Pavilion Lake Microbialites: Morphological, Molecular and Biochemical Evidence for a Cold-Water Transition to Colonial Aggregates



Abstract
:
1. Introduction
2. Observations
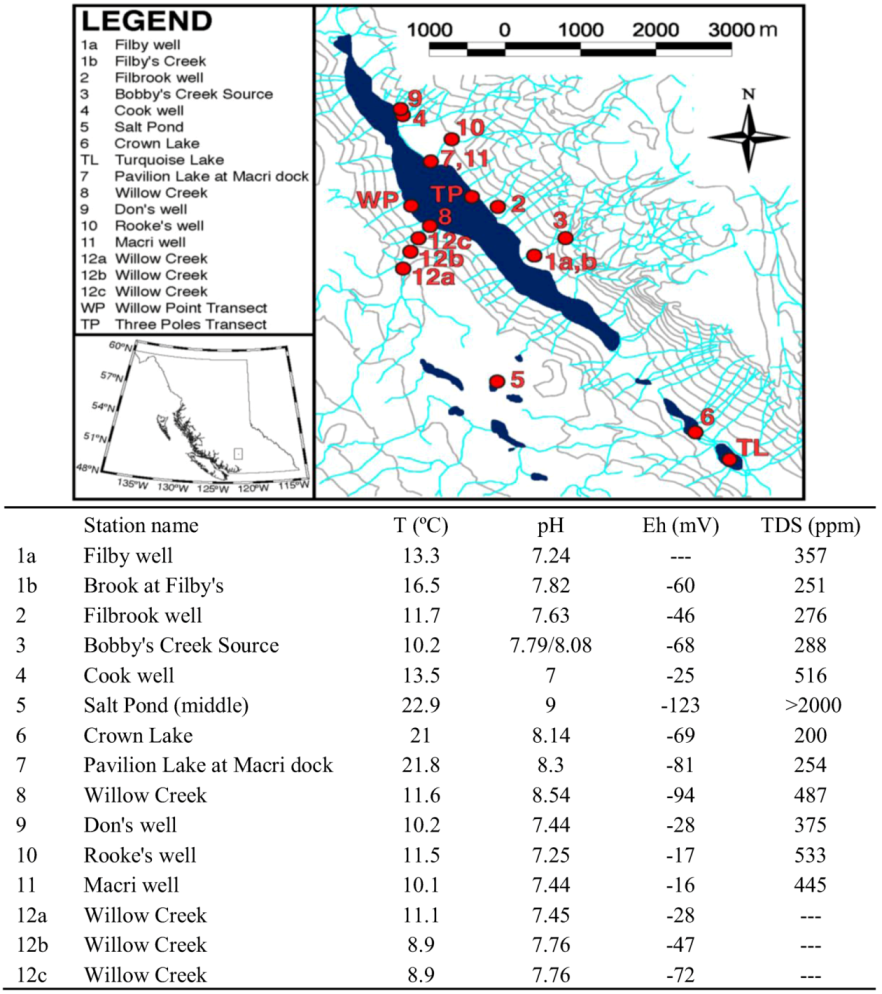
2.1. Morphology


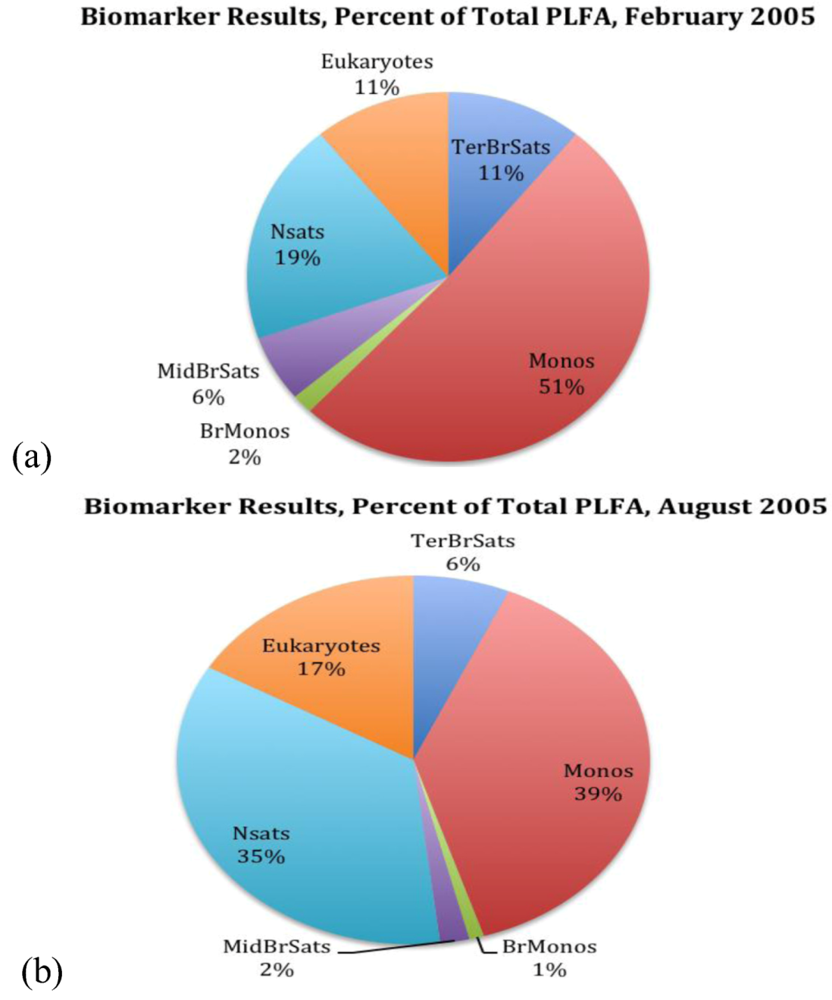
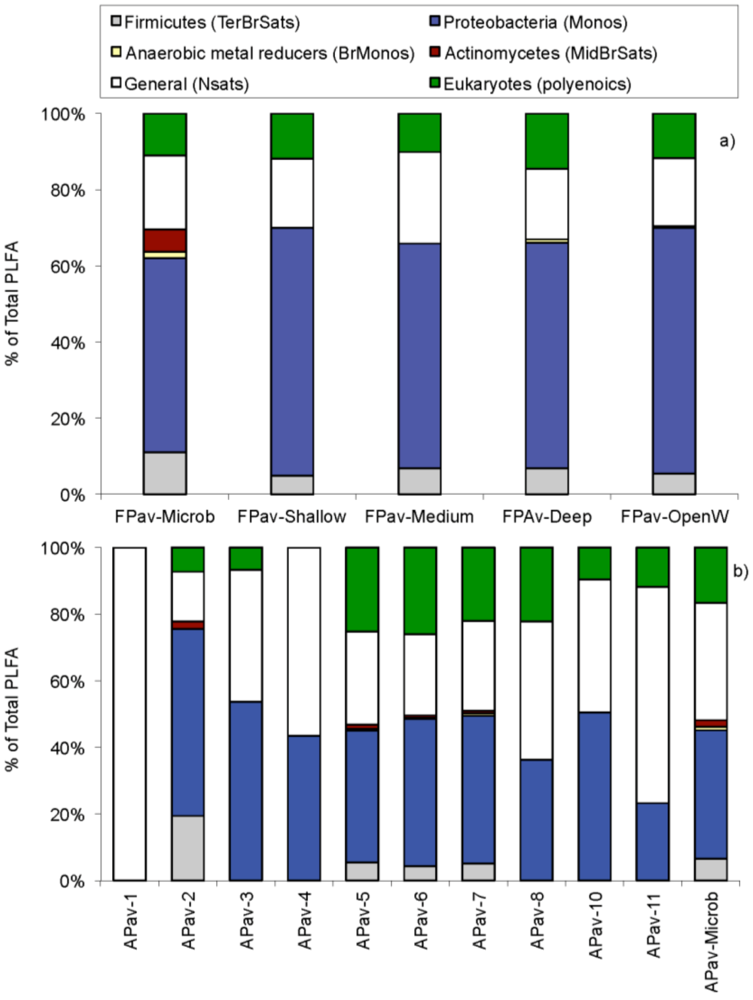
2.2. Biochemistry

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
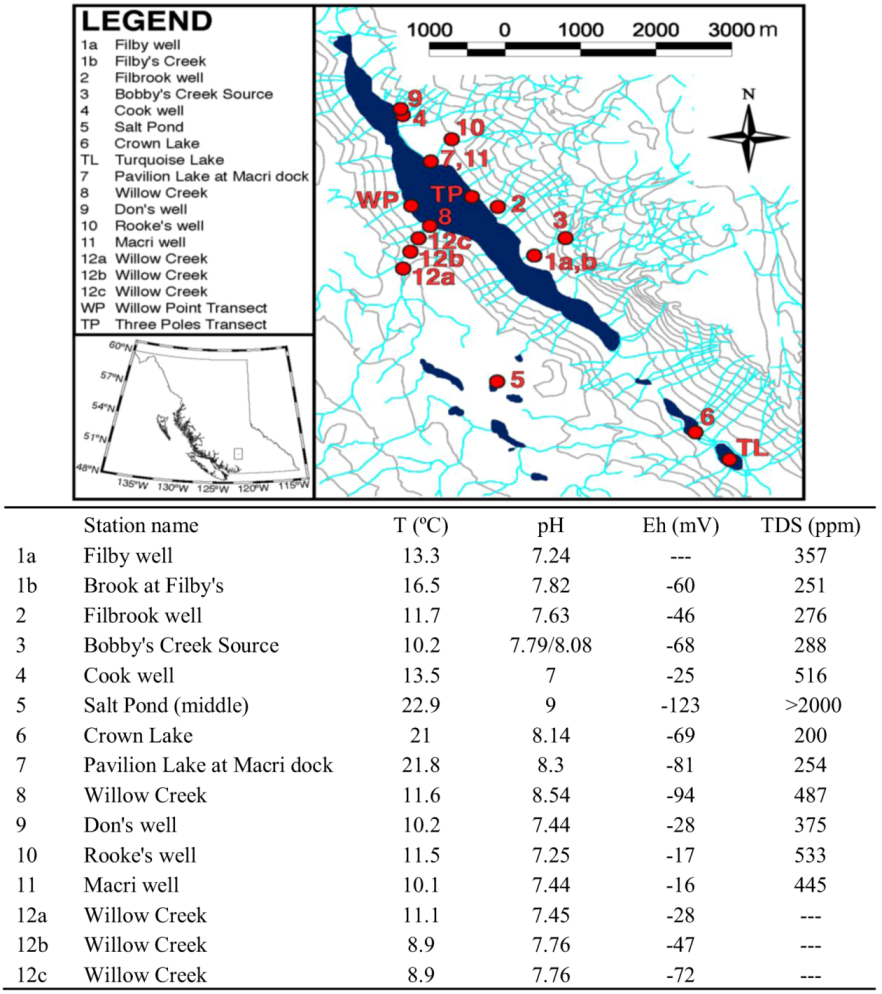
{kind=link}
| Biomarkers | ECL1 | February’05 | August’05 |
|---|---|---|---|
| Terminally Branched Saturates (TerBrSats) | |||
| i15:0 | 14.64 | 2.6 | 2.1 |
| a17:0 | 16.73 | 2.5 | 1 |
| Total | 11 | 6.6 | |
| Monoenoics (Monos) | |||
| 16:1w7c | 15.77 | 17.9 | 10.4 |
| 18:1w7c | 17.79 | 13 | 15.9 |
| Total | 51.1 | 38.6 | |
| Branched Monoenoics (BrMonos) | |||
| i17:1w7c | 16.38 | 1.1 | 0.4 |
| br19:1a | 18.05 | 0.6 | 0.6 |
| Total | 1.7 | 1 | |
| Mid-Chain Branched Saturates (MidBrSats) | |||
| 10me16:0 | 16.45 | 3.9 | 1.4 |
| 12me16:0 | 16.48 | 1.6 | 0.5 |
| Total: | 5.8 | 2 | |
| Normal Saturates (NSats) | |||
| 14:00 | 14 | 0.7 | 6.2 |
| 16:00 | 16 | 15.4 | 26.8 |
| Total: | 19.4 | 35.2 | |
| Eukaryotes | |||
| 18:2w6 | 17.64 | 3.6 | 7.3 |
| 18:3w3 | 17.68 | 2.6 | 6.3 |
| 20:5w3 | 19.32 | 3.3 | 1.8 |
| Total | 11 | 16.6 | |
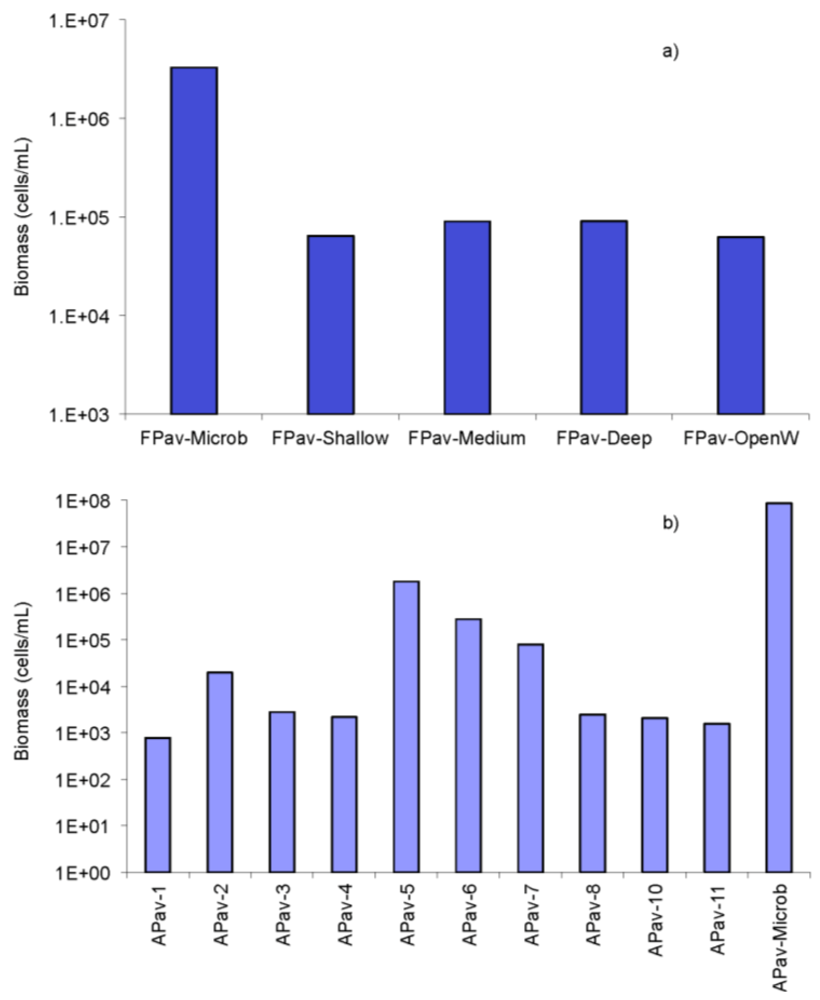
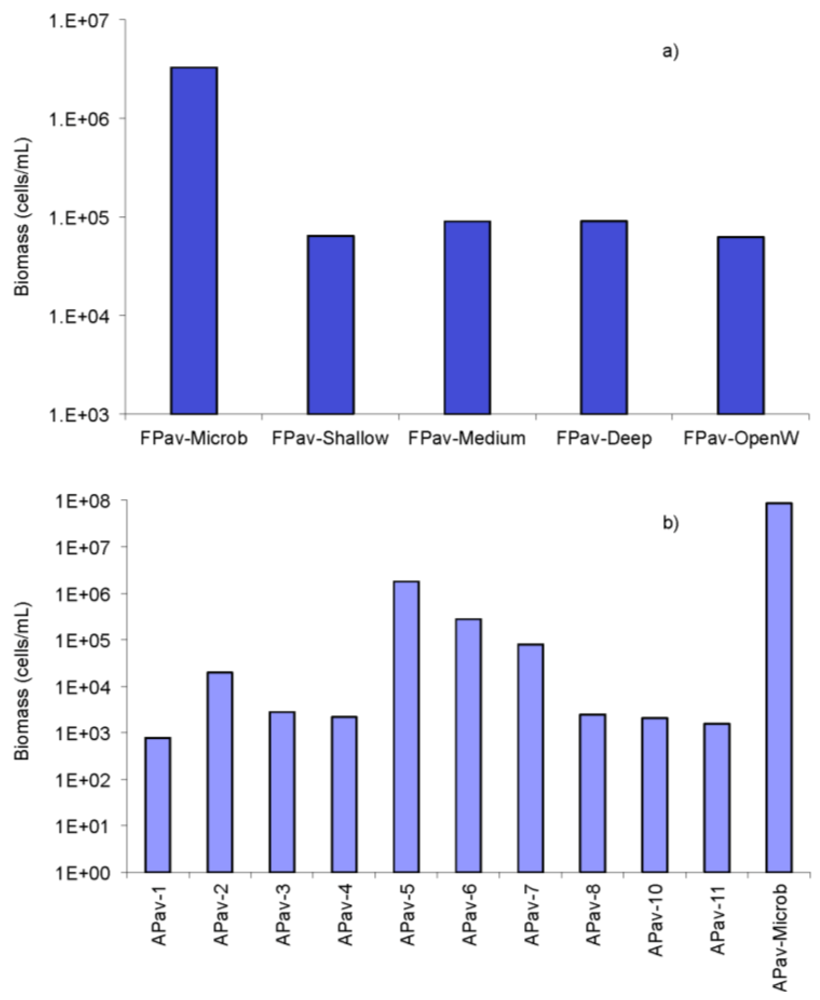
2.3. Biomass
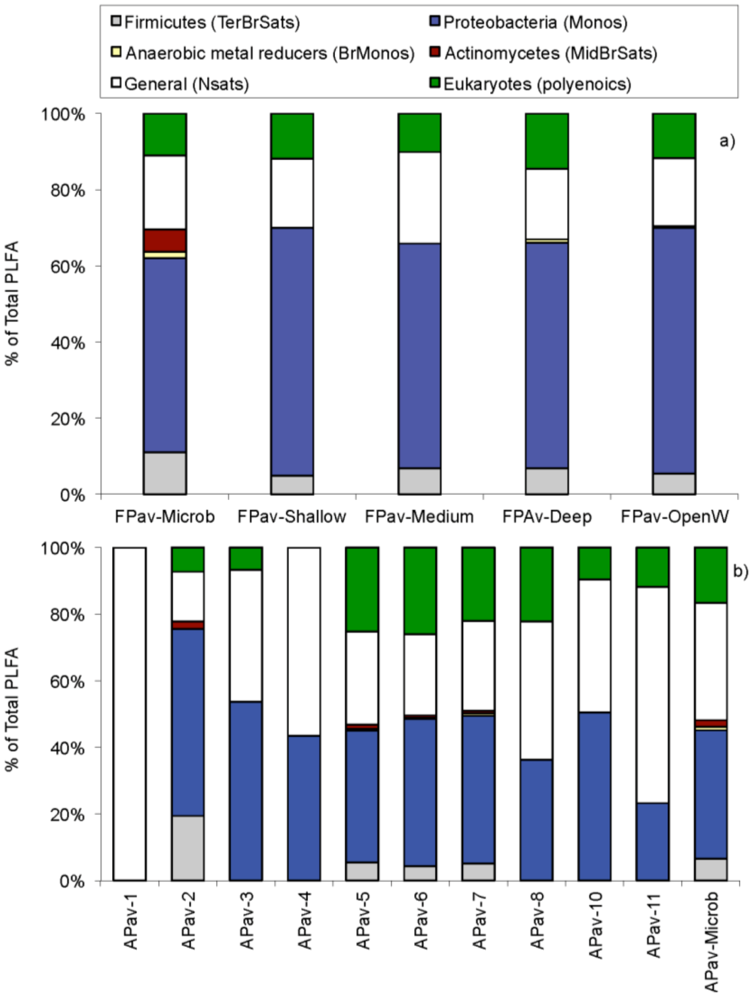
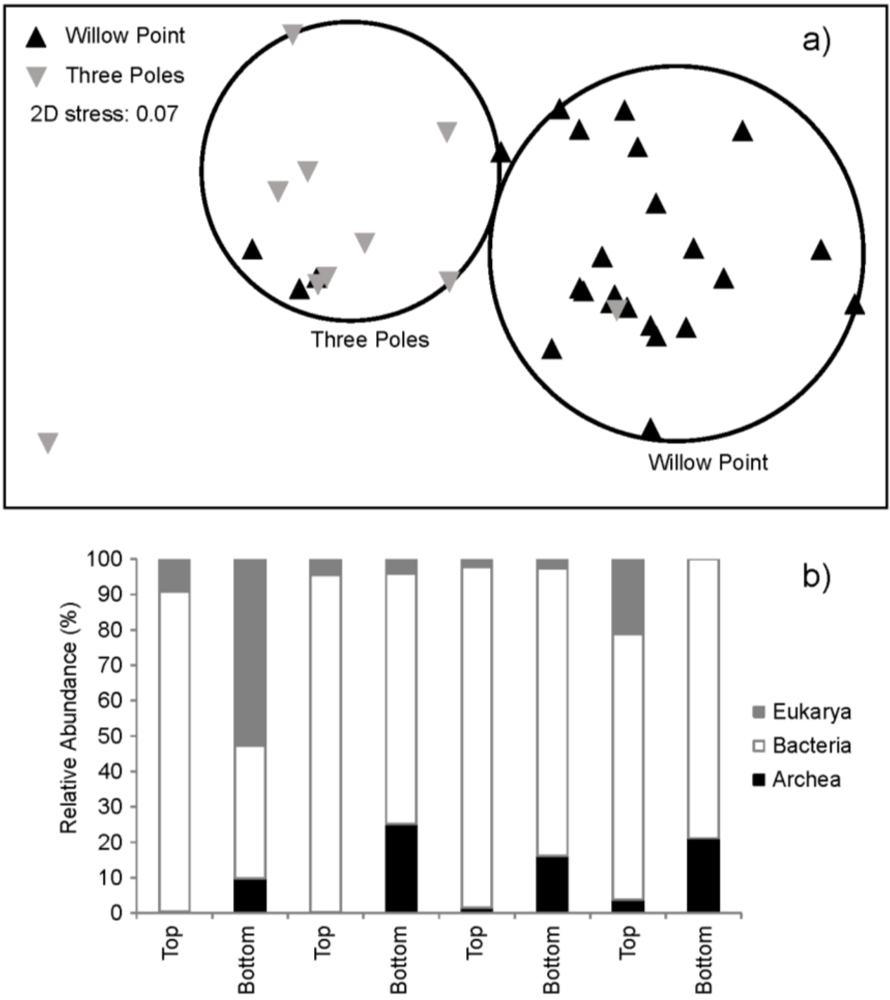
2.4. Community Structure
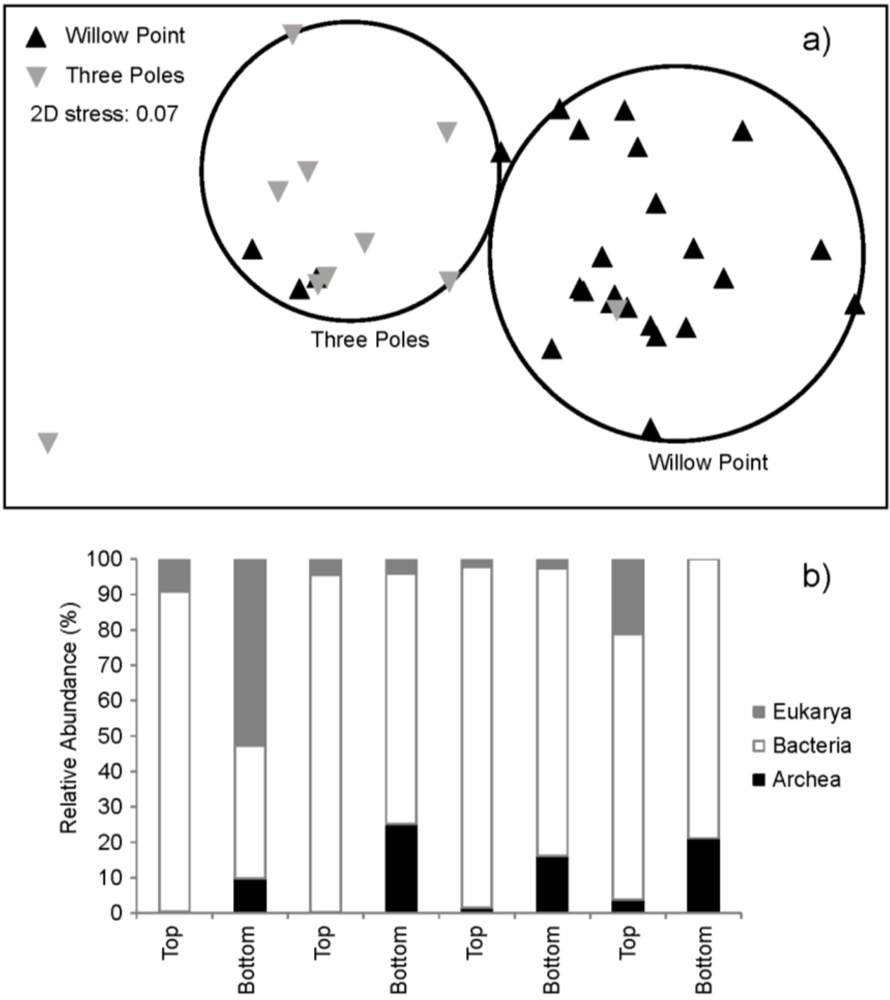
2.5. Analysis of Quorum Sensing Molecules
| Signal(s) | Microbe(s) | Synthesis | Precursor(s) | Receptor(s) | Regulated function(s) |
|---|---|---|---|---|---|
| Acyl homoserine lactones (AHLs) | Proteo-bacteria | -LuxI-type enzymes | -SAM, acyl-ACP | -LuxR-type proteins | Various processes |
| -AinS-type enzymes | -SAM, acyl-ACP, acyl-CoA | -two-component kinase | |||
| Linear oligopeptides | Gram-positive organisms | Genetically encoded | Secreted prepeptide | Two-component systems, phosphorelays | Various processes |
| Cyclical oligopeptides | Gram-positive organisms | Genetically encoded | Secreted post-syn mod. Prepeptide | Two-component systems, phosphorelays | Virulence genes |
| γ-Butyro-lactones (GBLs) | Streptomyces spp. | AfsA-type GBL synthase | Acyl-ACP, glycerol | ArpA-type repressor, GBL-binding proteins | Secondary metabolism; antibiotics and sporulation |
| Furanosyl diester (+/- boron); AI-2 | Diverse taxa (no a- proteo- bacteria) | LuxS AI-2 synthase | Methionine slavage (DPD) | Vibrio spp. LuxP/LuxQ/LuxO phosphorelay; unknown for other systems | Luminescence and diverse processes |
| Cis-11-Methyl-2-dodecenoic acid (DSF) | Xanthomonas spp., perhaps others | RpfB,RpfF | Ayl-CoA | RpfC, RpfH | Virulence and pigmentation |
| 4-Hydroxy-2-alkyl quinolines (PQS,HAQs) | Pseudomonads | pqs ABCDE, pqsH | Anthranilic acid | Unknown | Global regulation, virulence |
3. Discussion and Conclusions
References
- Lim, D.S.S.; Laval, B.E.; Salter, G.; Antoniades, D.; Forrest, A.L.; Pike, W.; Pieters, R.; Saffari, M.; Reid, D.; Schulze-Makuch, D.; Andersen, D.; McKay, C.P. Limnology of Pavilion Lake, B.C., Canada—Characterization of a microbialite forming environment. Fundam. Appl. Limnol. 2009, 173, 329–351. [Google Scholar]
- Laval, B.; Cady, S.L.; Pollack, J.C.; McKay, C.P.; Bird, J.S.; Grotzinger, J.P.; Ford, D.C.; Bohm, H.R. Modern freshwater microbialite analogues for ancient dendritic reef structures. Nature 2000, 407, 626–629. [Google Scholar]
- Brady, A.L.; Slater, G.F.; Omelon, C.R.; Southam, G.; Druschel, G.; Andersen, D.T.; Hawes, I.; Laval, B.; Lim, D.S.S. Photosynthetic isotope biosignatures in laminated micro-stromatolitic and non-laminated nodules associated with modern, freshwater microbialites in Pavilion Lake, B.C. Chem. Geol. 2010, 274, 56–67. [Google Scholar]
- Lim, D.S.S.; Brady, A.L.; Abercromby, A.F.; Andersen, D.T.; Andersen, M.; Arnold, R.R.; Bird, J.S.; Bohm, H.R.; Booth, L.; Cady, S.L.; Cardman, Z.; Chan, A.M.; Chan, O.; Chénard, C.; Cowie, B.R.; Davila, A.; Deans, M.C.; Dearing, W.; Delaney, M.; Downs, M.; Fong, T.; Forrest, A.; Gernhardt, M.L.; Gutsche, J.R.; Hadfield, C.; Hamilton, A.; Hawes, I.; Hansen, J.; Heaton, J.; Imam, Y.; Laval, B.L.; Lees, D.; Leoni, L.; Looper, C.; Love, S.; Marinova, M.M.; McCombs, D.; McKay, C.P.; Mireau, B.; Mullins, G.; Nebel, S.H.; Nuytten, P.; Pendery, R.; Pike, W.; Pointing, S.B.; Pollack, J.; Raineault, N.; Reay, M.; Reid, D.; Sallstedt, T.; Schulze-Makuch, D.; Seibert, M.; Shepard, R.; Slater, G.F.; Stonehouse, J.; Sumner, D.Y.; Suttle, C.A.; Trembanis, A.; Turse, C.; Wilhelm, M.; Wilkinson, N.; Williams, D.; Winget, D.M.; Winter, C. A historical overview of the Pavilion Lake Research Project—Analog science and exploration in an underwater environment. Geol. Soc. Am. 2011, 483, 85–116. [Google Scholar]
- Lim, D.S.S.; Harwood, C.; Sumner, D.; Omelon, C.; Nienow, J.; Russell, J.; Biddle, J.; Brady, A.L.; Reid, D.; McKay, C.P. Deep water microbialites of Pavilion Lake, Canada. Geobiology 2012. submitted. [Google Scholar]
- Brady, A.; Slater, G.F.; Laval, B.; Lim, D.S.S. Constraining carbon sources and growth rates of freshwater microbialites in Pavilion Lake using 14C analysis. Geobiology 2009, 7, 544–555. [Google Scholar] [CrossRef]
- White, D.C.; Bobbie, R.J.; Nichols, J.S.; Davis, W.M.; Fazio, S.D. Nonselective biochemical methods for the determination of fungal mass and community structure in estuarine detrital microflora. Botanica marina 1980, 23, 239–250. [Google Scholar]
- Gregory, A.D. Microbial Insights. Technical Report for Washington State University, Pavilion Lake Project: Washington, DC, USA, 2005; pp. 1–14. [Google Scholar]
- Edlund, A.; Nichols, P.D.; Roffey, R.; White, D.C. Extractable and lipopolysaccharide fatty acid and hydroxy acid profiles from Desulfovibrio species. J. Lipid Res. 1985, 26, 982–988. [Google Scholar]
- Dowling, N.J.E.; Widdel, F.; White, D.C. Phospholipid ester-linked fatty acid biomarkers of acetate-oxidizing sulfate reducers and other sulfide-forming bacteria. J. Gen. Microbiol. 1986, 132, 1815–1825. [Google Scholar]
- Parker, J.H.; Smith, G.A.; Fredrickson, H.L.; Vestal, J.R.; White, D.C. Sensitive assay, based on hydroxy-fatty acids from lipopolysaccharide lipid A for gram negative bacteria in sediments. Appl. Environ. Microbiol. 1982, 44, 1170–1177. [Google Scholar]
- Bhat, R.U.; Carlson, R.W. A new method for the analysis of amide-linked hydroxy fatty acids in lipid-A from gram-negative bacteria. Glycobiology 1992, 2, 535–539. [Google Scholar] [CrossRef]
- Smith, G.A.; Nickels, J.S.; Kerger, R. Quantitative characterization of microbial biomass and community structure in subsurface material: a prokaryotic consortium responsive to organic contamination. Can. J. Microbiol. 1986, 32, 104–111. [Google Scholar] [CrossRef]
- Schulze-Makuch, D.; Kennedy, J.F. Microbiological and chemical characterization of hydrothermal fluids at Tortugas Mountain Geothermal Area, southern New Mexico, USA. Hydrol. J. 2000, 8, 295–309. [Google Scholar]
- Schulze-Makuch, D.; Goodell, P.; Kretzschmar, T.; Kennedy, J.F. Microbial and chemical characterization of a groundwater flow system in an intermontane basin of southern New Mexico, USA. Hydrol. J. 2003, 11, 401–412. [Google Scholar]
- Kenyon, C.N. Fatty acid composition of unicellular strains of blue-green algae. J. Bacteriol. 1972, 109, 827–834. [Google Scholar]
- Kenyon, C.N.; Rippka, R.; Stanier, R.Y. Fatty acid composition and physiological properties of some filamentous blue-green algae. Arch. Microbiol. 1972, 83, 216–236. [Google Scholar]
- Caudales, R.; Wells, J.M.; butterfield, J.E. Cellular fatty acid composition of cyanobacteria assigned to subsection II, order Pleurocapsales. Int. J. Syst. Evol. Microbiol. 2000, 50, 1029–1034. [Google Scholar]
- Fang, J.; Chan, O.; Joeckel, R.M.; Huang, Y.; Wang, Y.; Bazylinski, D.A.; Moorman, T.B.; Clement, B.J.A. Biomarker analysis of microbial diversity in sediments of a saline groundwater seep of Salt Basin, Nebraska. Org. Geochem. 2006, 37, 912–931. [Google Scholar] [CrossRef]
- Bühring, S.I.; Smittenberg, R.H.; Sachse, D.; Lipp, J.S.; Golubic, S.; Sachs, J.P.; Hinrichs, K.-U.; Summons, R.E. A hypersaline microbial mat from the Pacific Atoll Kiritimati: Insights into composition and carbon fixation using biomarker analyses and a 13C-labeling approach. Geobiology 2009, 7, 308–323. [Google Scholar] [CrossRef]
- Guckert, J.B.; Antworth, C.P.; Nichols, P.D.; White, D.C. Phospholipid, ester-linked fatty acid profiles as reproducible assays for changes in prokaryotic community structure of estuarine sediment. FEMS. Microbiol. Ecol. 1985, 31, 147–158. [Google Scholar]
- Guckert, J.B.; Hood, M.A.; White, D.C. Phospholipid, ester-linked fatty acid profile changes during nutrient deprivation of Vibrio cholerae: increases in the trans/cis ratio and proportions of cyclopropyl fatty acids. Appl. Environ. Microbiol. 1986, 52, 147–158. [Google Scholar]
- Heipieper, H.J.; Diefenbach, R.; Keweloh, H. Conversion of cis unsaturated fatty acids to trans, a possible mechanism for the protection of phenol degrading Pseudomonas putida P8 from substrate toxicity. Appl. Environ. Microbiol. 1992, 58, 1847–1852. [Google Scholar]
- Tsitko, I.V.; Zaitsev, G.M.; Lobanok, A.G.; Salkinoja-Salonen, M.S. Effect of Aromatic Compounds on Cellular Fatty Acid Composition of Rhodococcus opacus. Appl. Environ. Microbiol. 1999, 65, 853–855. [Google Scholar]
- White, D.C.; Ringelberg, D.B. Utility of the signature lipid biomarker analysis in determining the in-situ viable biomass, community structure and nutritional/physiologic status of deep subsurface microbiota. In The Microbiology of the Terrestrial Deep Subsurface; Amy, P.S., Haldeman, D.L., Eds.; CRC Press: Boca Raton, FL, USA, 1997; pp. 119–136. [Google Scholar]
- Balkwill, D.L.; Leach, F.R.; Wilson, J.T.; McNabb, J.F.; White, D.C. Equivalence of microbial biomass measures based on membrane lipid and cell wall components, adenosine triphosphate and direct counts in subsurface aquifer sediments. Microb. Ecol. 1988, 16, 73–84. [Google Scholar] [CrossRef]
- Foster, J.S.; Green, S.J.; Ahrendt, S.R.; Golubic, S., Reid; Hetherington, K.L.; Bebout, L. Molecular and morphological characterization of cyanobacterial diversity in the stromatolites of Highborne Cay, Bahamas. ISME J. 2009, 3, 573–587. [Google Scholar]
- Goh, F.; Allen, M.A.; Leuko, S.; Kawaguchi, T.; Decho, A.W.; Burns, B.P.; Neilan, P.A. Determining the specific microbial populations and their spatial distribution within the stromatolite ecosystem of Shark Bay. ISME J. 2009, 3, 383–396. [Google Scholar] [CrossRef]
- Visick, K.L.; Fuqua, C. Decoding microbial chatter: Cell-cell communication in bacteria. J. Bacteriol. 2005, 187, 5507–5519. [Google Scholar] [CrossRef]
- Fuqua, W.C.; Winans, S.C.; Greenberg, E.P. Quorum sensing in bacteria: the LuxR-LuxI family of cell density-responsive transcriptional regulators. J. Bacteriol. 1994, 176, 269–275. [Google Scholar]
- Whitehead, N.A.; Barnard, A.M.; Slater, H.; Simpson, N.J.; Salmond, G.P. Quorum-sensing in Gram-negative bacteria. FEMS Microbiol. Rev. 2001, 25, 365–404. [Google Scholar] [CrossRef]
- Smith, J.N.; Ahmer, B.M. Detection of other microbial species by Salmonella: expression of the SdiA regulon. J. Bacteriol. 2003, 185, 1357–1366. [Google Scholar] [CrossRef]
- Wagner-Döbler, I.; Thiel, V.; Allgaier, M.; Bodor, A.; Meyer, S.; Ebner, S.; Hennig, A.; Pukall, R.; Schulz, S. Discovery of complex mixtures of novel long-chain quorum sensing signals in free-living and host-associated marine alphaproteobacteria. Chembiochem 2005, 6, 2195–2206. [Google Scholar] [CrossRef]
- Sun, J.; Daniel, R.; Wagner-Döbler, I.; Zeng, A.P. Is autoinducer-2 a universal signal for interspecies communication: A comparative genomic and phylogenetic analysis of the synthesis and signal transduction pathways. BMC Evol. Biol. 2004, 4, 36–47. [Google Scholar]
- NC-UIB; Nomenclature for Incompletely Specified Bases in Nucleic Acid Sequences. Available online: http://www.chem.qmul.ac.uk/iubmb/misc/naseq.html (accessed on 20 August 2012).
- Don, R.H.; Cox, P.T.; Wainwright, B.J.; Baker, K.; Mattick, J.S. Touchdown PCR to circumvent spurious priming during gene amplification. Nucleic Acids Res. 1991, 19, 4008. [Google Scholar] [CrossRef]
- Hecker, K.H.; Roux, K.H. High and Low annealing temperatures increase both specificity and yield in touchdown and stepdown pcr. Biotechniques 1996, 20, 478–485. [Google Scholar]
- Korbie, D.J.; Mattick, J.S. Touchdown PCR for increased specificity and sensitivity in PCR amplification. Nat. Protoc. 2008, 3, 1452–1456. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Schulze-Makuch, D.; Lim, D.; Laval, B.; Turse, C.; António, M.R.d.S.; Chan, O.; Pointing, S.B.; Brady, A.; Reid, D.; Irwin, L.N. Pavilion Lake Microbialites: Morphological, Molecular and Biochemical Evidence for a Cold-Water Transition to Colonial Aggregates. Life 2013, 3, 21-37. https://doi.org/10.3390/life3010021
Schulze-Makuch D, Lim D, Laval B, Turse C, António MRdS, Chan O, Pointing SB, Brady A, Reid D, Irwin LN. Pavilion Lake Microbialites: Morphological, Molecular and Biochemical Evidence for a Cold-Water Transition to Colonial Aggregates. Life. 2013; 3(1):21-37. https://doi.org/10.3390/life3010021
Chicago/Turabian StyleSchulze-Makuch, Dirk, Darlene Lim, Bernard Laval, Carol Turse, Marina Resendes de Sousa António, Olivia Chan, Stephen B. Pointing, Allyson Brady, Donnie Reid, and Louis N. Irwin. 2013. "Pavilion Lake Microbialites: Morphological, Molecular and Biochemical Evidence for a Cold-Water Transition to Colonial Aggregates" Life 3, no. 1: 21-37. https://doi.org/10.3390/life3010021
APA StyleSchulze-Makuch, D., Lim, D., Laval, B., Turse, C., António, M. R. d. S., Chan, O., Pointing, S. B., Brady, A., Reid, D., & Irwin, L. N. (2013). Pavilion Lake Microbialites: Morphological, Molecular and Biochemical Evidence for a Cold-Water Transition to Colonial Aggregates. Life, 3(1), 21-37. https://doi.org/10.3390/life3010021