
Fungi Can Be More Effective than Bacteria for the Bioremediation of Marine Sediments Highly Contaminated with Heavy Metals

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area and Sampling
2.2. Sediment Remediation Experimental Setup, Microbial Strains Used, and pH Determination
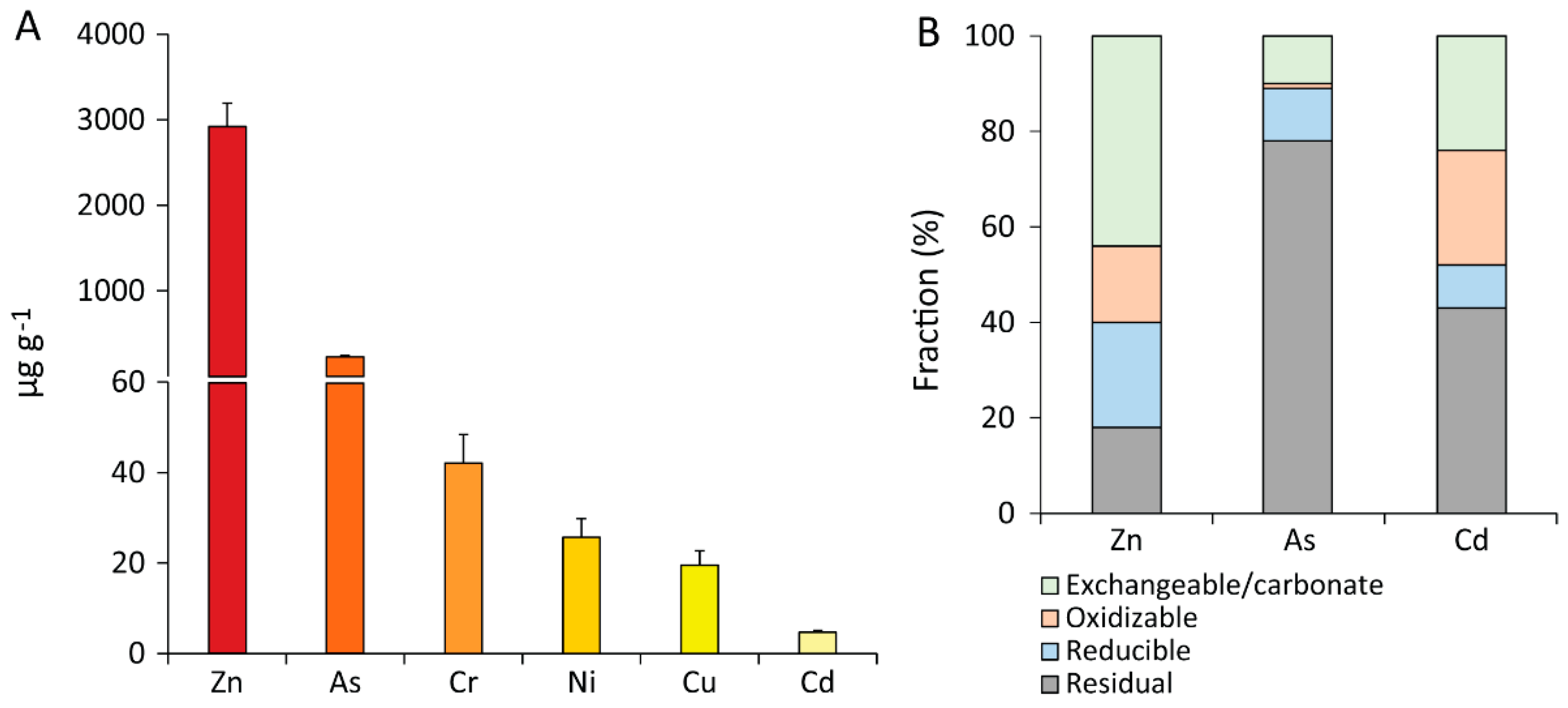
2.3. Determination of HM Concentrations and HM Bioremediation Yields
2.4. Statistical Analyses
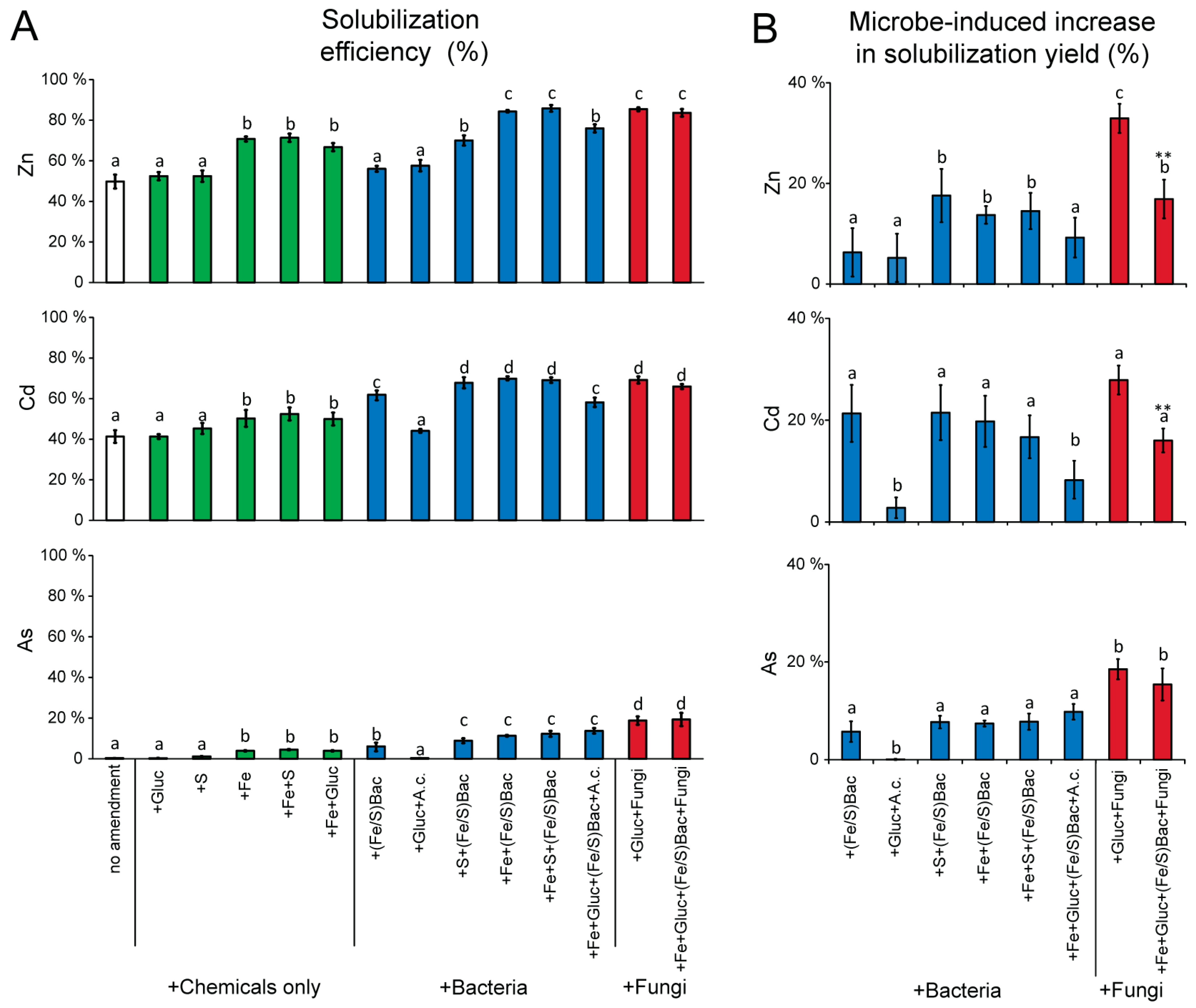
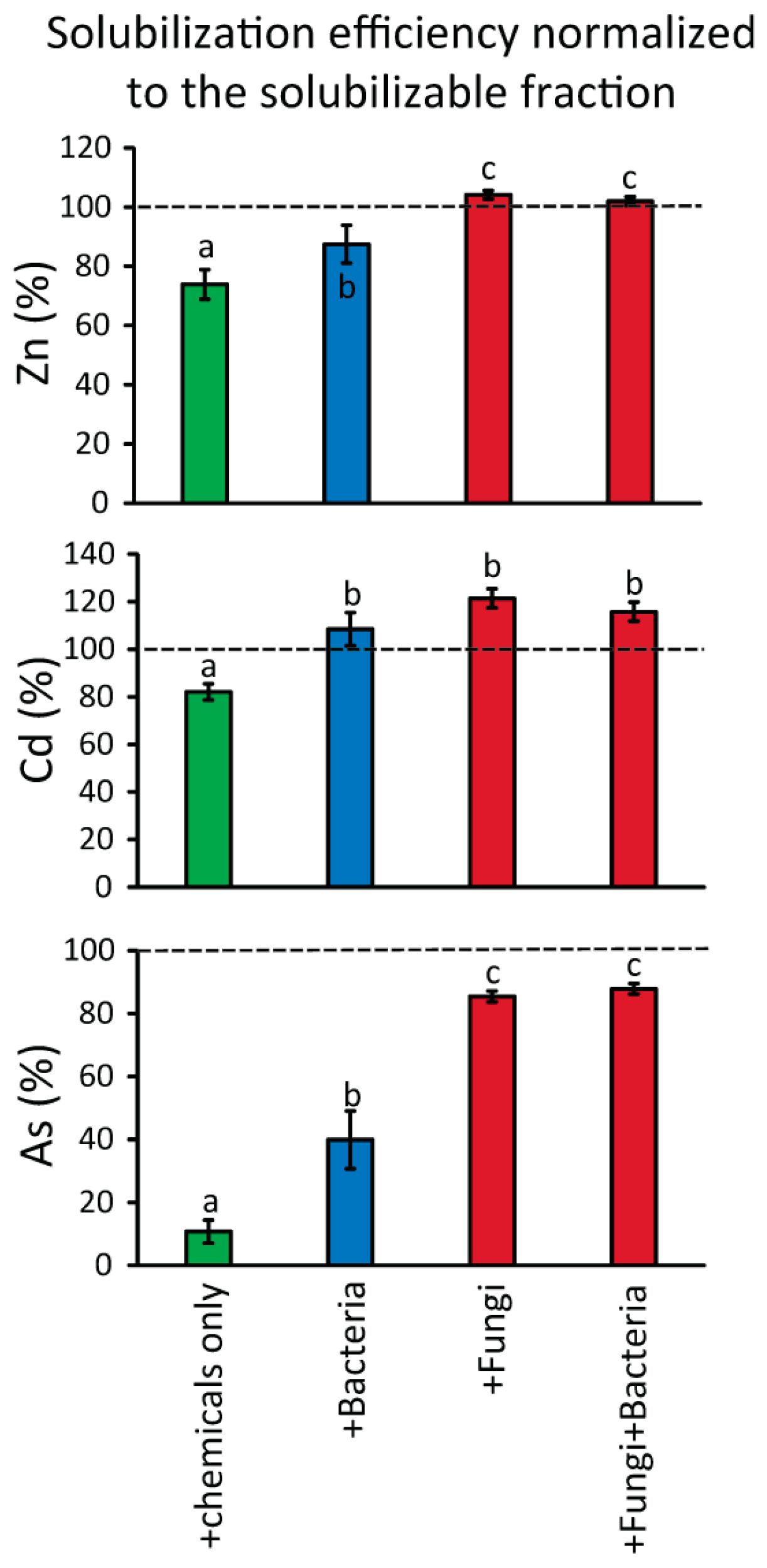
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fonti, V.; Dell’Anno, A.; Beolchini, F. Influence of biogeochemical interactions on metal bioleaching performance in contaminated marine sediment. Water Res. 2013, 47, 5139–5152. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Wang, C. Natural and human factors affect the distribution of soil heavy metal pollution: A review. Water Air Soil Pollut. 2020, 231, 350. [Google Scholar] [CrossRef]
- Ayangbenro, A.S.; Babalola, O.O. A new strategy for heavy metal polluted environments: A review of microbial biosorbents. Int. J. Environ. Res. Public Health 2017, 14, 94. [Google Scholar] [CrossRef] [PubMed]
- Kalaimurugan, D.; Balamuralikrishnan, B.; Durairaj, K.; Vasudhevan, P.; Shivakumar, M.S.; Kaul, T.; Chang, S.W.; Ravindran, B.; Venkatesan, S. Isolation and characterization of heavy-metal-resistant bacteria and their applications in environmental bioremediation. Int. J. Environ. Sci. Technol. 2020, 17, 1455–1462. [Google Scholar] [CrossRef]
- Priyadarshanee, M.; Das, S. Biosorption and removal of toxic heavy metals by metal tolerating bacteria for bioremediation of metal contamination: A comprehensive review. J. Environ. Chem. Eng. 2021, 9, 104686. [Google Scholar] [CrossRef]
- Ahemad, M.; Kibret, M. Recent trends in microbial biosorption of heavy metals: A review. Biochem. Mol. Biol. 2013, 1, 19–26. [Google Scholar] [CrossRef]
- Abdu, N.; Abdullahi, A.A.; Abdulkadir, A. Heavy metals and soil microbes. Environ. Chem. Lett. 2017, 15, 65–84. [Google Scholar] [CrossRef]
- U.S. Environmental Protection Agency. 2015 Update to the 1998 U.S. EPA Supplemental Environmental Projects Policy. Available online: https://www.epa.gov/enforcement/2015-update-1998-us-epa-supplemental-environmental-projects-policy (accessed on 6 May 2022).
- Bădescu, I.S.; Bulgariu, D.; Ahmad, I.; Bulgariu, L. Valorisation possibilities of exhausted biosorbents loaded with metal ions—A review. J. Environ. Manag. 2018, 224, 288–297. [Google Scholar] [CrossRef]
- Beolchini, F.; Dell’Anno, A.; De Propris, L.; Ubaldini, S.; Cerrone, F.; Danovaro, R. Auto- and heterotrophic acidophilic bacteria enhance the bioremediation efficiency of sediments contaminated by heavy metals. Chemosphere 2009, 74, 1321–1326. [Google Scholar] [CrossRef]
- Pasciucco, F.; Pecorini, I.; Di Gregorio, S.; Pilato, F.; Iannelli, R. Recovery Strategies of Contaminated Marine Sediments: A Life Cycle Assessment. Sustainability 2021, 13, 8520. [Google Scholar] [CrossRef]
- Renella, G. Recycling and Reuse of Sediments in Agriculture: Where Is the Problem? Sustainability 2021, 13, 1648. [Google Scholar] [CrossRef]
- Wang, Q.; Li, J.S.; Xue, Q.; Poon, C.S. Immobilization and recycling of contaminated marine sediments in cement-based materials incorporating iron-biochar composites. J. Hazard. Mater. 2022, 435, 128971. [Google Scholar] [CrossRef] [PubMed]
- Roeters, P.B. Large scale treatment of contaminated sediments in the Netherlands, the feasibility study. Water Sci. Technol. 1998, 37, 291–298. [Google Scholar] [CrossRef]
- Rienks, J. Comparison of results for chemical and thermal treatment of contaminated dredged sediments. Water Sci. Technol. 1998, 37, 355–362. [Google Scholar] [CrossRef]
- Mulligan, C.N.; Yong, R.N.; Gibbs, B.F. An evaluation of technologies for the heavy metal remediation of dredged sediments. J. Hazard. Mater. 2001, 85, 145–163. [Google Scholar] [CrossRef]
- Jones, K.W.; Feng, H.; Stern, E.A.; Lodge, J.; Clesceri, N.L. Dredged material decontamination demonstration for the port of New York/New Jersey. J. Hazard. Mater. 2001, 85, 127–143. [Google Scholar] [CrossRef]
- Meegoda, J.N.; Perera, R. Ultrasound to decontaminate heavy metals in dredged sediments. J. Hazard. Mater. 2001, 85, 73–89. [Google Scholar] [CrossRef]
- Akcil, A.; Erust, C.; Ozdemiroglu, S.; Fonti, V.; Beolchini, F. A review of approaches and techniques used in aquatic contaminated sediments: Metal removal and stabilization by chemical and biotechnological processes. J. Clean. Prod. 2015, 86, 24–36. [Google Scholar] [CrossRef]
- Peng, W.; Li, X.; Xiao, S.; Fan, W. Review of remediation technologies for sediments contaminated by heavy metals. J. Soils Sediments 2018, 18, 1701–1719. [Google Scholar] [CrossRef]
- Haynes, R.J.; Zhou, Y.F. Retention of heavy metals by dredged sediments and their management following land application. Adv. Agron. 2021, 171, 191–254. [Google Scholar]
- U.S. Environmental Protection Agency. Innovative Treatment Technologies: Overview and Guide to Information Sources EPA/540/9-91/002. 1991. Available online: https://nepis.epa.gov/Exe/ZyNET.exe/2000KFUQ.txt?ZyActionD=ZyDocument&Client=EPA&Index=1991%20Thru%201994&Docs=&Query=&Time=&EndTime=&SearchMethod=1&TocRestrict=n&Toc=&TocEntry=&QField=&QFieldYear=&QFieldMonth=&QFieldDay=&UseQField=&IntQFieldOp=0&ExtQFieldOp=0&XmlQuery=&File=D%3A%5CZYFILES%5CINDEX%20DATA%5C91THRU94%5CTXT%5C00000015%5C2000KFUQ.txt&User=ANONYMOUS&Password=anonymous&SortMethod=h%7C-&MaximumDocuments=1&FuzzyDegree=0&ImageQuality=r75g8/r75g8/x150y150g16/i425&Display=hpfr&DefSeekPage=x&SearchBack=ZyActionL&Back=ZyActionS&BackDesc=Results%20page&MaximumPages=1&ZyEntry=250&slide (accessed on 6 May 2022).
- Zoubeir, L.; Adeline, S.; Laurent, C.S.; Yoann, C.; Truc, H.T.; Benoit, L.G.; Federico, A. The use of the Novosol process for the treatment of polluted marine sediment. J. Hazard. Mater. 2007, 148, 606–612. [Google Scholar] [CrossRef] [PubMed]
- Pazos, M.; Plaza, A.; Martín, M.; Lobo, M.C. The impact of electrokinetic treatment on a loamy-sand soil property. Chem. Eng. J. 2012, 183, 231–237. [Google Scholar] [CrossRef]
- Rosas, J.M.; Vicente, F.; Santos, A.; Romero, A. Soil remediation using soil washing followed by Fenton oxidation. Chem. Eng. J. 2013, 220, 125–132. [Google Scholar] [CrossRef]
- Tsang, D.C.; Lo, M.C.; Surampalli, R.Y. Design, implementation, and economic/societal considerations of chelant-enhanced soil washing. In Chelating Agents for Land Decontamination Technologies; American Society of Civil Engineers: Reston, VA, USA, 2012; p. 1. [Google Scholar]
- Fashola, M.O.; Ngole-Jeme, V.M.; Babalola, O.O. Heavy Metal Immobilization Potential of Indigenous Bacteria Isolated from Gold Mine Tailings. Int. J. Environ. Res. 2020, 14, 71–86. [Google Scholar] [CrossRef]
- Dell’Anno, F.; Rastelli, E.; Tangherlini, M.; Corinaldesi, C.; Sansone, C.; Brunet, C.; Balzano, S.; Ianora, A.; Musco, L.; Montereali, M.R.; et al. Highly contaminated marine sediments can host rare bacterial taxa potentially useful for bioremediation. Front. Microbiol. 2021, 12, 326. [Google Scholar] [CrossRef] [PubMed]
- Dell’Anno, F.; Rastelli, E.; Sansone, C.; Brunet, C.; Ianora, A.; Dell’Anno, A. Bacteria, fungi and microalgae for the bioremediation of marine sediments contaminated by petroleum hydrocarbons in the omics era. Microorganisms 2021, 9, 1695. [Google Scholar] [CrossRef]
- Cecchi, G.; Vagge, G.; Cutroneo, L.; Greco, G.; Di Piazza, S.; Faga, M.; Zotti, M.; Capello, M. Fungi as potential tool for polluted port sediment remediation. Environ. Sci. Pollut. Res. 2019, 26, 35602–35609. [Google Scholar] [CrossRef]
- Magan, N.; Gouma, S.; Fragoeiro, S.; Shuaib, M.E.; Bastos, A.C. Bacterial and fungal bioremediation strategies. In Microbial Biodegradation and Bioremediation; Elsevier: Amsterdam, The Netherlands, 2022; pp. 193–212. [Google Scholar]
- Peng, J.F.; Song, Y.H.; Yuan, P.; Cui, X.Y.; Qiu, G.L. The remediation of heavy metals contaminated sediment. J. Hazard. Mater. 2009, 161, 633–640. [Google Scholar] [CrossRef]
- Gan, M.; Song, Z.; Zhu, J.; Liu, X. Efficient bioleaching of heavy metals from contaminated sediment in batch method coupled with the assistance of heterotrophic microorganisms. Environ. Earth Sci. 2016, 75, 1–10. [Google Scholar] [CrossRef]
- Hassan, A.; Periathamby, A.; Ahmed, A.; Innocent, O.; Hamid, F.S. Effective bioremediation of heavy metal–contaminated landfill soil through bioaugmentation using consortia of fungi. J. Soils Sediments 2020, 20, 66–80. [Google Scholar] [CrossRef]
- Dell’Anno, F.; Brunet, C.; van Zyl, L.J.; Trindade, M.; Golyshin, P.N.; Dell’Anno, A.; Ianora, A.; Sansone, C. Degradation of hydrocarbons and heavy metal reduction by marine bacteria in highly contaminated sediments. Microorganisms 2020, 8, 1402. [Google Scholar] [CrossRef] [PubMed]
- Tamegiai, H.; Kai, M.; Fukumori, Y.; Yamanaka, T. Two membrane-bound c-type cytochromes of Thiobacillus ferrooxidans: Purification and properties. FEMS Microb. Lett. 1994, 119, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Ohmura, N.; Sasaki, K.; Matsumoto, N.; Saiki, H. Anaerobic Respiration Using Fe3+, S0, and H2 in the Chemolithoautotrophic Bacterium Acidithiobacillus ferrooxidans. J. Bacteriol. 2002, 184, 2081–2087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Zhang, X.; Ni, Y.; Yang, X.; Li, H. Bioleaching of arsenic from medicinal realgar by pure and mixed cultures. Process. Biochem. 2007, 42, 1265–1271. [Google Scholar] [CrossRef]
- Fonti, V.; Dell’Anno, A.; Beolchini, F. Does bioleaching represent a biotechnological strategy for remediation of contaminated sediments? Sci. Total Environ. 2016, 563, 302–319. [Google Scholar] [CrossRef]
- Sher, S.; Rehman, A. Use of heavy metals resistant bacteria—A strategy for arsenic bioremediation. Appl. Microbiol. Biotechnol. 2019, 103, 6007–6021. [Google Scholar] [CrossRef]
- Sun, W.; Cheng, K.; Sun, K.Y.; Ma, X. Microbially mediated remediation of contaminated sediments by heavy metals: A critical review. Curr. Pollut. Rep. 2021, 7, 201–212. [Google Scholar] [CrossRef]
- Nguyen, T.H.; Won, S.; Ha, M.G.; Nguyen, D.D.; Kang, H.Y. Bioleaching for environmental remediation of toxic metals and metalloids: A review on soils, sediments, and mine tailings. Chemosphere 2021, 282, 131108. [Google Scholar] [CrossRef]
- Inoue, T.; Kamimura, K.; Suogi, T. Isolation and some properties of a mesophilic and mixotrophic iron-oxidizing bacterium, OKM-9. Biosci. Biotechnol. Biochem. 2000, 64, 2059–2067. [Google Scholar] [CrossRef]
- Spring, S.; Kämpfer, P.; Schleifer, K.H. Limnobacter thiooxidans gen. nov., sp. nov., a novel thiosulfate-oxidizing bacterium isolated from freshwater lake sediment. Int. J. Syst. Evol. Microbiol. 2001, 51, 1463–1470. [Google Scholar] [CrossRef]
- Chen, S.; Lin, J. Bioleaching of heavy metals from contaminated sediment by indigenous sulphur-oxiding bacteria in air lift bioreactor: Effects of sulphur concentration. Water Res. 2004, 38, 3205–3214. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Lin, J. Bioleaching of heavy metals from sediment: Significance of pH. Chemosphere 2001, 44, 1093–1102. [Google Scholar] [CrossRef]
- Finneran, K.T.; Johnsen, C.V.; Lovley, D.R. Rhodoferax ferrireducens sp. nov., a psychrotolerant, facultatively anaerobic bacteriumthat oxidizes acetate with the reduction of Fe(III). Int. J. Syst. Evol. Microbiol. 2003, 53, 669–673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumbhar, P.; Savla, N.; Banerjee, S.; Mathuriya, A.S.; Sarkar, A.; Khilari, S.; Jadhav, D.A.; Pandit, S. Microbial electrochemical heavy metal removal: Fundamental to the recent development. In Wastewater treatment; Elsevier: Amsterdam, The Netherlands, 2021; pp. 521–542. [Google Scholar]
- Kusël, K.; Dorsch, T.; Acker, G.; Stackebrandt, E. Microbial Reduction of Fe(III) in Acidic Sediments: Isolation of Acidiphilium cryptum JF-5 Capable of Coupling the Reduction of Fe(III) to the Oxidation of Glucose. Appl. Environ. Microbiol. 1999, 65, 3633–3640. [Google Scholar] [CrossRef] [Green Version]
- Rawlings, D.E. Relevance of cell physiology and genetic adaptability of biomining microorganisms to industrial processes. In Biomining; Springer: Berlin/Heidelberg, Germany, 2007; pp. 177–198. [Google Scholar]
- Nancucheo, I.; Johnson, D.B. Characteristics of an Iron-Reducing, Moderately Acidophilic Actinobacterium Isolated from Pyritic Mine Waste, and Its Potential Role in Mitigating Mineral Dissolution in Mineral Tailings Deposits. Microorganisms 2020, 8, 990. [Google Scholar] [CrossRef]
- Fournier, D.; Lemieux, R.; Couillard, D. Essential interactions between Thiobacillus ferrooxidans and heterotrophic microorganisms during a wastewater sludge bioleaching process. Environ. Pollut. 1998, 101, 303–309. [Google Scholar] [CrossRef]
- González-Toril, E.; Llobet-Brossa, E.; Casamayor, E.O.; Amann, R.; Amils, R. Microbial Ecology of an Extreme Acidic Environment, the Tinto River. Appl. Environ. Microbiol. 2003, 69, 4853–4865. [Google Scholar] [CrossRef] [Green Version]
- Xiao, C.Q.; Chi, R.A.; Fang, Y.J. Effects of Acidiphilium cryptum on biosolubilization of rock phosphate in the presence of Acidithiobacillus ferrooxidans. Trans. Nonferr. Met. Soc. China 2013, 23, 2153–2159. [Google Scholar] [CrossRef]
- Zhu, J.Y.; Zhang, J.X.; Qian, L.I.; Tao, H.A.N.; Hu, Y.H.; Liu, X.D.; Qin, W.Q.; Chai, L.Y.; Qiu, G.Z. Bioleaching of heavy metals from contaminated alkaline sediment by auto-and heterotrophic bacteria in stirred tank reactor. Trans. Nonferr. Met. Soc. China 2014, 24, 2969–2975. [Google Scholar] [CrossRef]
- Xu, M.; Liu, Y.; Deng, Y.; Zhang, S.; Hao, X.; Zhu, P.; Zhou, J.; Yin, H.; Liang, Y.; Liu, H.; et al. Bioremediation of cadmium-contaminated paddy soil using an autotrophic and heterotrophic mixture. RSC Adv. 2020, 10, 26090–26101. [Google Scholar] [CrossRef]
- Dursun, A.; Uslu, G.; Cuci, Y.; Aksu, Z. Bioaccumulation of copper(II), lead(II) and chromium(VI) by growing Aspergillus niger. Process. Biochem. 2003, 38, 1647–1651. [Google Scholar] [CrossRef]
- Vala, A.K. Tolerance and removal of arsenic by a facultative marine fungus Aspergillus candidus. Bioresour. Technol. 2010, 101, 2565–2567. [Google Scholar] [CrossRef] [PubMed]
- Vala, A.K.; Sachaniya, B.; Dave, B.P. Marine-Derived Fungi: Promising Candidates for Enhanced Bioremediation. In Nanomaterial Biointeractions at the Cellular, Organismal and System Levels; Springer Science and Business Media LLC: Berlin/Heidelberg, Germany, 2018; pp. 281–300. [Google Scholar]
- Adams, P.; Lynch, J.M.; De Leij, F.A.A.M. Desorption of zinc by extracellularly produced metabolites of Trichoderma harzianum, Trichoderma reesei and Coriolus versicolor. J. Appl. Microbiol. 2007, 103, 2240–2247. [Google Scholar] [CrossRef]
- Kacprzak, M.J.; Rosikon, K.; Fijalkowski, K.; Grobelak, A. The effect of Trichoderma on heavy metal mobility and uptake by Miscanthus giganteus, Salix sp., Phalaris arundinacea, and Panicum virgatum. Appl. Environ. Soil Sci. 2014, 2014, 506142. [Google Scholar] [CrossRef] [Green Version]
- Dusengemungu, L.; Kasali, G.; Gwanama, C.; Mubemba, B. Overview of fungal bioleaching of metals. Environ. Adv. 2021, 5, 100083. [Google Scholar] [CrossRef]
- Sabra, N.; Dubourguier, H.C.; Duval, M.N.; Hamieh, T. Study of canal sediments contaminated with heavy metals: Fungal versus bacterial bioleaching techniques. Environ. Technol. 2011, 32, 1307–1324. [Google Scholar] [CrossRef]
- Zeng, X.; Wei, S.; Sun, L.; Jacques, D.A.; Tang, J.; Lian, M.; Ji, Z.; Wang, J.; Zhu, J.; Xu, Z. Bioleaching of heavy metals from contaminated sediments by the Aspergillus niger strain SY1. J. Soils Sediments 2015, 15, 1029–1038. [Google Scholar] [CrossRef]
- Zeng, X.; Twardowska, I.; Wei, S.; Sun, L.; Wang, J.; Zhu, J.; Cai, J. Removal of trace metals and improvement of dredged sediment dewaterability by bioleaching combined with Fenton-like reaction. J. Hazard. Mater. 2015, 288, 51–59. [Google Scholar] [CrossRef]
- Mestre, N.C.; Rocha, T.L.; Canals, M.; Cardoso, C.; Danovaro, R.; Dell’Anno, A.; Gambi, C.; Regoli, F.; Sanchez-Vidal, A.; Bebianno, M.J. Environmental hazard assessment of a marine mine tailings deposit site and potential implications for deep-sea mining. Environ. Pollut. 2017, 228, 169–178. [Google Scholar] [CrossRef]
- Marchand, E.A.; Silverstein, J. The role of enhanced heterotrophic bacterial growth on iron oxidation by Acidithiobacillus ferrooxidans. Geomicrobiol. J. 2003, 20, 231–244. [Google Scholar] [CrossRef]
- Guenet, B.; Danger, M.; Abbadie, L.; Lacroix, G. Priming effect: Bridging the gap between terrestrial and aquatic ecology. Ecology 2010, 91, 2850–2861. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Liu, P.; Shu, W.; Li, C.; Li, H.; Liu, S.; Xia, J.; Noorman, H. Dynamic response of Aspergillus niger to single pulses of glucose with high and low concentrations. BIOB 2019, 6, 16. [Google Scholar] [CrossRef]
- Dell’Anno, A.; Beolchini, F.; Corinaldesi, C.; Amato, A.; Becci, A.; Rastelli, E.; Hekeu, M.; Regoli, F.; Astarita, E.; Greco, S.; et al. Assessing the efficiency and eco-sustainability of bioremediation strategies for the reclamation of highly contaminated marine sediments. Mar. Environ. Res. 2020, 162, 105101. [Google Scholar] [CrossRef] [PubMed]
- Sabra, N.; Dubourguier, H.C.; Hamieh, T. Fungal bioleaching of heavy metals from sediments dredged from the Deûle Canal. France. Adv. Chem. Eng. Sci. 2012, 2, 16702. [Google Scholar] [CrossRef] [Green Version]
- Karlfeldt Fedje, K.; Ekberg, C.; Skarnemark, G.; Steenari, B.M. Removal of hazardous metals from MSW fly ash—An evaluation of ash bioleaching methods. J. Hazard. Mater. 2010, 173, 310–317. [Google Scholar] [CrossRef]
- El-Kassas, H.Y.; El-Taher, E.M. Optimization of batch process parameters by response surface methodology for mycoremediation of chrome-VI by a chromium resistant strain of marine Trichoderma viride. Am.-Eurasian J. Agric. Environ. Sci. 2009, 5, 676–681. [Google Scholar]
- Kartal, S.N.; Katsumata, N.; Imamura, Y. Removal of copper, chromium, and arsenic from CCA-treated wood by organic acids released by mold and staining fungi. For. Prod. J. 2006, 56, 33–37. [Google Scholar]
- Quevauviller, P.; Lachica, M.; Barahona, E.; Gomez, A.; Rauret, G.; Ure, A.; Muntau, H. Certified reference material for the quality control of EDTA-and DTPA-extractable trace metal contents in calcareous soil (CRM 600). Fresenius’ J. Anal. Chem. 1998, 360, 505–511. [Google Scholar] [CrossRef]
- Anderson, D.R. Model Based Inference in the Life Sciences: A Primer on Evidence; Springer: New York, NY, USA, 2008; Volume 31. [Google Scholar]
- Buchman, M.F. NOAA Screening Quick Reference Tables 1999. Available online: https://repository.library.noaa.gov/view/noaa/8310 (accessed on 6 May 2022).
- Italian DM 173/2016. Ministero dell’Ambiente e Della Tutela del Territorio e del Mare, Supplemento Ordinario alla Gazzetta Ufficiale, n. 208 del 6 Settembre 2016-Serie Generale. Regolamento Recante Modalità e Criteri Tecnici Per l′Autorizzazione All’immersione in Mare dei Materiali di Escavo di Fondali Marini. Available online: https://www.gazzettaufficiale.it/eli/id/2016/09/06/16G00184/sg (accessed on 6 May 2022).
- Long, E.R.; MacDonald, D.D.; Smith, S.L.; Calder, F.D. Incidence of adverse biological effects within ranges of chemical concentrations in marine and estuarine sediments. Environ. Manag. 1995, 19, 81–97. [Google Scholar] [CrossRef]
- Tangherlini, M.; Corinaldesi, C.; Rastelli, E.; Musco, L.; Armiento, G.; Danovaro, R.; Dell’Anno, A. Chemical contamination can promote turnover diversity of benthic prokaryotic assemblages: The case study of the Bagnoli-Coroglio bay (southern Tyrrhenian Sea). Mar. Environ. Res. 2020, 160, 105040. [Google Scholar] [CrossRef]
- Schippers, A.; Jørgensen, B.B. Biogeochemistry of pyrite and iron sulfide oxidation in marine sediments. Geochim. Cosmochim. Acta 2002, 66, 85–92. [Google Scholar] [CrossRef]
- Yuan, L.I.U. Feasibility study of bioleaching for low-grade ore in Tongkuaangyu Copper Mine. Min. Metal. Explor. 2004, 13, 26–29. [Google Scholar]
- Jafari, M.; Shafaei, S.Z.; Abdollahi, H.; Gharabaghi, M.; Chehreh Chelgani, S. Effect of flotation reagents on the activity of L. ferrooxidans. Miner. Process. Extr. Metall. Rev. 2018, 39, 34–43. [Google Scholar] [CrossRef]
- Ma, Y.; Lin, C. Microbial Oxidation of Fe2+ and Pyrite Exposed to Flux of Micromolar H2O2 in Acidic Media. Sci. Rep. 2013, 3, 1979. [Google Scholar] [CrossRef] [Green Version]
- Ko, M.S.; Park, H.S.; Kim, K.W.; Lee, J.U. The role of Acidithiobacillus ferrooxidans and Acidithiobacillus thiooxidans in arsenic bioleaching from soil. Environ. Geochem. Health 2013, 35, 727–733. [Google Scholar] [CrossRef]
- Huber, F.; Blasenbauer, D.; Mallow, O.; Lederer, J.; Winter, F.; Fellner, J. Thermal co-treatment of combustible hazardous waste and waste incineration fly ash in a rotary kiln. J. Waste Manag. 2016, 58, 181–190. [Google Scholar] [CrossRef]
- Zhou, S.G.; Zhou, L.X.; Fang, D. Enhancing metal removal by coaddition of Fe2+ and S0 as substrates of Acidithiobacillus ferrooxidans for sewage sludge bioleaching. J. Hazard. Toxic Radioact. Waste 2008, 12, 159–164. [Google Scholar] [CrossRef]
- Hao, X.; Zhu, P.; Zhang, H.; Liang, Y.; Yin, H.; Liu, X.; Bai, L.; Liu, H.; Jiang, H. Mixotrophic acidophiles increase cadmium soluble fraction and phytoextraction efficiency from cadmium contaminated soils. Sci. Total Environ. 2019, 655, 347–355. [Google Scholar] [CrossRef]
- Chang, C.Y.; Chen, S.Y.; Klipkhayai, P.; Chiemchaisri, C. Bioleaching of heavy metals from harbor sediment using sulfur-oxidizing microflora acclimated from native sediment and exogenous soil. Environ. Sci. Pollut. Res. 2019, 26, 6818–6828. [Google Scholar] [CrossRef]
- Cecchi, G.; Cutroneo, L.; Di Piazza, S.; Besio, G.; Capello, M.; Zotti, M. Port Sediments: Problem or Resource? A Review Concerning the Treatment and Decontamination of Port Sediments by Fungi and Bacteria. Microorganisms 2021, 9, 1279. [Google Scholar] [CrossRef]
- Seh-Bardan, B.J.; Othman, R.; Wahid, S.A.; Husin, A.; Sadegh-Zadeh, F. Bioleaching of heavy metals from mine tailings by Aspergillus fumigatus. Bioremediat. J. 2012, 16, 57–65. [Google Scholar] [CrossRef]
- Thompson, I.P.; Van Der Gast, C.J.; Ciric, L.; Singer, A.C. Bioaugmentation for bioremediation: The challenge of strain selection. Environ. Microbiol. 2005, 7, 909–915. [Google Scholar] [CrossRef] [PubMed]
- Urík, M.; Farkas, B.; Miglierini, M.B.; Bujdoš, M.; Mitróová, Z.; Kim, H.; Matúš, P. Mobilisation of hazardous elements from arsenic-rich mine drainage ochres by three Aspergillus species. J. Hazard. Mater. 2021, 409, 124938. [Google Scholar] [CrossRef] [PubMed]
- Govarthanan, M.; Mythili, R.; Selvankumar, T.; Kamala-Kannan, S.; Kim, H. Myco-phytoremediation of arsenic-and lead-contaminated soils by Helianthus annuus and wood rot fungi, Trichoderma sp. isolated from decayed wood. Ecotoxicol. Environ. Saf. 2018, 151, 279–284. [Google Scholar] [CrossRef]
- Gleyzes, C.; Tellier, S.; Astruc, M. Fractionation studies of trace elements in contaminated soils and sediments: A review of sequential extraction procedures. Trends Anal. Chem. 2002, 21, 451–467. [Google Scholar] [CrossRef]
- Fang, D.; Zhou, L.X. Effect of sludge dissolved organic matter on oxidation of ferrous iron and sulfur by Acidithiobacillus ferrooxidans and Acidithiobacillus thiooxidans. Water Air Soil Poll. 2006, 171, 81–94. [Google Scholar] [CrossRef]
- Tsiropoulos, I.; Cok, B.; Patel, M.K. Energy and greenhouse gas assessment of European glucose production from corn–a multiple allocation approach for a key ingredient of the bio-based economy. J. Clean. Prod. 2013, 43, 182–190. [Google Scholar] [CrossRef]
- Dhillon, G.S.; Brar, S.K.; Verma, M.; Tyagi, R.D. Utilization of different agro-industrial wastes for sustainable bioproduction of citric acid by Aspergillus niger. Biochem. Eng. J. 2011, 54, 83–92. [Google Scholar] [CrossRef]
- Peng, T.J.; Shi, L.J.; Yu, R.L.; Gu, G.H.; Dan, Z.; Miao, C.N.; Zeng, W.M. Effects of processing pH stimulation on cooperative bioleaching of chalcopyrite concentrate by free and attached cells. Trans. Non-Ferr. Met. Soc. China 2016, 26, 2220–2229. [Google Scholar] [CrossRef]
- Sajjad, W.; Zheng, G.; Zhang, G.; Ma, X.; Xu, W.; Khan, S. Bioleaching of copper-and zinc-bearing ore using consortia of indigenous iron-oxidizing bacteria. Extremophiles 2018, 22, 851–863. [Google Scholar] [CrossRef]
- Ghavidel, A.; Naji Rad, S.; Alikhani, H.A.; Sharari, M.; Ghanbari, A. Bioleaching of heavy metals from sewage sludge, direct action of Acidithiobacillus ferrooxidans or only the impact of pH? J. Mater. Cycles Waste Manag. 2018, 20, 1179–1187. [Google Scholar] [CrossRef]
- Pathak, A.; Dastidar, M.G.; Sreekrishnan, T.R. Bioleaching of heavy metals from sewage sludge: A review. J. Environ. Manag. 2009, 90, 2343–2353. [Google Scholar] [CrossRef] [PubMed]
- Ilyas, S.; Ru’an, C.H.I.; Lee, J.C.; Bhatti, H.N. One step bioleaching of sulphide ore with low concentration of arsenic by Aspergillus niger and Taguchi orthogonal array optimization. Chin. J. Chem. Eng. 2012, 20, 923–929. [Google Scholar] [CrossRef]
- Liang, X.; Gadd, G.M. Metal and metalloid biorecovery using fungi. Microb. Biotechnol. 2017, 10, 1199–1205. [Google Scholar] [CrossRef] [Green Version]
- Varrella, S.; Barone, G.; Tangherlini, M.; Rastelli, E.; Dell’Anno, A.; Corinaldesi, C. Diversity, ecological role and biotechnological potential of antarctic marine fungi. J. Fungi 2021, 7, 391. [Google Scholar] [CrossRef]
- Barone, G.; Varrella, S.; Tangherlini, M.; Rastelli, E.; Dell’Anno, A.; Danovaro, R.; Corinaldesi, C. Marine fungi: Biotechnological perspectives from deep-hypersaline anoxic basins. Diversity 2019, 11, 113. [Google Scholar] [CrossRef] [Green Version]
- Danovaro, R.; Manini, E.; Dell’Anno, A. Higher abundance of bacteria than of viruses in deep Mediterranean sediments. Appl. Environ. Microbiol. 2002, 68, 1468–1472. [Google Scholar] [CrossRef] [Green Version]
- Barone, G.; Rastelli, E.; Corinaldesi, C.; Tangherlini, M.; Danovaro, R.; Dell’Anno, A. Benthic deep-sea fungi in submarine canyons of the Mediterranean Sea. Prog. Oceanogr. 2018, 168, 57–64. [Google Scholar] [CrossRef]
- Kempf, V.A.J.; Trebesius, K.; Autenrieth, I.B. Fluorescent in situ hybridization allows rapid identification of microorganisms in blood cultures. J. Clin. Microbiol. 2000, 38, 830–838. [Google Scholar] [CrossRef] [Green Version]
- Menden-Deuer, S.; Lessard, E.J. Carbon to volume relationships for dinoflagellates, diatoms, and other protist plankton. Limnol. Oceanogr. 2000, 45, 569–579. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Sample ID | Fe (g/L) | S (g/L) | Glucose (g/L) |
|---|---|---|---|---|
| Incubation of original sediments (no additions) | No amendement | 0 | 0 | 0 |
| Addition of Chemicals Only | +Gluc | 0 | 0 | 0.1 |
| +S | 0 | 10 | 0 | |
| +Fe | 4.5 | 0 | 0 | |
| +Fe+S | 4.5 | 10 | 0 | |
| +Fe+Gluc | 4.5 | 0 | 0.1 | |
| Addition of Bacteria (autotrophic Fe/S oxidising bacteria -(Fe/S) Bac-, and/or heterotrophic bacteria -A.c., Acidiphilium cryptum-) | +(Fe/S) Bac | 0 | 0 | 0 |
| +Gluc+A.c. | 0 | 0 | 0.1 | |
| +S+(Fe/S) Bac | 0 | 10 | 0 | |
| +Fe+(Fe/S) Bac | 4.5 | 0 | 0 | |
| +Fe+S+(Fe/S) Bac | 4.5 | 10 | 0 | |
| +Fe+Glu+(Fe/S) Bac+A.c. | 4.5 | 0 | 0.1 | |
| Addition of Fungi | Gluc+Fungi | 0 | 0 | 0.1 |
| Addition of Fungi and Fe/S oxidising bacteria | +Fe+Glu+(Fe/S) Bac+Fungi | 4.5 | 0 | 0.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dell’Anno, F.; Rastelli, E.; Buschi, E.; Barone, G.; Beolchini, F.; Dell’Anno, A. Fungi Can Be More Effective than Bacteria for the Bioremediation of Marine Sediments Highly Contaminated with Heavy Metals. Microorganisms 2022, 10, 993. https://doi.org/10.3390/microorganisms10050993
Dell’Anno F, Rastelli E, Buschi E, Barone G, Beolchini F, Dell’Anno A. Fungi Can Be More Effective than Bacteria for the Bioremediation of Marine Sediments Highly Contaminated with Heavy Metals. Microorganisms. 2022; 10(5):993. https://doi.org/10.3390/microorganisms10050993
Chicago/Turabian StyleDell’Anno, Filippo, Eugenio Rastelli, Emanuela Buschi, Giulio Barone, Francesca Beolchini, and Antonio Dell’Anno. 2022. "Fungi Can Be More Effective than Bacteria for the Bioremediation of Marine Sediments Highly Contaminated with Heavy Metals" Microorganisms 10, no. 5: 993. https://doi.org/10.3390/microorganisms10050993