
Interaction between Microbes, Minerals, and Fluids in Deep-Sea Hydrothermal Systems


Deep Sea Science Division, Institute of Deep-Sea Science and Engineering, Chinese Academy of Sciences, Sanya 572000, China
*
Author to whom correspondence should be addressed.
Minerals 2021, 11(12), 1324; https://doi.org/10.3390/min11121324
Submission received: 15 October 2021
/
Revised: 13 November 2021
/
Accepted: 17 November 2021
/
Published: 26 November 2021
(This article belongs to the Special Issue Microorganisms and Minerals in Natural and Engineered Environments)
Abstract
:The discovery of deep-sea hydrothermal vents in the late 1970s widened the limits of life and habitability. The mixing of oxidizing seawater and reduction of hydrothermal fluids create a chemical disequilibrium that is exploited by chemosynthetic bacteria and archaea to harness energy by converting inorganic carbon into organic biomass. Due to the rich variety of chemical sources and steep physico-chemical gradients, a large array of microorganisms thrive in these extreme environments, which includes but are not restricted to chemolithoautotrophs, heterotrophs, and mixotrophs. Past research has revealed the underlying relationship of these microbial communities with the subsurface geology and hydrothermal geochemistry. Endolithic microbial communities at the ocean floor catalyze a number of redox reactions through various metabolic activities. Hydrothermal chimneys harbor Fe-reducers, sulfur-reducers, sulfide and H2-oxidizers, methanogens, and heterotrophs that continuously interact with the basaltic, carbonate, or ultramafic basement rocks for energy-yielding reactions. Here, we briefly review the global deep-sea hydrothermal systems, microbial diversity, and microbe–mineral interactions therein to obtain in-depth knowledge of the biogeochemistry in such a unique and geologically critical subseafloor environment.
1. Introduction
It has become increasingly apparent that microbial communities are not limited to shallow near-seafloor environments but could extend into the deep subsurface. Hydrothermal vents form one such habitat, abundant in microbial communities that harbor energy through processes largely different from the surface photosynthetic biosphere. These structures are formed when superheated, mineral-rich hot water percolate cracks, fissures, and comes into contact with cold seawater. Deep hydrothermal vents are located in areas with high tectonic activity, mid-ocean ridges, and seamounts. With their first discovery in the Galapagos rift [1], it uncovered new potential habitats previously unknown to us and changed our view of life on Earth. In the complete absence of light, chemosynthesis is a process in which chemical energy is used to convert carbon dioxide into sugar, which is the predominant form for organic carbon production in this environment [2].
To date, about 600 active vent sites are known, mostly along ridge boundaries [3]. Hydrothermal vents globally occur at sea-floor spreading zones and along the spreading zones [4,5] (Figure 1), with the discovery of new vent systems being studied in detail. While during eruptions some vents produce black smokers (such as the Mid Atlantic Ridge system (MAR), temperature ~400 °C) with diverse microbial communities on the vent exteriors, others such as the Lost City Hydrothermal Field (temperature of ~90–150 °C), in contrast, are dominated by carbonates and profoundly different than black smokers in terms of their geochemistry and microbial community structures depending on the temperature [6].
The active vents are characterized by the presence of different rock types, such as basalts, peridotites, and felsic rocks. The vented fluids exhibit large chemical variability depending on the mineralogical differences in the underlying rocks [7]. Hydrothermal fluids may be nearly colorless and have low concentrations of chemical species of dissolved minerals, or may be white or black, characterized by high concentrations of minerals. Sulfides and other sulfur minerals are common in this environment. The vent ecosystems host a lively habitat and complex ecosystem characterized by steep temperatures and geochemical gradients [8]. These deep-sea chemosynthetic ecosystems are fueled by chemical reactions between oxidizing components in seawater (e.g., O2 and SO4) and reducing components originating from/beneath the sea-floor (e.g., H2, CH4, and H2S). Permeable mineral structures and the continued mixing of both reduced vent fluids and oxidized seawater provide favorable growth conditions for microbial communities [9]. Additionally, termed as chemolithoautotrophs, these microorganisms synthesize organic matter that form the base of the food chain in these ecosystems. Abiotic redox reactions in the mixing zones also play a potentially vital role in hydrothermal fields. Hydrocarbon formation at the Piccard field was shown to be contributed by biomass pyrolysis, while abiotic sulfate reductions produced large depletions in H2 [10]. Since chemolithotrophs commonly colonize hydrothermal vents, a combination of biogenic and thermogenic processes is likely to contribute to the production of H2, CH4, and other organic compounds, such as in the MAR and in the South West Indian Ocean (SWIR) systems [11,12].
Gaps in the understanding of hydrothermal vent ecosystems [13] have been partially filled in the last ten years of research through the application of both culture-dependent and independent studies. Combined with advanced geochemical analyses, we now have a better perspective of the microbial communities inhabiting such ecosystems e.g., [14,15,16,17]. Lipid biomarkers were also used as proxies to reconstruct the community structure of vent systems. Branched fatty acids (iso/anteiso-C15 and C17) were detected in a SWIR sulfide chimney sample, indicating the presence of sulfate-reducing bacteria [18]. Recently, brGDGTs were detected from both active and inactive hydrothermal chimneys from the SWIR (Dasgupta et al. unpublished data) and were considered to be associated with autochthonous thermophilic bacteria [19,20]. Studies have shown that geochemical differences within vent systems are the strongest driving forces in structuring the microbial communities of a particular vent field [21]. The role of geochemistry is particularly critical when comparing sites with different host rock geology. Sulfur oxidation is the prevalent electron donor in basalt-hosted fast-spreading ridges, while hydrogen and methane are the key electron sources for the microbial metabolism of ultramafic systems, with slow and ultraslow spreading rates [5].
The manifestation of chemical sources for lithotrophs along mid-oceanic ridges (MOR) occurs in three ways. First, when seawater circulates through hot subsurface igneous rocks, the resulting hydrothermal fluid becomes enriched in reduced compounds such as H2S, H2, Fe, and CH4, including other metals. The chemical reactions that follow between the reduced components and oxidized seawater are promoted by microbes to derive energy. Evaluation of the metabolic energy sources during fluid-mixing suggests that oxidation of H2S and H2 supply the highest levels of energy at >90%, the rest being derived from methanogenesis, sulfate reduction, and oxidation of Fe, Mn, or other metals [22,23,24]. The second source of energy is derived when oxidized seawater comes into contact with seafloor rocks. Basaltic rocks exposed at the seafloor are rich in reduced Fe, S, and other elements. Further precipitates from hydrothermal fluids form an extensive accumulation of reduced sulfides and other metals. Lithoautotrophs promote oxidative surface reactions when these reduced compounds are exposed to seawater. Oxidation of Fe and S by these microorganisms is one of the major energy sources for lithoautotrophs that can use mineral substrates in the seafloor environment [22,23,24]. The third type of energy source has been suggested to occur at high temperature hydrothermal vents [25,26,27]. In this process, hydrogen acts as the primary energy source, generated by the reaction of seawater with reduced metals in basaltic glass or Fe-rich minerals. This hydrogen can be combined with SO4−2 by sulfate reducers or with CO2 by methanogens for metabolic energy gain.
In this article, we review, in brief, the dominant microbial diversity and community structure of deep-sea hydrothermal vents, microbe–mineral interactions, and the biogeochemistry involved in such unique ecosystems.
2. Microbial Diversity in Deep-Sea Hydrothermal Vent Systems
2.1. Sulfur Oxidizers and Sulfate Reducers
Sulfide and thiosulfate oxidation in sulfide-rich basalt-hosted hydrothermal systems provide the greatest source of energy yields [28]. Bacterial sulfur-oxidation pathways include sulfide oxidation to elemental sulfur by sulfide dehydrogenase (FccAB) or sulfide-quinone reductase, which is further oxidized to sulfate. The reverse sulfate reduction is another alternative pathway in which thiosulfate can be oxidized to sulfate.
Campylobacterota is highly abundant in hydrothermal vent fields [14,29,30,31,32]; (refer to Table 1 for microbial groups identified in specific hydrothermal fields). Takai et al. [33,34,35] first reported sulfur-oxidizers from deep-sea hydrothermal vents, such as in the Suiyo Seamount hydrothermal system. Other mesophilic S-oxidizers isolated from these environments include those from genus Sulfurimonas [36], Sulfurovum [37], and Lebetimonas [38]. A majority of strains of the genus Sulfurovum grow chemolithoautotrophically using sulfur as an energy source. Similar communities were also found in cool, diffusive flows at the Axial Seamount on the Juan de Fuca Ridge (JFR) [39] and at the chimney walls’ biofilms at the Loki’s Castle vent field [31]. Ding et al. (2017) [15], in their study, recovered both Sulfurovum and Sulfurimonas from all active chimneys of SWIR and inactive sulfides in the East Pacific Rise (EPR) [40]. Thus, these groups possibly represent the most widely distributed species at active sulfide vents. The Sulfurimonas species have also been isolated from the Kairei and Pelagia active chimneys from the Central Indian Ridge (CIR) system [16]. Several sulfur-oxidizing bacteria (SOB) belonging to the class Gammaproteobacteria have been isolated from deep hydrothermal habitats. In a study at the Edmond vent field, called the CIR system, [41] observed that the bacterial phylotypes in the Edmond microbial mats are primarily affiliated ( > 90%) with the epsilon and gamma subclasses of the Proteobacteria. Furthermore, a number of samples from the Rainbow and Lucky Strike hydrothermal fields in the MAR system were dominated by Gammaproteobacteria (Rb-2, -3) that included mesophilic sulfide-oxidizing Thiomicrospira and several genera within Methylococcaceae [21]. Gammaproteobacteria were also present in all the chimney samples of the SWIR system [24], with the SOBs within the genera of Thiotrichaceae, Ectothiorhodospiraceae, Thiohalophilus, and Piscirickettsiaceae [15]. Both subclasses are closely related to sulfur oxidation.
Sulfate reduction is one of the primary processes in hydrothermal vents at a wide temperature range (4–90 °C) in the anoxic areas of the vent. Isolated genera include Caminibacter, Nautilia, Lebetimonas, and Cetia. Some deltaproteobacteria, such as Desulfovibrio, Desulfonauticus, and Desulfothermas, were isolated from sites at 13° EPR, SWIR, and Southern Okinawa Trough (SOT) [21,42,43,44]. Within the aquificales class, sulfur-reducing bacteria (SRB) including Desulfurobacterium indicum was isolated from an Indian Ocean hydrothermal vent [43] and from SWIR [24]. Another obligate anaerobic chemolithotroph Thermosulfidibacter takaii was isolated from the SOT that could couple hydrogen oxidation to sulfate reduction [45]. Cha et al. (2013) [46] isolated Desulfotomaculum tongense sp., a novel moderately thermophilic SRB belonging to phylum Firmicutes, from the Tofua Arc in the Tonga Trench. Firmicutes have also been identified from the Lost City hydrothermal vents [5]. A few reports of SRBs in inactive chimneys indicate that sulfate-reducing Nitrospirae dominate such hydrothermal environments [47]. Furthermore, in the SWIR, the phylotypes of deltaproteobacteria were primarily similar to a sulfate-reducing benzene-mineralizing consortium clone and a mud volcano microbial mat clone [24].
Archaeal diversity has also been reported in deep-sea hydrothermal vents both from phylum Euryarchaeo-ta and Crenarchaeota. For example, Pyrococcus yayanosii [48,49] and Pyrococcus kukulkanii [50], both from phylum Euryarchaeota, were isolated from deep-sea vents. Pyrolobus fumarii, an archaeon in the phylum Crenarchaeota, was also isolated from high-temperature chimneys [51]
2.2. Methanogens and Methane Oxidizers
Methanogens are one of the most important groups in high-temperature vent areas [52]. While known methanogens (methanococcaceae and methanocaldococcaceae; refer to Table 1 for details) were detected in the Rainbow hydrothermal field, they are strikingly absent at the Lucky Strike of MAR [21]. Methanococcales were also detected in Kairei and Pelagia, CIR [16]. Actively venting carbonate chimneys in the Lost City hydrothermal field (LCHF) are dominated by a novel phylotype of anaerobic methanogens from the Methanosarcinales order [5,26,53]. Some other species isolated from deep hydrothermal vents include Methanocaldococcus fervens [54], M. infernus [55], M. villosus [56], M. vulcanius [54], M. bathoardescens [57], M. jannaschii [58], Methanopyrus kandleri [59], and Methanothermococcus okinawensis [60]. Among methane oxidizers, the anaerobic oxidation of methane (AOM) is prevalent around some hydrothermal vents and may consume up to 75% of the methane produced by the vent [61]. Archaea of phylum Euryarchaeota are also found in hydrothermal vents, commonly referred to as anaerobic methane-oxidizing lineages (ANME). Anaerobic methanotrophic clade ANME-1 has been recovered from the LCHF carbonate chimneys [53].
2.3. Hydrogen-Oxidizers
Although different types of microorganisms oxidize hydrogen, only a few are capable of effectively using the energy gain for CO2 fixation [8]. Such chemoautotrophs have been isolated from various deep-sea hydrothermal fields, including aquificales, delta and gammaproteobacteria, campylobacteria, desulfurococcales, thermodesulfobacteriales, methanococcales, and deferribacterales [62] (refer to Table 1 for details). Hydrogen has also been shown to be an important energy source in vent fluids at the Logatchev and Rainbow fields in the MAR [52]. Deltaproteobacteria contribute to H-oxidation in anoxic vent habitats [63]. Desulfonauticus, a genus within Deltaproteobacteria, is a moderately thermophilic hydrogen-oxidizing sulfate reducer and was detected from the Rainbow field, MAR [21]. Two hydrogen-oxidizing Caminibacter species isolated from the Rainbow field showed differences in their oxygen tolerance as C. mediatlanticus is a strict anaerobe while C. profundus grows optimally with 0.5% oxygen [64].
2.4. Ammonium-Oxidizers and Nitrate-Reducers
Dissolved inorganic nitrogen in the form of ammonium is common in hydrothermal environments. Ammonia oxidation to nitrite is the first step of nitrification by ammonia-oxidizing bacteria and/or archaea (AOB/AOA), followed by oxidation to nitrate by nitrite-oxidizing bacteria (NOB). Thaumarchaeota was assigned as the major AOA in two high-temperature vent systems in the SWIR, apart from other AOB within Nitrococcus, Nitrospira, and Nitrosomonas (Table 1). AOA was also found to be more abundant than AOB in samples from the MAR and South Atlantic Ocean [65]. Affiliates of the group Nitrosopumilius, an autotrophic AOA, have been mostly detected in inactive chimneys, although some related sequences were also found in active chimneys [16,66], and accounted for 33% of the archaeal library in chimney samples from the Lau Basin [67]. Furthermore, crenarchaeol, a GDGT specific to ammonia-oxidizing archaea of Marine group I (Thaumarchaeota) [68,69] was recently detected in active and inactive chimney samples from the SWIR hydro-thermal field (Dasgupta et al. unpublished data). Fewer studies reported the presence of denitrifying communities in hydrothermal environments. Sulfurimonas sp. [36,52], Caminibacter sp. [70], and Sulfurovum sp. [37,71], all from phylum Campylobacterota, have been isolated as denitrifiers. A number of isolates in the phylum proteobacteria [72] and aquificae [70] have also been reported from the Juan de Fuca Ridge (JFR) and other hydrothermal fields.
2.5. Iron-Oxidizers and Iron-Reducers
Reduced metals such as Fe, Mn, Cu, etc., are potential energy sources for hydrothermal microbial communities. Iron-oxidizing bacteria proliferate at deep-hydrothermal systems, forming microbial mats [73]. Common Fe-oxidizing bacteria (FeOB) include chemolithotrophic Zetaproteobacteria and mixotrophic Gammaproteobacteria. Zetaproteobacteria Mariprofundus ferrooxydans [74,75] from the Loihi Seamount and Lau Basin [67], M. micogutta ET2 from the Izu-Ogasawara Arc [76], and several Alpha and gammaproteobacteria strains are reported as the chemoautotrophic Fe(II) oxidizers described from deep-sea vents [77] (Table 1). Sequences closely related to FeOBs in low-temperature hydrothermal vents were also detected from the SWIR [78]. Deep-sea hydrothermal fields can be ecological niches for Fe-reducers because they are abundant in oxidized iron [79]. While a few Fe-reducers have been detected in hydrothermal fields, a majority of them are from phylum proteobacteria [80] and deferribacteria [81]. Among archeae, most Fe-reducers belong to the families Archaeoglobacae and Pyrodictiaceae. Few other species were also isolated from the JFR [82] vent chimney on the Endeavor Segment of the North East Pacific Ocean.
3. Biomineralization and Microbe–Mineral Interactions
3.1. Iron-Metabolizers and Biomineralization
Minerals can serve either as electron acceptors or as sources of energy and electrons for microbes. Reduced metals and sulfur precipitate from hydrothermal fluids to form large reserves of metal sulfide minerals at the seafloor and are by far the most abundant source of energy available to microbes that can use mineral substrata in seafloor environments. During the exposure of these reduced compounds to seawater, lithoautotrophs can promote oxidative surface reactions for energy gain. Microbial surfaces can provide a large number of available nucleation sites for Fe-sulfides [83]. Additionally, a variety of biological structures are observed in the cell walls of the outer reactive surfaces of bacterial cell walls that are predominantly negatively charged and lead to the sorption of positive ions from solution as well as to the nucleation of metal sulfides.
Biotic reactions responsible for the formation of biogenic Fe-oxides include the microbial oxidation of Fe(II) to Fe(III) by a wide range of microorganisms under both acidic and neutral pH, as well as under oxic and anoxic conditions [84,85]. Characteristic ultrastructures, such as tubular sheaths, helical or twisted stalks, and irregular filaments, are generally considered as reliable indicators of biomineralization involving the FeOBs [74,86]. Such ultrastructures have been observed in the Lau Basin hydrothermal vent in the Western Pacific Ocean, where dominant mineralogy contained amorphous or poor crystalline minerals from the two-line ferrihydrite to opal-A [67,87]. Dense Fe-rich ultrastructures in the form of twisted stalks, branched stalks, rod-like sheaths, and contorted filaments detected in the Lau Basin are considered the most authentic biosignatures of FeOB species such as Mariprofundus sp., Gallionella sp., and Leptothrix sp. [88,89]. Alternatively, biogenic encrustation of Fe-sulfides around microbes is also feasible. Sulfate-reducing bacteria that supply reactive sulfide ions could be the key to the formation of sulfide minerals [90]. These microbes can further oxidize organic compounds or methane using O2 under aerobic conditions and SO42− in anoxic environments as terminal electron acceptors. More processes of FeOB–mineral interactions suggested that organic matter provides nucleation sites for the reaction of FeS to FeS2 [91]. The fact that many microbes were encrusted with pyrite and marcasite from the SWIR [24] supports this possible theory of nucleation. Biomineralization was observed in the microbial mats attached to the inner surface of a sulfide chimney (Figure 2) in the Edmond vent field of the CIR [41]. However, no Fe-oxidizing microorganisms were detected in their samples from molecular phylogenetic analysis. Nevertheless, a large number of species detected in the Edmond mat samples were sulfur-metabolizers, a few of which could also survive as Fe-oxidizers [92], that induce the extracellular nucleation of Fe-minerals.
Another possible onset of intracellular mineralization is the viability of cells. It is known that microbes can produce a variety of organic and inorganic intracellular polymers [93,94], and intracellular magnetic iron minerals (magnetite and greigite) can occur in magnetotactic bacteria [95,96,97]. Fe-reducing bacteria have also been shown to produce intracellular iron oxides. Although Peng et al. (2010) [41] identified magnetotactic bacteria and Fe-reducing archaea that can produce intracellular iron oxides in the Edmond field, the exact mechanisms for the high concentrations of iron (10.5–13.9 mmol/kg) remain arguable. Iron assimilatory reactions [98,99,100,101], by which many microbes could undergo intracellular mineralization in high iron solutions, is thought to be a possible mechanism. Iron dissimilatory reactions, where the metal ion is used as an electron donor or acceptor during respiration and is typically not incorporated into the cell, could be another potential mechanism [102,103] for the intracellular mineralization in Edmond microbial mats.
Some thermophilic H-oxidizers in hydrothermal fluids can also interact with the minerals through energy conservation mechanisms [104]. For example, hyperthermophilic Fe-reducers from sulfide chimneys oxidize H2 and transform ferrihydrite to magnetite, as well as other forms of Fe-minerals [105].
To examine whether mineral compositions have a direct relationship with bacterial and archaeal diversities in the SWIR [16], the proportions of chalcopyrite, pyrite, and sphalerite relative to the microbial community compositions were analyzed. No significant correlation with mineral species and with the abundance of microbial communities could be established. However, based on the nature of the inferred metabolism in the type of microbe detected, the availability of specific sulfide species could be relevant. For example, in the SWIR, while species such as Sulfurimonas use reduced sulfur compounds as electron donors, Desulfobulbus are sulfate-reducers. Thus, the presence of specific microbial species could indicate an abundance of related mineral groups in the hydrothermal systems.
3.2. Role of Microbes in the Precipitation of Mn-Oxides and Si in SWIR Hydrothermal Deposits
Precipitates of Mn-oxides in the hydrothermal deposits in the SWIR field are commonly attributed to Mn-oxidizing bacteria [24,78,106]. Mineralized ultrastructures of Fe-oxides, such as sheaths, twisted stalks, etc., in the hydrothermal fields are well reported [16,24,75], whereas reports of biogenic Mn-oxides are less frequent. These occur as fibrillar-like aggregates with a porous structure or reticulated filaments [107,108]. Li et al. (2013) [78] detected birnessite (a layered Mn-oxide) in two low-temperature deposits of the SWIR and speculated at least a partial biogenic origin of such a compound based on two observations: several putative Mn-oxidizing bacteria were present in their study and Mn-oxides with a fibrillar-like structure resembling a crumpled tissue are typical of the morphology of biogenic Mn-oxides [107].
Greenish clay mineral nontronite deposits are reported from deep hydrothermal vents, such as in the SE slope of the Eolo Seamount, Tyrrhenian Sea [109], and Okinawa Trough [110]. The tube-like filamentous structure of nontronites indicate the bacteria-assisted precipitation of these deposits. At the Galapagos Rift and Mariana Trough, correlation clay minerals and biological conditions at the hydrothermal chimneys suggested that Fe-oxidizing, sheath-forming bacteria play a decisive role in the nontronite formation at these sites [111]. Mineralized structures composed of silica were also observed in the SWIR deposits. Li et al. (2013) [78] suspects that these structures are abiogenic in origin, controlled by processes such as silica supersaturation and polymerization [112,113]. However, microorganisms present in the vent site could act as substrates and provide nucleation sites for Si-precipitation. Now, additional cooling in hydrothermal sites is essential to precipitate amorphous Si. Large accumulation of FeOB-induced ultrastructures, which locally separate hydrothermal fluid from ambient seawater and largely restrict their mixing, could lead to conductive heat removal of hydrothermal fluid rather than sole cooling through full mixing with seawater [114,115]. This would result in local Si supersaturation and more precipitation as grains of mineral spherules. Thus, there could be multiple indirect ways in which microbe–mineral interactions could facilitate the formation of Si-precipitates in deep hydrothermal deposits.
4. Conclusions
Since the discovery of hydrothermal vents more than four decades ago, it has been established that deep hydrothermal vents are characterized by unique habitats that support their growth by deriving energy from the reduced chemicals available in the hydrothermal fluids or altered bedrock. The steep chemical and thermal gradients prevailing in such environments flourish a rich diversity of microorganisms, with a range of metabolic activities and adaptation mechanisms. A general model of a deep-sea hydrothermal system has been given in Figure 3, depicting the biogeochemical diversity of such vent systems. In terms of converting inorganic carbon to organic biomass, the contribution of hydrothermal vent communities to the global sink is considered negligible. However, endolithic microbial communities in the sub-seafloor environment could have a major impact on chemosynthetic biomass production [6]. The common proposition that mineral particles form abiotically in such systems has been replaced by growing evidence that suggests that microorganisms are also actively moderating the formation of mineral particles within the vent and plumes. Nevertheless, caution must be exercised since mineral precipitates often resemble microbial morphologies. The interaction of microbes and minerals in deep hydrothermal fields can result in the accumulation of deposits and ultrastructures.
Interestingly, modern hydrothermal systems have long been considered as similar, to some extent, to early Earth’s conditions and hence can be considered as a place of interest for the search of the origin of life. Microenvironments, such as mineral surfaces, favor adsorption, the concentration of organics, and chemical reactions, and the hydrothermal niche provides physicochemical gradients and non-equilibrium conditions typical for the emergence of living organism [116]. The serpentinization process is also considered as a likely energy source essential for life to have emerged on the primitive Earth [117,118]. Furthermore, alkaline hydrothermal systems, such as the Lost City hydrothermal field, are thought to be more relevant to Archean hydrothermal vents, which can provide insights into the past mantle geochemistry and present a better understanding of the chemical constraints that existed during the onset of life [12]. Few other studies have also proposed a possible relation between low temperature shallow hydrothermal conditions [119], subsurface filamentous fabrics (SFF) [120], and the quest for Martian life.
In terms of technological advancements, a series of advanced analytical technologies or methods are used to study biomineralization. For instance, Fe isotopic compositions, together with nano SIMS (secondary ion mass spectroscopy) and FISH (fluorescence in situ hybridization), have been applied to study the formation mechanisms and preservation of hydrothermal mineralized microbial mats at the Loihi Seamount [121,122]. However, understanding the real depth of the adaptation mechanisms of vent microorganisms, how these microbes interact with minerals biochemically, the thermodynamic constraints, and how deep hydrothermal vents shape the global carbon cycle are some of the future challenges that need to be addressed.
Author Contributions
Conceptualization, S.D. and X.P.; Writing-original draft preparation, S.D.; Writing—review and editing, S.D. and X.P.; Resources, S.D., K.T. and X.P.; Supervision, X.P.; Funding acquisition, X.P. and S.D. All authors have read and agreed to the published version of the manuscript.
Funding
Completion of the manuscript was aided by grants from the National Key Research and Development Plan of China (2018YFC0309902) and the National Natural Science Foundation of China (42006061).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Lonsdale, P. Clustering of suspension feeding macrobenthos near abyssal hydrothermal vents at oceanic spreading centers. Deep Sea Res. A 1977, 24, 857–858. [Google Scholar] [CrossRef]
- Jannasch, H.W.; Wirsen, C.O. Chemosynthetic primary production at East Pacific sea floor spreading centers. Bioscience 1979, 29, 592–598. [Google Scholar] [CrossRef]
- Beaulieu, S.E.; Baker, E.T.; German, C.R. Where are the undiscovered hydrothermal vents on oceanic spreading ridges? Deep Sea Res. Part II Top Stud. Oceanogr. 2015, 121, 202–212. [Google Scholar] [CrossRef] [Green Version]
- Baker, E.T.; German, C.R. Mid-Ocean Ridges: Hydrothermal Interactions between the Litho-Sphere And Oceans; German, C., Lin, J., Parson, L.M., Eds.; American Geophysical Union: Washington, DC, USA, 2004; pp. 245–266. [Google Scholar]
- Martin, W.; Baross, J.; Kelley, D.; Russell, M.J. Hydrothermal vents and the origin of life. Nat. Rev. Microbiol. 2008, 6, 805–814. [Google Scholar] [CrossRef] [PubMed]
- Edwards, K.J.; Bach, W.; McCollom, T.M. Geomicrobiology in oceanography: Microbe–mineral interactions at and below the seafloor. Trends Microbiol. 2005, 13, 449–456. [Google Scholar] [CrossRef] [PubMed]
- Amend, J.P.; Mccollom, T.M.; Hentscher, M.; Bach, W. Catabolic and anabolic energy for chemo-lithoautotrophs in deep-sea hydrothermal systems hosted in different rock types. Geochim. Cosmochim. Acta 2011, 75, 5736–5748. [Google Scholar] [CrossRef]
- Jannasch, H.W.; Mottl, M.J. Geomicrobiology of deep-sea hydrothermal vents. Science 1985, 229, 717–725. [Google Scholar] [CrossRef]
- Frank, K.L.; Rogers, D.R.; Olins, H.C.; Vidoudez, C.; Girguis, P.R. Characterizing the distribution and rates of microbial sulfate reduction at Middle Valley hydrothermal vents. ISME J. 2013, 7, 1391–1401. [Google Scholar] [CrossRef]
- McDermott, J.M.; Sylva, S.P.; Ono, S.; German, C.R.; Seewald, J.S. Abiotic redox reactions in hydrothermal mixing zones: Decreased energy availability for the subsurface biosphere. Proc. Natl. Acad. Sci. USA 2020, 117, 20453–20461. [Google Scholar] [CrossRef]
- Kelley, D.S. Methane-bearing fluids in the oceanic crust: Gabbro-hosted fluid inclusions from the southwest Indian ridge. J. Geophys. Res. 1996, 101, 2943–2962. [Google Scholar] [CrossRef]
- Konn, C.; Charlou, J.L.; Holm, N.G.; Mousis, O. The production of methane, hydrogen, and organic compounds in ultramafic-hosted hydrothermal vents of the Mid-Atlantic Ridge. Astrobiology 2015, 15, 381–399. [Google Scholar] [CrossRef]
- Sievert, S.M.; Vetriani, C. Chemoautotrophy at deep-sea vents: Past, present, and future. Oceanography 2012, 25, 218–233. [Google Scholar] [CrossRef]
- Anderson, R.E.; Sogin, M.L.; Baross, J.A. Biogeography and ecology of the rare and abundant mi-crobial lineages in deep-sea hydrothermal vents. FEMS Microbiol. Ecol. 2015, 91, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, J.; Zhang, Y.; Jiang, H.; Leng, H.; Xiao, X. Microbial community structure of deep-sea hydro-thermal vents on the ultraslow spreading Southwest Indian Ridge. Front. Microbiol. 2017, 8, 1012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, Y.; Gonnella, G.; Adam, N.; Schippers, A.; Burkhardt, L.; Kurtz, S.; Schwarz-Schampera, U.; Franke, H.; Perner, M. Hydrothermal chimneys host habitat-specific microbial communities: Analogues for studying the possible impact of mining seafloor massive sulfide deposits. Sci. Rep. 2018, 8, 10386. [Google Scholar] [CrossRef]
- Dick, G.J. The microbiomes of deep-sea hydrothermal vents: Distributed globally, shaped locally. Nat. Rev. Microbiol. 2019, 17, 271–283. [Google Scholar] [CrossRef] [PubMed]
- Lei, J.; Chu, F.; Yu, X.; Li, X.; Tao, C. Lipid biomarkers reveal microbial communities in hydrothermal chimney structures from the 49.6ºE hydrothermal vent field at the Southwest Indian Ocean Ridge. Geomicrobiol. J. 2016, 34, 557–566. [Google Scholar] [CrossRef]
- Hu, J.F.; Meyers, P.A.; Chen, G.K.; Peng, P.A.; Yang, Q.H. Archaeal and bacterial glycerol dialkyl glycerol tetraethers in sediments from the Eastern Lau Spreading Center, South Pacific Ocean. Org. Geochem. 2012, 43, 162–167. [Google Scholar] [CrossRef]
- Jaeschke, A.; Eickmann, B.; Lang, S.Q.; Bernasconi, S.M.; Strauss, H.; Früh-Green, G.L. Biosignatures in chimney structures and sediment from the Loki’s Castle low-temperature hydrothermal vent field at the Arctic Mid-Ocean Ridge. Extremophiles 2014, 18, 545–560. [Google Scholar] [CrossRef] [PubMed]
- Flores, G.E.; Campbell, J.H.; Kirshtein, J.D.; Meneghin, J.; Podar, M.; Steinberg, J.I.; Seewald, J.S.; Tivey, M.K.; Voytek, M.A.; Yang, Z.K.; et al. Microbial community structure of hydrothermal deposits from geochemically different vent fields along the Mid-Atlantic Ridge. Environ. Microbiol. 2011, 13, 2158–2171. [Google Scholar] [CrossRef]
- McCollom, T.M. Geochemical constraints on primary productivity in submarine hydrothermal vent plumes. Deep-Sea Res. I 2000, 47, 85–101. [Google Scholar] [CrossRef]
- Shock, E.L.; Holland, M.E. Geochemical energy sources that support the subsurface biosphere. In The Subsurface Biosphere at Mid-Ocean Ridges; Wilcock, R.W., et al., Eds.; American Geophysical Union: Washington, DC, USA, 2004; pp. 153–166. [Google Scholar]
- Peng, X.T.; Chen, S.; Zhou, H.Y.; Zhang, L.X.; Wu, Z.J.; Li, J.T.; Li, J.W.; Xu, H.C. Diversity of biogenic minerals in low-temperature Si-rich deposits from a newly discovered hydrothermal field on the ultraslow spreading Southwest Indian Ridge. J. Geophys. Res. 2011, 116, G03030. [Google Scholar] [CrossRef] [Green Version]
- Takai, K.; Gamo, T.; Tsunogai, U.; Nakayama, N.; Hirayama, H.; Nealson, K.H.; Horikoshi, K. Geochemical and microbiological evidence for a hydrogen-based, hyperthermophilic subsurface lithoautotrophic microbial ecosystem (HyperSLiME) beneath an active deep-sea hydrothermal field. Extremophile 2004, 8, 269–282. [Google Scholar] [CrossRef] [PubMed]
- Kelley, D.S.; Karson, J.A.; Früh-Green, G.L.; Yoerger, D.R.; Shank, T.M.; Butterfield, D.A.; Hayes, J.M.; Schrenk, M.O.; Olson, E.J.; Proskurowski, G. A Serpentinite-Hosted Ecosystem: The Lost City Hydrothermal Field. Science 2005, 307, 1428–1434. [Google Scholar] [CrossRef] [PubMed]
- Ivarsson, M.; Lindblom, S.; Broman, C.; Holm, H.G. Fossilized microorganisms associated with zeolite–carbonate interfaces in sub-seafloor hydrothermal environments. Geobiology 2008, 6, 155–170. [Google Scholar] [CrossRef] [PubMed]
- McCollom, T.M. Geochemical constraints on source of metabolic energy for chemolithoautotrophy in ultramafic-hosted deep-sea hydrothermal systems. Astrobiology 2007, 7, 933–950. [Google Scholar] [CrossRef]
- Takai, K.; Nunoura, T.; Horikoshi, K.; Shibuya, T.; Nakamura, K.; Suzuki, Y.; Stott, M.; Massoth, G.J.; Christenson, B.W.; de Ronde, C.E.J.; et al. Variability in microbial communities in black smoker chimneys at the NW caldera vent field, Brothers volcano, Kermadec arc. Geomicrobiol. J. 2009, 26, 552–569. [Google Scholar] [CrossRef]
- Zhou, H.; Li, J.; Peng, X.; Meng, J.; Wang, F.; Ai, Y. Microbial diversity of a sulfide black smoker in main endeavour hydrothermal vent field, Juan de Fuca Ridge. J. Microbiol. 2009, 47, 235–247. [Google Scholar] [CrossRef] [PubMed]
- Dahle, H.; Roalkvam, I.; Thorseth, I.H.; Pedersen, R.B.; Steen, I.H. The versatile in situ gene expression of an Epsilonproteobacteria-dominated biofilm from a hydrothermal chimney. Environ. Microbiol. Rep. 2013, 5, 282–290. [Google Scholar] [CrossRef]
- Perner, M.; Gonnella, G.; Kurtz, S.; LaRoche, J. Handling temperature bursts reaching 464 °C: Different microbial strategies in the Sisters Peak hydrothermal chimney. Appl. Environ. Microbiol. 2014, 80, 4585–4598. [Google Scholar] [CrossRef] [Green Version]
- Takai, K.; Kobayashi, H.; Nealson, K.H.; Horikoshi, K. Deferribacter desulfuricans sp. nov., a novel sulfur-, nitrate-and arsenate-reducing thermophile isolated from a deep-sea hydrothermal vent. Int. J. Syst. Evol. 2003, 53, 839–846. [Google Scholar] [CrossRef]
- Takai, K.; Nakagawa, S.; Sako, Y.; Horikoshi, K. Balnearium lithotrophicum gen. nov., sp. nov., a novel thermophilic, strictly anaerobic, hydrogen-oxidizing chemolithoautotroph isolated from a black smoker chimney in the Suiyo Seamount hydrothermal system. Int. J. Syst. Evol. 2003, 53, 1847–1854. [Google Scholar] [CrossRef] [PubMed]
- Takai, K.; Inagaki, F.; Nakagawa, S.; Hirayama, H.; Nunoura, T.; Sako, Y.; Nealson, K.H.; Horikoshi, K. Isolation and phylogenetic diversity of members of previously uncultivated ε-Proteobacteria in deep-sea hydrothermal fields. FEMS Microbiol. Lett. 2003, 218, 167–174. [Google Scholar]
- Inagaki, F.; Takai, K.; Kobayashi, H.; Nealson, K.H.; Horikoshi, K. Sulfurimonas autotrophica gen. nov., sp. nov., a novel sulfuroxidizing ε-proteobacterium isolated from hydrothermal sediments in the Mid-Okinawa Trough. Int. J. Syst. Evol. Microbiol. 2003, 53, 1801–1805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inagaki, F.; Takai, K.; Nealson, K.H.; Horikoshi, K. Sulfurovum lithotrophicum gen. nov., sp. nov., a novel sulfur-oxidizing chemolithoautotroph within the ε-Proteobacteria isolated from Okinawa Trough hydrothermal sediments. Int. J. Syst. Evol. Microbiol. 2004, 54, 1477–1482. [Google Scholar] [CrossRef] [PubMed]
- Takai, K.; Hirayamam, H.; Nakagawa, T.; Suzuki, Y.; Nealson, K.H.; Horikoshi, K. Lebetimonas acidiphila gen. nov., sp. nov., a novel thermophilic, acidophilic, hydrogen-oxidizing chemolithoautotroph within the “Epsilonproteobacteria”, isolated from a deep-sea hydrothermal fumarole in the Mariana Arc. Int. J. Syst. Evol. Microbiol. 2005, 55, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Akerman, N.H.; Butterfield, D.A.; Huber, J.A. Phylogenetic diversity and functional gene patterns of sulfur-oxidizing subseafloor Epsilonproteobacteria in diffuse hydrothermal vent fluids. Front. Microbiol. 2013, 4, 185. [Google Scholar] [CrossRef] [Green Version]
- Sylvan, J.B.; Toner, B.M.; Edwards, K.J. Life and death of deep-sea vents: Bacterial diversity and ecosystem succession on inactive hydrothermal sulfides. MBio 2012, 3, e00279-11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, X.T.; Zhou, H.Y.; Li, J.T.; Li, J.W.; Chen, S.; Yao, H.Q.; Wu, Z.J. Intracellular and extracellular mineralization of a microbial community in the Edmond deep-sea vent field environment. Sediment. Geol. 2010, 229, 193–206. [Google Scholar] [CrossRef]
- Nunoura, T.; Oida, H.; Miyazaki, M.; Suzuki, Y.; Takai, K.; Horikoshi, K. Desulfothermus okinawensis sp. nov., a thermophilic and heterotrophic sulfate-reducing bacterium isolated from a deep-sea hydrothermal field. Int. J. Syst. Evol. Microbiol. 2007, 57, 2360–2364. [Google Scholar] [CrossRef]
- Cao, J.; Birien, T.; Gayet, N.; Huang, Z.; Shao, Z.; Jebbar, M.; Alain, K. Desulfurobacterium indicum sp. nov., a thermophilic sulfur reducing bacterium from the Indian Ocean. Int. J. Syst. Evol. Microbiol. 2017, 67, 1665–1668. [Google Scholar] [CrossRef] [PubMed]
- Hou, J.; Sievert, S.; Wang, Y.; Seewald, J.S.; Wang, F.; Xiao, X. Microbial succession during the transition from active to inactive stages of deep-sea hydrothermal vent sulfide chimneys. Microbiome 2020, 8, 102. [Google Scholar] [CrossRef] [PubMed]
- Nunoura, T.; Miyazaki, M.; Suzuki, Y.; Takai, K.; Horikoshi, K. Hydrogenivirga okinawensis sp. nov., a thermophilic sulfur-oxidizing chemolithoautotroph isolated from a deep-sea hydrothermal field, Southern Okinawa Trough. Int. J. Syst. Evol. Microbiol. 2008, 58, 676–681. [Google Scholar] [CrossRef] [PubMed]
- Cha, I.; Roh, S.W.; Kim, S.; Hong, H.; Lee, H.; Lim, W.; Rhee, S. Desulfotomaculum tongense sp. nov., a moderately thermophilic sulfate-reducing bacterium isolated from a hydrothermal vent sediment collected from the Tofua Arc in the Tonga Trench. Antonie Van Leeuwenhoek 2013, 104, 1185–1192. [Google Scholar] [CrossRef] [PubMed]
- Meier, D.; Pjevac, P.; Bach, W.; Markert, S.; Schweder, T.; Jamieson, J.; Petersen, S.; Amann, R.; Meyerdierks, A. Microbial metal sulfide oxidation in inactive hydrothermal vent chimneys suggested by metagenomic and metaproteomic analyses. Environ. Microbiol. 2019, 21, 682–701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Birrien, J.; Zeng, X.; Jebbar, M.; Cambonbonavita, M.; Querellou, J.; Oger, P.; Bienvenu, N.; Xiao, X.; Pri-eur, D. Pyrococcus yayanosii sp. nov., an obligate piezophilic hyperthermophilic archaeon isolated from a deep-sea hydrothermal vent. Int. J. Syst. Evol. Microbiol. 2011, 61, 2827–2881. [Google Scholar] [CrossRef] [Green Version]
- Zeng, X.; Birrien, J.; Fouquet, Y.; Cherkashov, G.; Jebbar, M.; Querellou, J.; Oger, P.; Cambon-Bonavita, M.A.; Xiao, X.; Prieur, D. Pyrococcus CH1, an obligate piezophilic hyperthermophile: Extending the upper pressure–temperature limits for life. ISME J. 2009, 3, 873–876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Callac, N.; Oger, P.; Lesongeur, F.; Rattray, J.E.; Vannier, P.; Michoud, G.; Beauverger, M.; Gayet, N.; Rouxel, O.; Jebbar, M.; et al. Pyrococcus kukulkanii sp. nov., a hyperthermophilic, piezophilic archaeon isolated from a deep-sea hydrothermal vent. Int. J. Syst. Evol. Microbiol. 2016, 66, 3142–3149. [Google Scholar] [CrossRef] [Green Version]
- Blochl, E.; Rachel, R.; Burggraf, S.; Hafenbradl, D.; Jannasch, H.W.; Stetter, K.O. Pyrolobus fumarii, gen. and sp. nov., represents a novel group of archaea, extending the upper temperature limit for life to 113 °C. Extremophiles 1997, 1, 14–21. [Google Scholar]
- Takai, K.; Suzuki, M.; Nakagawa, S.; Miyazaki, M.; Suzuki, Y.; Inagaki, F.; Horikoshi, K. Sulfurimonas paralvinellae sp. nov., a novel mesophilic, hydrogen- and sulfur-oxidizing chemolithoautotroph within the Epsilonproteobacteria isolated from a deep-sea hydrothermal vent polychaete nest, reclassification of Thiomicrospira denitrificans as Sulfurimonas denitrificans comb. nov. and emended description of the genus Sulfurimonas. Int. J. Syst. Evol. Microbiol. 2006, 56, 1725–1733. [Google Scholar]
- Brazelton, W.; Nelson, B.; Schrenk, M. Metagenomic evidence for H2 oxidation and H2 production by serpentinite-hosted subsurface microbial communities. Front. Microbiol. 2012, 2, 268. [Google Scholar] [CrossRef] [Green Version]
- Jeanthon, C.; Lharidon, S.; Reysenbach, A.; Corre, E.; Vernet, M.; Messner, P.; Sleytr, U.B.; Prieur, D. Methanococcus vulcanius sp. nov., a novel hyperthermophilic methanogen isolated from East Pacif-ic Rise, and identification of Methanococcus sp. DSM 4213T as Methanococcus fervens sp. nov. Int. J. Syst. Evol. Microbiol. 1999, 49, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Jeanthon, C.; L’Haridon, S.; Reysenbach, A.L.; Vernet, M.; Messner, P.; Sleytr, U.B.; Prieur, D. Methanococcus infernus sp. nov., a novel hyperthermophilic lithotrophic methanogen isolated from a deep-sea hydrothermal vent. Int. J. Syst. Bacteriol. 1998, 48, 913–919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bellack, A.; Huber, H.; Rachel, R.; Wanner, G.; Wirth, R. Methanocaldococcus villosus sp. nov., a heavily flagellated archaeon that adheres to surfaces and forms cell–cell contacts. Int. J. Syst. Evol. Microbiol. 2011, 61, 1239–1245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stewart, L.C.; Jung, J.; Kim, Y.; Kwon, S.; Park, C.; Holden, J.F. Methanocaldococcus bathoardescens sp. nov., a hyperthermophilic methanogen isolated from a volcanically active deep-sea hydrothermal vent. Int. J. Syst. Evol. Microbiol. 2015, 65, 1280–1283. [Google Scholar] [CrossRef] [Green Version]
- Jones, W.J.; Leigh, J.A.; Mayer, F.; Woese, C.R.; Wolfe, R.S. Methanococcus jannaschii sp. nov., an extremely thermophilic methanogen from a submarine hydrothermal vent. Arch. Microbiol. 1983, 136, 254–261. [Google Scholar] [CrossRef]
- Kurr, M.; Huber, R.; Konig, H.; Jannasch, H.W.; Fricke, H.; Trincone, A.; Kristjansson, J.K.; Stetter, K.O. Methanopyrus kandleri, gen. and sp. nov. represents a novel group of hyperthermophilic methanogens, growing at 110 C. Arch. Microbiol. 1991, 156, 239–247. [Google Scholar] [CrossRef]
- Takai, K.; Inoue, A.; Horikoshi, K. Methanothermococcus okinawensis sp. nov., a thermophilic, methane-producing archaeon isolated from a Western Pacific deep-sea hydrothermal vent system. Int. J. Syst. Evol. Microbiol. 2002, 52, 1089–1095. [Google Scholar]
- Wankel, S.D.; Adams, M.M.; Johnston, D.T.; Hansel, C.M.; Joye, S.B.; Girguis, P.R. Anaerobic methane oxidation in metalliferous hydrothermal sediments: Influence on carbon flux and decoupling from sulfate reduction. Environ. Microbiol. 2012, 14, 2726–2740. [Google Scholar] [CrossRef]
- Nakagawa, S.; Takai, K. Deep-sea vent chemoautotrophs: Diversity, biochemistry and ecological significance. FEMS Microbiol. Ecol. 2008, 65, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Adam, N.; Perner, M. Microbially mediated hydrogen cycling in deep-sea hydrothermal vents. Front. Microbiol. 2018, 9, 2873. [Google Scholar] [CrossRef]
- Miroshnichenko, M.L.; Lharidonm, S.; Schumann, P.; Spring, S.; Bonch-Osmolovskaya, E.A.; Jeanthon, C.; Stackebrandt, E. Caminibacter profundus sp. nov., a novel thermophile of Nautiliales ord. nov. within the class ‘Epsilonproteobacteria’, isolated from a deep-sea hydrothermal vent. Int. J. Syst. Evol. Microbiol. 2004, 54, 41–45. [Google Scholar] [CrossRef]
- Xu, W.; Li, M.; Ding, J.; Gu, J.; Luo, Z. Bacteria dominate the ammonia-oxidizing community in a hydrothermal vent site at the mid-atlantic ridge of the South Atlantic Ocean. Appl. Microbiol. Biotechnol. 2014, 98, 7993–8004. [Google Scholar] [CrossRef]
- Christakis, C.A.; Polymenakou, P.N.; Mandalakis, M.; Nomikou, P.; Kristoffersen, J.B.; Lampridou, D.; Kotoulas, G.; Magoulas, A. Microbial community differentiation between active and inactive sulfide chimneys of the Kolumbo submarine volcano, Hellenic Volcanic Arc. Extremophiles 2018, 22, 13–27. [Google Scholar] [CrossRef]
- Li, J.; Zhou, H.; Peng, X.; Wu, Z.; Chen, S.; Fang, J. Microbial diversity and biomineralization in low-temperature hydrothermal iron-silica-rich precipitates of the Lau Basin hydrothermal field. FEMS Microbiol. Ecol. 2012, 81, 205–216. [Google Scholar] [CrossRef] [Green Version]
- Pitcher, A.; Rychlik, N.; Hopmans, E.C.; Spieck, E.; Rijpstra, W.I.C.; Ossebaar, J.; Schouten, S.; Wagner, M.; Sinninghe Damsté, J.S. Crenarchaeol dominates the membrane lipids of Candidatus Nitrososphaera gargensis, a thermophilic group I.1b Archaeon. ISME J. 2010, 4, 542–552. [Google Scholar] [CrossRef] [PubMed]
- Pitcher, A.; Wuchter, C.; Siedenberg, K.; Schouten, S.; Sinninghe Damsté, J.S. Crenarchaeol tracks winter blooms of ammonia-oxidizing Thaumarchaeota in the coastal North Sea. Limnol. Oceanogr. 2011, 56, 2308–2318. [Google Scholar] [CrossRef] [Green Version]
- Alain, K.; Rolland, S.; Crassous, P.; Lesongeur, F.; Zbinden, M.; Le Gall, C.; Godfroy, A.; Page, A.; Juniper, S.K.; Cambonbonavita, M.; et al. Desulfurobacterium crinifex sp.nov., a novel thermophilic, pinkish-streamer forming, chemolithoautotrophic bacterium isolated from a Juan de Fuca Ridge hydrothermal vent and amendment of the genus Desulfurobacterium. Extremophiles 2003, 7, 361–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mori, K.; Yamaguchi, K.; Hanada, S. Sulfurovum denitrificans sp. nov., an obligately chemolithoautotrophic sulfur-oxidizing epsilonproteobacterium isolated from a hydrothermal field. Int. J. Syst. Evol. Microbiol. 2018, 68, 2183–2187. [Google Scholar] [CrossRef]
- Cao, J.; Gayet, N.; Zeng, X.; Shao, Z.; Jebbar, M.; Alain, K. Pseudodesulfovibrio indicus gen. nov., sp. nov., a piezophilic sulfate reducing bacterium from the Indian Ocean and reclassification of four spe-cies of the genus Desulfovibrio. Int. J. Syst. Evol. Microbiol. 2016, 66, 3904–3911. [Google Scholar] [CrossRef] [PubMed]
- Chan, C.S.; McAllister, S.; Leavitt, A.; Emerson, D.; Moyer, C.L.; Glazer, B.T. Fe-oxidizing microbes are hydrothermal vent ecosystem engineers at the Loihi Seamount. In AGU Fall Meeting Abstracts; Abstract B14B-01; American Geophysical Union: Washington, DC, USA, 2013. [Google Scholar]
- Emerson, D.; Moyer, C.L. Neutrophilic Fe-oxidizing bacteria are abundant at the Loihi Seamount hydrothermal vents and play a major role in Fe oxide deposition. Appl. Environ. Microbiol. 2002, 68, 3085–3093. [Google Scholar] [CrossRef] [Green Version]
- Emerson, D.; Rentz, J.A.; Liburn, T.G.; Davis, R.E.; Aldrich, H.; Chan, C.; Moyer, C.L. A novel line-age of Proteobacteria involved in formation of marine Fe-oxidizing microbial mat communities. PLoS ONE 2007, 2, e667. [Google Scholar] [CrossRef] [Green Version]
- Makita, H.; Tanaka, E.; Mitsunobu, S.; Miyazaki, M.; Nunoura, T.; Uematsu, K.; Takaki, Y.; Nishi, S.; Shimamura, S.; Takai, K. Mariprofundus micogutta sp. nov., a novel ironoxidizing zetaproteobacterium isolated from a deep-sea hydrothermal field at the Bayonnaise knoll of the Izu-Ogasawara arc, and a description of Mariprofundales ord. nov. and Zetaproteobacteria classis nov. Arch. Microbiol. 2017, 199, 335–346. [Google Scholar] [CrossRef] [PubMed]
- Edwards, K.J.; Rogers, D.R.; Wirsen, C.O.; McCollom, T.M. Isolation and characterization of novel psychrophilic, neutrophilic, Fe-oxidizing, chemolithoautotrophic α- and γ-Proteobacteria from the deep sea. Appl. Environ. Microbiol. 2003, 69, 2906–2913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Peng, X.; Zhou, H.; Li, J.; Sun, Z. Molecular evidence for microorganisms participating in Fe, Mn, and S biogeochemical cycling in two low-temperature hydrothermal fields at the Southwest Indian Ridge. J. Geophys. Res. Biogeosci. 2013, 118, 665–679. [Google Scholar] [CrossRef]
- Toner, B.M.; Marcus, M.A.; Edwards, K.J.; Rouxel, O.; German, C.R. Measuring the form of iron in hydrothermal plume particles. Oceanography 2012, 25, 209–212. [Google Scholar] [CrossRef]
- Li, J.; Sun, Y.; Fang, J.; Xie, W.; Peng, X.; Dong, L.; Wu, Z.; Zhou, H. Aerobic and anaerobic ammonia-oxidizing microorganisms in low-temperature hydrothermal Fe-Si-rich precipitates of the south-western Pacific Ocean. Geomicrobiol. J. 2013, 31, 42–52. [Google Scholar] [CrossRef]
- Slobodkina, G.; Reysenbach, A.L.; Panteleeva, A.; Kostrikina, N.; Wagner, I.; Bonch-Osmolovskaya, E.; Slobodkin, A.I. Deferrisoma camini gen. nov., sp. nov., a moderately thermophilic, dissimilatory iron (III)-reducing bacterium from a deep-sea hydrothermal vent that forms a distinct phylogenetic branch in the Deltaproteobacteria. Int. J. Syst. Evol. Microbiol. 2012, 62, 2463–2468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kashefi, K.; Lovley, D.R. Extending the upper temperature limit for life. Science 2003, 301, 934. [Google Scholar] [CrossRef] [PubMed]
- Ferris, F.G.; Fyfe, W.W.; Beveridge, T.J. Bacteria as nucleation sites for authigenic minerals in a metal-contaminated lake sediment. Chem. Geol. 1987, 63, 225–232. [Google Scholar] [CrossRef]
- Emerson, D.; Moyer, C.L. Isolation and characterization of novel iron-oxidizing bacteria that grow at circumneutral pH. Appl. Environ. Microbiol. 1997, 63, 4784–4792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emerson, D.; Weiss, J.V. Bacterial iron oxidation in circumneutral freshwater habitats: Findings from the field and the laboratory. Geomicrobiol. J. 2004, 21, 405–414. [Google Scholar] [CrossRef]
- Chan, C.S.; Fakra, S.C.; Emerson, D.; Fleming, E.J.; Edwards, K.J. Lithotrophic iron-oxidizing bacteria produce organic stalks to control mineral growth: Implication for biosignature formation. ISME J. 2011, 5, 717–727. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Su, L.; Wang, F.; Yang, J.; Gu, L.; Sun, M.; Li, Q.; Zhou, H.; Fang, J. Elucidating the biomineralization of low-temperature hydrothermal precipitates with varying Fe, Si contents: Indication from ultrastructure and microbiological analyses. Deep-Sea Res. I 2020, 157, 103208. [Google Scholar] [CrossRef]
- Emerson, D.; Fleming, E.J.; McBeth, J.M. Iron-oxidizing bacteria: An environmental and genomic perspective. Annu. Rev. Microbiol. 2010, 64, 561–583. [Google Scholar] [CrossRef] [PubMed]
- Fleming, E.J.; Cetinic, I.; Chan, C.S.; King, D.W.; Emerson, D. Ecological succession among iron-oxidizing bacteria. ISME J. 2014, 8, 804–815. [Google Scholar] [CrossRef]
- Frankel, R.B.; Bazylinski, D.A. Biologically induced mineralization by bacteria. Rev. Mineral. Geochem. 2003, 54, 95–114. [Google Scholar] [CrossRef]
- Grimes, S.T.; Brock, F.; Rickard, D.; Davies, K.L.; Edwards, D.; Briggs, D.E.G.; Parkes, R.J. Under-standing fossilization: Experimental pyritization of plants. Geology 2001, 29, 123–126. [Google Scholar] [CrossRef]
- Vidyalakshmi, R.; Paranthaman, R.; Bhakyaraj, R. Sulphur oxidizing bacteria and pulse nutrition—A review. World J. Agric. Sci. 2009, 5, 270–278. [Google Scholar]
- Fortin, D.; Langley, S. Formation and occurrence of biogenic iron-rich minerals. Earth Sci. Rev. 2005, 72, 1–19. [Google Scholar] [CrossRef]
- Edwards, K.J.; Bazylinski, D.A. Intracellular minerals and metal deposits in prokaryotes. Geobiology 2008, 6, 309–317. [Google Scholar] [CrossRef] [PubMed]
- Blakemore, R. Magnetotactic bacteria. Science 1975, 190, 377–379. [Google Scholar] [CrossRef] [PubMed]
- Petersen, N.; Dobeneck, T.; Vali, H. Fossil bacterial magnetite in deep sea sediments from the South Atlantic Ocean. Nature 1986, 320, 611–615. [Google Scholar] [CrossRef]
- Dong, Y.; Li, J.; Zhang, W.; Zhang, W.; Zhao, Y.; Xiao, T.; Wu, L.-F.; Pan, H. The detection of magnetotactic bacteria in deep sea sediments from the east Pacific Manganese Nodule Province. Environ. Microbiol. Rep. 2016, 8, 239–249. [Google Scholar] [CrossRef] [PubMed]
- Vainshtein, M.; Suzina, N.; Kudryashova, E.; Ariskina, E. New magnet-sensitive structures in bacterial and archaeal cells. Biol. Cell 2002, 94, 29–35. [Google Scholar] [CrossRef] [Green Version]
- Vainshtein, M.; Suzina, N.; Sorokin, V. A new type of magnet-sensitive inclusions in cells of photosynthetic bacteria. Syst. Appl. Microbiol. 1997, 20, 182–186. [Google Scholar] [CrossRef]
- Vadas, A.; Monbouquette, H.G.; Johnson, E.; Schröder, I. Identification and characterization of a novel ferric reductase from the hyperthermophilic archaeon Archaeoglobus fulgidus. J. Biol. Chem. 1999, 274, 36715–36721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiu, H.-J.; Johnson, E.; Schröder, I.; Rees, D.C. Crystal structures of a novel ferric reductase from the hyperthermophilic archaeon Archaeoglobus fulgidus and its complex with NADPþ. Structure 2001, 9, 311–319. [Google Scholar] [CrossRef] [Green Version]
- Juniper, S.K.; Martineau, P.; Sarrazin, J.; Gelinas, Y. Microbial-mineral floc associated with nascent hydrothermal activity on CoAxial segment, Juan de Fuca Ridge. Geophys. Res. Lett. 1995, 22, 179–182. [Google Scholar] [CrossRef]
- Holden, J.F.; Adams, M.W.W. Microbe–metal interactions in marine hydrothermal environments. Curr. Opin. Chem. Biol. 2003, 7, 160–165. [Google Scholar] [CrossRef]
- Takai, K.; Nakamura, K. Compositional physiological and metabolic variability in microbial communities associated with geochemically diverse deep-sea hydrothermal vent fluids. In Geomicrobiology: Molecular and Environmental Perspective; Barton, L.L., Mandl, M., Loy, A., Eds.; Springer: Berlin, Germany, 2010; pp. 251–283. [Google Scholar]
- Ver Eecke, H.C.; Kelley, D.S.; Holden, J.F. Abundances of hyperthermophilic autotrophic Fe(III) oxide reducers and heterotrophs in hydrothermal sulfide chimneys of the northeastern Pacific Ocean. Appl. Environ. Microbiol. 2009, 75, 242–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mandernack, K.W.; Tebo, B.M. Manganese scavenging and oxidation at hydrothermal vents and in vent plumes. Geochim. Cosmochim. Acta 1993, 57, 3907–3923. [Google Scholar] [CrossRef]
- Spilde, M.N.; Northup, D.E.; Boston, P.J.; Schelble, R.T.; Dano, K.E.; Crossey, L.J.; Dahm, C.N. Geomicrobiology of cave ferromanganese deposits: A field and laboratory investigation. Geomicrobiol. J. 2005, 22, 99–116. [Google Scholar] [CrossRef]
- Miller, A.Z.; Dionísio, A.; Braga, M.S.; Hernández-Mariné, M.; Afonso, M.J.; Muralha, V.S.F.; Herrera, L.K.; Raabe, J.; Fernandez-Cortes, A.; Cuezva, S.; et al. Biogenic Mn oxide minerals coating in a subsurface granite environment. Chem. Geol. 2012, 322–323, 181–191. [Google Scholar] [CrossRef] [Green Version]
- Dekov, V.M.; Kamenov, G.D.; Stmmeyer, J.; Thirty, M.; Savelli, C.; Shanks, W.C.; Fortin, D.; Kuzmann, E.; Vertes, A. Hydrothermal nontronite formation at Eolo Seamount (aeolian volcanic arc, Tyrrhenian Sea). Chem. Geol. 2007, 245, 103–119. [Google Scholar] [CrossRef]
- Ueshima, M.; Tazaki, K. Possible role of microbial polysaccharides nontronite formation. Clays Clay Miner. 2001, 49, 292–299. [Google Scholar] [CrossRef]
- Köhler, B.; Singer, A.; Stoffers, O. Biogenic nontronite from marine white smoker chimneys. Clays Clay Miner. 1994, 42, 689–701. [Google Scholar] [CrossRef]
- Konhauser, K.O.; Jones, B.; Phoenix, V.R.; Ferris, F.G.; Renaut, R.W. The microbial role in hot spring silicification. AMBIO 2004, 33, 552–558. [Google Scholar] [CrossRef] [PubMed]
- Jones, B.; de Ronde, C.E.J.; Renaut, R.W. Mineralized microbes from Giggenbach submarine volcano. J. Geophys. Res. 2008, 113, B08S05. [Google Scholar] [CrossRef] [Green Version]
- Halbach, M.; Halbach, P.; Lüders, V. Sulfide-impregnated and pure silica precipitates of hydrothermal origin from the Central Indian Ocean. Chem. Geol. 2002, 182, 357–375. [Google Scholar] [CrossRef]
- Jones, B.; Konhauser, K.O.; Renaut, R.W.; Wheeler, R.S. Microbial silicification in Iodine Pool, Waimangu geothermal area, North Island, New Zealand: Implications for recognition and identification of ancient silicified microbes. J. Geol. Soc. 2004, 161, 983–993. [Google Scholar] [CrossRef]
- Kompanichenko, V.N. Changeable hydrothermal media as potential cradle of life on a planet. Planet. Space Sci. 2009, 57, 468–476. [Google Scholar] [CrossRef]
- Russell, M.J.; Hall, A.J.; Turner, D. In vitro growth of iron sulfide chimneys: Possible culture chambers for origin-of-life experiments. Terra Nova 1989, 1, 238–241. [Google Scholar] [CrossRef]
- Russell, M.J.; Hall, A.J.; Martin, W. Serpentinization as a source of energy for the origin of life. Geobiology 2010, 8, 355–371. [Google Scholar] [CrossRef] [PubMed]
- Al-Hanbali, H.S.; Sowerby, S.J.; Holm, N.G. Biogenicity of silicified microbes from a hydrothermal system: Relevance to the search for evidence of life on earth and other planets. Earth Planet. Sci. Lett. 2001, 191, 213–218. [Google Scholar] [CrossRef]
- Hofmann, B.A.; Farmer, J.D.; Von Blanckenburg, F.; Fallick, A.E. Subsurface Filamentous Fabrics: An evaluation of origins based on morphological and geochemical criteria, with implications for exopaleontology. Astrobiology 2008, 8, 87–117. [Google Scholar] [CrossRef] [PubMed]
- Rouxel, O.; Toner, B.; Germain, Y.; Glazer, B. Geochemical and iron isotopic insights into hydrothermal iron oxyhydroxide deposit formation at Loihi Seamount. Geochim. Cosmochim. Acta 2018, 220, 449–482. [Google Scholar] [CrossRef] [Green Version]
- Dauphus, N.; John, S.G.; Rouxel, O. Iron isotope systematics. Rev. Mineral. Geochem. 2017, 82, 415–510. [Google Scholar] [CrossRef]
Figure 1.
Global distribution of major deep-sea hydrothermal systems. CIR: Central Indian Ridge and SWIR: South West Indian Ridge.
Figure 1.
Global distribution of major deep-sea hydrothermal systems. CIR: Central Indian Ridge and SWIR: South West Indian Ridge.

Figure 2.
Biomineralization in the chimney at the Edmond field, Central Indian Ridge hydrothermal system. (a) Yellow sulfide layers at the chimney exterior; (b) SEM image of biomineralized Fe stalks and spheroidal particles within the deposits.
Figure 2.
Biomineralization in the chimney at the Edmond field, Central Indian Ridge hydrothermal system. (a) Yellow sulfide layers at the chimney exterior; (b) SEM image of biomineralized Fe stalks and spheroidal particles within the deposits.

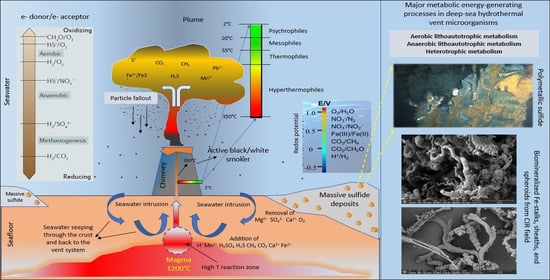
Figure 3.
Simplified representation of vent morphology and characteristics in terms of microbial communities with respect to temperature and redox potentials, and a list of the major energy-yielding processes used by microbes. The model is a generalized representation of hydrothermal vent systems and does not depict any particular location [6,17].
Figure 3.
Simplified representation of vent morphology and characteristics in terms of microbial communities with respect to temperature and redox potentials, and a list of the major energy-yielding processes used by microbes. The model is a generalized representation of hydrothermal vent systems and does not depict any particular location [6,17].


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Orders and genera of bacteria and archaea reported from deep-sea hydrothermal vents and their energy-yielding processes.
Table 1.
Orders and genera of bacteria and archaea reported from deep-sea hydrothermal vents and their energy-yielding processes.
| Categories (Bacteria and Archaea) | Main Energy-Yielding Process | Field Locations |
|---|---|---|
| Campylobacteria | S-oxidation, S-reduction, H-oxidation, and nitrate-reduction | |
| Sulfurimonas | S-oxidation, H-oxidation, and nitrate-reduction | JFR; Loki–s Castle, MAR; Suiyo Seamount; SWIR; EPR; and Kairei, Pelagia, CIR |
| Sulfurovum | ||
| Lebetimonas | ||
| Caminibacter | S-reduction, nitrate-reduction, and H-oxidation | EPR; SWIR; MAR; and JFR |
| Nautilia | ||
| Lebetimonas | ||
| Cetia | ||
| Gammaproteobacteria | S-oxidation, N-reduction, and H-oxidation | |
| Thiomicrospira | S-oxidation, N-reduction, and H-oxidation | CIR and MAR |
| Methylococcaceae | S-oxidation and N-reduction | |
| Thiotrichaceae | S-oxidation and N-reduction | SWIR |
| Ectothiorhodospiraceae | ||
| Thiohalophilus | ||
| Piscirickettsiaceae | ||
| Deltaproteobacteria | S-reduction, H-oxidation, and Fe-reduction, | |
| Desulfovibrio | S-reduction | EPR; SWIR; and SOT |
| Desulfonauticus | ||
| Desulfothermus | ||
| Desulfonauticus | H-oxidation | Rainbow, MAR |
| Deferrisoma | Fe-reduction | Eastern Lau Spreading Center |
| Zetaproteobacteria | Fe-oxidation | |
| Mariprofundus | Fe-oxidation | Loihi Seamount and Lau Basin |
| Betaproteobacteria | Ammonium-oxidation | |
| Nitrospira | Ammonium-oxidation | SWIR |
| Nitrococcus | ||
| Nitrosomonas | ||
| Aquificae | S-reduction, H-oxidation, and N-reduction | |
| Desulfobacterium | S-reduction | SWIR |
| H-oxidation | Eastern Lau Spreading Center | |
| N-reduction | JFR | |
| Firmicutes | S-reduction | |
| Desulfohalotomaculum | S-reduction | Tonga Trench |
| Euryarchaeota | Methanogenesis, methane-oxidation, and S-reduction | |
| Methanococcales | Methanogenesis | Kairei, Pelagia, CIR; Rainbow, MAR; and LCHF |
| Methanocaldococcus | ||
| Methanosarcinales | ||
| Methanopyrus | ||
| Pyrococcus | S-reduction | |
| ANME-1 | Methane-oxidation | LCHF |
| Crenarchaeota | S-reduction and Fe-reduction | |
| Pyrolobus | S-reduction | |
| Archaeoglobacae | Fe-reduction | JFR |
| Pyrodictiacceae | ||
| Thaumoarchaeota | Ammonium-oxidation | SWIR |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Dasgupta, S.; Peng, X.; Ta, K. Interaction between Microbes, Minerals, and Fluids in Deep-Sea Hydrothermal Systems. Minerals 2021, 11, 1324. https://doi.org/10.3390/min11121324
AMA Style
Dasgupta S, Peng X, Ta K. Interaction between Microbes, Minerals, and Fluids in Deep-Sea Hydrothermal Systems. Minerals. 2021; 11(12):1324. https://doi.org/10.3390/min11121324
Chicago/Turabian StyleDasgupta, Shamik, Xiaotong Peng, and Kaiwen Ta. 2021. "Interaction between Microbes, Minerals, and Fluids in Deep-Sea Hydrothermal Systems" Minerals 11, no. 12: 1324. https://doi.org/10.3390/min11121324
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.