The Effects of Grazing Systems on Plant Communities in Steppe Lands—A Case Study from Mongolia’s Pastoralists and Inner Mongolian Settlement Areas


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Grazing Systems of Inner Mongolia (China) and Mongolia
2.3. Field Sampling and Data Collection
2.4. The Plant Community Characteristics
2.5. Data Analysis
3. Results
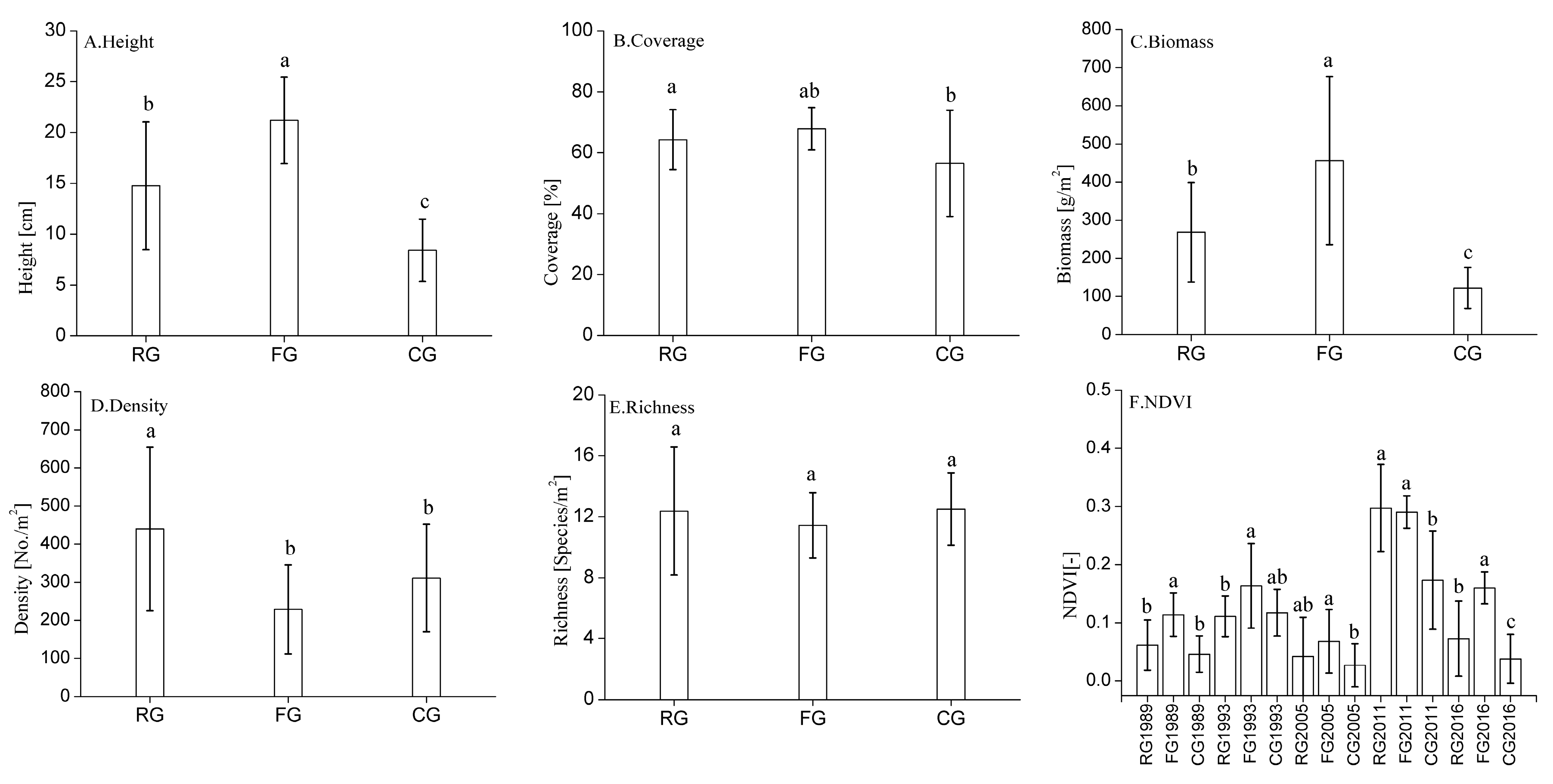
3.1. Changes in Plant Community Characteristics in Response to Different Grazing Systems
3.2. Changes in NDVI Value in Response to Different Grazing Systems
4. Discussion
4.1. Effects of Different Grazing Systems on Plant Community Characteristics
4.2. Effects of Different Grazing Systems on Changes in NDVI Value
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Milchunas, D.G.; Sala, O.E.; Lauenroth, W.K. A generalized model of the effects of grazing by large herbivores on grassland community structure. Am. Nat. 1988, 132, 87–106. [Google Scholar] [CrossRef]
- Briske, D.D.; Derner, J.D.; Brown, J.R.; Fuhlendorf, S.D.; Teague, W.R.; Havstad, K.M.; Gillen, R.L.; Ash, A.J.; Willms, W.D. Rotational grazing on rangelands: Reconciliation of perception and experimental evidence. Rangel. Ecol. Manag. 2008, 61, 3–17. [Google Scholar] [CrossRef]
- Conte, T.J.; Tilt, B. The effects of China’s grassland contract policy on pastoralists' attitudes towards cooperation in an Inner Mongolian banner. Hum. Ecol. 2014, 42, 837–846. [Google Scholar] [CrossRef]
- Han, G.; Hao, X.; Zhao, M.; Wang, M.; Ellert, B.H.; Willms, W.; Wang, M. Effect of grazing intensity on carbon and nitrogen in soil and vegetation in a meadow steppe in Inner Mongolia. Agric. Ecosyst. Environ. 2008, 125, 21–32. [Google Scholar] [CrossRef]
- Heitschmidt, R.K.; Dowhower, S.L.; Walker, J.W. Some effects of a rotational grazing treatment on quantity and quality of available forage and amount of ground litter. J. Range Manag. 1987, 40, 318–321. [Google Scholar] [CrossRef]
- Derner, J.D.; Gillen, R.L.; McCollum, F.T.; Tate, K.W. Little bluestem tiller defoliation patterns under continuous and rotational grazing. J. Range Manag. 1994, 47, 220–225. [Google Scholar] [CrossRef]
- Jacobo, E.J.; Rodríguez, A.M.; Bartoloni, N.; Deregibus, V.A. Rotational grazing effects on rangeland vegetation at a farm scale. Rangel. Ecol. Manag. 2006, 59, 249–257. [Google Scholar] [CrossRef]
- Wang, J.; Brown, D.G.; Chen, J. Drivers of the dynamics in net primary productivity across ecological zones on the mongolian plateau. Landsc. Ecol. 2013, 28, 725–739. [Google Scholar] [CrossRef]
- Hai, S. Study on the Relationship between Human and Land in Inner Mongolia Pasture; Inner Mongolian Education Press: Hohhot, China, 2013; pp. 131–143. [Google Scholar]
- Zhang, J.T. Sampling Methods and Community Characterization, 2nd ed.; Sciences Publisher: Beijing, China, 2011; pp. 9–25. [Google Scholar]
- Hoshino, B.; Yonemori, M.; Manayeva, K.; Karamalla, A.; Yoda, K.; Suliman, M.; Elgamri, M.; Nawata, H.; Mori, Y.; Yabuki, S.; et al. Remote sensing methods for the evaluation of the mesquite tree (Prosopis juliflora) environmental adaptation to semi-arid Africa. In Proceedings of the 2011 IEEE International Geoscience and Remote Sensing Symposium, Vancouver, BC, Canada, 24–29 July 2011; pp. 1910–1913. [Google Scholar]
- Tucker, C.J. Red and photographic infrared linear combinations for monitoring vegetation. Remote Sens. Environ. 1979, 8, 127–150. [Google Scholar] [CrossRef]
- Olff, H.; Ritchie, M.E. Effects of herbivores on grassland plant diversity. Trends Ecol. Evolut. 1998, 13, 261–265. [Google Scholar] [CrossRef]
- Metera, E.; Sakowski, T.; Sloniewski, K.; Romanowicz, B. Grazing as a tool to maintain biodiversity of grassland—A review. Anim. Sci. Pap. Rep. 2010, 28, 315–334. [Google Scholar]
- Huhta, A.-P.; Hellstrom, K.; Rautio, P.; Tuomi, J. Grazing tolerance of gentianella amarella and other monocarpic herbs: Why is tolerance highest at low damage levels? Plant Ecol. 2003, 166, 49–61. [Google Scholar] [CrossRef]
- Carrera, A.L.; Bertiller, M.B.; Larreguy, C. Leaf litterfall, fine-root production, and decomposition in shrublands with different canopy structure induced by grazing in the Patagonian Monte, Argentina. Plant Soil 2008, 311, 39–50. [Google Scholar] [CrossRef]
- Firincioglu, H.K.; Seefeldt, S.S.; Sahin, B. The effects of long-term grazing exclosures on range plants in the central Anatolian region of Turkey. Environ. Manag. 2007, 39, 326–337. [Google Scholar] [CrossRef] [PubMed]
- Deak, B.; Tothmeresz, B.; Valko, O.; Sudnik-Wojcikowska, B.; Moysiyenko, I.I.; Bragina, T.M.; Apostolova, I.; Dembicz, I.; Bykov, N.I.; Torok, P. Cultural monuments and nature conservation: A review of the role of kurgans in the conservation and restoration of steppe vegetation. Biodivers. Conserv. 2016, 25, 2473–2490. [Google Scholar] [CrossRef]
- Brinkert, A.; Hoelzel, N.; Sidorova, T.V.; Kamp, J. Spontaneous steppe restoration on abandoned cropland in Kazakhstan: Grazing affects successional pathways. Biodivers. Conserv. 2016, 25, 2543–2561. [Google Scholar] [CrossRef]
- Grime, J.P. Evidence for the existence of 3 primary strategies in plants and its relevance to ecological and evolutionary theory. Am. Nat. 1977, 111, 1169–1194. [Google Scholar] [CrossRef]
- Collins, S.L.; Bradford, J.A.; Sims, P.L. Succession and fluctuation in Artemisia dominated grassland. Vegetatio 1988, 73, 89–100. [Google Scholar] [CrossRef]
- Loeser, M.R.R.; Sisk, T.D.; Crews, T.E. Impact of grazing intensity during drought in an Arizona grassland. Conserv. Biol. 2007, 21, 87–97. [Google Scholar] [CrossRef] [PubMed]
- Guo, Q.; Berry, W.L. Species richness and biomass: Dissection of the hump-shaped relationships. Ecology 1998, 79, 2555–2559. [Google Scholar] [CrossRef]
- Oesterheld, M.; McNaughton, S.J. Effect of stress and time for recovery on the amount of compensatory growth after grazing. Oecologia (Berlin) 1990, 85, 305–313. [Google Scholar] [CrossRef] [PubMed]
- Bai, Y.; Han, X.; Wu, J.; Chen, Z.; Li, L. Ecosystem stability and compensatory effects in the Inner Mongolia grassland. Nature 2004, 431, 181–184. [Google Scholar] [CrossRef] [PubMed]
- Garcia, C.A.M.; Schellberg, J.; Ewert, F.; Brueser, K.; Canales-Prati, P.; Linstaedter, A.; Oomen, R.J.; Ruppert, J.C.; Perelman, S.B. Response of community-aggregated plant functional traits along grazing gradients: Insights from African semi-arid grasslands. Appl. Veg. Sci. 2014, 17, 470–481. [Google Scholar] [CrossRef]
- Martin, S.C.; Severson, K.E. Vegetation response to the Santa Rita grazing system. J. Range Manag. 1988, 41, 291–295. [Google Scholar] [CrossRef]
- Hao, J.; Dickhoefer, U.; Lin, L.; Mueller, K.; Glindemann, T.; Schoenbach, P.; Schiborra, A.; Wang, C.; Susenbeth, A. Effects of rotational and continuous grazing on herbage quality, feed intake and performance of sheep on a semi-arid grassland steppe. Arch. Anim. Nutr. 2013, 67, 62–76. [Google Scholar] [CrossRef] [PubMed]
- Tilman, D.; Wedin, D.; Knops, J. Productivity and sustainability influenced by biodiversity in grassland ecosystems. Nature 1996, 379, 718–720. [Google Scholar] [CrossRef]
- Sternberg, T.; Tsolmon, R.; Middleton, N.; Thomas, D. Tracking desertification on the Mongolian steppe through NDVI and field-survey data. Int. J. Digit. Earth 2011, 4, 50–64. [Google Scholar] [CrossRef]
- Zhang, M.A.; Borjigin, E.; Zhang, H. Mongolian nomadic culture and ecological culture: On the ecological reconstruction in the agro-pastoral mosaic zone in northern China. Ecol. Econ. 2007, 62, 19–26. [Google Scholar] [CrossRef]
- Michael, H.R.; Kothmann, M.M.; Taylor, C.A. Vegetation response to increased stocking rates in short-duration grazing. J. Range Manag. 1990, 43, 104–108. [Google Scholar]
- Savory, A.; Stanley, D.P. The savory grazing method. Rangelands 1980, 2, 234–237. [Google Scholar]
- Bailey, D.W.; Brown, J.R. Rotational grazing systems and livestock grazing behavior in shrub-dominated semi-arid and arid rangelands. Rangel. Ecol. Manag. 2011, 64, 1–9. [Google Scholar] [CrossRef]
- Heitschmidt, R.K.; Price, D.L.; Gordon, R.A.; Frasure, J.R. Short Duration Grazing at the Texas Experimental Ranch: Effects on Aboveground Net Primary Production and Seasonal Growth Dynamics. J. Range Manag. 1982, 35, 367–372. [Google Scholar] [CrossRef]
- McIntyre, S.; Heard, K.M.; Martin, T.G. The relative importance of cattle grazing in subtropical grasslands: Does it reduce or enhance plant biodiversity? J. Appl. Ecol. 2003, 40, 445–457. [Google Scholar] [CrossRef]
- Ruifrok, J.L.; Postma, F.; Olff, H.; Smit, C. Scale-dependent effects of grazing and topographic heterogeneity on plant species richness in a Dutch salt marsh ecosystem. Appl. Veg. Sci. 2014, 17, 615–624. [Google Scholar] [CrossRef]
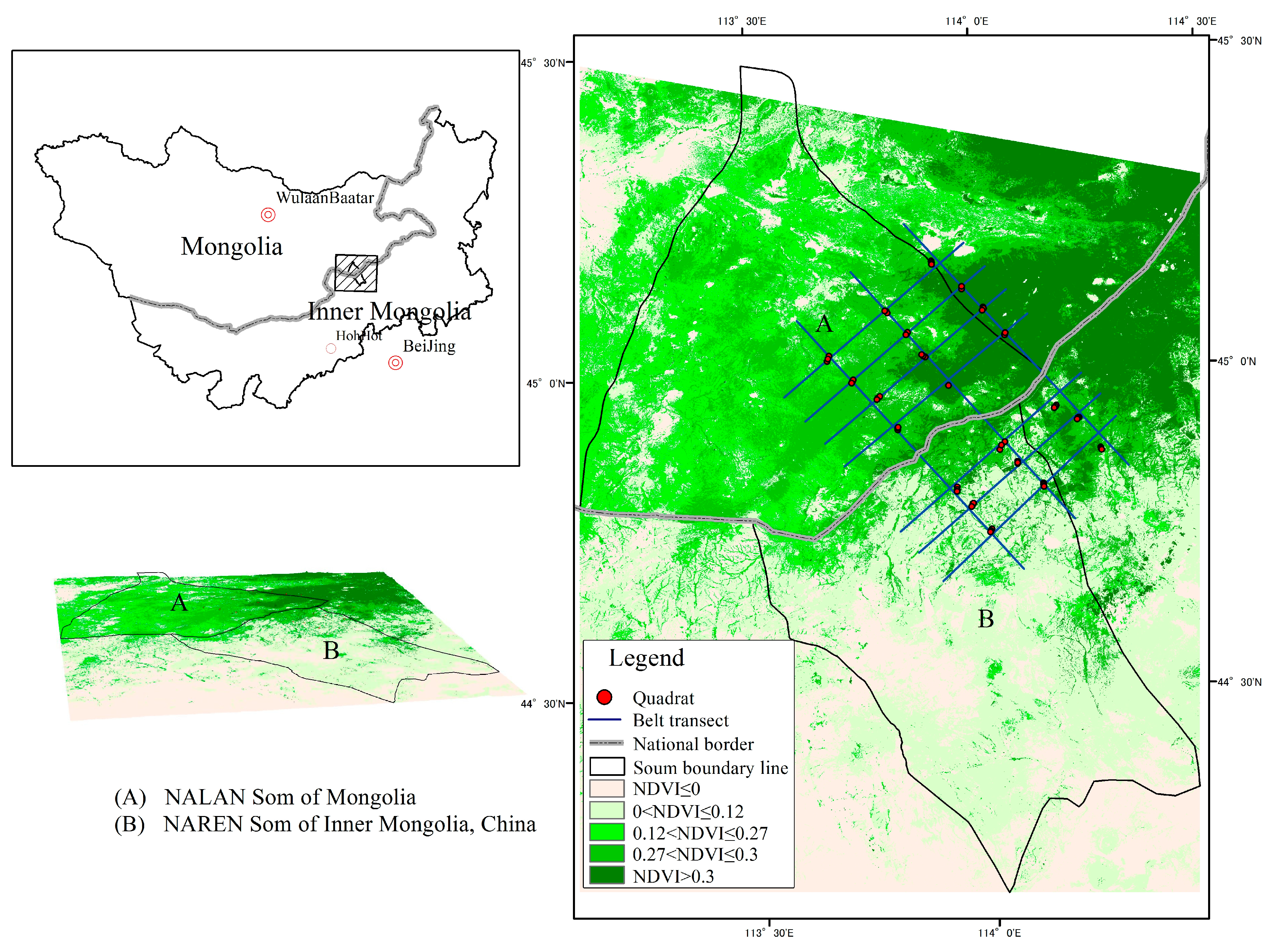
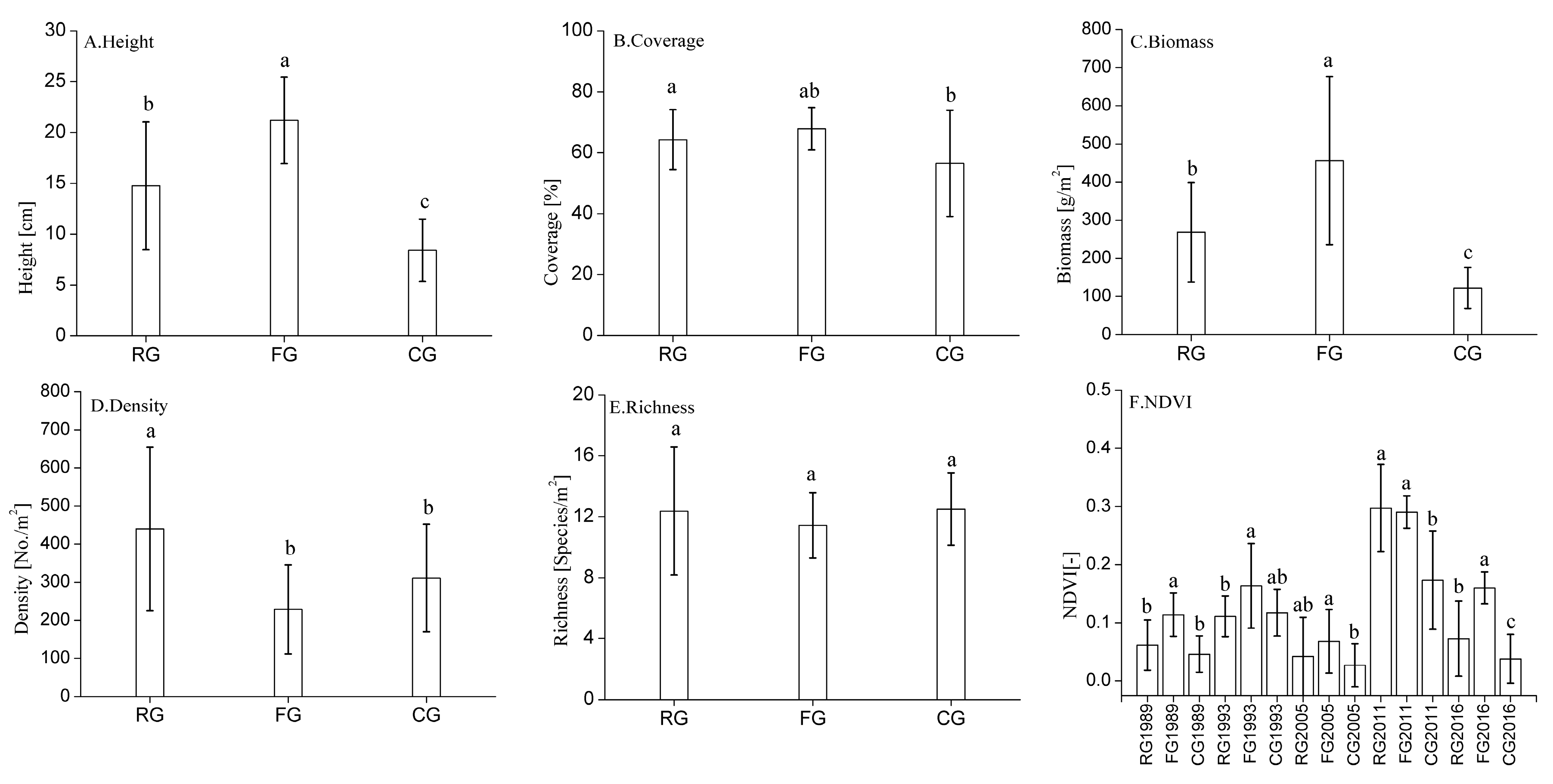

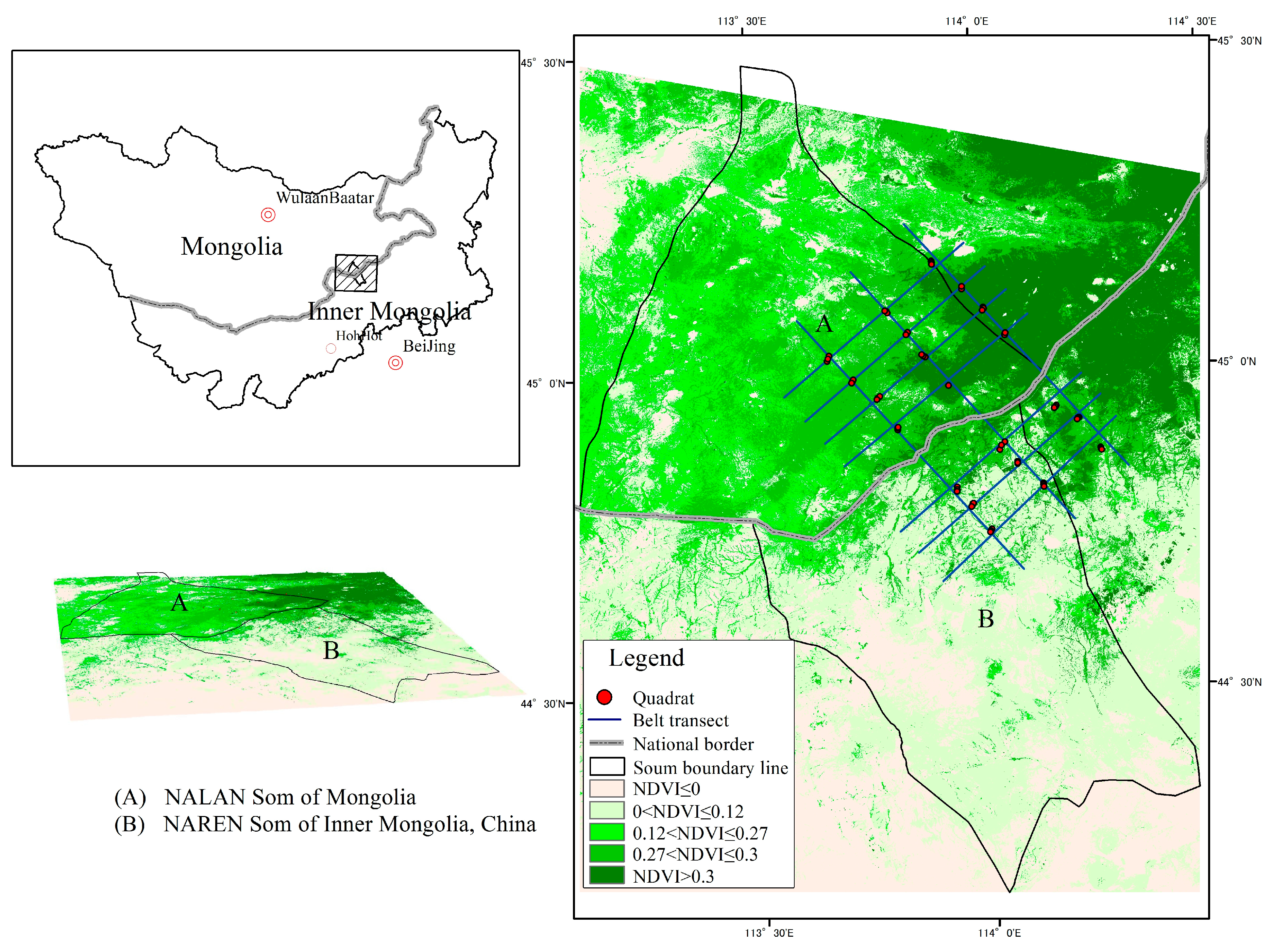{kind=link}
{kind=link}
| Nalan Soum | Naren Soum | |
|---|---|---|
| Annual mean temperature (°C) | 1.41a | 1.03a |
| Annual total precipitation (mm) | 216.80a | 224.49a |
| Altitude (m) | 1356.21a | 1346.39a |
| Average annual evaporation (mm/y) | 1505.14a | 1498.52a |
| Average stocking rate (sheep unit/km2) | 42a | 50a |
| Soil type | Chestnut soil | Chestnut soil |
| Soil humidity (VWC) | 7.5%a | 6.9%b |
| Country | Mongolia | Inner Mongolia | |||
|---|---|---|---|---|---|
| Time | 1958–1990 | 1990–Today | 1978–1990 | 1990–Today | |
| Livestock Ownership | Common | Private | Private | Private | |
| Pasture Ownership | Common | Common | Common | Private | |
| Grazing Method | Rotational Grazing | Rotational Grazing | Rotational Grazing | Continuous Grazing | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Na, Y.; Bao, S.; Hashimoto, K.; McCarthy, C.; Hoshino, B. The Effects of Grazing Systems on Plant Communities in Steppe Lands—A Case Study from Mongolia’s Pastoralists and Inner Mongolian Settlement Areas. Land 2018, 7, 10. https://doi.org/10.3390/land7010010
Na Y, Bao S, Hashimoto K, McCarthy C, Hoshino B. The Effects of Grazing Systems on Plant Communities in Steppe Lands—A Case Study from Mongolia’s Pastoralists and Inner Mongolian Settlement Areas. Land. 2018; 7(1):10. https://doi.org/10.3390/land7010010
Chicago/Turabian StyleNa, Yintai, Saixiyalt Bao, Kanji Hashimoto, Christopher McCarthy, and Buho Hoshino. 2018. "The Effects of Grazing Systems on Plant Communities in Steppe Lands—A Case Study from Mongolia’s Pastoralists and Inner Mongolian Settlement Areas" Land 7, no. 1: 10. https://doi.org/10.3390/land7010010
APA StyleNa, Y., Bao, S., Hashimoto, K., McCarthy, C., & Hoshino, B. (2018). The Effects of Grazing Systems on Plant Communities in Steppe Lands—A Case Study from Mongolia’s Pastoralists and Inner Mongolian Settlement Areas. Land, 7(1), 10. https://doi.org/10.3390/land7010010