
Infection with SARS-CoV-2 Is Associated with Elevated Levels of IP-10, MCP-1, and IL-13 in Sepsis Patients
 , , , , , , , and
, , , , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients and Sample Collection
2.2. Data Collection
2.3. Blood Culture
2.4. Coagulation-Related Parameters
2.5. Characterization of Extracellular Vesicles
2.6. Quantification of C5b-9
2.7. Quantification of Cytokines, Chemokines, and Growth Factors
2.8. Statistical Analysis
3. Results
3.1. Patient Characteristics
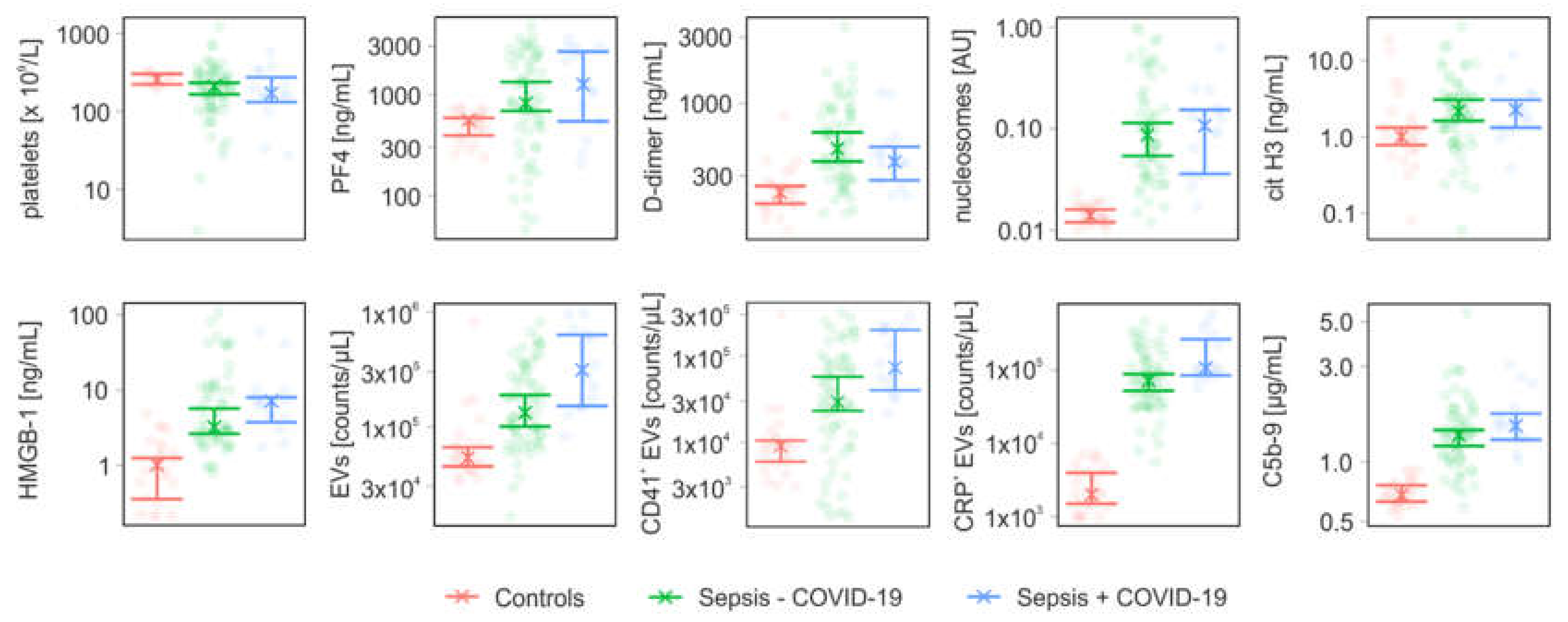
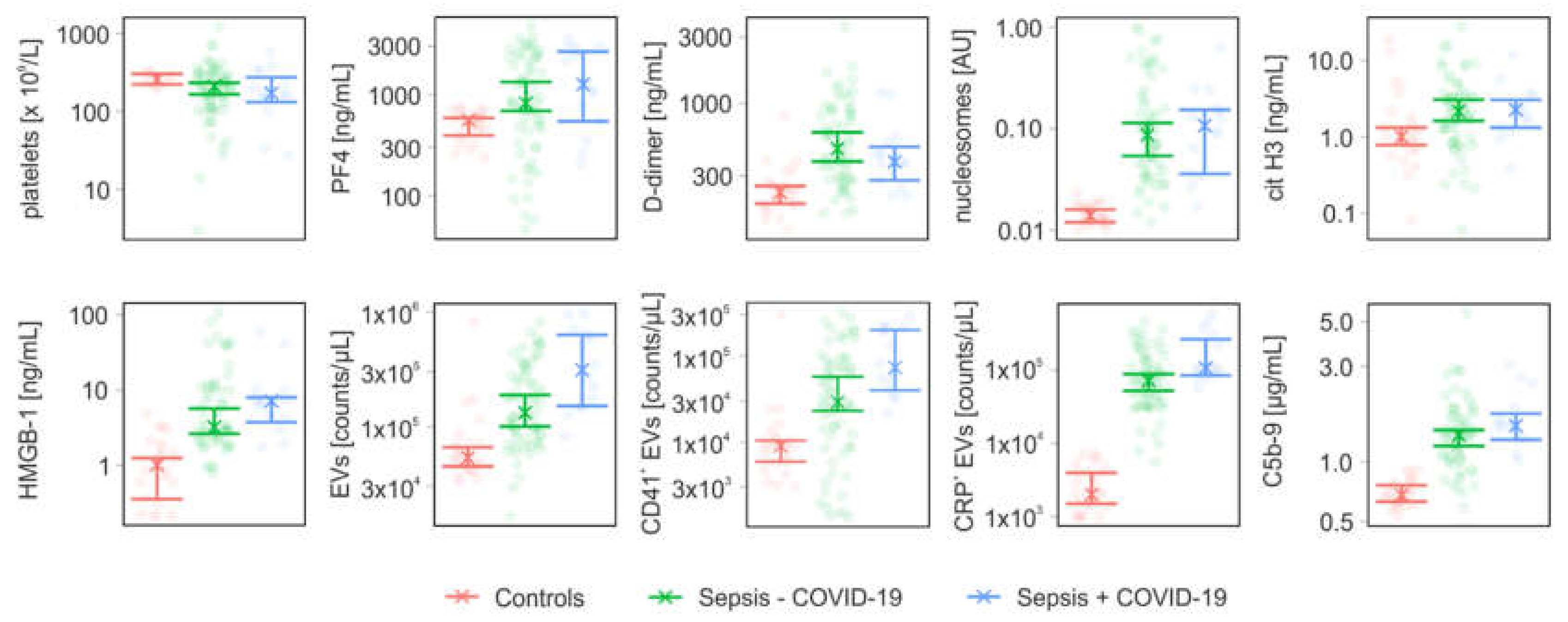
3.2. Coagulation- and Complement-Related Parameters
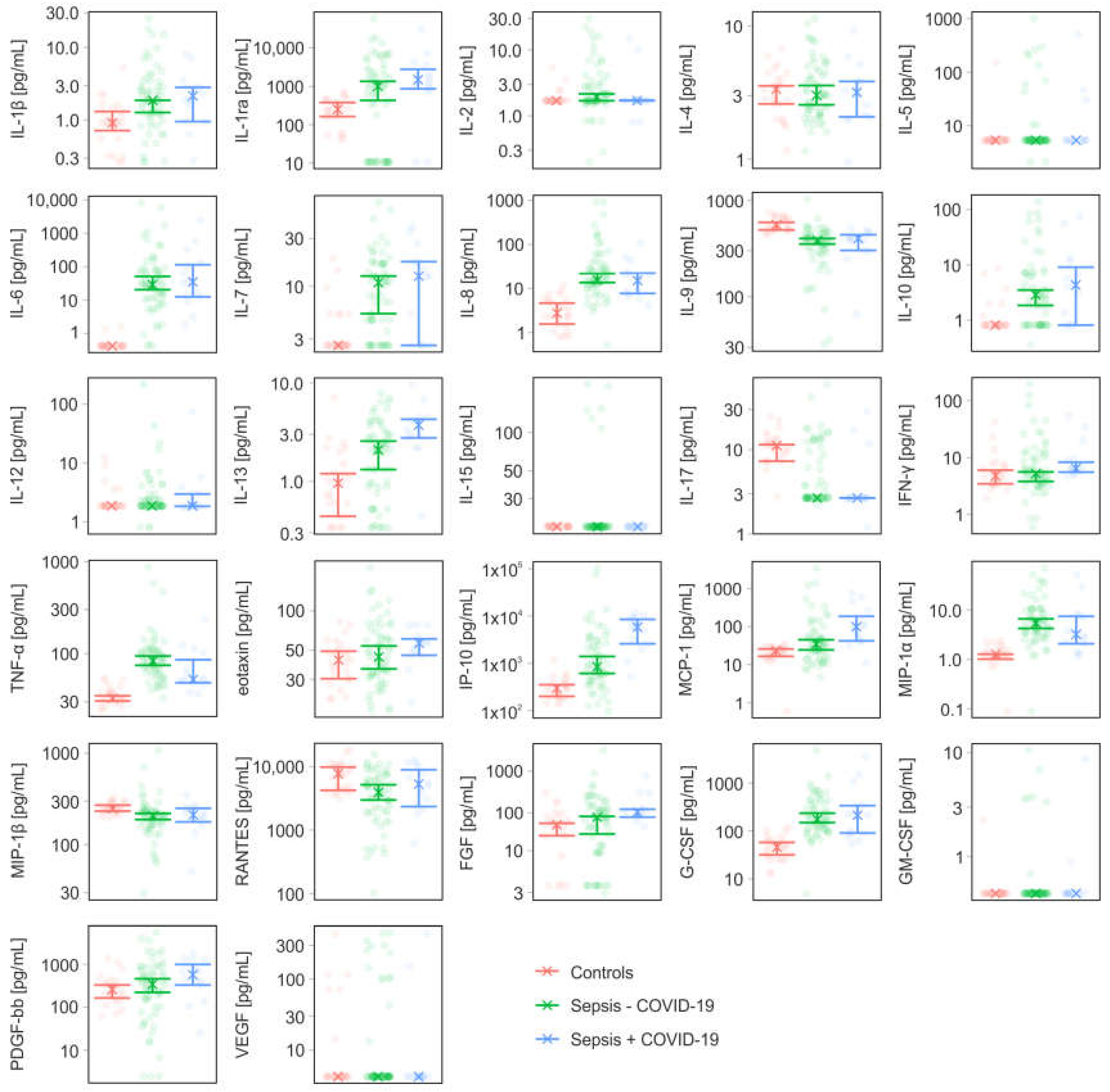
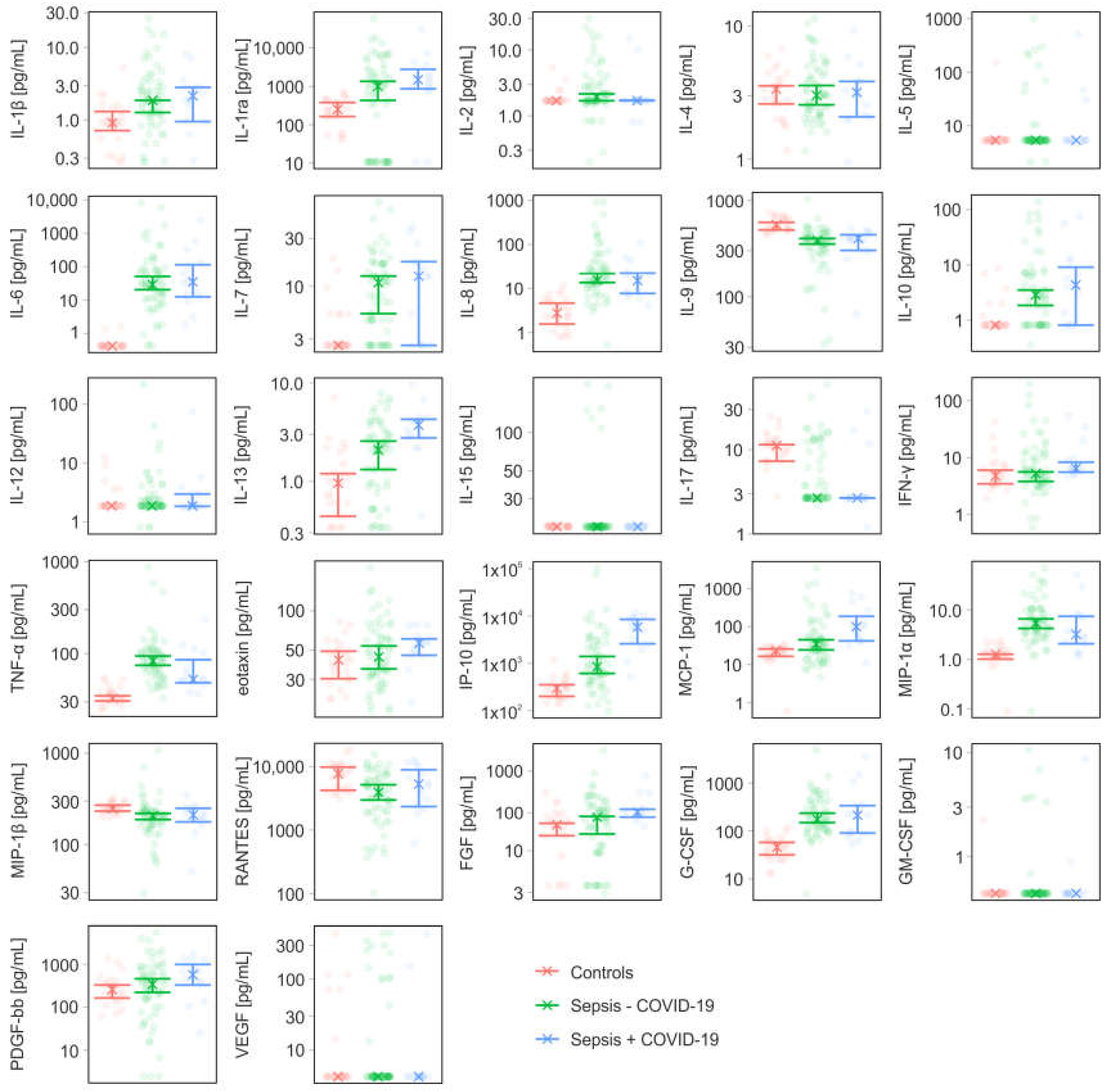
3.3. Cytokines, Chemokines, and Growth Factors
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Singer, M.; Deutschman, C.S.; Seymour, C.W.; Shankar-Hari, M.; Annane, D.; Bauer, M.; Bellomo, R.; Bernard, G.R.; Chiche, J.D.; Coopersmith, C.M.; et al. The Third International Consensus Definitions for Sepsis and Septic Shock (Sepsis-3). JAMA 2016, 315, 801–810. [Google Scholar] [CrossRef] [PubMed]
- Bonaventura, A.; Vecchie, A.; Dagna, L.; Martinod, K.; Dixon, D.L.; Van Tassell, B.W.; Dentali, F.; Montecucco, F.; Massberg, S.; Levi, M.; et al. Endothelial dysfunction and immunothrombosis as key pathogenic mechanisms in COVID-19. Nat. Rev. Immunol. 2021, 21, 319–329. [Google Scholar] [CrossRef] [PubMed]
- Iba, T.; Levy, J.H.; Levi, M.; Connors, J.M.; Thachil, J. Coagulopathy of Coronavirus Disease 2019. Crit. Care Med. 2020, 48, 1358–1364. [Google Scholar] [CrossRef] [PubMed]
- Olwal, C.O.; Nganyewo, N.N.; Tapela, K.; Djomkam Zune, A.L.; Owoicho, O.; Bediako, Y.; Duodu, S. Parallels in Sepsis and COVID-19 Conditions: Implications for Managing Severe COVID-19. Front. Immunol. 2021, 12, 602848. [Google Scholar] [CrossRef]
- Remmelink, M.; De Mendonca, R.; D’Haene, N.; De Clercq, S.; Verocq, C.; Lebrun, L.; Lavis, P.; Racu, M.L.; Trepant, A.L.; Maris, C.; et al. Unspecific post-mortem findings despite multiorgan viral spread in COVID-19 patients. Crit. Care 2020, 24, 495. [Google Scholar] [CrossRef]
- Engelmann, B.; Massberg, S. Thrombosis as an intravascular effector of innate immunity. Nat. Rev. Immunol. 2013, 13, 34–45. [Google Scholar] [CrossRef]
- Ackermann, M.; Verleden, S.E.; Kuehnel, M.; Haverich, A.; Welte, T.; Laenger, F.; Vanstapel, A.; Werlein, C.; Stark, H.; Tzankov, A.; et al. Pulmonary Vascular Endothelialitis, Thrombosis, and Angiogenesis in COVID-19. N. Engl. J. Med. 2020, 383, 120–128. [Google Scholar] [CrossRef]
- Skendros, P.; Mitsios, A.; Chrysanthopoulou, A.; Mastellos, D.C.; Metallidis, S.; Rafailidis, P.; Ntinopoulou, M.; Sertaridou, E.; Tsironidou, V.; Tsigalou, C.; et al. Complement and tissue factor-enriched neutrophil extracellular traps are key drivers in COVID-19 immunothrombosis. J. Clin. Investig. 2020, 130, 6151–6157. [Google Scholar] [CrossRef]
- Middleton, E.A.; He, X.Y.; Denorme, F.; Campbell, R.A.; Ng, D.; Salvatore, S.P.; Mostyka, M.; Baxter-Stoltzfus, A.; Borczuk, A.C.; Loda, M.; et al. Neutrophil extracellular traps contribute to immunothrombosis in COVID-19 acute respiratory distress syndrome. Blood 2020, 136, 1169–1179. [Google Scholar] [CrossRef]
- Gould, T.J.; Vu, T.T.; Swystun, L.L.; Dwivedi, D.J.; Mai, S.H.; Weitz, J.I.; Liaw, P.C. Neutrophil extracellular traps promote thrombin generation through platelet-dependent and platelet-independent mechanisms. Arterioscler. Thromb. Vasc. Biol. 2014, 34, 1977–1984. [Google Scholar] [CrossRef] [Green Version]
- Kambas, K.; Chrysanthopoulou, A.; Vassilopoulos, D.; Apostolidou, E.; Skendros, P.; Girod, A.; Arelaki, S.; Froudarakis, M.; Nakopoulou, L.; Giatromanolaki, A.; et al. Tissue factor expression in neutrophil extracellular traps and neutrophil derived microparticles in antineutrophil cytoplasmic antibody associated vasculitis may promote thromboinflammation and the thrombophilic state associated with the disease. Ann. Rheum. Dis. 2014, 73, 1854–1863. [Google Scholar] [CrossRef] [Green Version]
- Foley, J.H.; Conway, E.M. Cross Talk Pathways Between Coagulation and Inflammation. Circ. Res. 2016, 118, 1392–1408. [Google Scholar] [CrossRef]
- Martinod, K.; Deppermann, C. Immunothrombosis and thromboinflammation in host defense and disease. Platelets 2021, 32, 314–324. [Google Scholar] [CrossRef]
- Ebeyer-Masotta, M.; Eichhorn, T.; Weiss, R.; Laukova, L.; Weber, V. Activated Platelets and Platelet-Derived Extracellular Vesicles Mediate COVID-19-Associated Immunothrombosis. Front. Cell Dev. Biol. 2022, 10, 914891. [Google Scholar] [CrossRef]
- Slungaard, A. Platelet factor 4: A chemokine enigma. Int. J. Biochem. Cell Biol. 2005, 37, 1162–1167. [Google Scholar] [CrossRef]
- Vogel, S.; Bodenstein, R.; Chen, Q.; Feil, S.; Feil, R.; Rheinlaender, J.; Schaffer, T.E.; Bohn, E.; Frick, J.S.; Borst, O.; et al. Platelet-derived HMGB1 is a critical mediator of thrombosis. J. Clin. Investig. 2015, 125, 4638–4654. [Google Scholar] [CrossRef] [Green Version]
- Maugeri, N.; Campana, L.; Gavina, M.; Covino, C.; De Metrio, M.; Panciroli, C.; Maiuri, L.; Maseri, A.; D′Angelo, A.; Bianchi, M.E.; et al. Activated platelets present high mobility group box 1 to neutrophils, inducing autophagy and promoting the extrusion of neutrophil extracellular traps. J. Thromb. Haemost. 2014, 12, 2074–2088. [Google Scholar] [CrossRef]
- Hottz, E.D.; Azevedo-Quintanilha, I.G.; Palhinha, L.; Teixeira, L.; Barreto, E.A.; Pao, C.R.R.; Righy, C.; Franco, S.; Souza, T.M.L.; Kurtz, P.; et al. Platelet activation and platelet-monocyte aggregate formation trigger tissue factor expression in patients with severe COVID-19. Blood 2020, 136, 1330–1341. [Google Scholar] [CrossRef]
- Manne, B.K.; Denorme, F.; Middleton, E.A.; Portier, I.; Rowley, J.W.; Stubben, C.; Petrey, A.C.; Tolley, N.D.; Guo, L.; Cody, M.; et al. Platelet gene expression and function in patients with COVID-19. Blood 2020, 136, 1317–1329. [Google Scholar] [CrossRef]
- McDonald, B.; Urrutia, R.; Yipp, B.G.; Jenne, C.N.; Kubes, P. Intravascular neutrophil extracellular traps capture bacteria from the bloodstream during sepsis. Cell Host Microbe 2012, 12, 324–333. [Google Scholar] [CrossRef] [Green Version]
- Puhm, F.; Flamand, L.; Boilard, E. Platelet extracellular vesicles in COVID-19: Potential markers and makers. J. Leukoc. Biol. 2022, 111, 63–74. [Google Scholar] [CrossRef] [PubMed]
- Kerris, E.W.J.; Hoptay, C.; Calderon, T.; Freishtat, R.J. Platelets and platelet extracellular vesicles in hemostasis and sepsis. J. Investig. Med. 2020, 68, 813–820. [Google Scholar] [CrossRef] [PubMed]
- Tripisciano, C.; Weiss, R.; Eichhorn, T.; Spittler, A.; Heuser, T.; Fischer, M.B.; Weber, V. Different Potential of Extracellular Vesicles to Support Thrombin Generation: Contributions of Phosphatidylserine, Tissue Factor, and Cellular Origin. Sci. Rep. 2017, 7, 6522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Owens, A.P., 3rd; Mackman, N. Microparticles in hemostasis and thrombosis. Circ. Res. 2011, 108, 1284–1297. [Google Scholar] [CrossRef]
- World Health Organization. Clinical Management of COVID-19. Living Guideline. 2021. Available online: https://www.who.int/teams/health-care-readiness/covid-19 (accessed on 5 April 2022).
- EUCAST Disk Diffusion – Manual v 2.0 to v 7.0. Available online: https://www.eucast.org/ast_of_bacteria/previous_versions_of_documents/ (accessed on 21 July 2021).
- George, S.K.; Laukova, L.; Weiss, R.; Semak, V.; Fendl, B.; Weiss, V.U.; Steinberger, S.; Allmaier, G.; Tripisciano, C.; Weber, V. Comparative Analysis of Platelet-Derived Extracellular Vesicles Using Flow Cytometry and Nanoparticle Tracking Analysis. Int. J. Mol. Sci. 2021, 22, 3839. [Google Scholar] [CrossRef]
- Ebeyer-Masotta, M.; Eichhorn, T.; Weiss, R.; Semak, V.; Laukova, L.; Fischer, M.B.; Weber, V. Heparin-Functionalized Adsorbents Eliminate Central Effectors of Immunothrombosis, including Platelet Factor 4, High-Mobility Group Box 1 Protein and Histones. Int. J. Mol. Sci. 2022, 23, 1823. [Google Scholar] [CrossRef]
- Huber, S.; Massri, M.; Grasse, M.; Fleischer, V.; Kellnerova, S.; Harpf, V.; Knabl, L.; Knabl, L., Sr.; Heiner, T.; Kummann, M.; et al. Systemic Inflammation and Complement Activation Parameters Predict Clinical Outcome of Severe SARS-CoV-2 Infections. Viruses 2021, 13, 2376. [Google Scholar] [CrossRef]
- Wurzner, R.; Nitze, R.; Gotze, O. C7*9, a new frequent C7 allele detected by an allotype-specific monoclonal antibody. Complement. Inflamm. 1990, 7, 290–297. [Google Scholar] [CrossRef]
- Canty, A.R.; Ripley, B.D. boot: Bootstrap R (S-Plus) Functions. R package version 1.3-28. Available online: https://cran.r-project.org/web/packages/boot/index.html (accessed on 30 November 2022).
- Davison, A.C.; Hinkley, D.V. Bootstrap Methods and Their Applications; Cambridge University Press: Cambridge, UK, 1997. [Google Scholar]
- Fendl, B.; Weiss, R.; Eichhorn, T.; Linsberger, I.; Afonyushkin, T.; Puhm, F.; Binder, C.J.; Fischer, M.B.; Weber, V. Extracellular vesicles are associated with C-reactive protein in sepsis. Sci. Rep. 2021, 11, 6996. [Google Scholar] [CrossRef]
- Langford, B.J.; So, M.; Raybardhan, S.; Leung, V.; Westwood, D.; MacFadden, D.R.; Soucy, J.R.; Daneman, N. Bacterial co-infection and secondary infection in patients with COVID-19: A living rapid review and meta-analysis. Clin. Microbiol. Infect. 2020, 26, 1622–1629. [Google Scholar] [CrossRef]
- Musuuza, J.S.; Watson, L.; Parmasad, V.; Putman-Buehler, N.; Christensen, L.; Safdar, N. Prevalence and outcomes of co-infection and superinfection with SARS-CoV-2 and other pathogens: A systematic review and meta-analysis. PLoS ONE 2021, 16, e0251170. [Google Scholar] [CrossRef]
- Clerbaux, L.A.; Fillipovska, J.; Muñoz, A.; Petrillo, M.; Coecke, S.; Amorim, M.J.; Grenga, L. Mechanisms Leading to Gut Dysbiosis in COVID-19: Current Evidence and Uncertainties Based on Adverse Outcome Pathways. J. Clin. Med. 2022, 11, 5400. [Google Scholar] [CrossRef]
- Gang, J.; Wang, H.; Xue, X.; Zhang, S. Microbiota and COVID-19: Long-term and complex influencing factors. Front. Microbiol. 2022, 13, 963488. [Google Scholar] [CrossRef]
- Petrey, A.C.; Qeadan, F.; Middleton, E.A.; Pinchuk, I.V.; Campbell, R.A.; Beswick, E.J. Cytokine release syndrome in COVID-19: Innate immune, vascular, and platelet pathogenic factors differ in severity of disease and sex. J. Leukoc. Biol. 2021, 109, 55–66. [Google Scholar] [CrossRef]
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X.; et al. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef] [Green Version]
- Del Valle, D.M.; Kim-Schulze, S.; Huang, H.H.; Beckmann, N.D.; Nirenberg, S.; Wang, B.; Lavin, Y.; Swartz, T.H.; Madduri, D.; Stock, A.; et al. An inflammatory cytokine signature predicts COVID-19 severity and survival. Nat. Med. 2020, 26, 1636–1643. [Google Scholar] [CrossRef]
- Jarczak, D.; Nierhaus, A. Cytokine Storm-Definition, Causes, and Implications. Int. J. Mol. Sci. 2022, 23, 11740. [Google Scholar] [CrossRef]
- Osuchowski, M.F.; Winkler, M.S.; Skirecki, T.; Cajander, S.; Shankar-Hari, M.; Lachmann, G.; Monneret, G.; Venet, F.; Bauer, M.; Brunkhorst, F.M.; et al. The COVID-19 puzzle: Deciphering pathophysiology and phenotypes of a new disease entity. Lancet Respir. Med. 2021, 9, 622–642. [Google Scholar] [CrossRef]
- Chi, Y.; Ge, Y.; Wu, B.; Zhang, W.; Wu, T.; Wen, T.; Liu, J.; Guo, X.; Huang, C.; Jiao, Y.; et al. Serum Cytokine and Chemokine Profile in Relation to the Severity of Coronavirus Disease 2019 in China. J. Infect. Dis. 2020, 222, 746–754. [Google Scholar] [CrossRef]
- Monneret, G.; Benlyamani, I.; Gossez, M.; Bermejo-Martin, J.F.; Martin-Fernandez, M.; Sesques, P.; Wallet, F.; Venet, F. COVID-19: What type of cytokine storm are we dealing with? J. Med. Virol. 2021, 93, 197–198. [Google Scholar] [CrossRef]
- Kox, M.; Waalders, N.J.B.; Kooistra, E.J.; Gerretsen, J.; Pickkers, P. Cytokine Levels in Critically Ill Patients With COVID-19 and Other Conditions. JAMA 2020, 324, 1565–1567. [Google Scholar] [CrossRef] [PubMed]
- Leisman, D.E.; Ronner, L.; Pinotti, R.; Taylor, M.D.; Sinha, P.; Calfee, C.S.; Hirayama, A.V.; Mastroiani, F.; Turtle, C.J.; Harhay, M.O.; et al. Cytokine elevation in severe and critical COVID-19: A rapid systematic review, meta-analysis, and comparison with other inflammatory syndromes. Lancet Respir. Med. 2020, 8, 1233–1244. [Google Scholar] [CrossRef] [PubMed]
- Stolarski, A.E.; Kim, J.; Zhang, Q.; Remick, D.G. Cytokine Drizzle-The Rationale for Abandoning “Cytokine Storm”. Shock 2021, 56, 667–672. [Google Scholar] [CrossRef] [PubMed]
- Wilson, J.G.; Simpson, L.J.; Ferreira, A.M.; Rustagi, A.; Roque, J.; Asuni, A.; Ranganath, T.; Grant, P.M.; Subramanian, A.; Rosenberg-Hasson, Y.; et al. Cytokine profile in plasma of severe COVID-19 does not differ from ARDS and sepsis. JCI Insight 2020, 5, e140289. [Google Scholar] [CrossRef]
- Dufour, J.H.; Dziejman, M.; Liu, M.T.; Leung, J.H.; Lane, T.E.; Luster, A.D. IFN-gamma-inducible protein 10 (IP-10; CXCL10)-deficient mice reveal a role for IP-10 in effector T cell generation and trafficking. J. Immunol. 2002, 168, 3195–3204. [Google Scholar] [CrossRef] [Green Version]
- Punyadeera, C.; Schneider, E.M.; Schaffer, D.; Hsu, H.Y.; Joos, T.O.; Kriebel, F.; Weiss, M.; Verhaegh, W.F. A biomarker panel to discriminate between systemic inflammatory response syndrome and sepsis and sepsis severity. J. Emerg. Trauma Shock 2010, 3, 26–35. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Zhang, C.; Huang, F.; Yang, Y.; Wang, F.; Yuan, J.; Zhang, Z.; Qin, Y.; Li, X.; Zhao, D.; et al. Elevated plasma levels of selective cytokines in COVID-19 patients reflect viral load and lung injury. Natl. Sci. Rev. 2020, 7, 1003–1011. [Google Scholar] [CrossRef] [Green Version]
- Tegethoff, S.A.; Danziger, G.; Kuhn, D.; Kimmer, C.; Adams, T.; Heintz, L.; Metz, C.; Reifenrath, K.; Angresius, R.; Mang, S.; et al. TNF-related apoptosis-inducing ligand, interferon gamma-induced protein 10, and C-reactive protein in predicting the progression of SARS-CoV-2 infection: A prospective cohort study. Int. J. Infect. Dis. 2022, 122, 178–187. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, J.; Liu, C.; Su, L.; Zhang, D.; Fan, J.; Yang, Y.; Xiao, M.; Xie, J.; Xu, Y.; et al. IP-10 and MCP-1 as biomarkers associated with disease severity of COVID-19. Mol. Med. 2020, 26, 97. [Google Scholar] [CrossRef]
- Donlan, A.N.; Sutherland, T.E.; Marie, C.; Preissner, S.; Bradley, B.T.; Carpenter, R.M.; Sturek, J.M.; Ma, J.Z.; Moreau, G.B.; Donowitz, J.R.; et al. IL-13 is a driver of COVID-19 severity. JCI Insight 2021, 6, e150107. [Google Scholar] [CrossRef]
- Marone, G.; Granata, F.; Pucino, V.; Pecoraro, A.; Heffler, E.; Loffredo, S.; Scadding, G.W.; Varricchi, G. The Intriguing Role of Interleukin 13 in the Pathophysiology of Asthma. Front. Pharmacol. 2019, 10, 1387. [Google Scholar] [CrossRef] [Green Version]
- Loftus, T.J.; Ungaro, R.; Dirain, M.; Efron, P.A.; Mazer, M.B.; Remy, K.E.; Hotchkiss, R.S.; Zhong, L.; Bacher, R.; Starostik, P.; et al. Overlapping but Disparate Inflammatory and Immunosuppressive Responses to SARS-CoV-2 and Bacterial Sepsis: An Immunological Time Course Analysis. Front. Immunol. 2021, 12, 792448. [Google Scholar] [CrossRef]
- Maharaj, S.; Chang, S. Anti-PF4/heparin antibodies are increased in hospitalized patients with bacterial sepsis. Thromb. Res. 2018, 171, 111–113. [Google Scholar] [CrossRef]
- Comer, S.P.; Cullivan, S.; Szklanna, P.B.; Weiss, L.; Cullen, S.; Kelliher, S.; Smolenski, A.; Murphy, C.; Altaie, H.; Curran, J.; et al. COVID-19 induces a hyperactive phenotype in circulating platelets. PLoS Biol. 2021, 19, e3001109. [Google Scholar] [CrossRef]
- Wegrzyn, G.; Walborn, A.; Rondina, M.; Fareed, J.; Hoppensteadt, D. Biomarkers of Platelet Activation and Their Prognostic Value in Patients with Sepsis-Associated Disseminated Intravascular Coagulopathy. Clin. Appl. Thromb. Hemost. 2021, 27, 1076029620943300. [Google Scholar] [CrossRef]
- Traby, L.; Kollars, M.; Kussmann, M.; Karer, M.; Sinkovec, H.; Lobmeyr, E.; Hermann, A.; Staudinger, T.; Schellongowski, P.; Rossler, B.; et al. Extracellular Vesicles and Citrullinated Histone H3 in Coronavirus Disease 2019 Patients. Thromb. Haemost. 2022, 122, 113–122. [Google Scholar] [CrossRef]
- Bouck, E.G.; Denorme, F.; Holle, L.A.; Middelton, E.A.; Blair, A.M.; de Laat, B.; Schiffman, J.D.; Yost, C.C.; Rondina, M.T.; Wolberg, A.S.; et al. COVID-19 and Sepsis Are Associated With Different Abnormalities in Plasma Procoagulant and Fibrinolytic Activity. Arterioscler. Thromb. Vasc. Biol. 2021, 41, 401–414. [Google Scholar] [CrossRef]
- Leppkes, M.; Knopf, J.; Naschberger, E.; Lindemann, A.; Singh, J.; Herrmann, I.; Sturzl, M.; Staats, L.; Mahajan, A.; Schauer, C.; et al. Vascular occlusion by neutrophil extracellular traps in COVID-19. EBioMedicine 2020, 58, 102925. [Google Scholar] [CrossRef]
- Ng, H.; Havervall, S.; Rosell, A.; Aguilera, K.; Parv, K.; von Meijenfeldt, F.A.; Lisman, T.; Mackman, N.; Thalin, C.; Phillipson, M. Circulating Markers of Neutrophil Extracellular Traps Are of Prognostic Value in Patients With COVID-19. Arterioscler. Thromb. Vasc. Biol. 2021, 41, 988–994. [Google Scholar] [CrossRef]
- Yokoyama, Y.; Ito, T.; Yasuda, T.; Furubeppu, H.; Kamikokuryo, C.; Yamada, S.; Maruyama, I.; Kakihana, Y. Circulating histone H3 levels in septic patients are associated with coagulopathy, multiple organ failure, and death: A single-center observational study. Thromb. J. 2019, 17, 1. [Google Scholar] [CrossRef]
- Maruchi, Y.; Tsuda, M.; Mori, H.; Takenaka, N.; Gocho, T.; Huq, M.A.; Takeyama, N. Plasma myeloperoxidase-conjugated DNA level predicts outcomes and organ dysfunction in patients with septic shock. Crit. Care 2018, 22, 176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reid, V.L.; Webster, N.R. Role of microparticles in sepsis. Br. J. Anaesth. 2012, 109, 503–513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dix, C.; Zeller, J.; Stevens, H.; Eisenhardt, S.U.; Shing, K.; Nero, T.L.; Morton, C.J.; Parker, M.W.; Peter, K.; McFadyen, J.D. C-reactive protein, immunothrombosis and venous thromboembolism. Front. Immunol. 2022, 13, 1002652. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
| Characteristics | Controls (n = 25) | Sepsis − COVID-19 (n = 64) | Sepsis + COVID-19 (n = 14) |
|---|---|---|---|
| Age, years | 34.9 ± 11.7 | 63.6 ± 18.7 | 68.8 ± 17.3 |
| Male | 10 (40%) | 41 (64%) | 11 (79%) |
| Female | 15 (60%) | 23 (36%) | 3 (21%) |
| ICU admission | n.a. | 14 (22%) | 5 (36%) |
| Mortality | n.a. | 7 (11%) | 6 (43%) |
| Positive blood culture | n.a. | 8 (13%) | 1 (7%) |
| CRP [mg/dL] | n.a. | 9.78 (6.31; 12.08) | 11.31 (5.67; 15.11) |
| PCT [µg/L] | n.a. | 0.54 (0.31; 3.18) | 0.56 (0.16; 1.14) |
| Red blood cells [× 1012/L] | n.a. | 3.42 (3.16; 3.57) | 3.45 (3.11; 3.97) |
| White blood cells [× 109/L] | n.a. | 9.05 (7.55; 11.2) | 8.35 (6.15; 13.65) |
| Monocytes [× 109/L] | n.a. | 0.69 (0.48; 0.83) | 0.38 (0.22; 0.64) |
| Neutrophils [× 109/L] | n.a. | 9.4 (4.85; 11) | 9.85 (4.85; 13.6) |
| Lymphocytes [× 109/L] | n.a. | 1.03 (0.69; 1.16) | 0.98 (0.36; 1.62) |
| Neutrophil-to-lymphocyte ratio (NLR) | n.a. | 7.16 (5.53; 11.25) | 16.32 (1.74; 36.36) |
| Platelets [× 109/L] | 257 (222; 303) a | 212 (166; 232) | 170 (132; 274) |
| Comorbidities, n (%) | |||
| Infectious/parasitic diseases | n.a. | 19 (30) | 4 (29) |
| Neoplasms | n.a. | 4 (6) | 5 (36) |
| Blood/blood forming organ diseases | n.a. | 9 (14) | 3 (21) |
| Immune system diseases | n.a. | 3 (5) | 0 (0) |
| Endocrine/nutritional/metabolic diseases | n.a. | 24 (38) | 6 (43) |
| Mental/behavioral/neurodevelopmental disorders | n.a. | 9 (14) | 2 (14) |
| Nervous system diseases | n.a. | 3 (5) | 4 (29) |
| Visual system diseases | n.a. | 1 (2) | 1 (7) |
| Circulatory system diseases | n.a. | 34 (53) | 10 (71) |
| Respiratory system diseases | n.a. | 25 (39) | 13 (93) |
| Digestive system diseases | n.a. | 11 (17) | 5 (36) |
| Skin diseases | n.a. | 4 (6) | 1 (7) |
| Musculoskeletal/connective tissue diseases | n.a. | 5 (8) | 0 (0) |
| Genitourinary system diseases | n.a. | 29 (45) | 6 (43) |
| Other symptoms, signs, or clinical findings | n.a. | 4 (6) | 6 (43) |
| Injury/poisoning/consequences of external causes | n.a. | 5 (8) | 1 (7) |
| Factors influencing health status/contact with health services | n.a. | 10 (16) | 2 (14) |
| Parameter | Controls (n = 25) | Sepsis − COVID-19 (n = 64) | Sepsis + COVID-19 (n = 14) | |
|---|---|---|---|---|
| PF4 | [ng/mL] | 558 (394; 587) | 820 (688; 1330) | 1260 (544; 2657) |
| D-dimer | [ng/mL] | 227 (189; 253) | 471 (380; 617) | 378 (278; 485) |
| nucleosomes | [AU] | 0.01 (0.01; 0.02) | 0.09 (0.05; 0.11) | 0.11 (0.04; 0.15) |
| cit H3 | [ng/mL] | 1 (0.78; 1.32) | 2.14 (1.62; 3.06) | 2.27 (1.32; 3.04) |
| HMGB-1 | [ng/mL] | 1.01 (0.35; 1.25) | 3.22 (2.63; 5.69) | 7.16 (3.74; 8.01) |
| C5b-9 | [µg/mL] | 0.68 (0.63; 0.76) | 1.36 (1.2; 1.44) | 1.52 (1.29; 1.74) |
| EVs | [counts/µL] | 53,500 (45,000; 66,000) | 132,250 (100,250; 188,500) | 311,000 (151,000; 630,000) |
| CD41+ EVs | [counts/µL] | 9000 (5940; 10,440) | 28,945 (23,090; 57,195) | 73,085 (39,790; 198,860) |
| CD41+ EVs | [% of total EVs] | 15 (11; 18) | 27 (18; 34) | 31 (23; 35.5) |
| CRP+ EVs | [counts/µL] | 2000 (1500; 4000) | 71,660 (52,500; 88,500) | 106750 (85,000; 263,500) |
| CRP+ EVs | [% of total EVs] | 4 (2; 4) | 55 (46; 61) | 62 (40; 67.5) |
| IL-1β | [pg/mL] | 0.92 (0.72; 1.32) | 1.88 (1.28; 1.88) | 2.16 (0.96; 2.84) |
| IL-1ra | [pg/mL] | 251 (164; 379) | 1010 (431; 1353) | 1495 (866; 2805) |
| IL-2 | [pg/mL] | 1.7 (n.a.; n.a.) | 1.93 (1.7; 2.16) | 1.7 (1.7; 1.73) |
| IL-4 | [pg/mL] | 3.36 (2.6; 3.56) | 3.02 (2.56; 3.58) | 3.18 (2.08; 3.84) |
| IL-5 | [pg/mL] | 5.34 (n.a.; n.a.) | 5.34 (n.a.; n.a.) | 5.34 (n.a.; n.a.) |
| IL-6 | [pg/mL] | 0.42 (n.a.; n.a.) | 28.6 (20.12; 50.04) | 34.34 (12.32; 115) |
| IL-7 | [pg/mL] | 2.6 (n.a.; n.a.) | 10.86 (5.36; 12.64) | 12.64 (2.6; 17.68) |
| IL-8 | [pg/mL] | 2.76 (1.56; 4.64) | 15.44 (13.56; 21.84) | 15 (7.68; 22.3) |
| IL-9 | [pg/mL] | 557 (493; 591) | 383 (353; 402) | 402 (303; 440) |
| IL-10 | [pg/mL] | 0.81 (n.a.; n.a.) | 2.84 (1.84; 3.48) | 4.28 (0.81; 9.12) |
| IL-12 | [pg/mL] | 1.87 (n.a.; n.a.) | 1.87 (n.a.; n.a.) | 1.87 (1.83; 2.96) |
| IL-13 | [pg/mL] | 0.96 (0.44; 1.2) | 2.08 (1.32; 2.56) | 3.72 (2.76; 4.26) |
| IL-15 | [pg/mL] | 18.02 (n.a.; n.a.) | 18.02 (n.a.; n.a.) | 18.02 (n.a.; n.a.) |
| IL-17 | [pg/mL] | 11.24 (7.32; 11.48) | 2.67 (n.a.; n.a.) | 2.67 (2.67; 2.67) |
| IFN-γ | [pg/mL] | 4.8 (3.48; 6.04) | 5.28 (3.84; 5.62) | 6.48 (5.58; 8.34) |
| TNF-α | [pg/mL] | 32.72 (30.56; 34.84) | 82.92 (74.44; 93.96) | 52.38 (48.24; 85.52) |
| eotaxin | [pg/mL] | 42.12 (30.32; 49.12) | 44.48 (36; 53.92) | 56.58 (45.76; 61.08) |
| IP-10 | [pg/mL] | 297 (202; 354) | 846 (614; 1400) | 5806 (2593; 8493) |
| MCP-1 | [pg/mL] | 23.2 (16.4; 25.52) | 34.22 (24.12; 44.64) | 98.24 (41.52; 184) |
| MIP-1α | [pg/mL] | 1.28 (1.01; 1.28) | 5.42 (4.24; 6.6) | 3.2 (2.08; 7.42) |
| MIP-1β | [pg/mL] | 248 (233; 271) | 207 (188; 220) | 212 (177; 251) |
| RANTES | [pg/mL] | 7742 (4249; 9806) | 3940 (2989; 5182) | 5319 (2357; 8911) |
| FGF | [pg/mL] | 21.2 (15.4; 22.04) | 26.24 (16.16; 27.12) | 30.66 (26.48; 33.28) |
| G-CSF | [pg/mL] | 45.68 (31.6; 57.56) | 174 (148; 237) | 213 (90; 340) |
| GM-CSF | [pg/mL] | 0.44 (n.a.; n.a.) | 0.44 (n.a.; n.a.) | 0.44 (n.a.; n.a.) |
| PDGF-bb | [pg/mL] | 258 (164; 330) | 343 (222; 461) | 573 (329; 996) |
| VEGF | [pg/mL] | 4.22 (n.a.; n.a.) | 4.22 (n.a.; n.a.) | 4.22 (n.a.; n.a.) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eichhorn, T.; Huber, S.; Weiss, R.; Ebeyer-Masotta, M.; Lauková, L.; Emprechtinger, R.; Bellmann-Weiler, R.; Lorenz, I.; Martini, J.; Pirklbauer, M.; et al. Infection with SARS-CoV-2 Is Associated with Elevated Levels of IP-10, MCP-1, and IL-13 in Sepsis Patients. Diagnostics 2023, 13, 1069. https://doi.org/10.3390/diagnostics13061069
Eichhorn T, Huber S, Weiss R, Ebeyer-Masotta M, Lauková L, Emprechtinger R, Bellmann-Weiler R, Lorenz I, Martini J, Pirklbauer M, et al. Infection with SARS-CoV-2 Is Associated with Elevated Levels of IP-10, MCP-1, and IL-13 in Sepsis Patients. Diagnostics. 2023; 13(6):1069. https://doi.org/10.3390/diagnostics13061069
Chicago/Turabian StyleEichhorn, Tanja, Silke Huber, René Weiss, Marie Ebeyer-Masotta, Lucia Lauková, Robert Emprechtinger, Rosa Bellmann-Weiler, Ingo Lorenz, Judith Martini, Markus Pirklbauer, and et al. 2023. "Infection with SARS-CoV-2 Is Associated with Elevated Levels of IP-10, MCP-1, and IL-13 in Sepsis Patients" Diagnostics 13, no. 6: 1069. https://doi.org/10.3390/diagnostics13061069