
First Polyphasic Study of Cheffia Reservoir (Algeria) Cyanobacteria Isolates Reveals Toxic Picocyanobacteria Genotype
 ,
,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection of Cyanobacterial Samples and Isolation
2.2. Morphological Characterization
2.3. DNA Extraction, Amplification (PCR) and Sequencing
2.4. Sequence Analysis
2.5. PCR Amplification for Toxin Production
3. Results
3.1. Cyanobacterial Isolation and Morphological Identification
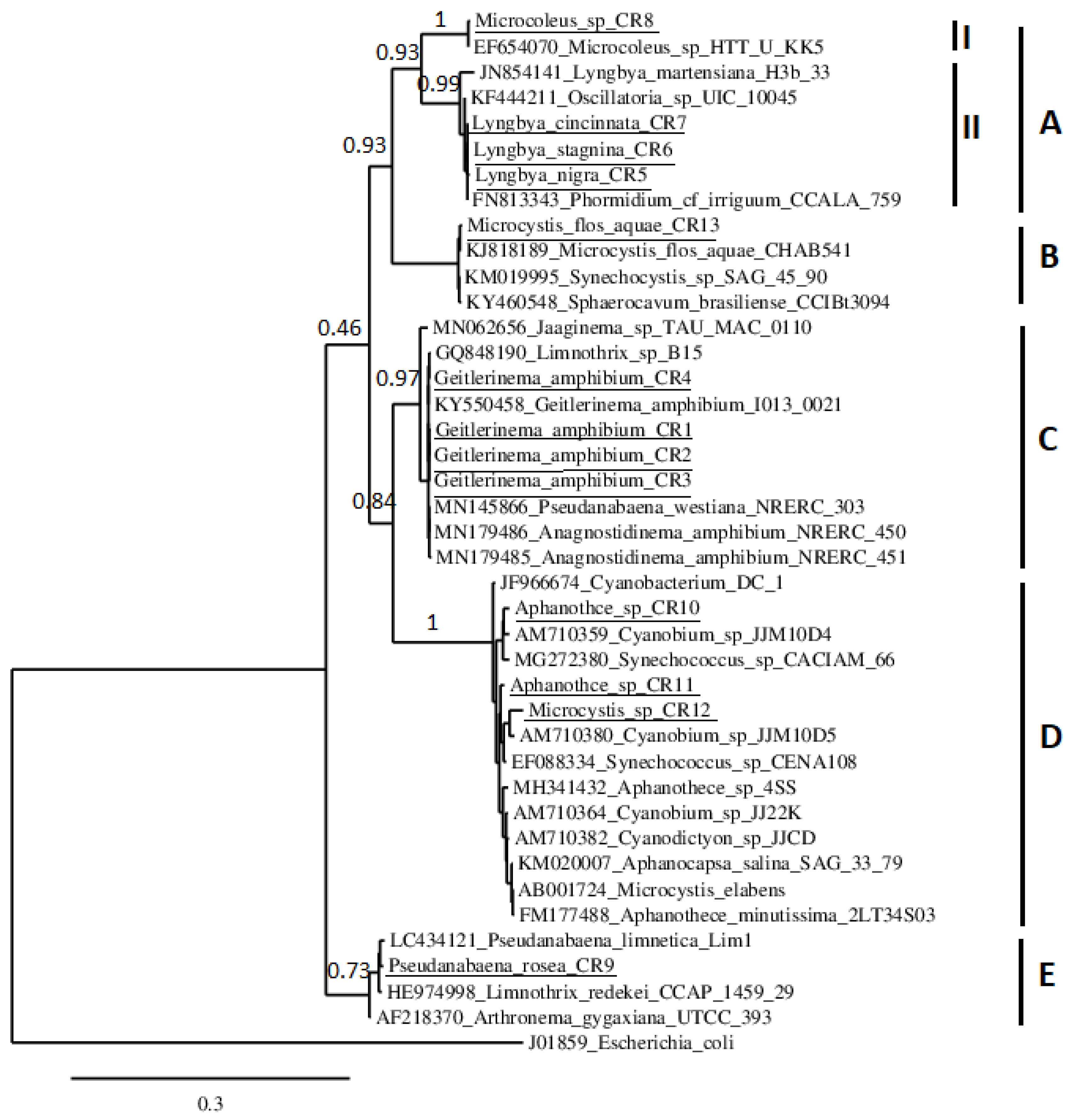
3.2. Phylogenetic Study
3.3. PCR Amplification of Toxin-Encoding Genes
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Svirčev, Z.; Chen, L.; Sántha, K.; Drobac Backović, D.; Šušak, S.; Vulin, A.; Palanački Malešević, T.; Codd, G.A.; Meriluoto, J. A Review and assessment of cyanobacterial toxins as cardiovascular health hazards. Arch. Toxicol. 2022, 96, 2829–2863. [Google Scholar] [CrossRef] [PubMed]
- Dittmann, E.; Fewer, D.P.; Neilan, B.A. Cyanobacterial toxins: Biosynthetic routes and evolutionary roots. FEMS Microbiol. Rev. 2013, 37, 23–43. [Google Scholar] [CrossRef] [PubMed]
- Jochimsen, E.M.; Carmichael, W.W.; An, J.; Cardo, D.M.; Cookson, S.T.; Holmes, C.E.M.; Antunes, M.B.; de Melo Filho, D.A.; Lyra, T.M.; Barreto, V.S.T.; et al. Liver failure and death after exposure to microcystins at a hemodialysis center in Brazil. N. Engl. J. Med. 1998, 338, 873–878. [Google Scholar] [CrossRef] [PubMed]
- Chorus, I.; Welker, M. Toxic Cyanobacteria in Water: A Guide to Their Public Health Consequences, Monitoring and Management, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2021. [Google Scholar] [CrossRef]
- Zamyadi, A.; Glover, C.M.; Yasir, A.; Stuetz, R.; Newcombe, G.; Crosbie, N.D.; Lin, T.F.; Henderson, R. Toxic cyanobacteria in water supply systems: Data analysis to map global challenges and demonstrate the benefits of multi-barrier treatment approaches. H2Open J. 2021, 4, 47–62. [Google Scholar] [CrossRef]
- Zamyadi, A.; Dorner, S.; Sauve, S.; Ellis, D.; Bolduc, A.; Bastien, C.; Prevost, M. Species-dependence of cyanobacteria removal efficiency by different drinking water treatment processes. Water Res. 2013, 47, 2689–2700. [Google Scholar] [CrossRef]
- Castenholz, R.W. Species usage, concept, and evolution in the cyanobacteria (Blue-Green Algae). J. Phycol. 1992, 28, 737–745. [Google Scholar] [CrossRef]
- Palinska, K.A.; Liesack, W.; Rhiel, E.; Krumbein, W.E. Phenotype variability of identical genotypes: The need for a combined approach in cyanobacterial taxonomy demonstrated on Merismopedia-like isolates. Arch. Microbiol. 1996, 166, 224–233. [Google Scholar] [CrossRef]
- Komárek, J. A polyphasic approach for the taxonomy of cyanobacteria: Principles and applications. Eur. J. Phycol. 2016, 51, 346–353. [Google Scholar] [CrossRef]
- El Semary, N.A. Modern methods for detection and elimination of microcystins toxins produced by cyanobacteria: Mini-review. J. Appl. Sci. 2010, 10, 1662–1666. [Google Scholar] [CrossRef]
- Yuan, J.; Yoon, K.-J. Overview of PCR methods applied for the identification of freshwater toxigenic cyanobacteria. In Cyanobacteria-Recent Advances in Taxonomy and Applications; IntechOpen: London, UK, 2021. [Google Scholar]
- Benredjem, L.; Berredjem, H.; Abdi, A.; Casero, M.C.; Quesada, A.; Fosso, B.; Marzano, M.; Pesole, G.; Azevedo, J.; Vasconcelos, V. Morphological, molecular, and biochemical study of cyanobacteria from a eutrophic Algerian Reservoir (Cheffia). SPR 2022, 29, 27624–27635. [Google Scholar] [CrossRef]
- Saoudi, A.; Brient, L.; Boucetta, S.; Ouzrout, R.; Bormans, M.; Bensouilah, M. Management of toxic cyanobacteria for drinking water production of Ain Zada Dam. Environ. Monit. Assess. 2017, 189, 361. [Google Scholar] [CrossRef] [PubMed]
- Touati, H.; Guellati, F.Z.; Arif, S.; Bensouilah, M. Cyanobacteria dynamics in a Mediterranean Reservoir of the North East of Algeria: Vertical and seasonal variability. JEE 2019, 20, 93–107. [Google Scholar] [CrossRef]
- Bryant, D.A. A brief history of cyanobacterial research: Past, present, and future prospects. In The Cell Biology of Cyanobacteria; Flores, E., Herrero, A., Eds.; Caister Academic Press: Norfolk, UK, 2014; pp. 1–6. [Google Scholar]
- Komárek, J.; Kaštovský, J.; Mareš, J.; Johansen, J.R. Taxonomic classification of cyanoprokaryotes (cyanobacterial genera) 2014, Using a Polyphasic Approach. Preslia 2014, 86, 295–335. [Google Scholar]
- Genuário, D.B.; Vieira Vaz, M.G.M.; Hentschke, G.S.; Sant’ Anna, C.L.; Fiore, M.F. Halotia gen. nov., a phylogenetically and physiologically coherent cyanobacterial genus isolated from marine coastal environments. Int. J. Syst. Evol. Microbiol. 2015, 65, 633–675. [Google Scholar] [CrossRef]
- Hauer, T.; Bohunická, M. Reassessment of the cyanobacterial family Microchaetaceae and establishment of new families Tolypothrichaceae and Godleyaceae. J. Phycol. 2014, 50, 1089–1100. [Google Scholar] [CrossRef]
- Mai, T.; Johansen, J.R.; Pietrasiak, N.; Bohunická, M.; Martin, M.P. Revision of the Synechococcales (cyanobacteria) through recognition of four families including Oculatellaceae fam. nov. and Trichocoleaceae fam. nov. and six new genera containing 14 species. Phytotaxa 2018, 365, 1–59. [Google Scholar] [CrossRef]
- Raabová, L.; Kovacik, L.; Elster, J.; Strunecký, O. Review of the genus Phormidesmis (Cyanobacteria) based on environmental, morphological, and molecular data with description of a new genus Leptodesmis. Phytotaxa 2019, 395, 1–16. [Google Scholar] [CrossRef]
- Rippka, E.; Deruelles, J.; Waterbury, N.B. Generic assignments, strain histories and properties of pure cultures of cyanobacteria. J. Gen. Microbiol. 1979, 111, 1–61. [Google Scholar] [CrossRef]
- Rippka, R. Isolation and Purification of Cyanobacteria. Methods. Enzymol. 1988, 167, 3–27. [Google Scholar]
- Boukerb, S. Vulnerability to Pollution of the Aquifer of the Dune Massif of Bououteldja (El-Taref). Master’s Thesis, Mouloud Mammeri University of Tizi-Ouzou, Tizi Ouzou, Algerie, 15 March 2021. [Google Scholar]
- Komárek, J.; Anagnostidis, K. Cyanoprokaryota 2. In Teil/2nd Part: Oscillatoriales; Büdel, B., Krienitz, L., Gärtner, G., Schagerl, M., Eds.; Elsevier/Spektrum: Heidelberg, Germany, 2005; p. 759. [Google Scholar]
- Komárek, J.; Anagnostidis, K. Cyanoprokaryota 1. In Teil: Chroococcales; Ettl, H., Gärtner, G., Heynig, H., Mollenhauer, D., Eds.; Spektrum Akademischer Verlag: Heidelberg, Germany, 1999; p. 548. [Google Scholar]
- Nu¨bel, U.; Nu¨bel, N.; Garcia-Pichel, F.; Muyzer, G. PCR primers to Amplify 16S rRNA genes from cyanobacteria. Appl. Environ. Microbiol. 1997, 63, 3327–3332. [Google Scholar] [CrossRef]
- Neilan, B.A.; Jacobs, D.; Therese, D.D.; Blackall, L.L.; Hawkins, P.R.; Cox, P.T.; Goodman4, A.E. rRNA sequences and evolutionary relationships among toxic and nontoxic cyanobacteria of the genus Microcystis. Int. J. Syst. Bacteriol. 1997, 47, 693–697. [Google Scholar] [CrossRef] [PubMed]
- Dereeper, A.; Guignon, V.; Blanc, G.; Audic, S.; Buffet, S.; Chevenet, F.; Dufayard, J.F.; Guindon, S.; Lefort, V.; Lescot, M.; et al. Phylogeny.Fr: Robust phylogenetic analysis for the non-specialist. Nucleic. Acids Res. 2008, 36, W465–W469. [Google Scholar] [CrossRef] [PubMed]
- Jungblut, A.D.; Neilan, B.A. Molecular identification and evolution of the cyclic peptide hepatotoxins, microcystin and nodulain, synthetase genes in three orders of cyanobacteria. Arch. Microbiol. 2006, 185, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Lopes, V.R.; Ramos, V.; Martins, A.; Sousa, M.; Welker, M.; Antunes, A.; Vasconcelos, V.M. Phylogenetic, chemical and morphological diversity of cyanobacteria from Portuguese temperate estuaries. Mar. Environ. Res. 2012, 73, 7–16. [Google Scholar] [CrossRef]
- Mihali, T.K.; Kellmann, R.; Muenchhoff, J.; Barrow, K.D.; Neilan, B.A. Characterization of the gene cluster responsible for cylindrospermopsin biosynthesis. Appl. Environ. Microbiol. 2008, 74, 716–722. [Google Scholar] [CrossRef]
- Rantala-Ylinen, A.; Känä, S.; Wang, H.; Rouhiainen, L.; Wahlsten, M.; Rizzi, E.; Berg, K.; Gugger, M.; Sivonen, K. Anatoxin-a synthetase gene cluster of the cyanobacterium Anabaena sp. Strain 37 and molecular methods to detect potential producers. Appl. Eviron. Microbiol. 2011, 77, 7271–7278. [Google Scholar] [CrossRef]
- Saker, M.L.; Welker, M.; Vasconcelos, V.M. Multiplex PCR for the detection of toxigenic cyanobacteria in dietary supplements poduced for human consumption. Appl. Microbiol. Biotechnol. 2007, 73, 1136–1142. [Google Scholar] [CrossRef]
- Savela, H.; Spoof, L.; Perälä, N.; Preede, M.; Lamminmäki, U.; Nybom, S.; Häggqvist, K.; Meriluoto, J.; Vehniäinen, M. Detection of cyanobacterial Sxt genes and paralytic shellfish toxins in freshwater lakes and brackish waters on Åland Islands, Finland. Harmful Algae 2015, 46, 1–10. [Google Scholar] [CrossRef]
- Hisbergues, M.; Christiansen, G.; Rouhiainen, L.; Sivonen, K.; Börner, T. PCR-based identification of microcystin-producing genotypes of different cyanobacterial genera. Arch. Microbiol. 2003, 180, 402–410. [Google Scholar] [CrossRef]
- Do Carmo Bittencourt-Oliveira, M.; Do Nascimento Moura, A.; De Oliveira, M.C.; Sidnei Massola, N. Geitlerinema species (Oscillatoriales, Cyanobacteria) revealed by cellular morphology, ultrastructure, and DNA sequencing. J. Phycol. 2009, 45, 716–725. [Google Scholar] [CrossRef]
- Bittencourt-Oliveira, M.d.C.; de Oliveira, M.C.; Bolch, C.J.S. Genetic variability of Brazilian strains of the Microcystis aeruginosa complex (Cyanobacteria/Cyanophyceae) using the phycocyanin intergenic spacer and flanking regions (Cpc BA) 1. J. Phycol. 2001, 37, 810–818. [Google Scholar] [CrossRef]
- Haande, S.; Ballot, A.; Rohrlack, T.; Fastner, J.; Wiedner, C.; Edvardsen, B. Diversity of Microcystis aeruginosa isolates (Chroocccales, Cyanobacteria) from East-African water bodies. Arch. Microbiol. 2007, 188, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Otsuka, S.; Suda, S.; Li, R.; Matsumoto, S.; Watanabe, M.M. Morphological variability of colonies of Microcystis morphospecies in culture. J. Gen. Appl. Microbiol. 2000, 46, 39–50. [Google Scholar] [CrossRef]
- Zapomělová, E.; Hrouzek, P.; Řeháková, K.; Sabacká, M.; Stibal, M.; Caisová, L.; Komárková, J.; Lukesová, A. Morphological variability in selected heterocystous cyanobacterial strains as a response to varied temperature, light intensity and medium composition. Folia Microbiol. 2008, 53, 333–341. [Google Scholar] [CrossRef] [PubMed]
- Miller, S.R.; Longley, R.; Hutchins, P.R.; Bauersachs, T. Cellular innovation of the cyanobacterial heterocyst by the adaptive loss of plasticity. Curr. Biol. 2020, 20, 344–350. [Google Scholar] [CrossRef] [PubMed]
- Holt, J.G.; Krieg, N.R.; Sneath, P.H.A.; Staley, J.T.; Williams, S.T. Group 11. Oxygenic phototrophic bacteria. In Bergey’s Manual of Determinative Bacteriology; Hensyl, W.R., Ed.; Williams & Wilkins: London, UK; Baltimore, MD, USA, 1994; pp. 377–425. [Google Scholar]
- Komárek, J. Recent changes (2008) in cyanobacteria taxonomy based on a combination of molecular background with phenotype and ecological consequences (genus and species concept). Hydrobiologia 2010, 639, 245–259. [Google Scholar] [CrossRef]
- Lee, E.; Ryan, U.M.; Monis, P.; McGregor, G.B.; Bath, A.; Gordon, C.; Paparini, A. Polyphasic identification of cyanobacterial isolates from Australia. Water Res. 2014, 59, 248–261. [Google Scholar] [CrossRef]
- Litvaitis, M.K. A molecular test of cyanobacterial phylogeny: Inferences from constraint analyses. Hydrobiologia 2002, 468, 135–145. [Google Scholar] [CrossRef]
- Willame, R.; Boutte, C.; Grubisic, S.; Wilmotte, A.; Komárek, J.; Hoffmann, L. Morphological and molecular characterization of planktonic cyanobacteria from Belgium and Luxembourg. J. Phycol. 2006, 42, 1312–1332. [Google Scholar] [CrossRef]
- Komárek, J. Cyanobacterial Taxonomy: Current problems and prospects for the integration of traditional and molecular approaches. Algae 2006, 21, 349–375. [Google Scholar] [CrossRef]
- Fox, G.E.; Wisotzkey, J.D.; Jurtshuk, P.J. How close is close: 16S rRNA sequence identity may not be sufficient to guarantee species identity. Int. J. Syst. Bacteriol. 1992, 42, 166–170. [Google Scholar] [CrossRef] [PubMed]
- Engene, N.; Cameron Coates, R.; Gerwick, W.H. 16S rRNA gene heterogeneity in the filamentous marine cyanobacterial genus Lyngbya. J. Phycol. 2010, 46, 591–601. [Google Scholar] [CrossRef]
- Fathalli, A.; Ben Rejeb Jenhani, A.; Moreira, C.; Welker, M.; Romdhane, M.; Antunes, A.; Vasconcelos, V. Molecular and phylogenetic characterization of potentially toxic cyanobacteria in Tunisian freshwaters. Syst. Appl. Microbiol. 2011, 34, 303–310. [Google Scholar] [CrossRef]
- Premanandh, J.; Priya, B.; Teneva, I.; Dzhambazov, B.; Prabaharan, D.; Uma, L. Molecular characterization of marine cyanobacteria from the Indian subcontinent deduced from sequence analysis of the phycocyanin operon (CpcB-IGS-CpcA) and 16S-23S ITS region. J. Microbiol. 2006, 44, 607–616. [Google Scholar] [PubMed]
- Strunecky, O.; Pavlovna Ivanova, A.; Mares, J. An updated classification of cyanobacterial orders and families based on phylogenomic and polyphasic analysis. J. Phycol. 2023, 59, 12–51. [Google Scholar] [CrossRef] [PubMed]
- Wanigatunge, R.P.; Magana-Arachchi, D.N.; Chandrasekharan, N.V.; Kulasooriya, S.A. Genetic diversity and molecular phylogeny of cyanobacteria from Sri Lanka based on 16S rRNA gene. Environ. Eng. Res. 2014, 19, 317–329. [Google Scholar] [CrossRef]
- Domingos, P.; Rubim, T.K.; Molica, R.J.R.; Azevedo, S.M.F.O.; Carmichael, W.W. First report of microcystin production by picoplanktonic cyanobacteria isolated from a Northeast Brazilian drinking water supply. Environ. Toxicol. 1999, 14, 3135. [Google Scholar] [CrossRef]
- Žegura, B. An overview of the mechanisms of microcystin-LR Genotoxicity and potential carcinogenicity. Mini Rev. Med. Chem. 2016, 16, 1042–1062. [Google Scholar] [CrossRef]
- Jakubowska, N.; Szeląg-Wasielewska, E. Toxic picoplanktonic cyanobacteria-review. Mar. Drugs 2015, 13, 1497–1518. [Google Scholar] [CrossRef]
- Śliwińska-Wilczewska, S.; Maculewicz, J.; Felpeto, A.B.; Latała, A. Allelopathic and bloom-forming picocyanobacteria in a changing world. Toxins 2018, 10, 48. [Google Scholar] [CrossRef]
- Schmidt, K.; Birchill, A.J.; Atkinson, A.; Brewin, R.J.W.; Clark, J.R.; Hickman, A.E.; Johns, D.G.; Lohan, M.C.; Milne, A.; Pardo, S.; et al. Increasing picocyanobacteria success in shelf waters contributes to long-term food web degradation. Glob. Chang. Biol. 2020, 26, 5574–5587. [Google Scholar] [CrossRef] [PubMed]
- Mazur-Marzec, H.; Sutryk, K.; Kobos, J.; Hebel, A.; Hohlfeld, N.; Błaszczyk, A.; Toruńska, A.; Kaczkowska, M.J.; Łysiak-Pastuszak, E.; Kraśniewski, W.; et al. Occurrence of cyanobacteria and cyanotoxin in the Southern Baltic Proper. Filamentous cyanobacteria versus single-celled picocyanobacteria. Hydrobiologia 2013, 701, 235–252. [Google Scholar] [CrossRef]
- Stockner, J.; Callieri, C.; Cronberg, G. Picoplankton and other non-bloom-forming cyanobacteria in lakes. In The Ecology of Cyanobacteria: Their Diversity in Time and Space; Whitton, B.A., Potts, M., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2000; pp. 195–231. [Google Scholar]
- Sorokin, Y.I.; Zakuskina, O.Y. Features of the Comacchio ecosystem transformed during persistent bloom of picocyanobacteria. J. Oceanogr. 2010, 66, 373–387. [Google Scholar] [CrossRef]
- Munari, C.; Rossi, R.; Mistri, M. Temporal trends in macrobenthos community structure and redundancy in a shallow Coastal Lagoon (Valli Di Comacchio, Northern Adriatic Sea). Hydrobiologia 2005, 550, 95–104. [Google Scholar] [CrossRef]
- Felpeto, A.B.; Śliwińska-Wilczewska, S.; Klin, M.; Konarzewska, Z.; Vasconcelos, V. Temperature-dependent impacts of allelopathy on growth, pigment, and lipid content between a subpolar strain of Synechocystis sp. CCBA MA-01 and coexisting microalgae. Hydrobiologia 2019, 835, 117–128. [Google Scholar] [CrossRef]
- Zua, J.A.; Legrand, C.; Farnelid, H. Seasonal dynamics in picocyanobacterial abundance and clade composition at coastal and offshore stations in the Baltic Sea. Sci. Rep. 2022, 12, 14330. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target | Primer Pair | Sequence (5′−3′) | Size (bp) | Reference |
|---|---|---|---|---|
| mcyA | mcyA-Cd1F mcyA-Cd1R | AAAATTAAAAGCCGTATCAAA AAAAGTGTTTTATTAGCGGCTCAT | 297 | [35] |
| mcyE | HEPF HEPR | TTTGGGGTTAACTTTTTTGGGCATAGTC AATTCTTGAGGCTGTAAATCGGGTTT | 472 | [29] |
| sxtA | sxtA855F sxtA1480R | GACTCGGCTTGTTGCTTCCCC GCCAAACTCGCAACAGGAGAAGG | 648 | [34] |
| sxtG | sxtG432F sxtG928R | AATGGCAGATCGCAACCGCTAT ACATTCAACCCTGCCCATTCACT | 519 | [34] |
| sxtI | sxtI 682F sxtI 877R | GGATCTCAAAGAAGATGGCA GCCAAACGCAGTACCACTT | 200 | [30] |
| cyrJ | cynsulF cylnamR | ACTTCTCTCCTTTCCCTATC GAGTGAAAATGCGTAGAACTTG | 584 | [31] |
| anaC | anaC-genF anaC-genR | TCTGGTATTCAGTCCCCTCTAT CCCAATAGCCTGTCATCAA | 366 | [32] |
| Isolates | Trichome Description | Cell Description | Dimensions (µm) |
|---|---|---|---|
| G. amphibium CR1; CR2; CR3; CR4 | Bright blue-green trichomes, motile, without sheaths, more or less straight, not constricted at cross walls | Cells cylindrical | L = 1.44–4.73 W = 1.38–2.92 |
| L. nigra CR5 | Straight or slightly curved trichomes, constricted at the cross-walls, with calyptras, isopolar with sheaths. Sheaths thin and firm, open at the end, colorless or bluish | Cells are short, discoid, with granular content and aerotopes | L = 1.95–5.15 W = 12.20–15.93 |
| L. stagnina CR6 | Trichomes isopolar. Sheaths thin and firm, distinct, colorless, open at the ends. Filaments constricted at cross-walls and motile by gliding | Cells grayish blue-green, distinctly shorter than wide | L = 1.9–5.2 W = 13.00–19.36 |
| L. cincinnata CR7 | Cylindrical, slightly waved trichomes, constrictions at cross wall, isopolar. Sheaths firm, thick, colorless, lamellated and opened | Cells discoid, cell content is granulated | L = 2.09–4.62 W = 13.46–19.66 |
| Microcoleus sp. CR8 | Wavy or screw-like coiled trichomes, constricted at cross walls, motile, isopolar and delimited by firm and thickened sheaths, homogenous, open at the ends, hyaline | Cells mostly isodiametric or slightly longer than wide. Apical cells are conical without calyptras | L = 3.09–6.02 W = 3.08–5.17 |
| P. rosea CR9 | Straight or slightly curved trichomes, reddish violet, without sheath, with constrictions at the cross walls | Cells cylindrical without aerotopes | L = 1.67–3.89 W = 1.61–2.67 |
| Isolates | Colony Description | Mucilage | Cell Description | Cell Diameter (µm) |
|---|---|---|---|---|
| Aphanothece sp. CR10 | Irregular, microscopic to macroscopic colonies, blue-green | Thin, colorless mucilage and distinct at the margin | Cells mainly spherical or oval, with gas vesicle, bright blue-green content, and delimited with individual envelopes. Envelopes are firm, colored in dark blue-green | 1.07–1.98 |
| Aphanothece sp. CR11 | Spherical colonies, rough in outline, microscopic, free-living, forming macroscopic granular agglomerations | Mucilage colorless and homogeneous, delimited at the margin, and follows the irregular outline of the colony, not diffluent, without a refractive outline | Cells spherical, pale or bright blue-green, slightly distant from one another, having fine granulation, without gas vesicles, and enveloped by thin individual layer | 0.90–1.36 |
| Microcystis sp. CR12 | Colonies macroscopic, lenticular, slightly elongate, three-dimensional, agglomerated in macroscopic, free-floating, gelatinous, blue-green masses | Narrow, colorless mucilage, distinctly delimited along cell agglomerations and forming refractive outline | Cells spherical, densely aggregated, with individual envelopes, content is homogeneous, olive green or brownish with aerotopes | 4.02–5.97 |
| M. flos-aquae CR13 | Spherical colonies, with irregular margins, microscopic to macroscopic, free-floating, compact, or clathrate, with densely irregularly arranged cells gathered in small agglomerations | Mucilage colorless, slightly distant from cell clusters, and delimited by slightly refractive outline | Cells spherical or hemispherical after division, with individual thick envelopes. Cell content appears granular, olive green, or brownish, with aerotopes | 3.98–5.77 |
| Isolates | Closest Match (Accession Number) | Query Coverage (%) | Percent Identity (%) |
|---|---|---|---|
| G. amphibium CR1 | Geitlerinema amphibium I013-0021 (KY550458) | 82 | 99.39 |
| Anagnostidinema amphibium NRERC-450 (MN179486) | 98 | 99.13 | |
| Pseudanabaena westiana NRERC-303 (MN145866) | 98 | 99.13 | |
| Limnothrix sp. B15 (GQ848190) | 98 | 99.13 | |
| G. amphibium CR2 | Geitlerinema amphibium I013-0021 (KY550458) | 83 | 99.74 |
| Pseudanabaena westiana NRERC-303 (MN145866) | 99 | 99.56 | |
| Limnothrix sp. B15 (GQ848190) | 100 | 99.56 | |
| Anagnostidinema amphibium NRERC-451 (MN179485) | 100 | 99.49 | |
| Jaaginema sp. TAU-MAC 0110 (MN062656) | 96 | 97.59 | |
| G. amphibium CR3 | Geitlerinema amphibium I013-0021 (KY550458) | 85 | 99.91 |
| Anagnostidinema amphibium NRERC-450 (MN179486) | 100 | 99.70 | |
| Pseudanabaena westiana NRERC-303 (MN145866) | 100 | 99.70 | |
| Limnothrix sp. B15 (GQ848190) | 100 | 99.70 | |
| Jaaginema sp. TAU-MAC 0110 (MN062656) | 97 | 97.71 | |
| G. amphibium CR4 | Geitlerinema amphibium I013-0021 (KY550458) | 85 | 99.91 |
| Limnothrix sp. B15 (GQ848190) | 100 | 99.85 | |
| Anagnostidinema amphibium NRERC-450 (MN179486) | 100 | 99.70 | |
| Pseudanabaena westiana NRERC-303 (MN145866) | 100 | 99.70 | |
| Jaaginema sp. TAU-MAC 0110 (MN062656) | 98 | 97.73 | |
| L. nigra CR5 | Phormidium cf. irriguum CCALA 759 (FN813343) | 99 | 99.85 |
| Oscillatoria sp. UIC 10045 (KF444211) | 93 | 99.35 | |
| Lyngbya martensiana H3b/33 (JN854141) | 95 | 97.78 | |
| L. stagnina CR6 | Phormidium cf. irriguum CCALA 759 (FN813343) | 97 | 99.85 |
| Oscillatoria sp. UIC 10045 (KF444211) | 90 | 99.28 | |
| Lyngbya martensiana H3b/33 (JN854141) | 92 | 97.73 | |
| L. cincinnata CR7 | Phormidium cf. irriguum CCALA 759 (FN813343) | 97 | 99.63 |
| Oscillatoria sp. UIC 10045 (KF444211) | 91 | 99.04 | |
| Lyngbya martensiana H3b/33 (JN854141) | 93 | 97.50 | |
| Microcoleus sp. CR8 | Microcoleus sp. HTT-U-KK5 (EF654070) | 99 | 99.40 |
| P. rosea CR9 | Pseudanabaena limnetica Lim1 (LC434121) | 100 | 99.27 |
| Limnothrix redekei CCAP 1459/29 (HE974998) | 100 | 99.05 | |
| Arthronema gygaxiana UTCC 393 (AF218370) | 100 | 98.39 | |
| Aphanothece sp. CR10 | Synechococcus sp. CACIAM 66 (MG272380) | 100 | 99.11 |
| Cyanobium sp. JJM10D4 (AM710359) | 100 | 98.96 | |
| Aphanothece minutissima 2LT34S03 (FM177488) | 100 | 97.41 | |
| Microcystis elabens (AB001724) | 100 | 97.26 | |
| Aphanothece sp. 4SS (MH341432) | 86 | 97.79 | |
| Aphanothece sp. CR11 | Synechococcus sp. CENA108 (EF088334) | 100 | 98.96 |
| Cyanobium sp. JJ22K (AM710364) | 100 | 98.30 | |
| Cyanobacterium DC-1 (JF966674) | 100 | 98.30 | |
| Cyanodictyon sp. JJCD (AM710382) | 100 | 98.22 | |
| Aphanocapsa salina SAG 33.79 (KM020007) | 100 | 98.07 | |
| Aphanothece sp. 4SS (MH341432) | 87 | 97.89 | |
| Microcystis sp. CR12 | Synechococcus sp. CENA108 (EF088334) | 100 | 98.08 |
| Cyanobium sp. JJM10D5 (AM710380) | 100 | 98.00 | |
| M. flos-aquae CR13 | Microcystis flos-aquae CHAB541 (KJ818189) | 100 | 99.77 |
| Sphaerocavum brasiliense CCIBt3094 (KY460548) | 100 | 99.32 | |
| Synechocystis sp. SAG 45.90 (KM019995) | 100 | 99.47 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Benredjem, L.; Morais, J.; Hentschke, G.S.; Abdi, A.; Berredjem, H.; Vasconcelos, V. First Polyphasic Study of Cheffia Reservoir (Algeria) Cyanobacteria Isolates Reveals Toxic Picocyanobacteria Genotype. Microorganisms 2023, 11, 2664. https://doi.org/10.3390/microorganisms11112664
Benredjem L, Morais J, Hentschke GS, Abdi A, Berredjem H, Vasconcelos V. First Polyphasic Study of Cheffia Reservoir (Algeria) Cyanobacteria Isolates Reveals Toxic Picocyanobacteria Genotype. Microorganisms. 2023; 11(11):2664. https://doi.org/10.3390/microorganisms11112664
Chicago/Turabian StyleBenredjem, Lamia, João Morais, Guilherme Scotta Hentschke, Akila Abdi, Hajira Berredjem, and Vitor Vasconcelos. 2023. "First Polyphasic Study of Cheffia Reservoir (Algeria) Cyanobacteria Isolates Reveals Toxic Picocyanobacteria Genotype" Microorganisms 11, no. 11: 2664. https://doi.org/10.3390/microorganisms11112664