
Characteristic Gut Bacteria in High Barley Consuming Japanese Individuals without Hypertension
 , , , , , , ,
, , , , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design and Implementation
2.2. DNA Extraction and 16S rRNA Gene Amplicon Sequencing
2.3. Bioinformatics Analysis
2.4. Statistical Analysis
2.4.1. Calculation of α-Diversity and Intergroup Comparison of Subjects
2.4.2. Comparative Analysis Excluding Participants Undergoing Treatment
2.4.3. Principal Coordinate Analysis
2.4.4. Logistic Regression Analysis
2.4.5. Random Forest Machine Learning
3. Results
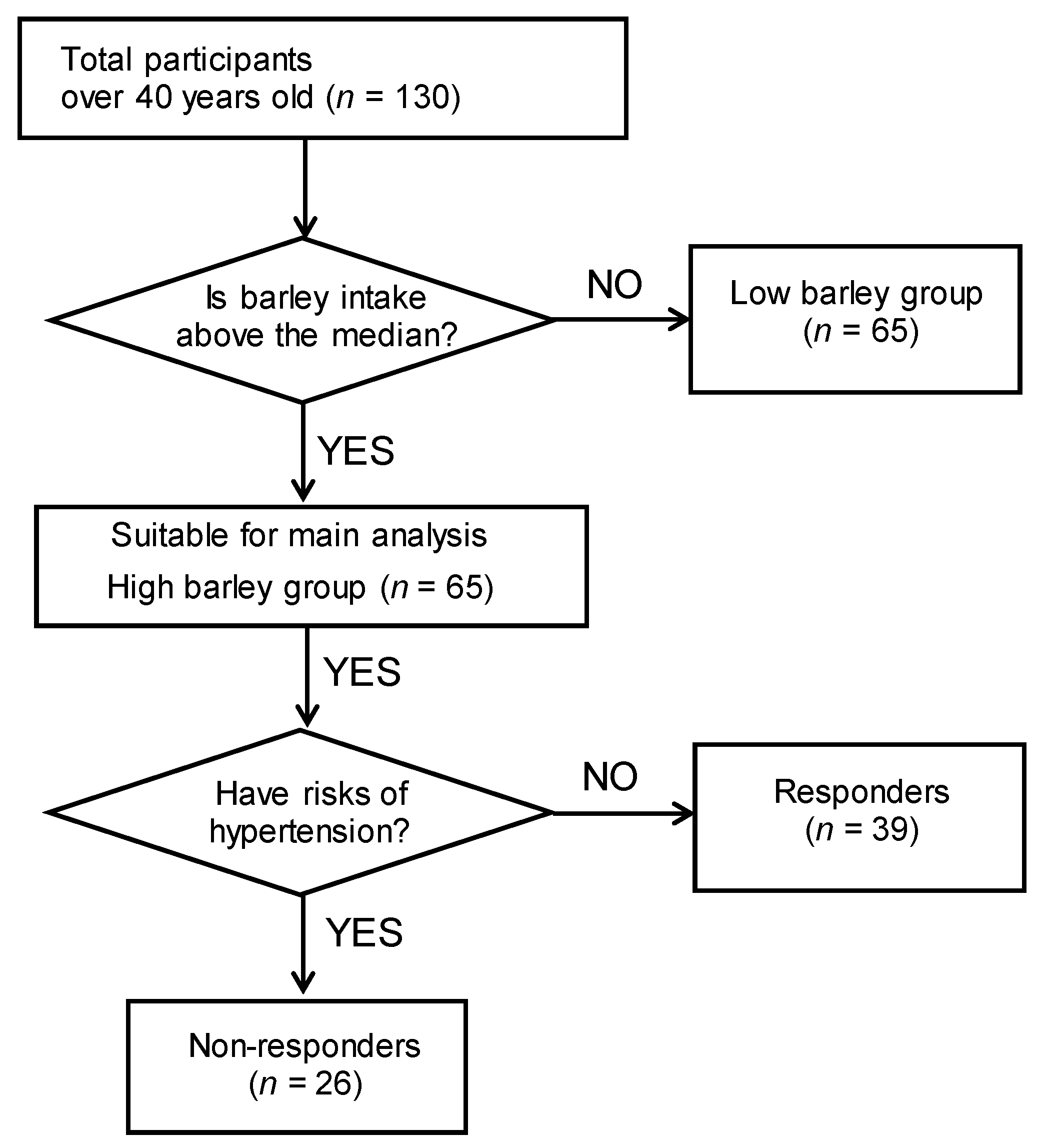
3.1. Subject Characteristics and Barley Responder/Non-Responder Definitions
3.2. Intestinal Bacteria Characteristics of Barley Responders
3.3. Adjusted Relationship between Responders and Characteristic Bacteria
3.4. Determination Model Based on the Gut Bacteria of Barley Responders
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Beaney, T.; Schutte, A.E.; Tomaszewski, M.; Ariti, C.; Burrell, L.M.; Castillo, R.R.; Charchar, F.J.; Damasceno, A.; Kruger, R.; Lackland, D.T.; et al. May Measurement Month 2017: An Analysis of Blood Pressure Screening Results Worldwide. Lancet Glob. Health 2018, 6, e736–e743. [Google Scholar] [CrossRef] [PubMed]
- GBD 2017 RiskFactor Collaborators. Global, Regional, and National Comparative Risk Assessment of 84 Behavioural, Environmental and Occupational, and Metabolic Risks or Clusters of Risks for 195 Countries and Territories, 1990–2017: A Systematic Analysis for the Global Burden of Disease Study 2017. Lancet 2018, 392, 1923–1994. [Google Scholar] [CrossRef]
- Roth, G.A.; Mensah, G.A.; Johnson, C.O.; Addolorato, G.; Ammirati, E.; Baddour, L.M.; Barengo, N.C.; Beaton, A.Z.; Benjamin, E.J.; Benziger, C.P.; et al. Global Burden of Cardiovascular Diseases and Risk Factors, 1990–2019: Update from the GBD 2019 Study. J. Am. Coll. Cardiol. 2020, 76, 2982–3021. [Google Scholar] [CrossRef] [PubMed]
- Giri, A.; Hellwege, J.N.; Keaton, J.M.; Park, J.; Qiu, C.; Warren, H.R.; Torstenson, E.S.; Kovesdy, C.P.; Sun, Y.V.; Wilson, O.D.; et al. Trans-ethnic Association Study of Blood Pressure Determinants in Over 750,000 Individuals. Nat. Genet. 2019, 51, 51–62. [Google Scholar] [CrossRef]
- GBD 2017 Diet Collaborators. Health Effects of Dietary Risks in 195 Countries, 1990–2017: A Systematic Analysis for the Global Burden of Disease Study 2017. Lancet 2019, 393, 1958–1972. [Google Scholar] [CrossRef] [PubMed]
- Cook, N.R.; Appel, L.J.; Whelton, P.K. Sodium Intake and All-Cause Mortality Over 20 Years in the Trials of Hypertension Prevention. J. Am. Coll. Cardiol. 2016, 68, 1609–1617. [Google Scholar] [CrossRef]
- Reynolds, A.; Mann, J.; Cummings, J.; Winter, N.; Mete, E.; Te Morenga, L. Carbohydrate Quality and Human Health: A Series of Systematic Reviews and Meta-Analyses. Lancet 2019, 393, 434–445. [Google Scholar] [CrossRef]
- Whelton, S.P.; Hyre, A.D.; Pedersen, B.; Yi, Y.; Whelton, P.K.; He, J. Effect of Dietary Fiber Intake on Blood Pressure: A Meta-analysis of Randomized, Controlled Clinical Trials. J. Hypertens. 2005, 23, 475–481. [Google Scholar] [CrossRef]
- Behall, K.M.; Scholfield, D.J.; Hallfrisch, J. Whole-Grain Diets Reduce Blood Pressure in Mildly Hypercholesterolemic Men and Women. J. Am. Diet. Assoc. 2006, 106, 1445–1449. [Google Scholar] [CrossRef]
- Marques, F.Z.; Mackay, C.R.; Kaye, D.M. Beyond Gut Feelings: How the Gut Microbiota Regulates Blood Pressure. Nat. Rev. Cardiol. 2018, 15, 20–32. [Google Scholar] [CrossRef]
- Xue, Y.; Cui, L.; Qi, J.; Ojo, O.; Du, X.; Liu, Y.; Wang, X. The Effect of Dietary Fiber (Oat Bran) Supplement on Blood Pressure in Patients with Essential Hypertension: A Randomized Controlled Trial. Nutr. Metab. Cardiovasc. Dis. 2021, 31, 2458–2470. [Google Scholar] [CrossRef] [PubMed]
- Santisteban, M.M.; Qi, Y.; Zubcevic, J.; Kim, S.; Yang, T.; Shenoy, V.; Cole-Jeffrey, C.T.; Lobaton, G.O.; Stewart, D.C.; Rubiano, A.; et al. Hypertension-Linked Pathophysiological Alterations in the Gut. Circ. Res. 2017, 120, 312–323. [Google Scholar] [CrossRef] [PubMed]
- Marques, F.Z.; Nelson, E.; Chu, P.Y.; Horlock, D.; Fiedler, A.; Ziemann, M.; Tan, J.K.; Kuruppu, S.; Rajapakse, N.W.; El-Osta, A.; et al. High-Fiber Diet and Acetate Supplementation Change the Gut Microbiota and Prevent the Development of Hypertension and Heart Failure in Hypertensive Mice. Circulation 2017, 135, 964–977. [Google Scholar] [CrossRef] [PubMed]
- Tilves, C.; Yeh, H.C.; Maruthur, N.; Juraschek, S.P.; Miller, E.; White, K.; Appel, L.J.; Mueller, N.T. Increases in Circulating and Fecal Butyrate are Associated with Reduced Blood Pressure and Hypertension: Results from the SPIRIT Trial. J. Am. Heart Assoc. 2022, 11, e024763. [Google Scholar] [CrossRef]
- Zheng, W.; Ma, Y.; Zhao, A.; He, T.; Lyu, N.; Pan, Z.; Mao, G.; Liu, Y.; Li, J.; Wang, P.; et al. Compositional and Functional Differences in Human Gut Microbiome with Respect to Equol Production and Its Association with Blood Lipid Level: A Cross-Sectional Study. Gut Pathog. 2019, 11, 20. [Google Scholar] [CrossRef]
- Lampe, J.W.; Navarro, S.L.; Hullar, M.A.J.; Shojaie, A. Inter-Individual Differences in Response to Dietary Intervention: Integrating Omics Platforms Towards Personalised Dietary Recommendations. Proc. Nutr. Soc. 2013, 72, 207–218. [Google Scholar] [CrossRef]
- Matsuoka, T.; Hosomi, K.; Park, J.; Goto, Y.; Nishimura, M.; Maruyama, S.; Murakami, H.; Konishi, K.; Miyachi, M.; Kawashima, H.; et al. Relationships Between Barley Consumption and Gut Microbiome Characteristics in a Healthy Japanese Population: A Cross-Sectional Study. BMC Nutr. 2022, 8, 23. [Google Scholar] [CrossRef]
- Goto, Y.; Nishimoto, Y.; Murakami, S.; Nomaguchi, T.; Mori, Y.; Ito, M.; Nakaguro, R.; Kudo, T.; Matsuoka, T.; Yamada, T.; et al. Metabologenomic Approach Reveals Intestinal Environmental Features Associated with Barley-Induced Glucose Tolerance Improvements in Japanese: A Randomized Controlled Trial. Nutrients 2022, 14, 3468. [Google Scholar] [CrossRef]
- Matsuoka, T.; Tsuchida, A.; Yamaji, A.; Kurosawa, C.; Shinohara, M.; Takayama, I.; Nakagomi, H.; Izumi, K.; Ichikawa, Y.; Hariya, N.; et al. Consumption of a Meal Containing Refined Barley Flour Bread is Associated with a Lower Postprandial Blood Glucose Concentration After a Second Meal Compared with One Containing Refined Wheat Flour Bread in Healthy Japanese: A Randomized Control Trial. Nutrition 2020, 72, 110637. [Google Scholar] [CrossRef]
- Osonoi, T.; Matsuoka, T.; Ofuchi, K.; Katoh, M.; Kobayashi, T.; Mochizuki, K. Effects of Barley Intake on Glycemic Control in Japanese Patients with Type 2 Diabetes Mellitus Undergoing Antidiabetic Therapy: A Prospective Study. Diabetol. Int. 2021, 13, 387–395. [Google Scholar] [CrossRef]
- He, L.X.; Zhao, J.; Huang, Y.S.; Li, Y. The Difference Between Oats and Beta-Glucan Extract Intake in the Management of HbA1c, Fasting Glucose and Insulin Sensitivity: A Meta-Analysis of Randomized Controlled Rrials. Food Funct. 2016, 7, 1413–1428. [Google Scholar] [CrossRef] [PubMed]
- Kovatcheva-Datchary, P.; Nilsson, A.; Akrami, R.; Lee, Y.S.; De Vadder, F.; Arora, T.; Hallen, A.; Martens, E.; Björck, I.; Bäckhed, F. Dietary Fiber-Induced Improvement in Glucose Metabolism Is Associated with Increased Abundance of Prevotella. Cell Metab. 2015, 22, 971–982. [Google Scholar] [CrossRef] [PubMed]
- Maruyama, S.; Matsuoka, T.; Hosomi, K.; Park, J.; Nishimura, M.; Murakami, H.; Konishi, K.; Miyachi, M.; Kawashima, H.; Mizuguchi, K.; et al. Classification of the Occurrence of Dyslipidemia Based on Gut Bacteria Related to Barley Intake. Front. Nutr. 2022, 9, 812469. [Google Scholar] [CrossRef]
- Verhaar, B.J.H.; Collard, D.; Prodan, A.; Levels, J.H.M.; Zwinderman, A.H.; Bäckhed, F.; Vogt, L.; Peters, M.J.L.; Muller, M.; Nieuwdorp, M.; et al. Associations between Gut Microbiota, Faecal Short-chain Fatty Acids, and Blood Pressure Across Ethnic Groups: The HELIUS Study. Eur. Heart J. 2020, 41, 4259–4267. [Google Scholar] [CrossRef] [PubMed]
- Onyszkiewicz, M.; Gawrys-Kopczynska, M.; Konopelski, P.; Aleksandrowicz, M.; Sawicka, A.; Koźniewska, E.; Samborowska, E.; Ufnal, M. Butyric Acid, a Gut Bacteria Metabolite, Lowers Arterial Blood Pressure Via Colon-vagus Nerve Signaling and GPR41/43 Receptors. Pflugers Arch. 2019, 471, 1441–1453. [Google Scholar] [CrossRef]
- Hosomi, K.; Ohno, H.; Murakami, H.; Natsume, K.Y.; Tanisawa, K.; Hirata, S.; Suzuki, H.; Nagatake, T.; Nishino, T.; Mizuguchi, K.; et al. Method for Preparing DNA from Feces in Guanidine Thiocyanate Solution Affects 16S rRNA-Based Profiling of Human Microbiota Diversity. Sci. Rep. 2017, 7, 4339. [Google Scholar] [CrossRef]
- Mohsen, A.; Park, J.; Chen, Y.A.; Kawashima, H.; Mizuguchi, K. Impact of Quality Trimming on the Efficiency of Reads Joining and Diversity Analysis of Illumina Paired-End Reads in the Context of QIIME1 and QIIME2 Microbiome Analysis Frameworks. BMC Bioinform. 2019, 20, 581. [Google Scholar] [CrossRef]
- Edgar, R.C. Search and Clustering Orders of Magnitude Faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA Ribosomal RNA Gene Database Project: Improved Data Processing and Web-Based Tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019; Available online: https://www.R-project.org (accessed on 6 March 2023).
- Section of the National Health and Nutrition Survey 2018. Available online: https://www.nibiohn.go.jp/eiken/kenkounippon21/en/eiyouchousa/ (accessed on 6 March 2023).
- Filippini, T.; Malavolti, M.; Whelton, P.K.; Naska, A.; Orsini, N.; Vinceti, M. Blood Pressure Effects of Sodium Reduction: Dose-Response Meta-Analysis of Experimental Studies. Circulation 2021, 143, 1542–1567. [Google Scholar] [CrossRef]
- Huang, L.; Trieu, K.; Yoshimura, S.; Neal, B.; Woodward, M.; Campbell, N.R.C.; Li, Q.; Lackland, D.T.; Leung, A.A.; Anderson, C.A.M.; et al. Effect of Dose and Duration of Reduction in Dietary Sodium on Blood Pressure Levels: Systematic Review and Meta-Analysis of Randomized Trials. BMJ 2020, 368, m315. [Google Scholar] [CrossRef]
- Bartolomaeus, H.; Balogh, A.; Yakoub, M.; Homann, S.; Markó, L.; Höges, S.; Tsvetkov, D.; Krannich, A.; Wundersitz, S.; Avery, E.G.; et al. Short-Chain Fatty Acid Propionate Protects From Hypertensive Cardiovascular Damage. Circulation 2019, 139, 1407–1421. [Google Scholar] [CrossRef]
- Karlsson, F.H.; Fåk, F.; Nookaew, I.; Tremaroli, V.; Fagerberg, B.; Petranovic, D.; Bäckhed, F.; Nielsen, J. Symptomatic Atherosclerosis Is Associated with an Altered Gut Metagenome. Nat. Commun. 2012, 3, 1245. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Jin, J.; Su, X.; Yin, X.; Gao, J.; Wang, X.; Zhang, S.; Bu, P.; Wang, M.; Zhang, Y.; et al. Intestinal Flora Modulates Blood Pressure by Regulating the Synthesis of Intestinal-Derived Corticosterone in High Salt-Induced Hypertension. Circ. Res. 2020, 126, 839–853. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhao, F.; Wang, Y.; Chen, J.; Tao, J.; Tian, G.; Wu, S.; Liu, W.; Cui, Q.; Geng, B.; et al. Gut Microbiota Dysbiosis Contributes to the Development of Hypertension. Microbiome 2017, 5, 14. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Santisteban, M.M.; Rodriguez, V.; Li, E.; Ahmari, N.; Carvajal, J.M.; Zadeh, M.; Gong, M.; Qi, Y.; Zubcevic, J.; et al. Gut Dysbiosis Is Linked to Hypertension. Hypertension 2015, 65, 1331–1340. [Google Scholar] [CrossRef]
- Qin, Y.; Zhao, J.; Wang, Y.; Bai, M.; Sun, S. Specific Alterations of Gut Microbiota in Chinese Patients with Hypertension: A Systematic Review and Meta-Analysis. Kidney Blood Press. Res. 2022, 47, 433–447. [Google Scholar] [CrossRef]
- Mushtaq, N.; Hussain, S.; Zhang, S.; Yuan, L.; Li, H.; Ullah, S.; Wang, Y.; Xu, J. Molecular Characterization of Alterations in the Intestinal Microbiota of Patients with Grade 3 Hypertension. Int. J. Mol. Med. 2019, 44, 513–522. [Google Scholar] [CrossRef]
- Pianta, A.; Arvikar, S.; Strle, K.; Drouin, E.E.; Wang, Q.; Costello, C.E.; Steere, A.C. Evidence of the Immune Relevance of Prevotella copri, a Gut Microbe, in Patients with Rheumatoid Arthritis. Arthritis Rheumatol. 2017, 69, 964–975. [Google Scholar] [CrossRef]
- Alpizar-Rodriguez, D.; Lesker, T.R.; Gronow, A.; Gilbert, B.; Raemy, E.; Lamacchia, C.; Gabay, C.; Finckh, A.; Strowig, T. Prevotella copri in Individuals At Risk for Rheumatoid Arthritis. Ann. Rheum. Dis. 2019, 78, 590–593. [Google Scholar] [CrossRef] [PubMed]
- Scher, J.U.; Sczesnak, A.; Longman, R.S.; Segata, N.; Ubeda, C.; Bielski, C.; Rostron, T.; Cerundolo, V.; Pamer, E.G.; Abramson, S.B.; et al. Expansion of Intestinal Prevotella copri Correlates with Enhanced Susceptibility to Arthritis. eLife 2013, 2, e01202. [Google Scholar] [CrossRef]
- Marungruang, N.; Tovar, J.; Björck, I.; Hållenius, F.F. Improvement in Cardiometabolic Risk Markers Following a Multifunctional Diet Is Associated with Gut Microbial Taxa in Healthy Overweight and Obese Subjects. Eur. J. Nutr. 2018, 57, 2927–2936. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Ames, N.P.; Tun, H.M.; Tosh, S.M.; Jones, P.J.; Khafipour, E. High Molecular Weight Barley β-Glucan Alters Gut Microbiota Toward Reduced Cardiovascular Disease Risk. Front. Microbiol. 2016, 7, 129. [Google Scholar] [CrossRef]
- Martin-Gallausiaux, C.; Marinelli, L.; Blottière, H.M.; Larraufie, P.; Lapaque, N. SCFA: Mechanisms and Functional Importance in the Gut. Proc. Nutr. Soc. 2021, 80, 37–49. [Google Scholar] [CrossRef] [PubMed]
- Natarajan, N.; Hori, D.; Flavahan, S.; Steppan, J.; Flavahan, N.A.; Berkowitz, D.E.; Pluznick, J.L. Microbial Short Chain Fatty Acid Metabolites Lower Blood Pressure via Endothelial G Protein-Coupled Receptor 41. Physiol. Genomics 2016, 48, 826–834. [Google Scholar] [CrossRef]
- Pluznick, J.L.; Protzko, R.J.; Gevorgyan, H.; Peterlin, Z.; Sipos, A.; Han, J.; Brunet, I.; Wan, L.X.; Rey, F.; Wang, T.; et al. Olfactory Receptor Responding to Gut Microbiota-Derived Signals Plays a Role in Renin Secretion and Blood Pressure Regulation. Proc. Natl. Acad. Sci. USA 2013, 110, 4410–4415. [Google Scholar] [CrossRef] [PubMed]
- Yan, Q.; Gu, Y.; Li, X.; Yang, W.; Jia, L.; Chen, C.; Han, X.; Huang, Y.; Zhao, L.; Li, P.; et al. Alterations of the Gut Microbiome in Hypertension. Front. Cell. Infect. Microbiol. 2017, 7, 381. [Google Scholar] [CrossRef]
- Lin, D.; Peters, B.A.; Friedlander, C.; Freiman, H.J.; Goedert, J.J.; Sinha, R.; Miller, G.; Bernstein, M.A.; Hayes, R.B.; Ahn, J. Association of Dietary Fibre Intake and Gut Microbiota in Adults. Br. J. Nutr. 2018, 120, 1014–1022. [Google Scholar] [CrossRef]
- Benus, R.F.J.; van der Werf, T.S.; Welling, G.W.; Judd, P.A.; Taylor, M.A.; Harmsen, H.J.M.; Whelan, K. Association Between Faecalibacterium prausnitzii and Dietary Fibre in Colonic Fermentation in Healthy Human Subjects. Br. J. Nutr. 2010, 104, 693–700. [Google Scholar] [CrossRef]
- Michalak, L.; Gaby, J.C.; Lagos, L.; La Rosa, S.L.; Hvidsten, T.R.; Tétard-Jones, C.; Willats, W.G.T.; Terrapon, N.; Lombard, V.; Henrissat, B.; et al. Microbiota-Directed Fibre Activates Both Targeted and Secondary Metabolic Shifts in the Distal Gut. Nat. Commun. 2020, 11, 5773. [Google Scholar] [CrossRef]
- Holmstrøm, K.; Collins, M.D.; Møller, T.; Falsen, E.; Lawson, P.A. Subdoligranulum variabile Gen. Nov., sp. Nov. From Human Feces. Anaerobe 2004, 10, 197–203. [Google Scholar] [CrossRef] [PubMed]
- Meehan, C.J.; Beiko, R.G. A Phylogenomic View of Ecological Specialization in the Lachnospiraceae, a Family of Digestive Tract-Associated Bacteria. Genome Biol. Evol. 2014, 6, 703–713. [Google Scholar] [CrossRef] [PubMed]
- Vanegas, S.M.; Meydani, M.; Barnett, J.B.; Goldin, B.; Kane, A.; Rasmussen, H.; Brown, C.; Vangay, P.; Knights, D.; Jonnalagadda, S.; et al. Substituting Whole Grains for Refined Grains in a 6-Wk Randomized Trial Has a Modest Effect on Gut Microbiota and Immune and Inflammatory Markers of Healthy Adults. Am. J. Clin. Nutr. 2017, 105, 635–650. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.; Liu, S.; Li, H.; Zhang, Z.; Zhang, Q.; Chen, L.; Zhao, Y.; Chen, Y.; Gu, J.; Min, L.; et al. Identification of Gut Microbiota and Metabolites Signature in Patients with Irritable Bowel Syndrome. Front. Cell. Infect. Microbiol. 2019, 9, 346. [Google Scholar] [CrossRef] [PubMed]
- Forbes, J.D.; Chen, C.Y.; Knox, N.C.; Marrie, R.A.; El-Gabalawy, H.; de Kievit, T.; Alfa, M.; Bernstein, C.N.; Van Domselaar, G.A. Comparative Study of the Gut Microbiota in Immune-Mediated Inflammatory Diseases-Does a Common Dysbiosis Exist? Microbiome 2018, 6, 221. [Google Scholar] [CrossRef] [PubMed]
- Wright, E.K.; Kamm, M.A.; Wagner, J.; Teo, S.M.; De Cruz, P.; Hamilton, A.L.; Ritchie, K.J.; Inouye, M.; Kirkwood, C.D. Microbial Factors Associated with Postoperative Crohn’s Disease Recurrence. J. Crohns Colitis 2017, 11, 191–203. [Google Scholar] [CrossRef] [PubMed]
- Strandwitz, P.; Kim, K.H.; Terekhova, D.; Liu, J.K.; Sharma, A.; Levering, J.; McDonald, D.; Dietrich, D.; Ramadhar, T.R.; Lekbua, A.; et al. GABA-Modulating Bacteria of the Human Gut Microbiota. Nat. Microbiol. 2019, 4, 396–403. [Google Scholar] [CrossRef]
- Maier, L.; Pruteanu, M.; Kuhn, M.; Zeller, G.; Telzerow, A.; Anderson, E.E.; Brochado, A.R.; Fernandez, K.C.; Dose, H.; Mori, H.; et al. Extensive Impact of Non-antibiotic Drugs on Human Gut Bacteria. Nature 2018, 555, 623–628. [Google Scholar] [CrossRef]
- Natali, A.; Ferrannini, E. Hypertension, Insulin Resistance, and the Metabolic Syndrome. Endocrinol. Metab. Clin. 2004, 33, 417–429. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Non-Responders (n = 26) | Responders (n = 39) | p Value (1) | |
|---|---|---|---|
| n (%) or Mean (SD) | n (%) or Mean (SD) | ||
| Male (n) | 24 (92%) | 28 (72%) | 0.09 (2) |
| Age (years) | 53.5 (6) | 48 (6) | 0.002 |
| Barley intake (g/1000 kcal) | 10.5 (5.7) | 8.4 (3.4) | 0.11 |
| Medications of hypertension drug (n) | 7 (27%) | 0 (0%) | <0.001 (2) |
| Parents with hypertension (n) | 21 (81%) | 15 (38%) | 0.002 (2) |
| Medical checkup | |||
| Weight (kg) | 71.9 (9.7) | 65.6 (13.4) | 0.03 |
| BMI (kg/m2) | 24.9 (3.4) | 22.8 (3.4) | 0.02 |
| SBP (mmHg) | 138 (16) | 113 (10) | <0.001 |
| DBP (mmHg) | 92 (8) | 73 (9) | <0.001 |
| Fasting blood glucose (mg/dL) | 100 (17) | 89 (7) | 0.004 |
| HbA1c (%) | 5.7 (0.5) | 5.5 (0.3) | 0.15 |
| Triglycerides (mg/dL) | 149 (75) | 111 (92) | 0.07 |
| HDL-cholesterol (mg/dL) | 57 (15) | 61 (19) | 0.27 |
| LDL-cholesterol (mg/dL) | 133 (27) | 115 (29) | 0.01 |
| Nutrients | |||
| Energy (kcal/d) | 1815 (569) | 1746 (439) | 0.60 |
| Protein (g/d) | 60 (22) | 59 (17) | 0.90 |
| Fat (g/d) | 48 (20) | 52 (18) | 0.34 |
| Carbohydrate (g/d) | 236 (99) | 224 (68) | 0.60 |
| Sodium chloride (g/d) | 10.1 (3.1) | 9.6 (2.4) | 0.49 |
| Lifestyle | |||
| Smoking (present, past, never) (n) | 5, 14, 7 (19%, 54%, 27%) | 12, 12, 15 (31%, 31%, 39%) | 0.18 (2) |
| With physical activity (3) (n) | 4 (23%) | 22 (85%) | 0.66 (2) |
| Non-Responders (n = 26) | Responders (n = 39) | p Value (1) | |
|---|---|---|---|
| Median [Interquartile Range] | Median [Interquartile Range] | ||
| α-Diversity | |||
| Chao1 | 1028 [806, 1331] | 997 [895, 1210] | 0.009 |
| Shannon | 3.47 [3.22, 3.80] | 3.66 [3.32, 3.91] | 0.07 |
| Simpson | 0.92 [0.89, 0.95] | 0.94 [0.90, 0.95] | 0.20 |
| Genus | |||
| Faecalibacterium | 1.74 [0.54, 5.28] | 5.86 [2.09, 9.28] | 0.02 |
| Lachnoclostridium | 2.12 [0.93, 2.84] | 1.13 [0.71, 1.75] | 0.02 |
| Ruminococcaceae UCG-013 | 0.08 [0.003, 0.19] | 0.21 [0.08, 0.50] | 0.03 |
| Lachnospira | 0.15 [0.003, 0.52] | 0.62 [0.22, 1.37] | 0.02 |
| Prevotella 9 | 0.01 [0.00, 24.19] | 0.00 [0.00, 0.01] | 0.03 |
| Subdoligranulum | 0.26 [0.003, 1.39] | 1.69 [0.09, 2.58] | 0.04 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maruyama, S.; Matsuoka, T.; Hosomi, K.; Park, J.; Nishimura, M.; Murakami, H.; Konishi, K.; Miyachi, M.; Kawashima, H.; Mizuguchi, K.; et al. Characteristic Gut Bacteria in High Barley Consuming Japanese Individuals without Hypertension. Microorganisms 2023, 11, 1246. https://doi.org/10.3390/microorganisms11051246
Maruyama S, Matsuoka T, Hosomi K, Park J, Nishimura M, Murakami H, Konishi K, Miyachi M, Kawashima H, Mizuguchi K, et al. Characteristic Gut Bacteria in High Barley Consuming Japanese Individuals without Hypertension. Microorganisms. 2023; 11(5):1246. https://doi.org/10.3390/microorganisms11051246
Chicago/Turabian StyleMaruyama, Satoko, Tsubasa Matsuoka, Koji Hosomi, Jonguk Park, Mao Nishimura, Haruka Murakami, Kana Konishi, Motohiko Miyachi, Hitoshi Kawashima, Kenji Mizuguchi, and et al. 2023. "Characteristic Gut Bacteria in High Barley Consuming Japanese Individuals without Hypertension" Microorganisms 11, no. 5: 1246. https://doi.org/10.3390/microorganisms11051246
APA StyleMaruyama, S., Matsuoka, T., Hosomi, K., Park, J., Nishimura, M., Murakami, H., Konishi, K., Miyachi, M., Kawashima, H., Mizuguchi, K., Kobayashi, T., Ooka, T., Yamagata, Z., & Kunisawa, J. (2023). Characteristic Gut Bacteria in High Barley Consuming Japanese Individuals without Hypertension. Microorganisms, 11(5), 1246. https://doi.org/10.3390/microorganisms11051246