The Origins of Human Modernity

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
This does not mean that there was a single female from whom we are all descended, but rather that out of a population numbering perhaps several thousand, by chance, only one set of mitochondrial genes was passed on (This finding, perhaps the most surprising to us, is the least disputed by population geneticists and others familiar with genetic drift and other manifestations of the laws of probability.).[12]
2. The African Hoax


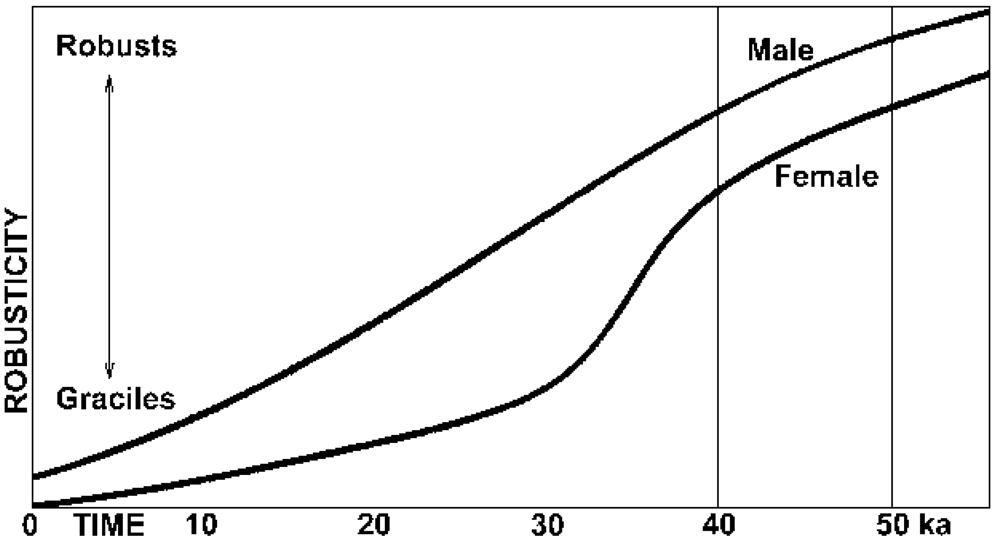
3. The Gracilization of Humans


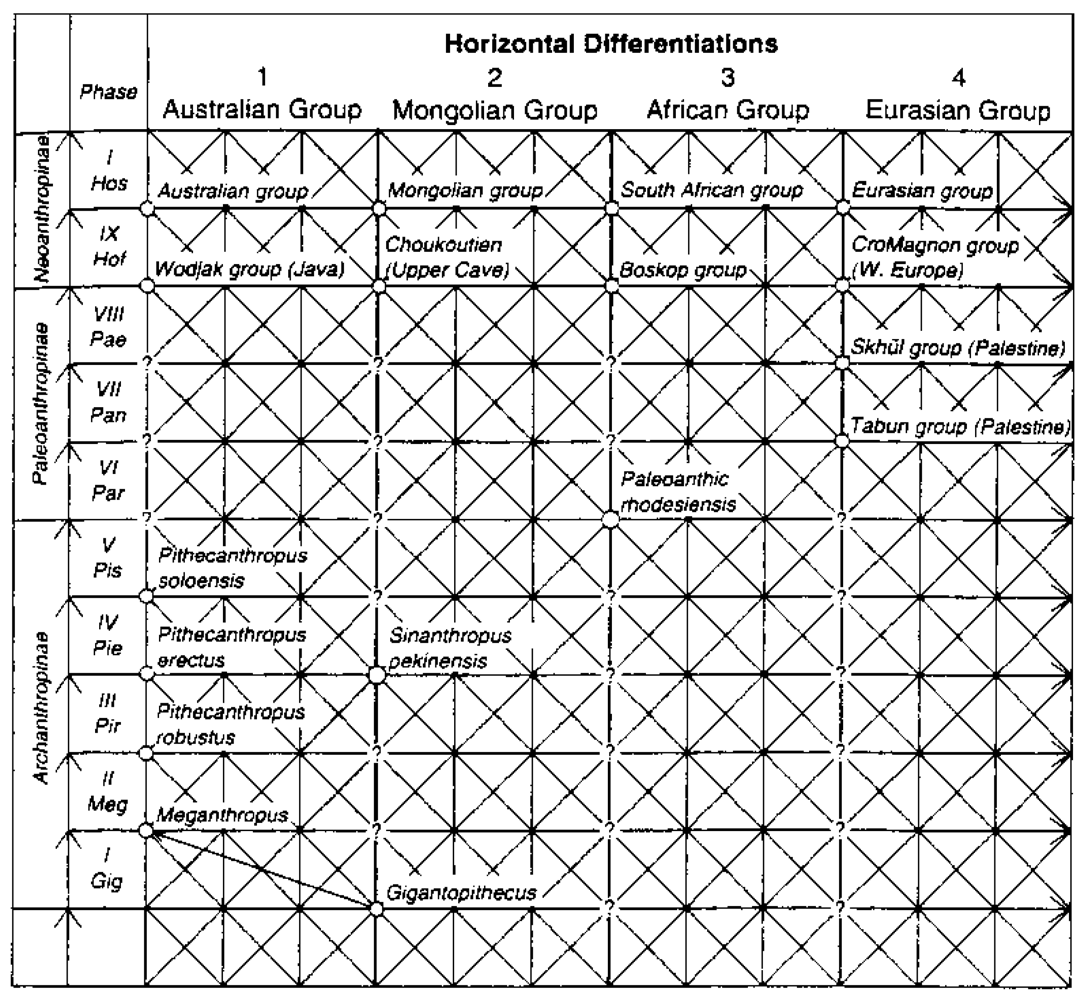
4. Alternative Models

5. The Fetalized Human

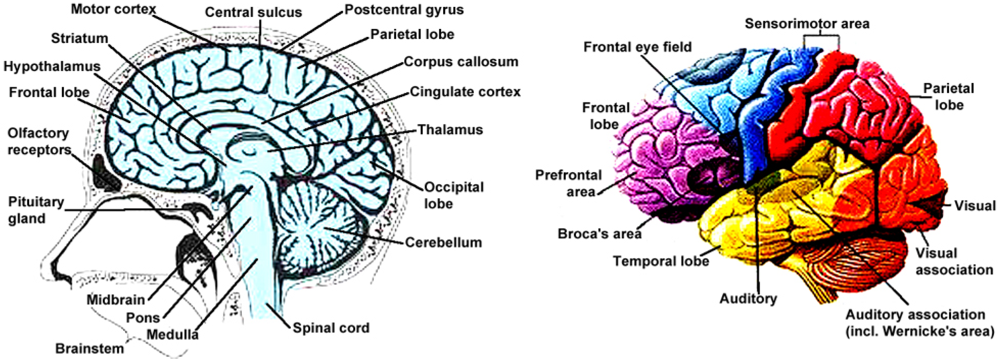
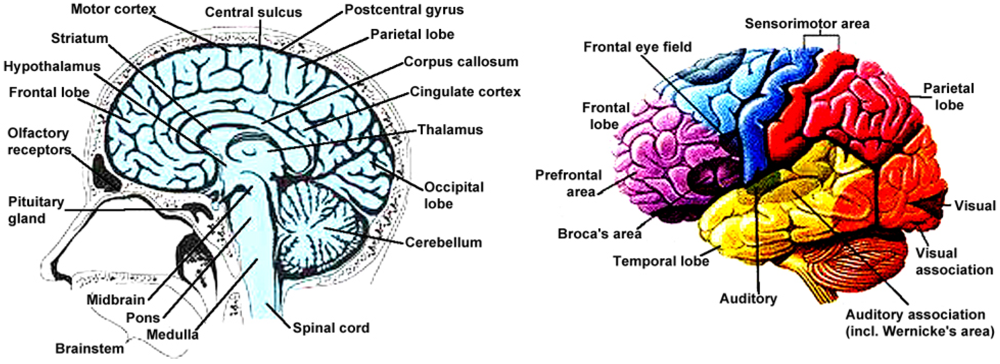
6. Susceptibility of the Human Brain to Illness
7. Conclusions

References
- P.V. Tobias. “The bearing of fossils and mitochondrial DNA on the evolution of modern humans, with a critique of the ‘mitochondrial Eve’ hypothesis.” S. Afr. Archaeol. Bull. 50 (1995): 155–167. [Google Scholar] [CrossRef]
- J.R. Searle. The Construction of Social Reality. London, UK: Allen Lane, 1995. [Google Scholar]
- D. Bickerton. Adam’s Tongue: How Humans Made Language, How Language Made Humans. New York, NY, USA: Hill and Wang, 2010. [Google Scholar]
- M. Schulz. “Die Regeln mache ich.” Der Spiegel 34 (2004): 128–131. [Google Scholar]
- L. Harding. “History of modern man unravels as German scholar is exposed as fraud.” The Guardian; London, UK: Guardian News and Media, 2005. Available online: http://www.guardian.co.uk/science/2005/feb/19/science.sciencenews (accessed on 25 August 2011).
- R.R.P. Von Zieten. “The Dating of Upper-Pleistocene Subsaharan Fossil Hominids and Their Place in Human Evolution: With Morphological and Archaeological Implications.” In Ph.D. Thesis. Los Angeles, CA, USA: University of California, 1973. [Google Scholar]
- R. Protsch. “The absolute dating of Upper Pleistocene sub-Saharan fossil hominids and their place in human evolution.” J. Hum. Evol. 4 (1975): 297–322. [Google Scholar]
- R. Protsch, and H. Glowatzki. “Das absolute Alter des paläolithischen Skeletts aus der Mittleren Klause bei Neuessing, Kreis Kelheim, Bayern.” Anthropol. Anz. 34 (1974): 140–144. [Google Scholar]
- R. Protsch, and A. Semmel. “Zur Chronologie des Kelsterbach-Hominiden.” Eiszeitalter Ggw. 28 (1978): 200–210. [Google Scholar]
- W. Henke, and R. Protsch. “Die Paderborner Calvaria—Ein diluvialer Homo sapiens.” Anthropol. Anz. 36 (1978): 85–108. [Google Scholar]
- R.L. Cann, M. Stoneking, and A.C. Wilson. “Mitochondrial DNA and human evolution.” Nature 325 (1987): 31–36. [Google Scholar]
- H. Curtis, and N.S. Barnes. Biology, 5th ed. New York, NY, USA: Worth Publishers Inc., 1989. [Google Scholar]
- T. Kuhn. The Structure of Scientific Revolutions. Chicago, IL, USA: University of Chicago Press, 1962. [Google Scholar]
- I. Davidson, and W. Noble. “The archaeology of perception: Traces of depiction and language.” Curr. Anthropol. 30 (1989): 125–155. [Google Scholar]
- I. Davidson, and W. Noble. “Tools, humans and evolution—The relevance of the Upper Palaeolithic.” In Tools, Language and Intelligence: Evolutionary Implications. Cascais, Portugal: Wenner-Gren Foundation, 1990, pp. 1–21. [Google Scholar]
- W. Noble, and I. Davidson. Human Evolution, Language and Mind: A Psychological and Archaeological Inquiry. Cambridge, UK: Cambridge University Press, 1996. [Google Scholar]
- F. D’Errico, and A. Nowell. “A new look at the Berekhat Ram figurine: Implications for the origins of symbolism.” Camb. Archaeol. J. 10 (2000): 123–167. [Google Scholar]
- G. Bräuer. “Präsapiens-Hypothese oder Afro-europäische Sapiens-Hypothese? ” Z. Morphol. Anthropol. 75 (1984): 1–25. [Google Scholar]
- C.B. Stringer, and P. Andrews. “Genetic and fossil evidence for the origin of modern humans.” Science 239 (1988): 1263–1268. [Google Scholar]
- V. Eswaran. “A diffusion wave out of Africa.” Curr. Anthropol. 43 (2002): 749–774. [Google Scholar] [CrossRef]
- F.H. Smith, I. Janković, and I. Karavanić. “The assimilation model, modern human origins in Europe, and the extinction of Neandertals.” Quat. Int. 137 (2005): 7–19. [Google Scholar] [CrossRef]
- F. Weidenreich. Apes, Giants, and Man. Chicago, IL, USA: University of Chicago Press, 1946. [Google Scholar]
- J.H. Relethford. “Absence of regional affinities of Neandertal DNA with living humans does not reject multiregional evolution.” Am. J. Phys. Anthropol. 115 (2002): 95–98. [Google Scholar] [CrossRef]
- J.H. Relethford, and L.B. Jorde. “Genetic evidence for larger African population size during recent human evolution.” J. Phys. Anthropol. 108 (1999): 251–260. [Google Scholar] [CrossRef]
- E.H. Bryant, S.A. McComas, and L.M. Combs. “The effect of an experimental bottleneck on quantitative genetic variation in the housefly.” Genetics 114 (1986): 1191–1211. [Google Scholar]
- L.R. Berger, S.E. Churchill, B. de Klerk, and R.L. Quinn. “Small-bodied humans from Palau, Micronesia.” PLoS One 3 (2008): e1780. [Google Scholar]
- J. Hawks, S.-H. Lee, K. Hunley, and M. Wolpoff. “Population bottlenecks and Pleistocene human evolution.” Mol. Biol. Evol. 17 (2000): 2–22. [Google Scholar] [CrossRef]
- R.G. Bednarik, and M. Kuckenburg. Nale Tasih: Eine Floßfahrt in Die Steinzeit. Stuttgart, Germany: Jan Thorbecke, 1999. [Google Scholar]
- M.F. Hammer. “A recent common ancestry for human Y chromosomes.” Nature 378 (1995): 376–378. [Google Scholar] [CrossRef]
- U. Gyllensten, D. Wharton, A. Josefsson, and A.C. Wilson. “Paternal inheritance of mitochondrial DNA in mice.” Nature 352 (1991): 255–257. [Google Scholar]
- P. Awadalla, A. Eyre-Walker, and J. Maynard Smith. “Linkage disequilibrium and recombination in hominid mitochondrial DNA.” Science 286 (1999): 2524–2525. [Google Scholar]
- A.A.M. Morris, and R.N. Lightowlers. “Can paternal mtDNA be inherited? ” Lancet 355 (2000): 1290–1291. [Google Scholar] [CrossRef]
- R.S. Williams. “Another surprise from the mitochondrial genome.” N. Engl. J. Med. 347 (2002): 609–611. [Google Scholar] [CrossRef]
- M. Schwartz, and J. Vissing. “Paternal inheritance of mitochondrial DNA.” N. Engl. J. Med. 347 (2002): 576–580. [Google Scholar]
- F. Rodriguez-Trelles, R. Tarrio, and F.J. Ayala. “Erratic overdispersion of three molecular clocks: GPDH, SOD, and XDH.” Proc. Natl. Acad. Sci. USA 98 (2001): 11405–11410. [Google Scholar] [CrossRef]
- F. Rodriguez-Trelles, R. Tarrio, and F.J. Ayala. “A methodological bias toward overestimation of molecular evolutionary time scales.” Proc. Natl. Acad. Sci. USA 99 (2002): 8112–8115. [Google Scholar] [CrossRef]
- K.K. Kidd, J.R. Kidd, S.A. Pakstis, C.M. Tishkoff, C.M. Castiglione, and G. Strugo. “Use of linkage disequilibrium to infer population histories.” Am. J. Phys. Anthropol. 22 (1996): 138. [Google Scholar]
- A. Gibbons. “Calibrating the mitochondrial clock.” Science 279 (1998): 28–29. [Google Scholar] [CrossRef]
- K. Christopherson, E. Ullian, C. Stokes, C. Mullowney, J. Hill, A. Agah, J. Lawler, D. Mosher, P. Brownstein, and B. Barres. “Thrombospondins are astrocyte-secreted proteins that promote CNS synaptogenesis.” Cell 120 (2005): 421–433. [Google Scholar] [CrossRef]
- M. Cáceres, C. Suwyn, M. Maddox, J.W. Thomas, and T.M. Preuss. “Increased cortical expression of two synaptogenic thrombospondins in human brain evolution.” Cereb. Cortex 17 (2007): 2312–2321. [Google Scholar]
- E. Jablonka, and M. Lamb. Evolution in Four Dimensions: Genetic, Epigenetic, Behavioral, and Symbolic Variation in the History of Life. Cambridge, MA, USA: MIT Press, 2005. [Google Scholar]
- G. Gutierrez, D. Sanchez, and A. Marin. “A reanalysis of the ancient mitochondrial DNA sequences recovered from Neandertal bones.” Mol. Biol. Evol. 19 (2002): 1359–1366. [Google Scholar] [CrossRef]
- M.W. Walberg, and D.A. Clayton. “Sequence and properties of the human KB cell and mouse L cell D-loop regions of mitochondrial DNA.” Nucleic Acids Res. 9 (1981): 5411–5421. [Google Scholar] [CrossRef]
- A. Torroni, M.T. Lott, M.F. Cabell, Y.-S. Chen, L. Lavergne, and D.C. Wallace. “MtDNA and the origin of Caucasians: Identification of ancient Caucasian-specific haplogroups, one of which is prone to a recurrent somatic duplication in the D-loop region.” Am. J. Hum. Genet. 55 (1994): 760–752. [Google Scholar]
- H. Zischler, H. Geisert, A. von Haeseler, and S. Pääbo. “A nuclear ‘fossil’ of the mitochondrial D-loop and the origin of modern humans.” Nature 378 (1995): 489–492. [Google Scholar]
- M. Pruvost, R. Schwarz, B.V. Correia, S. Champlot, S. Braguier, N. Morel, Y. Fernandez-Jalvo, T. Grange, and E.-M. Geigl. “Freshly excavated fossil bones are best for amplification of ancient DNA.” Proc. Natl. Acad. Sci. USA 104 (2007): 739–744. [Google Scholar]
- J. Hardy, A. Pittman, A. Myers, K. Gwinn-Hardy, H.C. Fung, R. de Silva, M. Hutton, and J. Duckworth. “Evidence suggesting that Homo neanderthalensis contributed the H2 MAPT haplotype to Homo sapiens.” Biochem. Soc. Trans. 33 (2005): 582–585. [Google Scholar] [CrossRef]
- D. Garrigan, Z. Mobasher, T. Severson, J.A. Wilder, and M.F. Hammer. “Evidence for archaic Asian ancestry on the human X chromosome.” Mol. Biol. Evol. 22 (2005): 189–192. [Google Scholar]
- A.R. Templeton. “Haplotype trees and modern human origins.” Yearb. Phys. Anthropol. 48 (2005): 33–59. [Google Scholar]
- R.E. Green, J. Krause, S.E. Ptak, A.W. Briggs, M.T. Ronan, J.F. Simons, L. Du, M. Egholm, J.M. Rothberg, M. Paunovic, and S. Pääbo. “Analysis of one million base pairs of Neanderthal DNA.” Nature 444 (2006): 330–336. [Google Scholar]
- R.E. Green, J. Krause, A.W. Briggs, T. Maricic, U. Stenzel, M. Kircher, N. Patterson, H. Li, W. Zhai, Hsi-Yang, and et al. “A draft sequence of the Neandertal genome.” Science 328 (2010): 710–722. [Google Scholar]
- A. Gibbons. “Close encounters of the prehistoric kind.” Science 328 (2010): 680–684. [Google Scholar]
- M. Krings, A. Stone, R.W. Schmitz, H. Krainitzki, M. Stoneking, and S. Pääbo. “Neandertal DNA sequences and the origin of modern humans.” Cell 90 (1997): 19–30. [Google Scholar] [CrossRef]
- S. Pääbo. “Extraction, characterization, molecular cloning, and enzymatic amplification.” Proc. Natl. Acad. Sci. USA 86 (1989): 1939–1943. [Google Scholar] [CrossRef]
- R. Higuchi, B. Bowman, M. Freiberger, O.A. Ryder, and A.C. Wilson. “DNA sequences from the quagga, and extinct member of the horse family.” Nature 312 (1984): 282–284. [Google Scholar]
- S. Pääbo. “Molecular cloning and ancient Egyptian mummy DNA.” Nature 314 (1985): 644–645. [Google Scholar]
- A. Cooper, C. Mourer-Chauvirp, G.K. Chambers, A. von Haeseler, A.C. Wilson, and S. Pääbo. “Independent origins of New Zealand moas and kiwis.” Proc. Natl. Acad. Sci. USA 89 (1992): 8741–8744. [Google Scholar]
- J.P. Noonan, M. Hofreiter, D. Smith, J.R. Priest, N. Rohland, G. Rabeder, J. Krause, J.C. Detter, S. Pääbo, and E.M. Rubin. “Genomic sequencing of Pleistocene cave bears.” Science 309 (2005): 597–599. [Google Scholar]
- S. Pääbo, H. Poinar, D. Serra, V. Jaenicke-Després, J. Hebler, N. Rohland, M. Kuch, J. Krause, L. Vigilant, and M. Hofreiter. “Genetic analyses from ancient DNA.” Annu. Rev. Genet. 38 (2004): 645–679. [Google Scholar] [CrossRef]
- M. Pruvost, T. Grange, and E.-M. Geigl. “Minimizing DNA contamination by using UNG-coupled quantitative real time PCR on degraded DNA samples: Application to DNA studies.” BioTechniques 38 (2005): 569–575. [Google Scholar]
- L. Orlando, and C. Hänni. “Du nouveau pour l’AND ancien.” Méd. Sci. 8-9 (2008): I-XVI. [Google Scholar]
- J.P. Jolivet, and M. Henry. De la Solution à L’oxyde, Condensation des Cations en Solution Aqueuse, Chimie de Surface des Oxides. Paris, France: InterEditions CNRS, 1994. [Google Scholar]
- T. Lindhal, and B. Nyberg. “Rate depurination of native deoxyribonucleic acid.” Biochemistry 11 (1972): 3610–3618. [Google Scholar]
- E.M. Golenberg, A. Bickel, and P. Weihs. “Effect of highly fragmented DNA on PCR.” Nucleic Acids Res. 24 (1996): 5026–5033. [Google Scholar] [CrossRef]
- G. Gutierrez, and A. Marin. “The most ancient DNA recovered from amber-preserved specimen may not be as ancient as it seems.” Mol. Biol. Evol. 15 (1998): 926–929. [Google Scholar] [CrossRef]
- E.-M. Geigl. “Why ancient DNA research needs taphonomy.” In Biosphere to Lithosphere: New Studies in Vertebrate Taphonomy. Edited by T. O’Connor. Oxford, UK: Oxbow Books, 2005, pp. 79–86. [Google Scholar]
- L. Carlier, J. Couprie, A. le Maire, L. Guilhaudis, I. Milazzo, M. Gondry, D. Davoust, B. Gilquin, and S. Zinn-Justin. “Solution structure of the region 51-160 of human KIN17 reveals an atypical winged helix domain.” Protein Sci. 16 (2007): 2750–2755. [Google Scholar] [CrossRef]
- R.G. Bednarik. “Only time will tell: A review of the methodology of direct rock art dating.” Archaeometry 38 (1996): 1–13. [Google Scholar] [CrossRef]
- R.G. Bednarik. “The dating of rock art: A critique.” J. Archaeol. Sci. 29 (2002): 1213–1233. [Google Scholar] [CrossRef]
- A. Watchman. “A universal standard for reporting the ages of petroglyphs and rock paintings.” In Dating and the Earliest Known Rock Art. Edited by M. Strecker and P. Bahn. Oxford, UK: Oxbow Books, 1999, pp. 1–3. [Google Scholar]
- J. Walser, and A. Furano. “The mutational spectrum of non-CpG DNA varies with CpG content.” Genome Res., 2010. [Google Scholar] [CrossRef]
- E. Strauss. “Can mitochondrial clocks keep time? ” Science 283 (1999): 1435–1438. [Google Scholar] [CrossRef]
- A.R. Templeton. “Gene lineages and human evolution.” Science 272 (1996): 1363. [Google Scholar]
- A. Templeton. “Out of Africa again and again.” Nature 416 (2002): 45–51. [Google Scholar] [CrossRef]
- H.-P. Schulz. “The lithic industry from layers IV-V, Susiluola Cave, western Finland, dated to the Eemian interglacial.” Préhistoire Européenne 16-17 (2002): 7–23. [Google Scholar]
- H.-P. Schulz, B. Eriksson, H. Hirvas, P. Huhta, H. Jungner, P. Purhonen, P. Ukkonen, and T. Rankama. “Excavations at Susiluola Cave.” Suomen Mus. 109 (2002): 5–45. [Google Scholar]
- P. Pavlov, J.I. Svendsen, and S. Indrelid. “Human presence in the European Arctic nearly 40,000 years ago.” Nature 413 (2001): 64–67. [Google Scholar] [CrossRef]
- C. Gamble. The Palaeolithic Societies of Europe. Cambridge, UK: Cambridge University Press, 1999. [Google Scholar]
- R.G. Bednarik. “Antiquity and authorship of the Chauvet rock art.” Rock Art Res. 24 (2007): 21–34. [Google Scholar]
- R.G. Bednarik. “Palaeoart and archaeological myths.” Camb. Archaeol. J. 2 (1992): 27–43. [Google Scholar] [CrossRef]
- R.G. Bednarik. “The Pleistocene art of Asia.” J. World Prehistory 8 (1994): 351–375. [Google Scholar] [CrossRef]
- R.G. Bednarik. “The earliest evidence of palaeoart.” Rock Art Res. 20 (2003): 89–135. [Google Scholar]
- F. Felgenhauer. “Das Paläolithikum von Willendorf in der Wachau, Niederösterreich. Vorbericht über die monographische Bearbeitung.” Forsch. Fortschr. 33 (1959): 152–155. [Google Scholar]
- V. Cabrera Valdés, and J. Bischoff. “Accelerator 14C dates for early Upper Palaeolithic (basal Aurignacian) at El Castillo Cave (Spain).” J. Archaeol. Sci. 16 (1989): 577–584. [Google Scholar] [CrossRef]
- J.L. Bischoff, K.R. Ludwig, J.F. Garcia, E. Carbonell, M. Vaquero, T.W. Stafford, and A.J.T. Jull. “Dating of the basal Aurignacian sandwich at Abric Romani (Catalunya, Spain) by radiocarbon and uranium series.” J. Archaeol. Sci. 21 (1994): 541–551. [Google Scholar] [CrossRef]
- J. González Echegaray, L.G. Freeman, I. Barandiaran, J.M. Apellaniz, K. Butzer, C. Fuentes Vidarte, B. Madariaga, J.A. Gonzalez Morales, and A. Leroi-Gourhan. La Cueva de El Pendo. Madrid, Spain: Bibliotheca Praehistorica Hispana, 1980. [Google Scholar]
- F. Bordes, and J. Labrot. “La stratigraphie du gisement de Roc de Combe (Lot) et ses implications.” Bull. Soc. Préhist. Franç. 64 (1967): 15–28. [Google Scholar]
- F. Champagne, and R. Espitalié. La Piage, Site Préhistorique Sur Lot; No. 15. Paris, France: Mémoires de la Société Préhistorique Française, 1981. [Google Scholar]
- M. Vaquero, M. Esteban, E. Allué, J. Vallverdú, E. Carbonell, and J.L. Bischoff. “Middle Palaeolithic refugium, or archaeological misconception? A new U-series and radiocarbon chronology of Abric Agut (Capellades, Spain).” J. Archaeol. Sci. 29 (2002): 953–958. [Google Scholar] [CrossRef]
- R.G. Bednarik. “Traces of cultural continuity in Middle and Upper Palaeolithic material evidence.” Origini 18 (1995): 47–67. [Google Scholar]
- A. Palma di Cesnola. “Le leptolithique archaïque en Italie.” In Périgordien et Gravettien en Europe. Edited by B. Klíma. Nice, France: Congrès IX, Colloque XV; UISPP, 1976, pp. 66–99. [Google Scholar]
- A. Palma di Cesnola. “L’Uluzzien: Faciès italien du Leptolithique archaïque.” L’Anthropologie 93 (1989): 783–811. [Google Scholar]
- S.L. Kuhn, and A. Bietti. “The late middle and early Upper Paleolithic in Italy.” In The Geography of Neandertals and Modern Humans in Europe and the Greater Mediterranean. Edited by O. Bar-Yosef and D. Pilbeam. Cambridge, MA, USA: Peabody Museum of Archaeology and Ethnology, 2000, pp. 49–76. [Google Scholar]
- S.L. Kuhn, and M.C. Stiner. “The antiquity of hunter-gatherers.” In Hunter-Gatherers: An Interdisciplinary Perspective. Edited by C. Panter-Brick, R.H. Layton and P. Rowley-Conwy. Cambridge, UK: Cambridge University Press, 2001, pp. 99–142. [Google Scholar]
- O. Abel. “Das Lebensbild der eiszeitlichen Tierwelt der Drachenhöhle bei Mixnitz.” In Die Drachenhöhle bei Mixnitz. Edited by O. Abel and G. Kyrle. Vienna, Austria: Speläologische Monographien, 1931, Volume 7-9, pp. 885–920. [Google Scholar]
- E. Bächler. Das Alpine Paläolithikum der Schweiz. Basle, Switzerland: Monographien zur Ur- und Frühgeschichte der Schweiz, 1940, Volume 2. [Google Scholar]
- J. Bayer. “Wildkirchlikultur.” Eiszeit und Urgeschichte 6 (1929): 142. [Google Scholar]
- R.G. Bednarik. “Wall markings of the cave bear.” Stud. Speleol. 9 (1993): 51–70. [Google Scholar]
- S. Brodar. “Zur Frage der Höhlenbärenjagd und des Höhlenbärenkults in den paläolithischen fundstellen Jugoslawiens.” Quartär 9 (1957): 147–159. [Google Scholar]
- H. Cramer. “Der Lebensraum des eiszeitlichen Höhlenbären und die ‘Höhlenbärenjagdkultur’.” Z. Dtsch. Geol. Ges. 93 (1941): 181–196. [Google Scholar]
- G. Kyrle. “Die Höhlenbärenjägerstation.” In Die Drachenhöhle bei Mixnitz. Edited by O. Abel and G. Kyrle. Vienna, Austria: Speläologische Monographien, 1931, pp. 804–962, Band 7-9. [Google Scholar]
- M. Malez. “Das Paläolithikum der Veternicahöhle und der Bärenkult.” Quartär 11 (1959): 171–188. [Google Scholar]
- L. Vértes. “Die Rolle des Höhlenbären im ungarischen Paläolithikum.” Quartär 11 (1959): 151–170. [Google Scholar]
- L.F. Zotz. Altsteinzeitkunde Mitteleuropas. Stuttgart, Germany: F. Enke, 1951. [Google Scholar]
- J. Svoboda. “The Bohunician.” In La Mutation. Edited by J.K. Kozłowski. Liège, Belgium: ERAUL, 1990, pp. 169–192. [Google Scholar]
- J. Svoboda. “The complex origin of the Upper Paleolithic in the Czech and Slovak Republics.” In Before Lascaux: The Complete Record of the Early Upper Paleolithic. Edited by H. Knecht, A. Pike-Tay and R. White. Boca Raton, FL, USA: CRC Press, 1993, pp. 23–36. [Google Scholar]
- M. Anikovich. “Early Upper Paleolithic cultures of eastern Europe.” J. World Prehistory 6 (2005): 205–245. [Google Scholar] [CrossRef]
- P.L. Allsworth-Jones. “The Szeletian: Main trends, recent results, and problems for resolution.” In The Pleistocene Perspective. Edited by M. Day, R. Foley and R. W. London, UK: Allen and Unwin, 1986, pp. 1–25. [Google Scholar]
- N. Kyparissi-Apostolika. Theopetra Cave. Twelve Years of Excavation and Research 1987-1998. Athens, Greece: Institute for Aegean Prehistory, 2000. [Google Scholar]
- Y. Facorellis, N. Kyparissi-Apostolika, and Y. Maniatis. “The cave of Theopetra, Kalambaka: Radiocarbon evidence for 50,000 years of human presence.” Radiocarbon 43 (2001): 1029–1048. [Google Scholar]
- J.K. Kozłowski. “A multiaspectual approach to the origins of the Upper Palaeolithic in Europe.” In The Emergence of Modern Humans. An Archaeological Perspective. Edited by P. Mellars. Edinburgh, UK: Edinburgh University Press, 1990, pp. 419–438. [Google Scholar]
- M.C. Stiner. Honor Among Thieves. A Zooarchaeological Study of Neandertal Ecology. Princeton, NJ, USA: Princeton University Press, 1994. [Google Scholar]
- S.L. Kuhn. Mousterian Lithic Technology. An Ecological Perspective. Princeton, NJ, USA: Princeton University Press, 1995. [Google Scholar]
- J. Riel-Salvatore, and G.A. Clark. “Grave markers. Middle and Early Upper Paleolithic burials and the use of chronotypology in contemporary Paleolithic research.” Curr. Anthropol. 42 (2001): 449–479. [Google Scholar] [CrossRef]
- F.G. Fedele, and B. Giaccio. “Paleolithic cultural change in western Eurasia across the 40,000 BP timeline: Continuities and environmental forcing.” In Exploring the Mind of Ancient Man. Festschrift to Robert G. Bednarik. Edited by P. Chenna Reddy. New Delhi, India: Research India Press, 2007, pp. 292–316. [Google Scholar]
- R.G. Bednarik, G. Kumar, A. Watchman, and R.G. Roberts. “Preliminary results of the EIP project.” Rock Art Res. 22 (2005): 147–197. [Google Scholar]
- X. Gao, and C.J. Norton. “A critique of the Chinese ‘Middle Palaeolithic’.” Antiquity 76 (2002): 397–412. [Google Scholar]
- P.B. Beaumont, and R.G. Bednarik. “On a search for ancestral rock art in the southeastern Kalahari, South Africa.” In Pré actes du congrès, IFRAO Ariège 2010, Pleistocene art of Africa, DVD. Lacombe, Tarascon-sur-Arège, France, 2010. (in press) [Google Scholar]
- G. Bräuer. “New evidence of the transitional period between Neanderthal and modern man.” J. Hum. Evol. 10 (1981): 467–474. [Google Scholar]
- G. Bräuer. “A craniological approach to the origin of anatomically modern Homo sapiens in Africa and implications for the appearance of modern Europeans.” In The Origins of Modern Humans: A World Survey of the Fossil Evidence. Edited by F.H. Smith and F. Spencer. New York, NY, USA: Alan R. Liss, 1984, pp. 327–410. [Google Scholar]
- G. Bräuer. “The evolution of modern humans: A comparison of the African and non-African evidence.” In The Human Revolution: Behavioural and Biological Perspectives on the Origins of Modern Humans. Edited by P. Mellars and C. Stringer. Edinburgh, UK: Edinburgh University Press, 1989, pp. 123–154. [Google Scholar]
- C.B. Stringer. “Human evolution and biological adaptation in the Pleistocene.” In Hominid Evolution and Community Ecology: Prehistoric Human Adaptation in Biological Perspective. Edited by R. Foley. London, UK: Academic Press, 1984, pp. 55–83. [Google Scholar]
- C.B. Stringer. “The fate of the Neanderthals.” Nat. Hist. 93 (1984): 6–12. [Google Scholar]
- C.B. Stringer. “Middle Pleistocene hominid variability and the origin of Late Pleistocene humans.” In Ancestors: The Hard Evidence. Edited by E. Delson. New York, NY, USA: Alan R. Liss, 1985, pp. 289–295. [Google Scholar]
- C.B. Stringer. “The origin of early modern humans: A comparison of the European and non-European evidence.” In The Human Revolution: Behavioural and Biological Perspectives on the Origins of Modern Humans. Edited by P. Mellars and C. Stringer. Edinburgh, UK: Edinburgh University Press, 1989, pp. 232–244. [Google Scholar]
- P. Mellars, and C. Stringer. Introduction. In The Human Revolution: Behavioural and Biological Perspectives on the Origins of Modern Humans. P. Mellars, and C. Stringer. Edinburgh, UK: Edinburgh University Press, 1989, pp. 1–14. [Google Scholar]
- J. Wainscoat. “Out of the garden of Eden.” Nature 325 (1987): 13. [Google Scholar] [CrossRef]
- J.S. Wainscoat, A.V.S. Hill, A.L. Boyce, J. Flint, M. Hernandez, S.L. Thein, J.M. Old, J.R. Lynch, A.G. Falusi, D.J. Weatherall, and et al. “Evolutionary relationships of human populations from an analysis of nuclear DNA polymorphisms.” Nature 319 (1986): 491–493. [Google Scholar]
- S.E. Churchill, and F.H. Smith. “A modern human humerus from the early Aurignacian of Vogelherdhöhle (Stetten, Germany).” Am. J. Phys. Anthropol. 112 (2000): 251–273. [Google Scholar] [CrossRef]
- S.E. Churchill, and F.H. Smith. “Makers of the early Aurignacian of Europe.” Am. J. Phys. Anthropol. 113 (2000): 61–115. [Google Scholar] [CrossRef]
- A. Czarnetzki. “Zur entwicklung des Menschen in Südwestdeutschland.” In Urgeschichte in Baden-Württemberg. Edited by H. Müller Beck. Stuttgart, Germany: Konrad Theiss, 1983, pp. 217–240. [Google Scholar]
- W. Gieseler. Die Fossilgeschichte des Menschen. Stuttgart, Germany: Konrad Theiss, 1974. [Google Scholar]
- N.J. Conard, P.M. Grootes, and F.H. Smith. “Unexpectedly recent dates for human remains from Vogelherd.” Nature 430 (2004): 198–201. [Google Scholar] [CrossRef]
- D. Sonneville-Bordes. “De Position-stratigraphique et chronologique relative des restes humains du Palaéolithique supérieur entre Loire et Pyrénées.” Ann. Paléontol. 45 (1959): 19–51. [Google Scholar]
- H.L. Movius. “The Abri of Cro-Magnon, Les Eyzies (Dordogne) and the probable age of the contained burials on the basis of the nearby Abri Pataud.” Anu. Estud. Atl. 15 (1969): 323–344. [Google Scholar]
- D. Henry-Gambier. “Les fossiles de Cro-Magnon (Les-Eyzies-de-Tayac, Dordogne): Nouvelles données sur leur position chronologique et leur attribution culturelle.” Bull. Mém. Soc. D’Anthropol. Paris 14 (2002): 89–112. [Google Scholar]
- R. White. “Comment on Bednarik, R.G., ‘Concept-mediated markings in the Lower Palaeolithic’.” Curr. Anthropol. 36 (1995): 623–625. [Google Scholar]
- R.G. Bednarik. “Concept-mediated marking in the Lower Palaeolithic.” Curr. Anthropol. 36 (1995): 605–634. [Google Scholar]
- J. Szombathy. “Die diluvialen Menschenreste aus der Fürst-Johanns-Höhle bei Lautsch in Mähren.” Die Eiszeit 2 (1925): 1-34, 73-95. [Google Scholar]
- J. Smyčka. “Litovel a okolí za pravĕku.” Pravĕk 3 (1907): 140–150. [Google Scholar]
- J. Knies. “Nový nález diluviálního človĕka u Mladče na Moravĕ.” Vĕstník klubu přírodovĕdného v Prostĕjovĕ 7 (1906): 1–19. [Google Scholar]
- J. Smyčka. “Nálezy diluviálního člověka v Mladči u Litovle na Moravě.” Obzor Prehistorický 1 (1922): 111–120. [Google Scholar]
- J. Smyčka. “Kdy přišel první člověk do litovelského kraje.” In Vlastivědný sborník střední a severní Moravy III. Litovel, Czech Republic: příloha Olomoucko a Litovelsko, 1925, pp. 6–7. [Google Scholar]
- E. Weiser. Reise und Wanderbuch, 2nd ed. Freudenthal, Germany: Ernst Feld, 1928. [Google Scholar]
- J. Jelínek. “Historie, identifikace a význam mladečských antropologických nálezů z počátku mladého paleolitu.” Anthropologie 25 (1987): 51–69. [Google Scholar]
- J.A. Svoboda, J. van der Plicht, and V. Kuzelka. “Upper Palaeolithic and Mesolithic human fossils from Moravia and Bohemia (Czech Republic): Some new 14C dates.” Antiquity 76 (2002): 957–962. [Google Scholar]
- R.G. Bednarik. “The cave art of Mladeč cave, Czech Republic.” Rock Art Res. 23 (2006): 207–216. [Google Scholar]
- E.M. Wild, M. Teschler-Nicola, W. Kutschera, P. Steier, E. Trinkaus, and W. Wanek. “Direct dating of early Upper Palaeolithic human remains from Mladeč.” Nature 435 (2005): 332–335. [Google Scholar]
- F.H. Smith. “Upper Pleistocene hominid evolution in south-central Europe: A review of the evidence and analysis of trends.” Curr. Anthropol. 23 (1982): 667–686. [Google Scholar]
- F.H. Smith. “Continuity and change in the origin of modern Homo sapiens.” Z. Morphol. Anthropol. 75 (1985): 197–222. [Google Scholar]
- D.W. Frayer. “Cranial variation at Mladeč and the relationship between Mousterian and Upper Palaeolithic hominids.” Anthropologie 23 (1986): 243–256. [Google Scholar]
- E. Trinkaus, and M. Le May. “Occipital bunning among Later Pleistocene hominids.” Am. J. Phys. Anthropol. 57 (1982): 27–35. [Google Scholar] [CrossRef]
- J. Jelínek, M.H. Wolpoff, and D.W. Frayer. “Evolutionary significance of the quarry cave specimens from Mladeč.” Anthropologie 43 (2005): 215–228. [Google Scholar]
- G. Bräuer. “Die morphologischen Affinitäten des jungpleistozänen Strinbeins aus dem Elbmündungsgebiet bei Hahnöfersand.” Z. Morphol. Anthropol. 71 (1980): 1–42. [Google Scholar]
- T. Terberger, and M. Street. “Jungpaläolithische Menschenreste im westlichen Mitteleuropa und ihr Kontex.” In Erkenntnisjäger: Kultur und Umwelt des Frühen Menschen. Edited by J.M. Burdukiewicz, L. Fiedler, W.-D. Heinrich, A. Justus and E. Brühl. Halle, Germany: Veröffentlichungen des Landesamtes für Archäologie Sachsen-Anhalt–Landesmuseum für Vorgeschichte, 2003, Volume 57/2, pp. 579–591. [Google Scholar]
- W. Henke, and H. Rothe. Paläoanthropologie. Berlin, Germany: Wiley-VCH Verlag GmbH, 1994. [Google Scholar]
- F.H. Smith, E. Trinkaus, P.B. Pettitt, I. Karavanić, and M. Paunović. “Direct radiocarbon dates for Vindija G1 and Velika Pećina Late Pleistocene hominid remains.” Proc. Natl. Acad. Sci. USA 96 (1999): 12281–12286. [Google Scholar] [CrossRef]
- E. Trinkaus, O. Moldovan, ş. Milota, A. Bîlgar, L. Sarcina, S. Athreya, S.E. Bailey, R. Rodrigo, G. Mircea, T. Higham, and et al. “An early modern human from the Peştera cu Oase, Romania.” Proc. Natl. Acad. Sci. USA 100 (2003): 11231–11236. [Google Scholar]
- R. Rodrigo, M. Gherase, L. Sarcină, O. Moldovan, R.G. Constantin, C. Franciscus, P.E. Zollikofer, M. Ponce de León, and et al. “Peştera cu Oase 2 and the cranial morphology of early modern Europeans.” Proc. Natl. Acad. Sci. USA 104 (2007): 1165–1170. [Google Scholar]
- A. Soficaru, A. Doboş, and E. Trinkaus. “Early modern humans from the Peştera Muierii, Baia de Fier, Romania.” Proc. Natl. Acad. Sci. USA 103 (2006): 17196–171201. [Google Scholar] [CrossRef]
- P. Geay. “Sur la découverte d’un squelette aurignacien en Charente-Maritime.” Bull. Soc. Préhistoroque Française 54 (1957): 193–197. [Google Scholar] [CrossRef]
- H. Klaatsch, and O. Hauser. “Homo Aurignaciensis Hauseri.” Prähistorische Z. 1 (1910): 273–338. [Google Scholar] [CrossRef]
- M. Perpère. “L’aurignacien en Poitou-Charentes (Étude des Collections D’industries Lithiques).” Ph.D. Thesis, University of Paris, Paris, France, 1971. [Google Scholar]
- G. Asmus. “Kritische Bemerkungen und neue Gesichtspunkte zur jungpaläolithischen Bestattung von Combe-Capelle, Périgord.” Eiszeitalter Ggw. 15 (1964): 181–186. [Google Scholar]
- M. Perpère. “Les grands gisements aurignaciens du Poitou.” L’Anthropologie 77 (1973): 683–716. [Google Scholar]
- F.V. Ramirez Rozzi, F. d’Errico, M. Vanhaeren, P.M. Grootes, B. Kerautret, and V. Dujardin. “Cutmarked human remains bearing Neandertal features and modern human remains associated with the Aurignacian at Les Rois.” J. Anthropol. Sci. 87 (2009): 153–185. [Google Scholar]
- E. Trinkaus. “European early modern humans and the fate of the Neandertals.” Proc. Natl. Acad. Sci. USA 104 (2007): 7367–7372. [Google Scholar] [CrossRef]
- H. Shang, H. Tong, S. Zhang, F. Chen, and E. Trinkaus. “An early modern human from Tianyuan Cave, Zhoukoudian, China.” Proc. Natl. Acad. Sci. USA 104 (2007): 6573–6578. [Google Scholar]
- I. McDougall, F.H. Brown, and J.G. Fleagle. “Stratigraphic placement and age of modern humans from Kibish, Ethiopia.” Nature 433 (2005): 733–736. [Google Scholar]
- T.D. White, B. Asfaw, D. DeGusta, H. Gilbert, G.D. Richards, G. Suwa, and F.C. Howell. “Pleistocene Homo sapiens from Middle Awash, Ethiopia.” Nature 425 (2003): 742–747. [Google Scholar]
- T. Terberger. “Endmesolithische Funde von Drigge, Lkr. Rügen—Kannibalen auf Rügen? ” Jahrb. Bodendenkmalpflege Mecklenburg-Vorpommern 46 (1998): 7–44. [Google Scholar]
- R. Lewin. Human Evolution: An Illustrated Introduction. Oxford, UK: Blackwell Publishing, 2005. [Google Scholar]
- J. Hawks. “Have Neanderthals left us their genes? ” In Human Evolution: Abstracts of Papers Presented at the 1997 Cold Spring Harbor Symposium on Human Evolution Arranged by L. L. Cavalli-Sforza and J. D. Watson. Edited by L. Cavalli-Sforza. Cold Spring Harbor, NY, USA: Cold Spring Harbor Laboratory, 1997, p. 81. [Google Scholar]
- D.W. Frayer. “Evolution at the European edge: Neanderthal and Upper Paleolithic relationships.” Préhist. Européenne 2 (1993): 9–69. [Google Scholar]
- D.W. Frayer. “Perspectives on Neandertals as ancestors.” In Conceptual Issues in Modern Human Origins Research. Edited by G.A. Clark and C.M. Willermet. New York, NY, USA: Aldine de Gruyter, 1998, pp. 220–234. [Google Scholar]
- D.W. Frayer, M.H. Wolpoff, A.G. Thorne, F.H. Smith, and G.G. Pope. “Getting it straight.” Am. Anthropol. 96 (1994): 424–438. [Google Scholar]
- R. White. “Technological and social dimensions of Aurignacian-age body ornaments across Europe.” In Before Lascaux: The Complex Record of the Early Upper Palaeolithic. Edited by H. Knecht, A. Pike-Tay and R. White. Boca Raton, FL, USA: CRC Press, 1993, pp. 277–299. [Google Scholar]
- J.-J. Hublin, F. Spoor, M. Braun, F. Zonneveld, and S. Condemi. “A late Neanderthal associated with Upper Palaeolithic artefacts.” Nature 381 (1996): 224–226. [Google Scholar]
- I. Karavanic, and H. Smith. “The Middle/Upper Palaeolithic interface and the relationship of Neanderthals and early modern humans in the Hrvatsko Zagorje, Croatia.” J. Hum. Evol. 34 (1998): 223–248. [Google Scholar] [CrossRef]
- V. Gábori-Csánk. Le Jankovichien: Une Civilisation Paléolithiques en Hongrie. Liège, Belgium: ERAUL 53, 1993. [Google Scholar]
- O.N. Bader. Sungir: Verkhnepaleoliticheskaya Stoyanka. Moscow, Russia: Izdatel’stvo “Nauka”, 1978. [Google Scholar]
- F.H. Smith, and G. Ranyard. “Evolution of the supraorbital region in Upper Pleistocene fossil hominids from south-central Europe.” Am. J. Phys. Anthropol. 53 (1980): 589–610. [Google Scholar] [CrossRef]
- M. Wolpoff. Paleoanthropology, 2nd ed. New York, NY, USA: McGraw-Hill, 1999. [Google Scholar]
- T. Higham, C.B. Ramsey, I. Karavanić, F.H. Smith, and E. Trinkaus. “Revised direct radiocarbon dating of the Vindija G1 Upper Paleolithic Neandertals.” Proc. Natl. Acad. Sci. USA 103 (2006): 553–557. [Google Scholar]
- J.C.M. Ahern, I. Karavanic, M. Paunović, I. Janković, and F.H. Smith. “New discoveries and interpretations of fossil hominids and artifacts from Vindija Cave, Croatia.” J. Hum. Evol. 46 (2004): 25–65. [Google Scholar]
- F. D’Errico. “Comment on Bednarik, R.G., ‘Concept-mediated markings of the Lower Palaeolithic’.” Curr. Anthropol. 36 (1995): 618–620. [Google Scholar]
- P. Mellars. “The impossible coincidence. A single-species model for the origins of modern human behavior in Europe.” Evol. Anthropol. 14 (2005): 12–27. [Google Scholar] [CrossRef]
- S. Oyama. Evolution’s Eye: A Systems View of the Biology-Culture Divide. Durham, NC, USA: Duke University Press, 2000. [Google Scholar]
- S. Oyama, P.E. Griffiths, and R.D. Gray. Cycles of Contingency: Developmental Systems and Evolution. Cambridge, MA, USA: MIT Press, 2001. [Google Scholar]
- F.J. Odling-Smee, K.N. Laland, and M.W. Feldman. Niche Construction: The Neglected Process in Evolution. Princeton, NJ, USA: Princeton University Press, 2003. [Google Scholar]
- R.G. Bednarik. “On the cognitive development of hominids.” Man Environ. 15 (1990): 1–7. [Google Scholar]
- K. Laland, J. Odling-Smee, and M.W. Feldman. “Niche construction, biological evolution and cultural change.” Behav. Brain Sci. 23 (2000): 131–175. [Google Scholar] [CrossRef]
- J.B. Silk. “Social component of fitness in primate groups.” Science 317 (2007): 1347–1351. [Google Scholar] [CrossRef]
- T. Dobzhansky. Mankind Evolving: The Evolution of the Human Species. New Haven, CT, USA: Yale University Press, 1962. [Google Scholar]
- T. Dobzhansky. “On the evolutionary uniqueness of man.” In Evolutionary Biology. Edited by T. Dobzhansky, M.K. Hecht and W.C. Steere. New York, NY, USA: Appleton-Century-Crofts, 1972, pp. 415–430. [Google Scholar]
- R. Boyd, and P.J. Richerson. The Origin and Evolution of Cultures. New York, NY, USA: Oxford University Press, 2005. [Google Scholar]
- P.J. Richerson, and R. Boyd. Not by Genes Alone: How Culture Transformed Human Evolution. Chicago, IL, USA: University of Chicago Press, 2005. [Google Scholar]
- A. Fuentes. Evolution of Human Behavior. New York, NY, USA; Oxford, UK: Oxford University Press, 2009. [Google Scholar]
- J.G. Mendel. “Versuche über Pflanzen-Hybriden.” Verh. Naturforsch. Ver. Brünn 4 (1866): 3–47. [Google Scholar]
- R.G. Bednarik. “The mythical moderns.” J. World Prehistory 21 (2008): 85–102. [Google Scholar] [CrossRef]
- R.G. Bednarik. “Children as Pleistocene artists.” Rock Art Res. 25 (2008): 173–182. [Google Scholar]
- R.G. Bednarik. “The domestication of humans.” Anthropologie 46 (2008): 1–17. [Google Scholar]
- L.C. Aiello, and P. Wheeler. “The expensive-tissue hypothesis: The brain and the digestive system in human and primate evolution.” Curr. Anthropol. 36 (1995): 199–221. [Google Scholar]
- M. Biesele. Women Like Meat. The Folklore and Foraging Ideology of the Kalahari Ju/’Hoan. Johannesburg, South Africa: Witwatersrand University Press, 1993. [Google Scholar]
- T. Deacon. The Symbolic Species. New York, NY, USA: Norton, 1997. [Google Scholar]
- C. Boehm. “Egalitarian society and reverse dominance hierarchy.” Curr. Anthropol. 34 (1993): 227–254. [Google Scholar]
- C. Boehm. Hierarchy of the Forest: The Evolution of Egalitarian Behavior. Cambridge, MA, USA: Harvard University Press, 1999. [Google Scholar]
- K. Grammer, and R. Thornhill. “Human facial attractiveness and sexual selection: The role of symmetry and averageness.” J. Comp. Psychol. 108 (1994): 233–242. [Google Scholar] [CrossRef]
- T.K. Shackelford, and R.J. Larsen. “Facial asymmetry as an indicator of psychological, emotional, and physiological distress.” J. Pers. Soc. Psychol. 72 (1997): 456–466. [Google Scholar] [CrossRef]
- D.M. Jones. “Sexual selection, physical attractiveness and facial neoteny: Cross-cultural evidence and implications.” Curr. Anthropol. 36 (1995): 723–748. [Google Scholar]
- D.M. Jones. “An evolutionary perspective on physical attractiveness.” Evol. Anthropol. 5 (1996): 97–109. [Google Scholar] [CrossRef]
- B. Delluc, and G. Delluc. “Les manifestations graphiques aurignaciens sur support rocheux des environs des Eyzies (Dordogne).” Gallia Préhistoire 21 (1978): 213–438. [Google Scholar]
- L. Chiotti, B. Delluc, and G. Delluc. “Art et parure aurignaciens de l’abri Pataud (Les Eyzies-de-Tayac, Dordogne, France) dans le contexte aurignacien Périgord.” In Les Chemins de L’art Aurignacien en Europe—Das Aurignacien und die Anfänge der Kunst in Europa. Edited by H. Floss and N. Rouquerol. Aurignac, France: Éditions Musée-forum Aurignac, 2007, pp. 171–186. [Google Scholar]
- R.G. Bednarik. “The Galgenberg figurine from Krems, Austria.” Rock Art Res. 6 (1989): 118–125. [Google Scholar]
- N.J. Conard. “A female figurine from the basal Aurignacian of Hohle Fels cave in southwestern Germany.” Nature 459 (2009): 248–252. [Google Scholar] [CrossRef]
- P. Mellars. “Origins of the female image.” Nature 459 (2009): 176–177. [Google Scholar] [CrossRef]
- K.N. Laland. “Sexual selection with a culturally transmitted mating preference.” Theor. Popul. Biol. 45 (1994): 1–15. [Google Scholar] [CrossRef]
- L.L. Cavalli-Sforza, and M.W. Feldman. “Cultural vs. biological inheritance.” Am. J. Hum. Genet. 25 (1973): 618–637. [Google Scholar]
- M.W. Feldman, and L.L. Cavalli-Sforza. “On the theory of evolution under genetic and cultural transmission with application to the lactose absorption problem.” In Mathematical Evolutionary Theory. Edited by M.W. Feldman. Princeton, USA, NJ: Princeton University Press, 1989, pp. 145–173. [Google Scholar]
- K. Aoki, and M.W. Feldman. “Recessive hereditary deafness, assortative mating, and persistence of a sign language.” Theor. Popul. Biol. 39 (1991): 358–372. [Google Scholar] [CrossRef]
- W.H. Durham. Coevolution: Genes, Culture, and Human Diversity. Stanford, CA, USA: Stanford University Press, 1991. [Google Scholar]
- J.B.S. Haldane. The Causes of Evolution. Edited by London Longmans Green & Co.:. New York, NY, USA: Harper Brothers, 1932. [Google Scholar]
- G.R. De Beer. Embryos and Ancestors. Oxford, UK: Oxford University Press, 1940. [Google Scholar]
- M.F. Ashley Montagu. An Introduction to Physical Anthropology. Springfield, IL, USA: Thomas, 1960. [Google Scholar]
- C.R. Badcock. The Psychoanalysis of Culture. Oxford, UK: Basil Blackwell, 1980. [Google Scholar]
- G.R. De Beer. Embryology and Evolution. Oxford, UK: Oxford University Press, 1930. [Google Scholar]
- W.R. Leonard. “Food for thought: Dietary change was a driving force in human evolution.” Sci. Am. 287 (2002): 106–115. [Google Scholar] [CrossRef]
- W.R. Leonard, and M.L. Robertson. “Nutritional requirements and human evolution: A bioenergetics model.” Am. J. Hum. Biol. 4 (1992): 179–195. [Google Scholar] [CrossRef]
- W.R. Leonard, and M.L. Robertson. “Evolutionary perspectives on human nutrition: The influence of brain and body size on diet and metabolism.” Am. J. Hum. Biol. 6 (1994): 77–88. [Google Scholar] [CrossRef]
- W.R. Leonard, and M.L. Robertson. “Comparative primate energetics and hominid evolution.” Am. J. Phys. Anthropol. 102 (1997): 265–281. [Google Scholar] [CrossRef]
- P. Andolfatto. “Adaptive hitchhiking effects on genome variability.” Curr. Opin. Genet. Dev. 11 (2001): 635–641. [Google Scholar] [CrossRef]
- J. Lu, T. Tang, H. Tang, J. Huang, S. Shi, and C.-I. Wu. “The accumulation of deleterious mutations in rice genomes: A hypothesis on the cost of domestication.” Trends Genet. 22 (2006): 126–131. [Google Scholar] [CrossRef]
- D.C. Rubinsztein, W. Amos, J. Leggo, S. Goodburn, R.S. Ramesar, J. Old, R. Dontrop, R. McMahon, D.E. Barton, and M.A. Ferguson-Smith. “Mutational bias provides a model for the evolution of Huntington’s disease and predicts a general increase in disease prevalence.” Nat. Genet. 7 (1994): 525–530. [Google Scholar] [CrossRef]
- L.C. Walker, and L.C. Cork. “The neurobiology of aging in nonhuman primates.” In Alzheimer’s Disease, 2nd. Edited by R.D. Terry, R. Katzman, K.L. Bick and S.S. Sisodia. New York, NY, USA: Lippincott Williams and Wilkins, 1999, pp. 233–243. [Google Scholar]
- M.V. Olson, and A. Varki. “Sequencing the chimpanzee genome: Insights into human evolution and disease.” Nat. Rev. Genet. 4 (2003): 20–28. [Google Scholar] [CrossRef]
- R.G. Bednarik, and P.A. Helvenston. “The nexus between neurodegeneration and advanced cognitive abilities.” Anthropos 106 (in press). (2011). [Google Scholar]
- C.C. Sherwood, A.D. Gordon, J.S. Allen, K.A. Phillips, J.M. Erwin, P.R. Hof, and W.D. Hopkins. “Aging of the cerebral cortex differs between humans and chimpanzees.” Proc. Natl. Acad. Sci. USA, 2011. [Google Scholar] [CrossRef]
- M. Marvanová, J. Ménager, E. Bezard, R.E. Bontrop, L. Pradier, and G. Wong. “Microarray analysis of nonhuman primates: Validation of experimental models in neurological disorders.” FASEB J. 17 (2003): 929–931. [Google Scholar]
- W. Enard, P. Khaitovich, J. Klose1, F. Heissig, S. Zöllner, P. Giavalisco, K. Nieselt-Struwe, E. Muchmore, A. Varki, R. Ravid, and et al. “Intra- and interspecific variation in primate gene expression patterns.” Science 296 (2002): 340–343. [Google Scholar]
- G. Arora, N. Polavarapu, and J.F. McDonald. “Did natural selection for increased cognitive ability in humans lead to an elevated risk for cancer? ” Med. Hypotheses 73 (2009): 453–436. [Google Scholar]
- D. Horrobin. The Madness of Adam and Eve: How Schizophrenia Shaped Humanity. London, UK: Bantam Press, 2002. [Google Scholar]
- S.W. Gangestad, and R.W. Yeo. “Behavioral genetic variation, adaptation and maladaptation: An evolutionary perspective.” Trends Cognit. Sci. 1 (1997): 103–108. [Google Scholar] [CrossRef]
- M.C. Keller, and G. Miller. “Resolving the paradox of common, harmful, heritable mental disorders: Which evolutionary genetic models work best? ” Beha. Brain Sci. 29 (2006): 385–452. [Google Scholar]
- M.C. Keller. “The evolutionary persistence of genes that increase mental disorders risk.” Curr. Dir. Psychol. Sci. 17 (2008): 395–399. [Google Scholar] [CrossRef]
- M.M. Brown, C. Kaplan, L.A. Jason, and C.B. Keys. “Subgroups of chronic fatigue syndrome based on psychiatric disorder onset and current psychiatric status.” Health 2 (2010): 90–96. [Google Scholar] [CrossRef]
- J.M. Castellano, J Kim, F.R. Stewart, H. Jiang, R.B. DeMattos, B.W. Patterson, A.M Fagan, J.C. Morris, K.G. Mawuenyega, C. Cruchaga, and et al. “Human apoE isoforms differentially regulate blain amyloid-β peptide clearance.” Sci. Transl. Med. 3 (2011): 89. [Google Scholar] [CrossRef]
- B.T. Hyman, G.W. van Hoesen, A.R. Damasio, and C.L. Barnes. “Alzheimer’s disease: Cell-specific pathology isolates the hippocampal formation.” Science 225 (1984): 1168–1170. [Google Scholar]
- D.A. Smith. “Imaging the progression of Alzheimer pathology through the brain.” Proc. Natl. Acad. Sci. USA 99 (2002): 4135–4137. [Google Scholar]
- W.W. Seeley, D.A. Carlin, and J.M. Allman. “Early frontotemporal dementia targets neurons unique to apes and humans.” Ann. Neurol. 60 (2006): 660–667. [Google Scholar]
- W.W. Seeley, J.M. Allman, D.A. Carlin, R.K. Crawford, M.N. Macedo, M.D. Greicius, S.J. Dearmond, and B.L. Miller. “Divergent social functioning in behavioral variant frontotemporal dementia and Alzheimer disease: Reciprocal networks and neuronal evolution.” Alzheimer Dis. Assoc. Disord. 21 (2007): S50–S57. [Google Scholar] [CrossRef]
- J.M. Schwartz, and S. Begley. The Mind and the Brain: Neuroplasticity and the Power of Mental Force. New York, NY, USA: HarperCollins, 2002. [Google Scholar]
- F.K. Goodwin, and K.R. Jamison. Manic Depressive Illness. Oxford, UK: Oxford University Press, 1990. [Google Scholar]
- N. Craddock, and I. Jones. “Genetics of bipolar disorder: Review article.” J. Med. Genet. 26 (1999): 585–594. [Google Scholar]
- J.J. Schildkraut. “The catecholamine hypothesis of affective disorders: A review of supporting evidence.” Am. J. Psychiatry 122 (1965): 509–528. [Google Scholar]
- M.P. Vawter, W.J. Freed, and J.E. Kleinman. “Neuropathology of bipolar disorder.” Biol. Psychiatr. 48 (2000): 486–504. [Google Scholar] [CrossRef]
- G. Rajkowska. “Reductions in neuronal and glial density characterize the dorsolateral prefrontal cortex in bipolar disorder.” Biol. Psychiatr. 49 (2009): 741–752. [Google Scholar] [CrossRef]
- J. Hwang, I.K. Lyoo, S.R. Dager, S.D. Friedman, J.S. Oh, J.Y. Lee, S.J. Kim, D.L. Dunner, and P.F. Renshaw. “Basal ganglia shape alterations in bipolar disorder.” Am. J. Psychiatr. 163 (2006): 276–285. [Google Scholar] [CrossRef]
- M. Preisig, F. Ferrero, and A. Malafosse. “Monoamine oxidase a and tryptophan hydroxylase gene polymorphisms: Are they associated in bipolar disorder? ” Am. J. Pharmacogenet. 5 (2005): 45–52. [Google Scholar] [CrossRef]
- N. Kuroki, M.E. Shenton, D.F. Salisbury, Y. Hirayasu, T. Onitsuka, H. Ersner-Hershfield, D. Yurgelun-Todd, R. Kikinis, F.A. Jolesz, and R.W. McCarley. “Middle and inferior temporal gyrus gray matter volume abnormalities in first-episode schizophrenia: An MRI study.” Am. J. Psychiatr. 163 (2006): 2103–2110. [Google Scholar] [CrossRef]
- P.J. Harrison. “The neuropathology of schizophrenia: A critical review of the data and their interpretation.” Brain 122 (1999): 593–624. [Google Scholar] [CrossRef]
- D.H. Mathalon, and J.M. Ford. “Divergent approaches converge on frontal lobe dysfunction in schizophrenia.” Am. J. Psychiatr. 154 (2008): 944–948. [Google Scholar] [CrossRef]
- J.M. McClellan, E. Susser, and M.C. King. “Schizophrenia: A common disease caused by multiple rare alleles.” Br. J. Psychiatr. 190 (2007): 194–199. [Google Scholar] [CrossRef]
- C.A. Tamminga, G.K. Thaker, R. Buchanan, B. Kirkpatrick, L.D. Alphs, T.N. Chase, and W.T. Carpenter. “Limbic system abnormalities identified in schizophrenia using PET with fluorodeoxyglucose.” Arch. Gen. Psychiatr. 49 (1992): 522–530. [Google Scholar] [CrossRef]
- H.J. Cho, I. Meira-Lima, Q. Cordeiro, L. Michelon, P.C. Sham, H. Vallada, and D.A. Collier. “Population-based and family-based studies on the serotonin transporter gene polymorphisms and bipolar disorder: A systematic review and meta-analysis.” Mol. Psychiatr. 10 (2005): 771–781. [Google Scholar] [CrossRef]
- D. Li, D.A. Collier, and L. He. “Meta-analysis shows strong positive association of the neuregulin 1 (NRG1) gene with schizophrenia.” Hum. Mol. Genet. 15 (2006): 1995–2002. [Google Scholar] [CrossRef]
- M. Sailor, B. Fischl, D. Salat, C. Tempelmann, E. Busa, N. Bodammer, H.-J. Heinzel, and A. Dale. “Focal cortical thinning of the cerebral cortex in multiple sclerosis.” Brain 126 (2003): 1734–1744. [Google Scholar] [CrossRef]
- C.E. Lumsden. “The neuropathology of multiple sclerosis.” In Handook of Clinical Neurology. Edited by P.J. Vinken and G.W. Bruyn. Amsterdam, The Netherlands: North-Holland, 1970, Volume 9, pp. 217–309. [Google Scholar]
- C. Wegner, and P.M. Mathews. “A new view of the cortex, new insights into multiple sclerosis.” Brain 126 (2003): 1810–1821. [Google Scholar]
- J.M. Allman, K.K. Watson, N.A. Tetreault, and A.Y. Hakeem. “Intuition and autism: A possible role for von Economo neurons.” Trends Cognit. Sci. 9 (2005): 367–373. [Google Scholar] [CrossRef]
- D.P. Kennedy, K. Semendeferi, and E. Courchesne. “No reduction of spindle neuron number in the frontoinsular cortex in autism.” Brain Cognit. 64 (2007): 124–129. [Google Scholar] [CrossRef]
- C. Hughes, I. Soares-Boucaud, J. Hochmann, and U. Frith. “Social behaviour in pervasive developmental disorders: Effects of informant, group and ‘theory-of-mind’.” Eur. Child Adolesc. Psychiatr. 6 (1997): 191–198. [Google Scholar]
- D.F. Benson, and D. Blumer. Psychiatric Aspects of Neurologic Disease. New York, NY, USA: Grune and Stratton, 1975. [Google Scholar]
- F.O. Volle, and P.A. Heron-Helvenston. Epilepsy and You. Springfield, IL, USA: Charles C. Thomas Publishing Company, 1979. [Google Scholar]
- A.R. Damasio, D. Tranel, and H. Damasio. “Individuals with sociopathic behavior caused by frontal damage fail to respond autonomically to social stimuli.” Behav. Brain Res. 41 (1990): 81–94. [Google Scholar] [CrossRef]
- K. Brodmann. “Neue Ergebnisse über die vergleichende histologische Lokalisation der Grosshirnrinde mit besonderer Berücksichtigung des Stirnhirns.” Anat. Anz. 41 (1912): 157–216. [Google Scholar]
- K. Semendeferi. “Advances in the study of hominoid brain evolution: Magnetic resonance imaging (MRI) and a 3-D reconstruction.” In Evolutionary Anatomy of the Primate Cerebral Cortex. Edited by D. Falk and K.R. Gibson. Cambridge, UK: Cambridge University Press, 2001. [Google Scholar]
- A.R. Damasio. “The frontal lobes.” In Clinical Neuropsychology. Edited by K. Heilman and H. Valenstein. Oxford, UK: Oxford University Press, 1985, pp. 339–375. [Google Scholar]
- D. Hodgson, and P.A. Helvenston. “The emergence of the representation of animals in palaeoart: Insights from evolution and the cognitive, limbic and visual systems of the human brain.” Rock Art Res. 23 (2006): 3–40. [Google Scholar]
- E.A. Nimchinsky, E. Gilissen, J.M. Allman, D.P. Perl, J.M. Erwin, and P. Hof. “A neuronal morphologic type unique to humans and great apes.” Proc. Natl. Acad. Sci. USA 96 (1999): 5268–5273. [Google Scholar]
- K.K. Watson, T.K. Kones, and J.M. Allman. “Dendritic architecture of the von Economo neurons.” Neuroscience 141 (2006): 1107–1112. [Google Scholar] [CrossRef]
- J. Allman, A. Hakeem, and K. Watson. “Two phylogenetic specializations in the human brain.” Neuroscientist 4 (2002): 335–345. [Google Scholar]
- M. Hayashi. “Spindle neurons in the anterior cingulate cortex of humans and great apes.” In Cognitive Development in Chimps. Edited by T. Matsuzawa, M. Tomonaga and M. Tanaka. Tokyo, Japan: Springer-Verlag, 2006, pp. 64–74. [Google Scholar]
- D. Sridharan, D.J. Levitin, and V. Menon. “A critical role for the right fronto-insular cortex in switching between central-executive and default-mode networks.” Proc. Natl. Acad. Sci. USA 105 (2008): 12569–12574. [Google Scholar] [CrossRef]
- T.M. Preuss, and G.Q. Coleman. “Human-specific organization of primary visual cortex: Alternating compartments of dense Cat-301 and calbindin immunoreacitivity in layer 4A.” Cereb. Cortex 12 (2002): 671–691. [Google Scholar] [CrossRef]
- D. Buxhoeveden, and M.F. Casanova. “The minicolumn and evolution of the brain.” Brain Behav. Evol. 60 (2002): 125–151. [Google Scholar] [CrossRef]
- C.C. Sherwood, J.K. Rilling, R.L. Holloway, and P.R. Hof. “Evolution of the brain in humans: Specializations in a comparative perspective.” In Encyclopedia of Neuroscience. Edited by M.D. Binder, N. Hirokawa and U. Windhorst. Berlin, Germany: Springer, 2009, pp. 1334–1338. [Google Scholar]
- E.M. Ullian, S.K. Sapperstein, K.S. Christopherson, and B. Barres. “Control of synapse number by glia.” Science 291 (2001): 569–570. [Google Scholar]
- E.M. Ullian, B.T. Harris, A. Wu, J.R. Chan, and B.A. Barres. “Schwann cells and astrocytes induce synapse formation by spinal motor neurons in culture.” Mol. Cell. Neurosci. 25 (2004): 241–251. [Google Scholar] [CrossRef]
- B.A. Barres. “The mystery and magic of glia: A perspective on their roles in health and disease.” Neuron 60 (2008): 430–440. [Google Scholar] [CrossRef]
- P.R. Hof, and E. van der Gucht. “Structure of the cerebral cortex of the humpback whale. Megaptera novaeangliae (Cetacea, Mysticeti, Balaenopteridae).” Anat. Rec. 290 (2007): 1–31. [Google Scholar] [CrossRef]
- C. Butti, C.C. Sherwood, A.Y. Hakeem, and J.M. Allman. “Total number and volume of von Economo neurons in the cerebral cortex of cetaceans.” J. Comp. Neurol. 515 (2009): 243–259. [Google Scholar] [CrossRef]
- A.Y. Hakeem, C.C. Sherwood, C.J. Bonar, C. Butti, P.R. Hof, and J.M. Allman. “Von Economo neurons in the elephant brain.” Anat. Rec. 292 (2009): 242–248. [Google Scholar] [CrossRef]
- J. Allman, A. Hakeem, J.M. Erwin, E. Nimchinsky, and P. Hof. “The anterior cingulate cortex: The evolution of an interface between emotion and cognition.” Ann. New York Acad. Sci. 935 (2001): 107–117. [Google Scholar]
- E.A. Nimchinsky, B.A. Vogt, J.H. Morrison, and P.R. Hof. “Spindle neurons of the human anterior cingulate cortex.” J. Comp. Neurol. 355 (1995): 27–37. [Google Scholar] [CrossRef]
- K. Semendeferi. “Evolution of the Hominoid Prefrontal Cortex: A Quantitative and Image Analysis of Areas 13 and 10.” Ph.D. Dissertation, University of Iowa, Iowa City, IA, USA, 1994. [Google Scholar]
- L. Heimer, G.W. Van Hoesen, M. Trimble, and D.S. Zahm. Anatomy of Neuropsychiatry: The New Anatomy of the Basal Forebrain and Its Implications for Neuropsychiatric Illness. Burlington, NJ, USA: Academic Press, 2007. [Google Scholar]
- M.S. Mega, and J. Cummings. “The cingulate and cingulate syndromes.” In Contemporary Behavioural Neurology. Edited by M.R. Trimble and J.L. Cummings. Oxford, UK: Butterworths, 1997, pp. 189–214. [Google Scholar]
- R. Adolphs, D. Tranel, H. Damasio, and A.R. Damasio. “Impaired recognition of emotion in facial expression following bilateral damage in the human amygdala.” Nature 372 (1994): 669–672. [Google Scholar] [CrossRef]
- R.D. Adolphs, D. Tranel, S. Hamann, A.W. Young, A.J. Calder, E.A. Phelps, A. Anderson, G.P. Lee, and A.R. Damasio. “Recognition of facial emotion in nine subjects with bilateral amygdala damage.” Neuropsychologia 37 (1999): 1111–1117. [Google Scholar] [CrossRef]
- L.M. Frank, E.N. Brown, and M.A. Wilson. “Trajectory encoding in the hippocampus and entorhinal cortex.” Neuron 27 (2000): 169–178. [Google Scholar] [CrossRef]
- P. Gloor. “Experiential phenomena of temporal lobe epilepsy.” Brain 113 (1990): 1673–1694. [Google Scholar] [CrossRef]
- P. Gloor. “Role of the amygdala in temporal lobe epilepsy.” In The Amygdala. Edited by A.P. Aggleton. New York, NY, USA: Wiley-Liss Press, 1992. [Google Scholar]
- T.M. Preuss. “The discovery of cerebral diversity: An unwelcome scientific revolution.” In Evolutionary Anatomy of the Primate Cerebral Cortex. Edited by D. Falk and K.R. Gibson. Cambridge, MA, USA: Cambridge University Press, 2001, pp. 138–164. [Google Scholar]
- T.M. Preuss, and J.H. Kaas. “Human brain evolution.” In Fundamental Neuroscience. Edited by F.E. Bloom, S.C. Landes, J.L. Robert, L.R. Squire and M.J. Zigmond. San Diego, CA, USA: Academic Press, 1999, pp. 1283–1311. [Google Scholar]
- R.L. Holloway. “Toward a synthetic theory of human brain evolution.” In Origins of the Human Brain. Edited by J.P. Changeaux and J. Chavillon. Oxford, UK: Clarendon Press, 1995, pp. 42–55. [Google Scholar]
- R.L. Holloway. “Evolution of the human brain.” In Handbook of Human Symbolic Evolution. Edited by A. Lock and C. Peters. Oxford, UK: Clarendon Press, 1996, pp. 74–116. [Google Scholar]
- R.L. Holloway. “Revisiting australopithecine visual striate cortex: Newer data from chimpanzee and human brains suggest it could have been reduced during australopithecine times.” In Evolutionary Anatomy of the Primate Cerebral Cortex. Edited by D. Falk and K.R. Gibson. New York, NY, USA: Cambridge University Press, 2001, pp. 177–186. [Google Scholar]
- R.-A. Rathelot, and P.L. Strick. “Subdivisions of primary motor cortex based on cortico-motoneuronal cells.” Proc. Natl. Acad. Sci. USA 106 (2009): 918–923. [Google Scholar] [CrossRef]
- H.C. Leiner, A.L. Leiner, and R.S. Dow. “Human brain map.” In The Cerebellum and Cognition. Edited by J.D. Schmahmann. San Diego, CA, USA: Academic Press, 1995, p. 244. [Google Scholar]
- R.-A. Mueller, and E. Courchesne. “The cerebellum: So much more.” Science 181 (1998): 879–880. [Google Scholar]
- N. Geschwind, and W. Levitsky. “Human brain: Left-right asymmetries in temporal speech region.” Science 161 (1968): 186–187. [Google Scholar]
- P.J. Gannon, R.L. Holloway, D.C. Broadfield, and A.R. Braun. “Asymmetry of chimpanzee planum temporale: Humanlike pattern of Wernicke’s brain language area homolog.” Science 279 (1998): 220–222. [Google Scholar] [CrossRef]
- P.J. Gannon, N.M. Kheck, and P.R. Hof. “Language areas of the hominoid brain: A dynamic communicative shift on the upper east side planum.” In Evolutionary Anatomy of the Primate Cerebral Cortex. Edited by D. Falk and K.R. Gibson. New York, NY, USA: Cambridge University Press, 2001, pp. 216–240. [Google Scholar]
- I. Stepniewska, T.M. Preuss, and J.H. Kaas. “Architectonics, somatotopic organization and ipsilateral cortical connections of the primary motor area (M1) of owl monkeys.” J. Comp. Neurol. 330 (1993): 238–271. [Google Scholar] [CrossRef]
- D. Buxhoeveden, W. Lefkowitz, P. Loats, and E. Armstrong. “The linear organization of cell columns in human and nonhuman anthropoid Tpt cortex.” Anat. Embryol. 194 (1996): 23–36. [Google Scholar]
- H.J. Jerison. Evolution of the Brain and Intelligence. New York, NY, USA: Academic Press, 1973. [Google Scholar]
- B. Kolb, and I.Q. Whishaw. Fundamentals of Human Neuropsychology, 6th ed. New York, NY, USA: Worth Publishing Co., 2008. [Google Scholar]
- R. Martin. “Scaling of the mammalian brain: The maternal energy hypothesis.” News Physiol. Sci. 11 (1996): 149–156. [Google Scholar]
- S.M. Blinkov, and I.I. Glezer. The Human Brain in Figures and Tables: A Quantitative Handbook. New York, NY, USA: Basic Books, Inc., Plenum Press, 1968. [Google Scholar]
- K. Semendeferi. “The evolution of the frontal lobes: A volumetric analysis based on three-dimensional reconstruction’s of magnetic resonance scans of human and ape brains.” J. Hum. Evol. 32 (1997): 375–388. [Google Scholar] [CrossRef]
- P.M. Kaskan, and B.L. Finley. “Encephalization and its developmental structure: How many ways can a brain get big.” In Evolutionary Anatomy of the Primate Cerebral Cortex. Edited by D. Falk and K.R. Gibson. New York, NY, USA: Cambridge University Press, 2001, pp. 14–29. [Google Scholar]
- P. Rakic, and D.R. Kornack. “Neocortical expansion and elaboration during primate evolution: A view from neuroembryology.” In Evolutionary Anatomy of the Primate Cerebral Cortex. Edited by D. Falk and K.R. Gibson. New York, NY, USA: Cambridge University Press, 2001, pp. 30–56. [Google Scholar]
- R. Ochse. Before the Gates of Excellence: The Determinants of Creative Genius. Cambridge, UK: Cambridge University Press, 1990. [Google Scholar]
- R.G. Bednarik. “Neurophysiology and palaeoart.” AURA Newsl. 25 (2008): 6–16. [Google Scholar]
- E.H. Hare. “Schizophrenia as a recent disease.” Br. J. Psychiatr. 153 (1988): 521–531. [Google Scholar] [CrossRef]
- B.F. Voight, S. Kudaravalli, X. Wen, and J.K. Pritchard. “A map of recent positive selection in the human genome.” PLoS Biol. 4 (2006): e72. [Google Scholar] [CrossRef]
- S. Wright. “The evolution of dominance.” Am. Nat. 63 (1929): 556–561. [Google Scholar]
- M. Kimura. “Evolutionary rate at the molecular level.” Nature 217 (1968): 624–626. [Google Scholar] [CrossRef]
- M. Kimura. The Neutral Theory of Molecular Evolution. Cambridge, UK: Cambridge University Press, 1983. [Google Scholar]
- D.T. Suzuki, A.J.F. Griffiths, J.H. Miller, and R.C. Lewontin. An Introduction to Genetic Analysis, 4th ed. New York, NY, USA: W. H. Freeman, 1989. [Google Scholar]
- R.G. Bednarik. “The initial peopling of Wallacea and Sahul.” Anthropos 92 (1997): 355–367. [Google Scholar]
- R.G. Bednarik. “The origins of navigation and language.” Artefact 20 (1997): 16–56. [Google Scholar]
- R.G. Bednarik. “The earliest evidence of ocean navigation.” Int. J. Nautic. Archaeol. 26 (1997): 183–191. [Google Scholar]
- R.G. Bednarik. “Maritime navigation in the Lower and Middle Palaeolithic.” C. R. L’Académie Sci. Series IIA 328 (1999): 559–563. [Google Scholar]
- R.G. Bednarik. “Pleistocene seafaring in the Mediterranean.” Anthropologie 37 (1999): 275–282. [Google Scholar]
- R.G. Bednarik. “Bednarik, R.G.” Quartär 49/50 (1999): 95–109. [Google Scholar]
- R.G. Bednarik. “Replicating the first known sea travel by humans: The Lower Pleistocene crossing of Lombok Strait.” Hum. Evol. 16 (2001): 229–242. [Google Scholar] [CrossRef]
- R.G. Bednarik. “Seafaring in the Pleistocene.” Camb. Archaeol. J. 13 (2003): 41–66. [Google Scholar] [CrossRef]
- R.G. Bednarik. “Early seafaring.” In Encyclopaedia of the History of Science, Technology, and Medicine in Non-Western Cultures. Edited by H. Selin. Dortrecht, The Netherlands: Springer, 2008, pp. 1978–1983. [Google Scholar]
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Bednarik, R.G. The Origins of Human Modernity. Humanities 2012, 1, 1-53. https://doi.org/10.3390/h1010001
Bednarik RG. The Origins of Human Modernity. Humanities. 2012; 1(1):1-53. https://doi.org/10.3390/h1010001
Chicago/Turabian StyleBednarik, Robert G. 2012. "The Origins of Human Modernity" Humanities 1, no. 1: 1-53. https://doi.org/10.3390/h1010001
APA StyleBednarik, R. G. (2012). The Origins of Human Modernity. Humanities, 1(1), 1-53. https://doi.org/10.3390/h1010001