, P.; González-Barrio, D.; Benito, D.; García, J.T.; Viñuela, J.; Zarazaga, M.; Ruiz-Fons, F.; Torres, C. Detection of methicillin-resistant S. aureus carrying the mecC gene in wild small mammals in Spain. J. Antimicrob. Chemother. 2014, 69, 2061–2064. [Google Scholar] [CrossRef] [PubMed]
- Strommenger, B.; Kehrenberg, C.; Kettlitz, C.; Cuny, C.; Verspohl, J.; Witte, W.; Schwarz, S. Molecular characterization of methicillin-resistant Staphylococcus aureus strains from pet animals and their relationship to human isolates. J. Antimicrob. Chemother. 2006, 57, 461–465. [Google Scholar] [CrossRef] [PubMed]
- Baptiste, K.E.; Williams, K.; Willams, N.J.; Wattret, A.; Clegg, P.D.; Dawson, S.; Corkill, J.E.; O’Neill, T.; Hart, C.A. Methicillin-resistant staphylococci in companion animals. Emerg. Infect. Dis. 2005, 11, 1942–1944. [Google Scholar] [CrossRef] [PubMed]
- Geneva: World Health Organization. Antimicrobial Resistance: Global Report on Surveillance 2014. Available online: http://www.who.int/drugresistance/documents/surveillancereport/en/ (accessed on 30 November 2015).
- Kimang’a, A.N. A situational analysis of antimicrobial drug resistance in Africa: Are we losing the battle? Ethiop. J. Health Sci. 2012, 22, 135–143. [Google Scholar] [PubMed]
- BoscoNdihokubwayo, J.; Ahmed Yahaya, A.; Tamer Desta, A.; Ki-Zerbo, G.; AssamoahOdei, E.; Keita, B.; PanaAssimawe, P.; Nkhoma, W. Antimicrobial Resistance in the African Region: Issues, Challenges and Actions Proposed. Available online: http://www.afro.who.int/en/clusters-a-programmes/hss/blood-safety-laboratories-a-health-technology/blt-highlights/3861-antimicrobial-resistance-in-the-african-region-issues-challenges-and-actions-proposed.html (accessed on 19 August 2015).
- Falagas, M.E.; Karageorgopoulos, D.E.; Leptidis, J.; Korbila, I.P. MRSA in Africa: Filling the global map of antimicrobial resistance. PLoS ONE 2013, 8, e68024. [Google Scholar]
- Schaumburg, F.; Alabi, A.S.; Peters, G.; Becker, K. New epidemiology of Staphylococcus aureus infection in Africa. Clin. Microbiol. Infect. 2014, 20, 589–596. [Google Scholar] [CrossRef] [PubMed]
- Abdulgader, S.M.; Shittu, A.O.; Nicol, M.P.; Kaba, M. Molecular epidemiology of Methicillin-resistant Staphylococcus aureus in Africa: A systematic review. Front. Microbiol. 2015. [Google Scholar] [CrossRef] [PubMed]
- Schaumburg, F.; Pauly, M.; Anoh, E.; Mossoun, A.; Wiersma, L.; Schubert, G.; Flammen, A.; Alabi, A.S.; Muyembe-Tamfum, J.J.; Grobusch, M.P.; et al. Staphylococcus aureus complex from animals and humans in three remote African regions. Clin. Microbiol. Infect. 2015, 21, e1–e8. [Google Scholar] [CrossRef] [PubMed]
- Schaumburg, F.; Mugisha, L.; Kappeller, P.; Fichtel, C.; Köck, R.; Köndgen, S.; Becker, K.; Boesch, C.; Peters, G.; Leendertz, F. Evaluation of non-invasive biological samples to monitor Staphylococcus aureus colonization in great apes and lemurs. PLoS ONE 2013, 8, e78046. [Google Scholar] [CrossRef]
- Schaumburg, F.; Alabi, A.S.; Köck, R.; Mellmann, A.; Kremsner, P.G.; Boesch, C.; Becker, K.; Leendertz, F.H.; Peters, G. Highly divergent Staphylococcus aureus isolates from African non-human primates. Environ. Microbiol. Rep. 2012, 4, 141–146. [Google Scholar] [CrossRef] [PubMed]
- Abdel-moein, K.A.; El-Hariri, M.; Samir, A. Methicillin-resistant Staphylococcus aureus: An emerging pathogen of pets in Egypt with a public health burden. Transbound. Emerg. Dis. 2012, 59, 331–335. [Google Scholar] [CrossRef] [PubMed]
- El-Jakee, J.; Nagwa, A.S.; Bakry, M.; Zouelfakar, S.A.; Elgabry, E.; Gad El-Said, W.A. Characteristics of Staphylococcus aureus Strains Isolated from Human and Animal Sources. Am. Eur. J. Agric. Environ. Sci. 2008, 4, 221–229. [Google Scholar]
- Nagel, M.; Dischinger, J.; Türck, M.; Verrier, D.; Oedenkoven, M.; Ngoubangoye, B.; Le Flohic, G.; Drexler, J.F.; Bierbaum, G.; Gonzalez, J.P. Human-associated Staphylococcus aureus strains within great ape populations in Central Africa (Gabon). Clin. Microbiol. Infect. 2013, 19, 1072–1077. [Google Scholar] [CrossRef] [PubMed]
- Adesiyun, A.A.; Lenz, W.; Schaal, K.P. Production of toxic shock syndrome toxin-1 (TSST-1) by Staphylococcus aureus strains isolated from humans, animals and foods in Nigeria. Microbiologica 1992, 15, 125–133. [Google Scholar] [PubMed]
- Akobi, B.; Aboderin, O.; Sasaki, T.; Shittu, A. Characterization of Staphylococcus aureus isolates from faecal samples of the Straw-Coloured Fruit Bat (Eidolon helvum) in Obafemi Awolowo University (OAU), Nigeria. BMC Microbiol. 2012. [Google Scholar] [CrossRef] [PubMed]
- Mai-siyama, I.B.; Okon, K.O.; Adamu, N.B.; Askira, U.M.; Isyaka, T.M.; Adamu, S.G.; Mohammed, A. Methicllin-resistant Staphylococcus aureus (MRSA) colonization rate among ruminant animals slaughtered for human consumption and contact persons in Maiduguri, Nigeria. Afr.J. Microbiol. Res. 2014, 8, 2643–2649. [Google Scholar] [CrossRef]
- Otalu, O.; Junaidu, K.; Chukwudi, O.E.; Jarlath, U.V. Multi-drug resistant coagulase positive Staphylococcus aureus from live and slaughtered chickens in Zaria, Nigeria. Int. J. Poult. Sci. 2011, 10, 871–875. [Google Scholar] [CrossRef]
- Fall, C.; Seck, A.; Richard, V.; Ndour, M.; Sembene, M.; Laurent, F.; Breurec, S. Epidemiology of Staphylococcus aureus in pigs and farmers in the largest farm in Dakar, Senegal. Foodborne Pathog. Dis. 2012, 9, 962–965. [Google Scholar] [CrossRef] [PubMed]
- Adegoke, A.A.; Okoh, A.I. Species diversity and antibiotic resistance properties of Staphylococcus of farm animal origin in Nkonkobe Municipality, South Africa. Folia Microbiol. Praha 2014, 59, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Schaumburg, F.; Mugisha, L.; Peck, B.; Becker, K.; Gillespie, T.R.; Peters, G.; Leendertz, F.H. Drug-resistant human Staphylococcus aureus in sanctuary apes pose a threat to endangered wild ape populations. Am. J. Primatol. 2012, 74, 1071–1075. [Google Scholar] [CrossRef] [PubMed]
- Tanih, N.F.; Sekwadi, E.; Ndip, R.N.; Bessong, P.O. Detection of pathogenic Escherichia coli and Staphylococcus aureus from cattle and pigs slaughtered in abattoirs in Vhembe District, South Africa. Sci. World J. 2015. [Google Scholar] [CrossRef] [PubMed]
- Elbir, H.; Feil, E.J.; Drancourt, M.; Roux, V.; El Sanousi, S.M.; Eshag, M.; Colque-Navarro, P.; Kühn, I.; Flock, J.I. Ovine clone ST1464: A predominant genotype of Staphylococcus aureus subsp. anaerobius isolated from sheep in Sudan. J. Infect. Dev. Ctries. 2010, 4, 235–238. [Google Scholar] [PubMed]
- Musa, N.O.; Babiker, A.; Eltom, K.; Rodwan, K.; El Sanousi, S.M. Prevalence of Staphylococcus aureus subsp. anaerobius in Sub-Clinical Abscess Cases of Sheep. Br. Microbiol. Res. J. 2012, 2, 131–136. [Google Scholar]
- Omer, M.M.; Abusalab, S.; Gumaa, M.M.; Mulla, S.A.; Osman, H.M.; Sabiel, Y.A.; Ahmed, A.M. Staphylococcus aureus isolated from a horse in a sudden death condition in Kassala state, eastern Sudan. Pak. J. Biol. Sci. 2008, 11, 2028–2031. [Google Scholar] [CrossRef] [PubMed]
- Gharsa, H.; Ben Slama, K.; Lozano, C.; Gómez-Sanz, E.; Klibi, N.; Ben Sallem, R.; Gómez, P.; Zarazaga, M.; Boudabous, A.; Torres, C. Prevalence, antibiotic resistance, virulence traits and genetic lineages of Staphylococcus aureus in healthy sheep in Tunisia. Vet. Microbiol. 2012, 156, 367–373. [Google Scholar] [CrossRef] [PubMed]
- Gharsa, H.; Ben Sallem, R.; Ben Slama, K.; Gómez-Sanz, E.; Lozano, C.; Jouini, A.; Klibi, N.; Zarazaga, M.; Boudabous, A.; Torres, C. High diversity of genetic lineages and virulence genes in nasal Staphylococcus aureus isolates from donkeys destined to food consumption in Tunisia with predominance of the ruminant associated CC133 lineage. BMC Vet. Res. 2012. [Google Scholar] [CrossRef] [PubMed]
- Gharsa, H.; Ben Slama, K.; Gómez-Sanz, E.; Lozano, C.; Zarazaga, M.; Messadi, L.; Boudabous, A.; Torres, C. Molecular characterization of Staphylococcus aureus from nasal samples of healthy farm animals and pets in Tunisia. Vector Borne Zoonotic Dis. 2015, 15, 109–115. [Google Scholar] [CrossRef] [PubMed]
- Pandey, G.S.; Nomura, Y.; Kobayashi, K.; Fujise, H.; Yamada, T. Cutaneous staphylococcal granuloma in a free living zebra (Equus burchelli) in Zambia. J. Vet. Med. Sci. 1998, 60, 137–138. [Google Scholar] [CrossRef] [PubMed]
- Youn, J.H.; Park, Y.H.; Hang’ombe, B.; Sugimoto, C. Prevalence and characterization of Staphylococcus aureus and Staphylococcus pseudintermedius isolated from companion animals and environment in the veterinary teaching hospital in Zambia, Africa. Comp. Immunol. Microbiol. Infect. Dis. 2014, 37, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Guinane, C.M.; Ben Zakour, N.L.; Tormo-Mas, M.A.; Weinert, L.A.; Lowder, B.V.; Cartwright, R.A.; Smyth, D.S.; Smyth, C.J.; Lindsay, J.A.; Gould, K.A.; et al. Evolutionary genomics of S. aureus reveals insights into the origin and molecular basis of ruminant host adaptation. Genome Biol. Evol. 2010, 2, 454–466. [Google Scholar] [CrossRef] [PubMed]
- García-Álvarez, L.; Holden, M.T.; Lindsay, H.; Webb, C.R.; Brown, D.F.; Curran, M.D.; Walpole, E.; Brooks, K.; Pickard, D.J.; Teale, C.; et al. Meticillin-resistant Staphylococcus aureus with a novel mecA homologue in human and bovine populations in the UK and Denmark: A descriptive study. Lancet Infect. Dis. 2011, 11, 595–603. [Google Scholar] [CrossRef]
- Loeffler, A.; Lloyd, D.H. Companion animals: A reservoir for methicillin-resistant Staphylococcus aureus in the community? Epidemiol. Infect. 2010, 138, 595–605. [Google Scholar] [CrossRef] [PubMed]
- Crombé, F.; Argudín, M.A.; Vanderhaeghen, W.; Hermans, K.; Haesebrouck, F.; Butaye, P. Transmission dynamics of Methicillin-Resistant Staphylococcus aureus in pigs. Front. Microbiol 2013. [Google Scholar] [CrossRef] [PubMed]
- Persoons, D.; van Hoorebeke, S.; Hermans, K.; Butaye, P.; de Kruif, A.; Haesebrouck, F.; Dewulf, J. Methicillin-resistant Staphylococcus aureus in poultry. Emerg. Infect. Dis. 2009, 15, 452–453. [Google Scholar] [CrossRef] [PubMed]
- Huber, H.; Koller, S.; Giezendanner, N.; Stephan, R.; Zweifel, C. Prevalence and characteristics of meticillin-resistant Staphylococcus aureus in humans in contact with farm animals, in livestock, and in food of animal origin, Switzerland, 2009. Euro Surveill. 2010, 15, 19542. Available online: http://www.eurosurveillance.org/images/dynamic/ee/v15n16/art19542.pdf (accessed on 30 November 2015). [Google Scholar] [PubMed]
- Elhani, D.; Gharsa, H.; Kalai, D.; Lozano, C.; Gomez, P.; Boutheina, J.; Aouni, M.; Barguellil, F.; Torres, C.; Ben Slama, K. Clonal lineages detected among tetracycline resistant MRSA isolates of a Tunisian Hospital, with detection of lineage ST398. J. Med. Microbiol. 2015. [Google Scholar] [CrossRef] [PubMed]
- Chairat, S.; Gharsa, H.; Lozano, C.; Gómez-Sanz, E.; Gómez, P.; Zarazaga, M.; Boudabous, A.; Torres, C.; Ben Slama, K. Characterization of Staphylococcus aureus from raw meat samples in tunisia: Detection of clonal lineage ST398 from the African Continent. Foodborne Pathog. Dis. 2015, 12, 686–692. [Google Scholar] [CrossRef] [PubMed]
- Paterson, G.K.; Harrison, E.M.; Holmes, M.A. The emergence of mecC methicillin-resistant Staphylococcus aureus. Trends Microbiol. 2014, 22, 42–47. [Google Scholar] [CrossRef] [PubMed]
- Paterson, G.K.; Morgan, F.J.; Harrison, E.M.; Cartwright, E.J.; Török, M.E.; Zadoks, R.N.; Parkhill, J.; Peacock, S.J.; Holmes, M.A. Prevalence and characterisation of human mecC methicillin-resistant S. aureus isolates in England. J. Antimicrob. Chemother. 2014, 69, 907–910. [Google Scholar] [CrossRef] [PubMed]
- Rabello, R.F.; Moreira, B.M.; Lopes, R.M.M.; Teixeira, L.M.; Riley, L.W.; Castro, A.C.D. Multilocus sequence typing of Staphylococcus aureus isolates recovered from cows with mastitis in Brazilian dairy herds. J. Med. Microbiol. 2007, 56, 1505–1511. [Google Scholar] [CrossRef] [PubMed]
- Hasman, H.; Moodley, A.; Guardabassi, L.; Stegger, M.; Skov, R.L.; Aarestrup, F.M. spa type distribution in Staphylococcus aureus originating from pigs, cattle and poultry. Vet. Microbiol. 2010, 141, 326–331. [Google Scholar] [CrossRef] [PubMed]
- Porrero, M.C.; Hasman, H.; Vela, A.I.; Fernández-Garayzábal, J.F.; Domínguez, L.; Aarestrup, F.M. Clonal diversity of Staphylococcus aureus originating from the small ruminants goats and sheep. Vet. Microbiol. 2012, 156, 157–161. [Google Scholar] [CrossRef] [PubMed]
- Lowder, B.V.; Guinane, C.M.; Ben Zakour, N.L.; Weinert, L.A.; Conway-Morris, A.; Cartwright, R.A.; Simpson, A.J.; Rambaut, A.; Nübel, U.; Fitzgerald, J.R. Recent human-to-poultry host jump, adaptation, and pandemic spread of Staphylococcus aureus. Proc. Natl. Acad. Sci. USA 2009, 106, 19545–19550. [Google Scholar] [CrossRef] [PubMed]
- Smyth, D.S.; Feil, E.J.; Meaney, W.J.; Hartigan, P.J.; Tollersrud, T.; Fitzgerald, J.R.; Enright, M.C.; Smyth, C.J. Molecular genetic typing reveals further insights into the diversity of animal-associated Staphylococcus aureus. J. Med. Microbiol. 2009, 58, 1343–1353. [Google Scholar] [CrossRef] [PubMed]
- Franco, A.; Hasman, H.; Iurescia, M.; Lorenzetti, R.; Stegger, M.; Pantosti, A.; Feltrin, F.; Ianzano, A.; Porrero, M.C.; Liapi, M.; et al. Molecular characterization of spa type t127, sequence type 1 methicillin-resistant Staphylococcus aureus from pigs. J. Antimicrob. Chemother. 2011, 66, 1231–1235. [Google Scholar] [CrossRef] [PubMed]
- Cui, S.; Li, J.; Hu, C.; Jin, S.; Li, F.; Guo, Y.; Ran, L.; Ma, Y. Isolation and characterization of methicillin-resistant Staphylococcus aureus from swine and workers in China. J. Antimicrob. Chemother. 2009, 64, 680–683. [Google Scholar] [CrossRef] [PubMed]
- Vestergaard, M.; Cavaco, L.M.; Sirichote, P.; Unahalekhaka, A.; Dangsakul, W.; Svendsen, C.A.; Aarestrup, F.M.; Hendriksen, R.S. SCCmec Type IX element in methicillin resistant Staphylococcus aureus spa type t337 (CC9) isolated from pigs and pork in Thailand. Front. Microbiol. 2012, 3, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Pomba, C.; Hasman, H.; Cavaco, L.M.; da Fonseca, J.D.; Aarestrup, F.M. First description of meticillin-resistant Staphylococcus aureus (MRSA) CC30 and CC398 from swine in Portugal. Int. J. Antimicrob. Agents 2009, 34, 193–194. [Google Scholar] [CrossRef] [PubMed]
- Vancraeynest, D.; Haesebrouck, F.; Deplano, A.; Denis, O.; Godard, C.; Wildemauwe, C.; Hermans, K. International dissemination of a high virulence rabbit Staphylococcus aureus clone. J. Vet. Med. B Infect. Dis. Vet. Public Health 2006, 53, 418–422. [Google Scholar] [CrossRef] [PubMed]
- Walther, B.; Monecke, S.; Ruscher, C.; Friedrich, A.W.; Ehricht, R.; Slickers, P.; Soba, A.; Wleklinski, C.G.; Wieler, L.H.; Lübke-Becker, A. Comparative molecular analysis substantiates zoonotic potential of equine methicillin-resistant Staphylococcus aureus. J. Clin. Microbiol. 2009, 47, 704–710. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Sanz, E.; Torres, C.; Lozano, C.; Fernández-Pérez, R.; Aspiroz, C.; Ruiz-Larrea, F.; Zarazaga, M. Detection, molecular characterization, and clonal diversity of methicillin-resistant Staphylococcus aureus CC398 and CC97 in Spanish slaughter pigs of different age groups. Foodborne Pathog. Dis. 2010, 7, 1269–1277. [Google Scholar] [CrossRef] [PubMed]
- Sakwinska, O.; Giddey, M.; Moreillon, M.; Morisset, D.; Waldvogel, A.; Moreillon, P. Staphylococcus aureus host range and human-bovine host shift. Appl. Environ. Microbiol. 2011, 77, 5908–5915. [Google Scholar] [CrossRef] [PubMed]
- Ben Slama, K.; Gharsa, H.; Klibi, N.; Jouini, A.; Lozano, C.; Gómez-Sanz, E.; Zarazaga, M.; Boudabous, A.; Torres, C. Nasal carriage of Staphylococcus aureus in healthy humans with different levels of contact with animals in Tunisia: genetic lineages, methicillin resistance, and virulence factors. Eur. J. Clin. Microbiol. Infect. Dis. 2011, 30, 499–508. [Google Scholar] [CrossRef] [PubMed]
- Abou, E.A. Bacteriological quality of ready to eat meals. J. Egypt Public Health Assoc. 1995, 70, 627–641. [Google Scholar] [PubMed]
- Geornaras, I.; de Jesus, A.; van Zyl, E.; von Holy, A. Microbiological survey of a South African poultry processing plant. J. Basic Microbiol. 1995, 35, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Abd El Aziz, T. Screening of some food poisoning bacteria in sausage and hamburger meat. J. Egypt Public Health Assoc. 1996, 71, 47–61. [Google Scholar] [PubMed]
- Karama, M.; de Jesus, A.E.; Veary, C.M. Microbial quality of ostrich carcasses produced at an export-approved South African abattoir. J. Food Prot. 2003, 66, 878–881. [Google Scholar] [PubMed]
- Nel, S.; Lues, J.F.; Buys, E.M.; Venter, P. Bacterial populations associated with meat from the deboning room of a high throughput red meat abattoir. Meat Sci. 2004, 66, 667–674. [Google Scholar] [CrossRef]
- Elhag, N.B.; Babiker, E.R.B.; Mahdi, A.A. Microbial profile of sausages in Khartoum State. J. Agri. Food Appl. Sci. 2014, 2, 206–219. [Google Scholar]
- Kivaria, F.M.; Noordhuizen, J.P.; Nielen, M. Interpretation of California mastitis test scores using Staphylococcus aureus culture results for screening of subclinical mastitis in low yielding smallholder dairy cows in the Dar es Salaam region of Tanzania. Prev. Vet. Med. 2007, 78, 274–285. [Google Scholar] [CrossRef] [PubMed]
- Almaw, G.; Zerihun, A.; Asfaw, Y. Bovine mastitis and its association with selected risk factors in smallholder dairy farms in and around Bahir Dar, Ethiopia. Trop. Anim. Health Prod. 2008, 40, 427–432. [Google Scholar] [CrossRef] [PubMed]
- Abera, M.; Demie, B.; Aragaw, K.; Regassa, F.G.; Regassa, A. Isolation and identification of Staphylococcus aureus from bovine mastitic milk and their drug resistance patterns in Adama town, Ethiopia. J. Vet. Med. Anim. Health 2010, 2, 29–34. [Google Scholar]
- Mengistu, F.; Molla, B.; Ali, A.I. Camel mastitis, associated bacterial pathogens and its impact on milk quality in Gewane district, afar regional state, Northeastern Ethiopia. Bull. Anim. Health Prod. Afr. 2010, 58, 241–247. [Google Scholar]
- Kadja, M.C.; Kpodekon, M.; Kane, Y.; Tchassou, K.; Kaboret, Y.; Mainil, J.; Taminiau, B. Typing of Staphylococcus aureus strains isolated from milk cows with subclinical mastitis in Dakar, Senegal. Bull. Anim. Health Prod. Afr. 2010, 58, 201–209. [Google Scholar] [CrossRef]
- Haftu, R.; Taddele, H.; Gugsa, G.; Kalayou, S. Prevalence, bacterial causes, and antimicrobial susceptibility profile of mastitis isolates from cows in large-scale dairy farms of Northern Ethiopia. Trop Anim Health Prod 2012, 44, 1765–1771. [Google Scholar] [CrossRef] [PubMed]
- Alemu, G.; Almaw, G.; Abera, M. Incidence rate of Staphylococcus aureus and Streptococcus agalactiae in subclinical mastitis at smallholder dairy cattle farms in Hawassa, Ethiopia. Afr. J. Microbiol. Res. 2014, 8, 252–256. [Google Scholar]
- Ibrahim, A.I.; Duprez, J.; Bada-Alambedji, R.; Moula, N.; Mainil, J.G.; Bardiau, M. Antibiotic resistance trend of Staphylococcus aureus isolated between 2010 and 2012 from mastitis cases in Azawak zebu in Niger. Afr. J. Microbiol. Res. 2014, 35, 3271–3275. [Google Scholar]
- Iceland Kasozi, K.; BoscoTingiira, J.; Vudriko, P. High prevalence of subclinical mastitis and multidrug resistant Staphylococcus aureus are a threat to dairy cattle production in Kiboga District (Uganda). Open J. Vet. Med. 2014, 4, 35–43. [Google Scholar] [CrossRef]
- Schaumburg, F.; Alabi, A.S.; Frielinghaus, L.; Grobusch, M.P.; Köck, R.; Becker, K.; Issifou, S.; Kremsner, P.G.; Peters, G.; Mellmann, A. The risk to import ESBL-producing Enterobacteriaceae and Staphylococcus aureus through chicken meat trade in Gabon. BMC Microbiol. 2014. [Google Scholar] [CrossRef] [PubMed]
- Yannick, N.; Rawlings, N.; Emmanuela, A. Assessment of bacteriological quality of cooked pork meat sold along the commercial streets of Nkwen through Bambili Metropolis, Cameroon. Afr. J. Food Sci. 2013, 7, 441–445. [Google Scholar] [CrossRef]
- Attien, P.S.; Moussaoui, W.; Dadié, T.; Chabi Sika, K.; Djéni, T.; Bankole, H.S.; Kotchoni, S.O.; Edoh, V.; Prévost, G.; Djè, M.; et al. Prevalence and antibiotic resistance of Staphylococcus strains isolated from meat products sold in Abidjan streets (Ivory Coast). Afr. J. Microbiol. Res. 2013, 7, 3285–3293. [Google Scholar]
- Mathieu, A.; Isigidi, B.K.; Devriese, L.A.; Godard, C.; Vanhoof, R. Characterization of Staphylococcus aureus and Salmonella species strains isolated from bovine meat in Zaire. Int. J. Food Microbiol. 1991, 14, 119–126. [Google Scholar] [CrossRef]
- Ezzeldeen, N.A.; Mansour, H.A.; Ahmed, A.A. Phenotypic and molecular identification of Staphylococcus aureus isolated from some egyptian salted fish. World Appl. Sci. J. 2011, 15, 1703–1712. [Google Scholar]
- Gwida, M.M.; EL-Gohary, F.A. Zoonotic bacterial pathogens isolated from raw milk with special reference to Escherichia coli and Staphylococcus aureus in Dakahlia Governorate, Egypt. Open Access Sci. Rep. 2013. [Google Scholar] [CrossRef]
- Khalifa, N.O.; Elhofy, F.I.; Fahmy, H.A.; Barakat, A.M.A. Epidemiological and genetic studies of enterotoxigenic Staphylococcus aureus isolated from goat and human. Am. J. Infect. Dis. Microbiol. 2015, 3, 32–37. [Google Scholar]
- Daka, D.; Silassie, S.; Yihdego, D. Antibiotic-resistance Staphylococcus aureus isolated from cow’s milk in the Hawassa area, South Ethiopia. Ann. Clin. Microbiol. Antimicrob. 2012. [Google Scholar] [CrossRef] [PubMed]
- Haileselassie, M.; Taddele, H.; Adhana, K.; Kalayou, S. Food safety knowledge and practices of abattoir and butchery shops and the microbial profile of meat in Mekelle city, Ethiopia. Asian Pac. J. Trop. Biomed. 2013, 3, 407–412. [Google Scholar] [CrossRef]
- Njage, P.M.K.; Dolci, S.; Jans, C.; Wangoh, J.; Lacroix, C.; Meile, L. Phenotypic and genotypic antibiotic resistance patterns of Staphylococcus aureus from raw and spontaneously fermented camel milk. Eur. J. Food Res. Rev. 2013, 3, 87–98. [Google Scholar] [CrossRef] [PubMed]
- Ombui, J.N.; Kimotho, A.M.; Nduhiu, J.G. Antimicrobial resistance patterns and plasmid profiles of Staphylococcus aureus isolated from milk and meat. East Afr. Med. J. 2000, 77, 463–467. [Google Scholar] [CrossRef] [PubMed]
- Shitandi, A.; Mwangi, M. Occurrence of miltiple antimicrobial resistance among Staphylococcus aureus isolates from Kenyan milk. J. Food Technol. Afr. 2004, 9, 23–25. [Google Scholar]
- Seeiso, T.M.; McCrindle, C.M. An investigation of the quality of meat sold in Lesotho. J.S. Afr. Vet. Assoc. 2009, 80, 237–242. [Google Scholar] [CrossRef] [PubMed]
- El Shrek, Y.M.; Ali, M.R. Microbiological study of spiced chicken burgers in Tripoli City, Libya. East Mediterr. Health J. 2012, 18, 653–662. [Google Scholar] [PubMed]
- Taulo, S.; Wetlesen, A.; Abrahamsen, R.; Kululanga, G.; Mkakosya, R.; Grimason, A. Microbiological hazard identification and exposure assessment of food prepared and served in rural households of Lungwena, Malawi. Int. J. Food Microbiol. 2008, 125, 111–116. [Google Scholar] [CrossRef] [PubMed]
- Abdellah, E.A.; Fouzia, R.F.; Bouchra, O. Prevalence and antibiogram study of Escherichia coli and Staphylococcus aureus in turkey meat in Morocco. Pharm. Anal. Acta 2013. [Google Scholar] [CrossRef]
- Bendahou, A.; Lebbadi, M.; Ennanei, L.; Essadqui, F.Z.; Abid, M. Characterization of Staphylococcus species isolated from raw milk and milk products (lben and jben) in North Morocco. J. Infect. Dev. Ctries. 2008, 2, 218–225. [Google Scholar] [PubMed]
- Cohen, N.; Ennaji, H.; Hassar, M.; Karib, H. The bacterial quality of red meat and offal in Casablanca (Morocco). Mol. Nutr. Food Res. 2006, 50, 557–562. [Google Scholar] [CrossRef] [PubMed]
- Beukes, E.M.; Bester, B.H.; Mostert, J.F. The microbiology of South African traditional fermented milks. Int. J. Food Microbiol. 2001, 63, 189–197. [Google Scholar] [CrossRef]
- Isara, A.R.; Isah, E.C.; Lofor, P.V.; Ojide, C.K. Food contamination in fast food restaurants in Benin City, Edo State, Nigeria: implications for food hygiene and safety. Public Health 2010, 124, 467–471. [Google Scholar] [CrossRef] [PubMed]
- Ndahi, M.D.; Kwaga, J.K.; Bello, M.; Kabir, J.; Umoh, V.J.; Yakubu, S.E.; Nok, A.J. Prevalence and antimicrobial susceptibility of Listeria monocytogenes and methicillin-resistant Staphylococcus aureus strains from raw meat and meat products in Zaria, Nigeria. Lett. Appl. Microbiol. 2014, 58, 262–269. [Google Scholar] [CrossRef] [PubMed]
- Sokari, T. Distribution of enterotoxigenic Staphylococcus aureus in ready-to-eat foods in eastern Nigeria. Int. J. Food Microbiol. 1991, 12, 275–279. [Google Scholar] [CrossRef]
- Ateba, C.N.; Mbewe, M.; Moneoang, M.S.; Bezuidenhout, C.C. Antibiotic-resistant Staphylococcus aureus isolated from milk in the Mafikeng Area, North West province, South Africa. S. Afr. J. Sci. 2010, 106, 1–6. [Google Scholar] [CrossRef]
- Mosupye, F.M.; von Holy, A. Microbiological hazard identification and exposure assessment of street food vending in Johannesburg, South Africa. Int. J. Food Microbiol. 2000, 61, 137–145. [Google Scholar] [CrossRef]
- Shuiep, E.S.; Kanbar, T.; Eissa, N.; Alber, J.; Lämmler, C.; Zschöck, M.; El Zubeir, I.E.; Weiss, R. Phenotypic and genotypic characterization of Staphylococcus aureus isolated from raw camel milk samples. Res. Vet. Sci. 2009, 86, 211–215. [Google Scholar] [CrossRef] [PubMed]
- Yagoub, S.O.; Awadalla, N.E.; El Zubeir, I.E.M. Incidence of some potential pathogens in raw milk in Khartoum North (Sudan) and their susceptinility to antimicrobial agents. J. Anim. Vet. Adv. 2005, 4, 341–344. [Google Scholar]
- Kivaria, F.M.; Noordhuizen, J.P.; Kapaga, A.M. Evaluation of the hygienic quality and associated public health hazards of raw milk marketed by smallholder dairy producers in the Dar es Salaam region, Tanzania. Trop. Anim. Health Prod. 2006, 38, 185–194. [Google Scholar] [CrossRef] [PubMed]
- Higenyi, J.; Kabasa, J.D. Microbial contamination load of hatching eggs in Butaleja, eastern Uganda. Anim. Vet. Sci. 2014, 2, 22–30. [Google Scholar] [CrossRef]
- Mhone, T.A.; Matope, G.; Saidi, P.T. Aerobic bacterial, coliform, Escherichia coli and Staphylococcus aureus counts of raw and processed milk from selected smallholder dairy farms of Zimbabwe. Int. J. Food Microbiol. 2011, 151, 223–228. [Google Scholar] [CrossRef] [PubMed]
- Lozano, C.; López, M.; Gómez-Sanz, E.; Ruiz-Larrea, F.; Torres, C.; Zarazaga, M. Detection of methicillin-resistant Staphylococcus aureus ST398 in food samples of animal origin in Spain. J. Antimicrob. Chemother. 2009, 64, 1325–1326. [Google Scholar] [CrossRef] [PubMed]
- Benito, D.; Gómez, P.; Lozano, C.; Estepa, V.; Gómez-Sanz, E.; Zarazaga, M.; Torres, C. Genetic lineages, antimicrobial resistance, and virulence in Staphylococcus aureus of meat samples in Spain: Analysis of immune evasion cluster (IEC) genes. Foodborne Pathog. Dis. 2014, 11, 354–356. [Google Scholar] [CrossRef] [PubMed]
- Stegger, M.; Wirth, T.; Andersen, P.S.; Skov, R.L.; de Grassi, A.; Simões, P.M.; Tristan, A.; Petersen, A.; Aziz, M.; Kiil, K.; et al. Origin and evolution of European community-acquired methicillin-resistant Staphylococcus aureus. MBio 2014. [Google Scholar] [CrossRef] [PubMed]
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lozano, C.; Gharsa, H.; Ben Slama, K.; Zarazaga, M.; Torres, C. Staphylococcus aureus in Animals and Food: Methicillin Resistance, Prevalence and Population Structure. A Review in the African Continent. Microorganisms 2016, 4, 12. https://doi.org/10.3390/microorganisms4010012
Lozano C, Gharsa H, Ben Slama K, Zarazaga M, Torres C. Staphylococcus aureus in Animals and Food: Methicillin Resistance, Prevalence and Population Structure. A Review in the African Continent. Microorganisms. 2016; 4(1):12. https://doi.org/10.3390/microorganisms4010012
Chicago/Turabian StyleLozano, Carmen, Haythem Gharsa, Karim Ben Slama, Myriam Zarazaga, and Carmen Torres. 2016. "Staphylococcus aureus in Animals and Food: Methicillin Resistance, Prevalence and Population Structure. A Review in the African Continent" Microorganisms 4, no. 1: 12. https://doi.org/10.3390/microorganisms4010012