Impacts of Logging Road Networks on Dung Beetles and Small Mammals in a Malaysian Production Forest: Implications for Biodiversity Safeguards

Abstract
:1. Introduction
2. Methods
2.1. Study Site
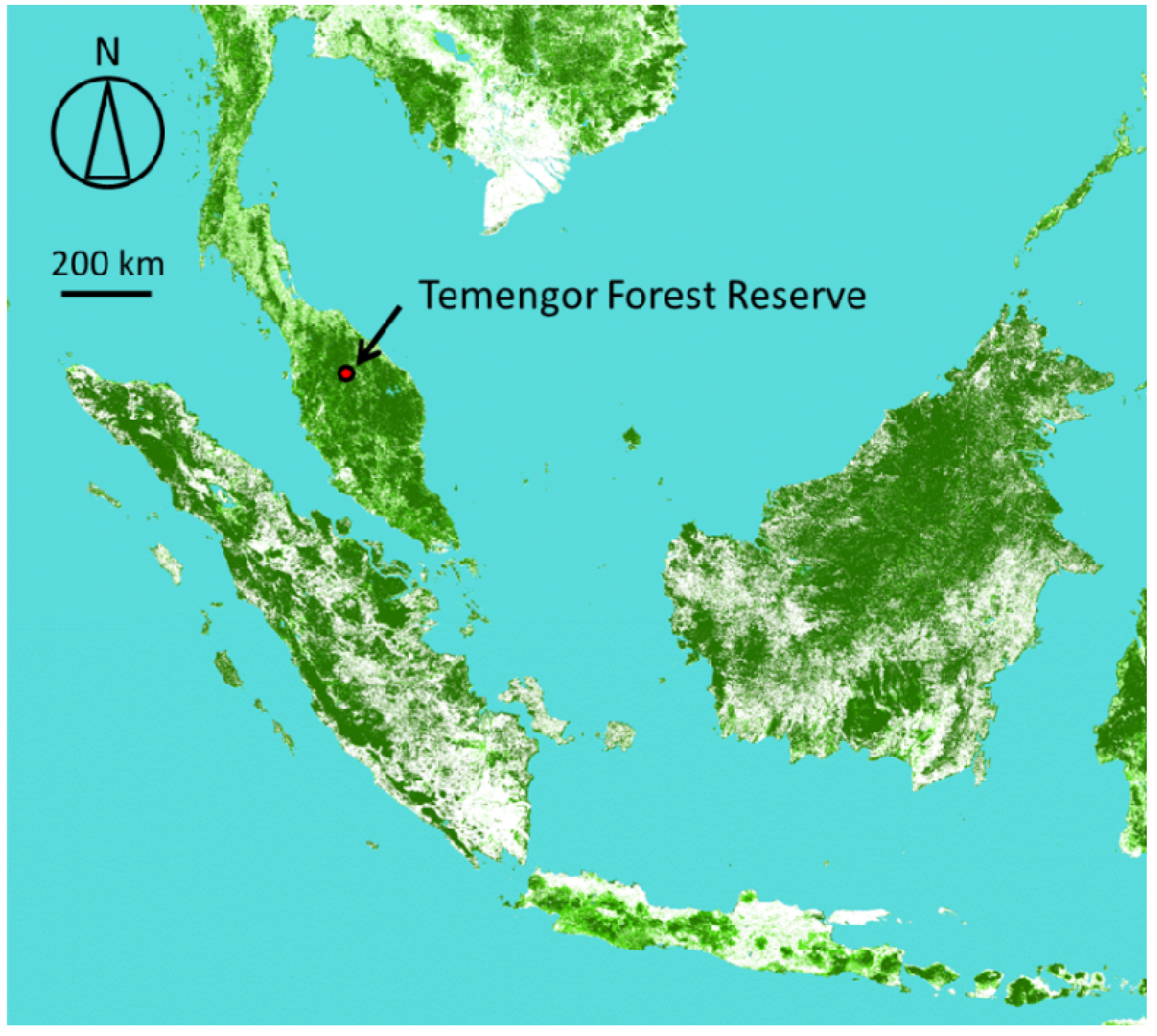
 ) in the Temengor Forest Reserve in northern Perak State, Malaysia, generated from the Global Map from the Geospatial Information Authority of Japan (GSI). The greener parts have the higher percent of tree covers. Our study site (the Temengor Forest Reserve) is indicated by an arrow.
) in the Temengor Forest Reserve in northern Perak State, Malaysia, generated from the Global Map from the Geospatial Information Authority of Japan (GSI). The greener parts have the higher percent of tree covers. Our study site (the Temengor Forest Reserve) is indicated by an arrow.
) in the Temengor Forest Reserve in northern Perak State, Malaysia, generated from the Global Map from the Geospatial Information Authority of Japan (GSI). The greener parts have the higher percent of tree covers. Our study site (the Temengor Forest Reserve) is indicated by an arrow.
) in the Temengor Forest Reserve in northern Perak State, Malaysia, generated from the Global Map from the Geospatial Information Authority of Japan (GSI). The greener parts have the higher percent of tree covers. Our study site (the Temengor Forest Reserve) is indicated by an arrow.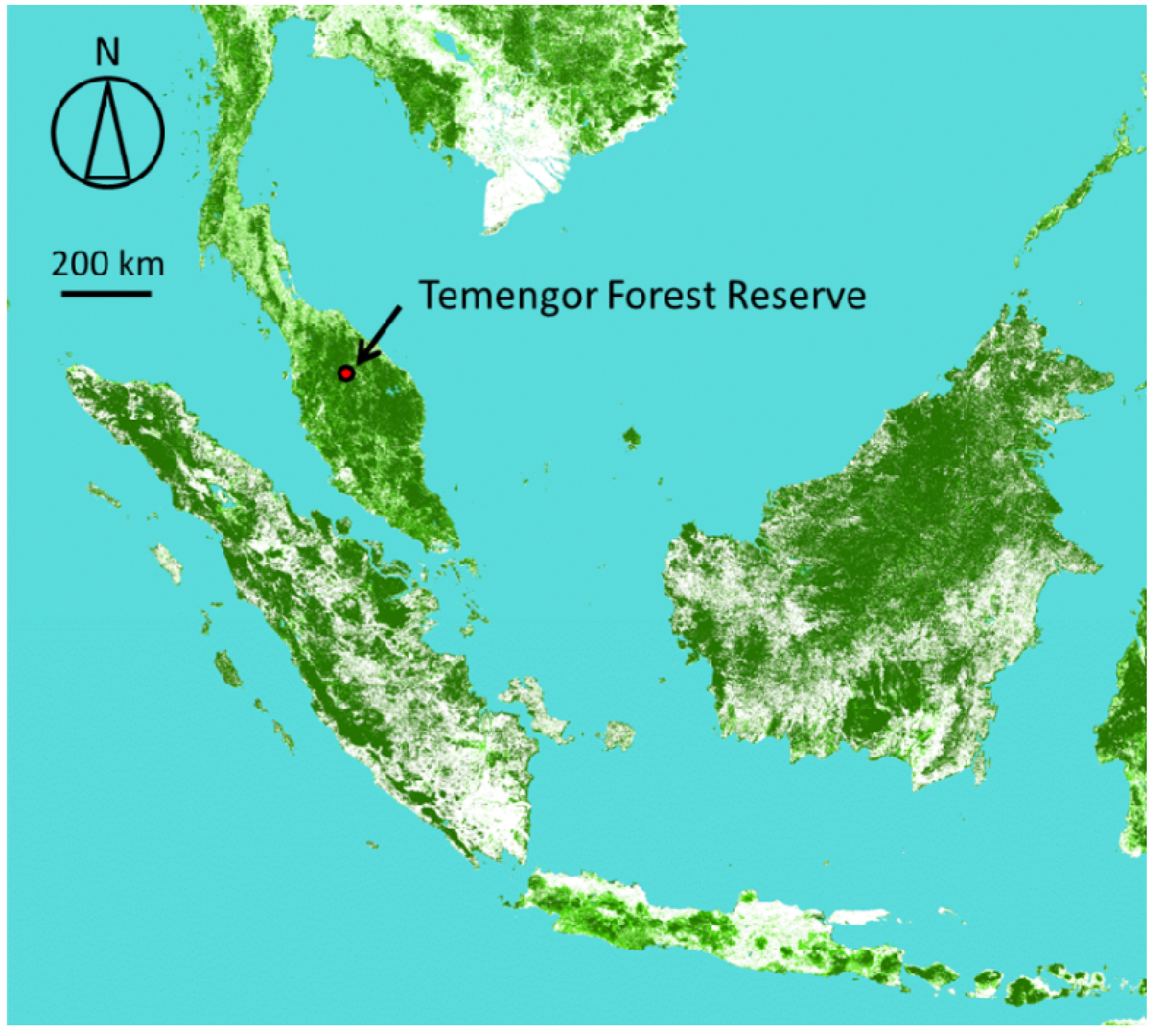
2.2. Field Methods


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Family | Scientific Name | Presence or Absence | |
|---|---|---|---|
| Logged Forest | Unlogged Forest | ||
| Erinaceidae | Hylomys suillus | ○ | ● |
| Soricidae | Crocidura fuliginosa | ○ | ○ |
| Tupaiidae | Tupaiaglis | ○ | ○ |
| Sciuridae | Sundasciuruslowii;Sundasciurustenius | ○ | ○ |
| Lariscus insignis | ○ | ○ | |
| Muridae | Rattusrattus; Rattus tiomanicus | ○ | ● |
| Sundamysmuelleri | ○ | ● | |
| Berylmysbowersi | ○ | ● | |
| Niviventercremoriventer | ○ | ○ | |
| Leopoldamyssabanus | ○ | ○ | |
| Maxomyssurifer; Maxomys whiteheadi | ○ | ○ | |
2.3. Analysis
3. Results
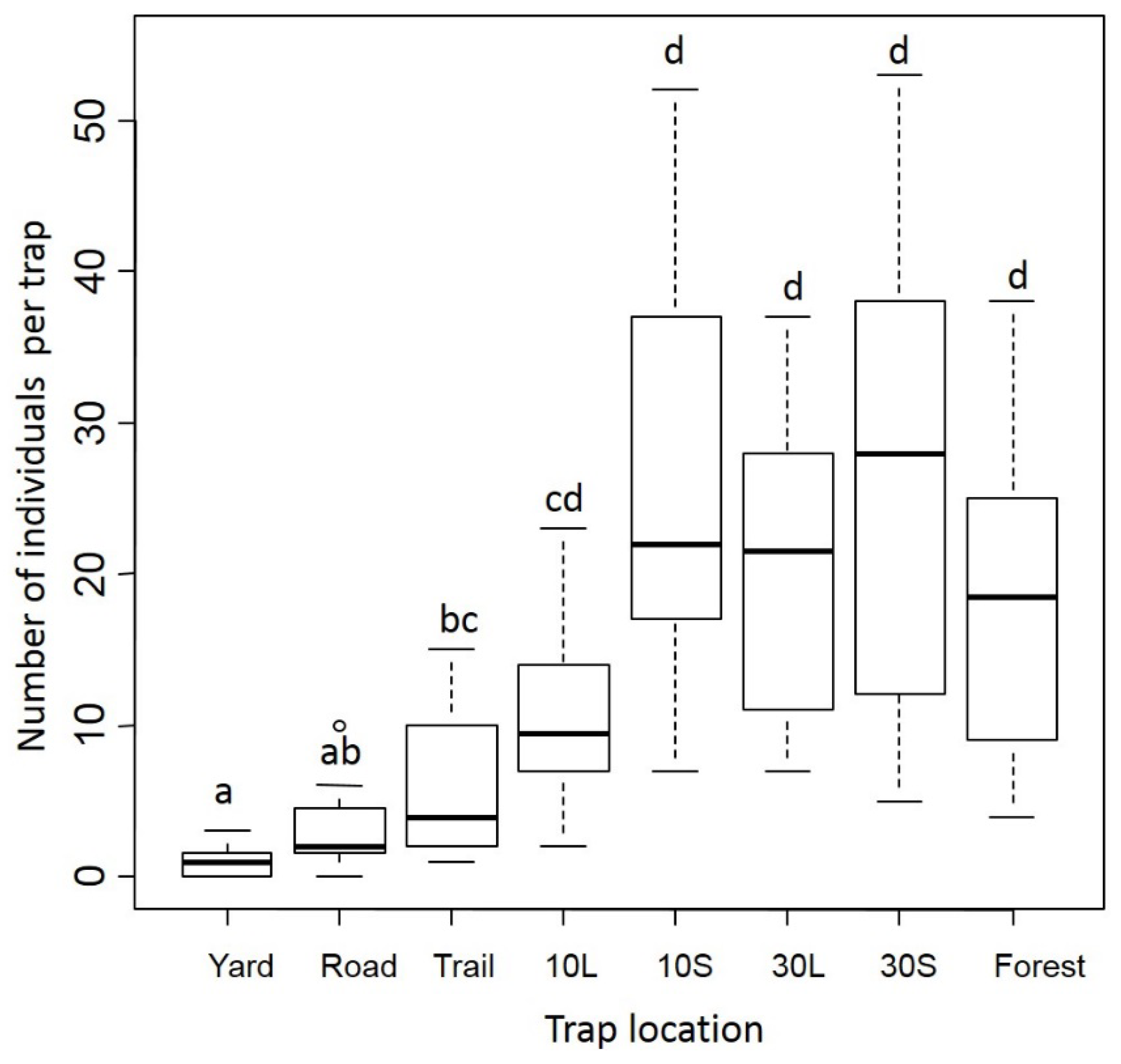
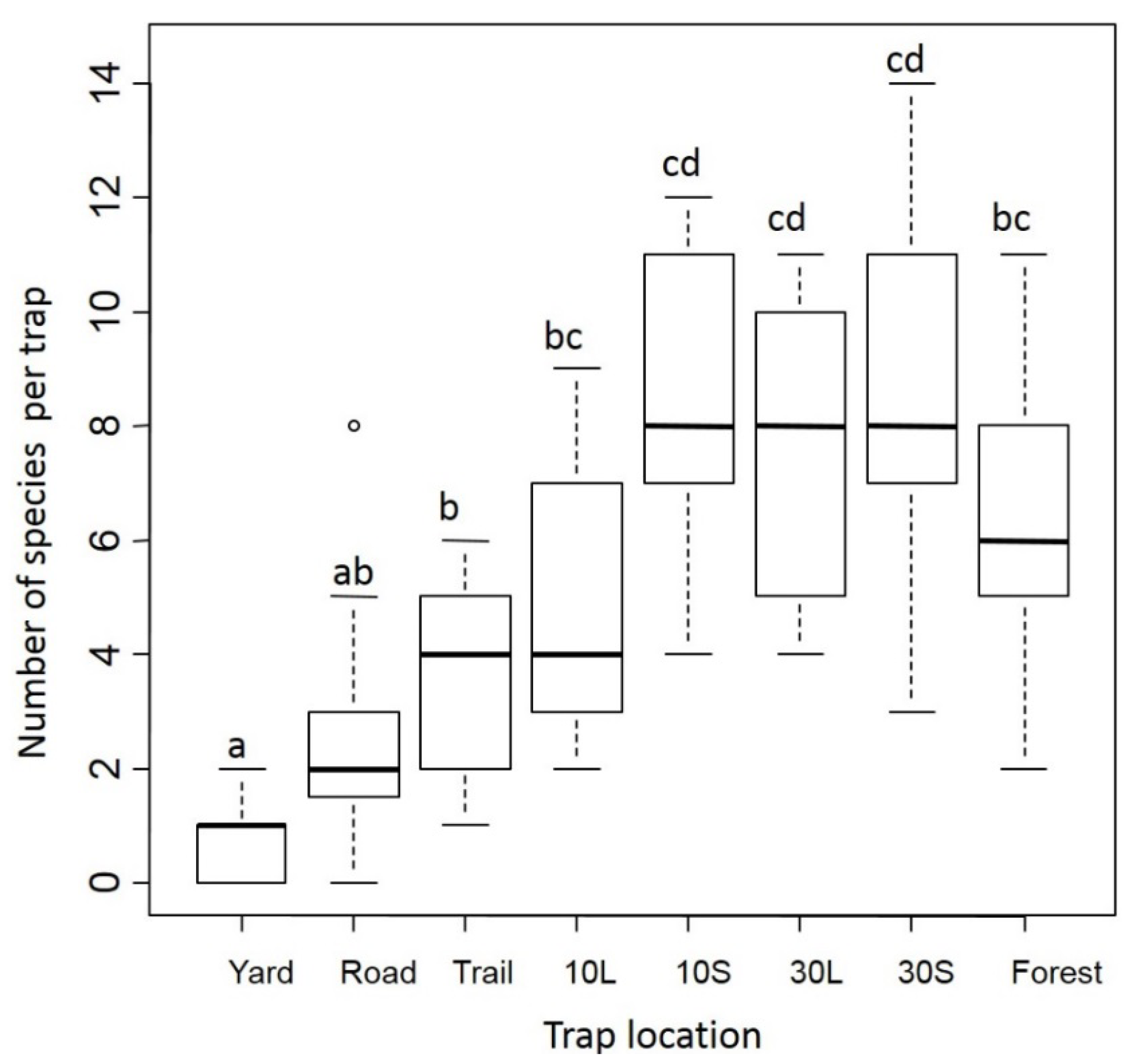
3.1. Dung Beetles
| Species | Presence or Absence | |||||||
|---|---|---|---|---|---|---|---|---|
| Yard | Road | Trail | 10 L | 10 S | 30 L | 30 S | Forest | |
| Caccobius bawangensis | ● | ● | ● | ● | ● | ● | ○ | ● |
| Caccobius unicornis | ● | ● | ● | ● | ● | ○ | ○ | ○ |
| Catharsius renaudpauliani | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ |
| Copris agnus | ● | ● | ○ | ○ | ○ | ○ | ○ | ○ |
| Copris spinator | ● | ● | ● | ● | ○ | ○ | ○ | ● |
| Microcopris doriae | ● | ● | ○ | ○ | ○ | ○ | ○ | ● |
| Microcopris hidakai | ● | ● | ● | ● | ○ | ● | ● | ● |
| Ochicanthon peninsularis | ● | ○ | ○ | ○ | ○ | ○ | ○ | ● |
| O. sp.A | ● | ○ | ● | ● | ○ | ● | ○ | ● |
| O. sp.B | ● | ○ | ● | ○ | ○ | ○ | ○ | ● |
| Onthophagus aphodioides | ● | ● | ○ | ● | ● | ● | ● | ● |
| Onthophagus deliensis | ● | ○ | ○ | ○ | ○ | ○ | ● | ● |
| Onthophagus falculatus | ● | ● | ● | ○ | ● | ● | ● | ● |
| Onthophagus kawaharai | ● | ● | ● | ○ | ● | ● | ● | ● |
| Onthophagus leusermontis | ● | ○ | ○ | ○ | ○ | ○ | ○ | ○ |
| Onthophagus liliputanus | ○ | ○ | ○ | ● | ● | ● | ● | ● |
| Onthophagus mentaveiensis | ● | ● | ● | ● | ○ | ● | ● | ● |
| Onthophagus nigriobscurior | ● | ● | ● | ● | ○ | ● | ● | ● |
| Onthophagus obscurior | ● | ○ | ○ | ○ | ○ | ○ | ○ | ● |
| Onthophagus orientalis | ● | ○ | ● | ● | ● | ● | ● | ● |
| Onthophagus roralius | ● | ● | ● | ● | ● | ○ | ○ | ● |
| Onthophagus roubali | ● | ○ | ● | ● | ● | ● | ● | ● |
| Onthophagus rudis | ● | ● | ○ | ○ | ● | ○ | ○ | ● |
| Onthophagus rugicollis | ● | ○ | ● | ● | ● | ○ | ● | ● |
| Onthophagus rutilans | ● | ● | ● | ○ | ○ | ○ | ○ | ● |
| Onthophagus semifex | ● | ● | ● | ● | ● | ○ | ● | ● |
| Onthophagus sepilokensis | ● | ○ | ○ | ○ | ● | ● | ● | ○ |
| Onthophagus ulugombakensis | ● | ○ | ● | ● | ○ | ○ | ● | ● |
| Onthophagus tsubakii | ● | ● | ○ | ● | ● | ○ | ● | ● |
| Onthophagus vethi | ● | ● | ○ | ● | ● | ● | ● | ● |
| Onthophagus viridicervicapra | ● | ● | ○ | ● | ○ | ● | ○ | ● |
| Onthophagus vulpes | ● | ○ | ○ | ○ | ○ | ○ | ○ | ○ |
| O. sp. A | ● | ● | ○ | ● | ○ | ● | ● | ● |
| O. sp. B | ● | ○ | ○ | ● | ● | ○ | ○ | ● |
| Paracopris ramosiceps | ● | ● | ● | ● | ● | ○ | ○ | ○ |
| Paragymnopleurus maurus | ● | ● | ● | ● | ○ | ○ | ○ | ● |
| Paragymnopleurus striatus | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ |
| Sisyphus thoracicus | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ |
| Synapsis ritsemae | ● | ● | ● | ● | ○ | ○ | ○ | ○ |
| Synapsis roslihashimi | ● | ● | ● | ● | ○ | ● | ○ | ● |
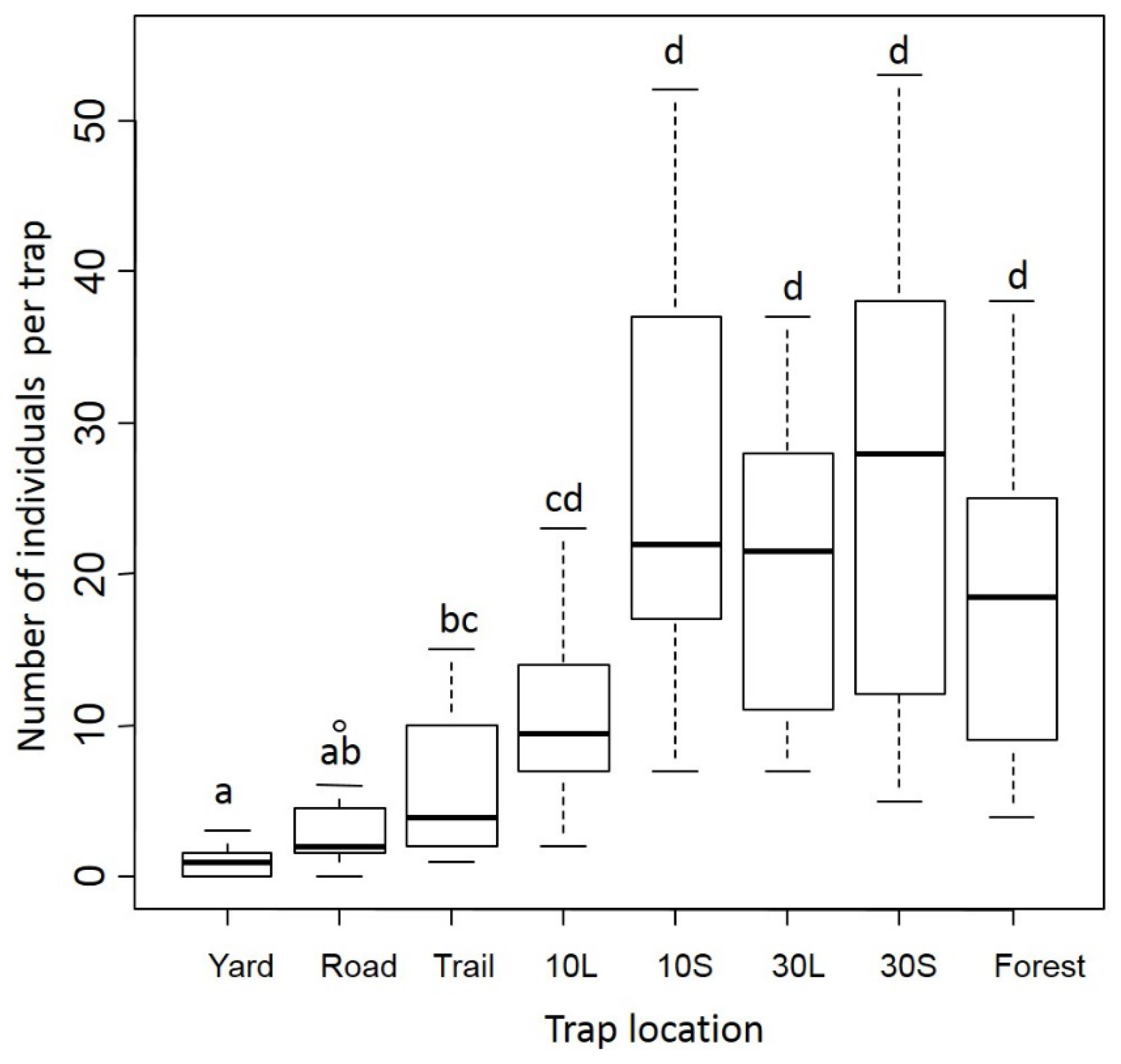
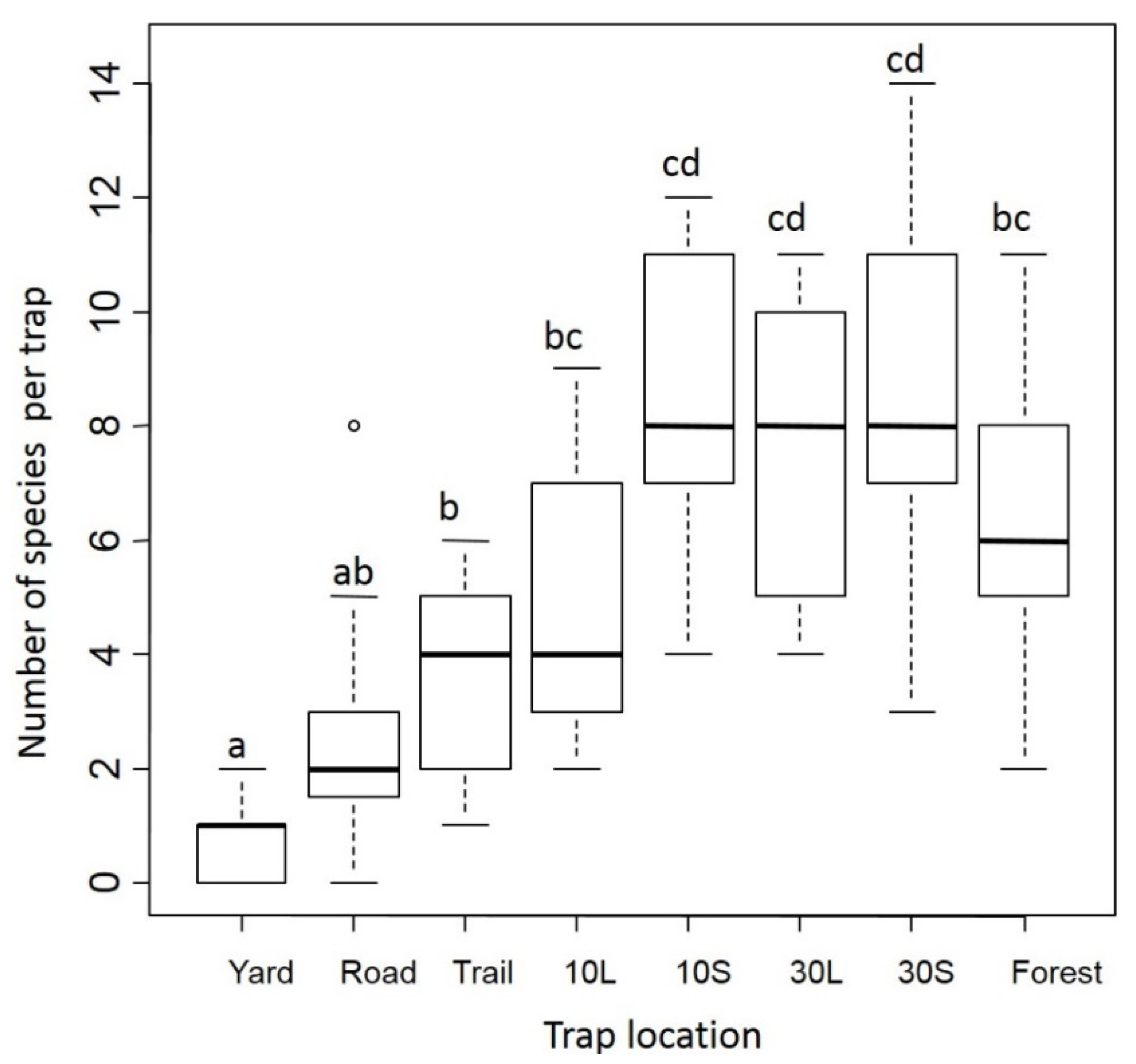
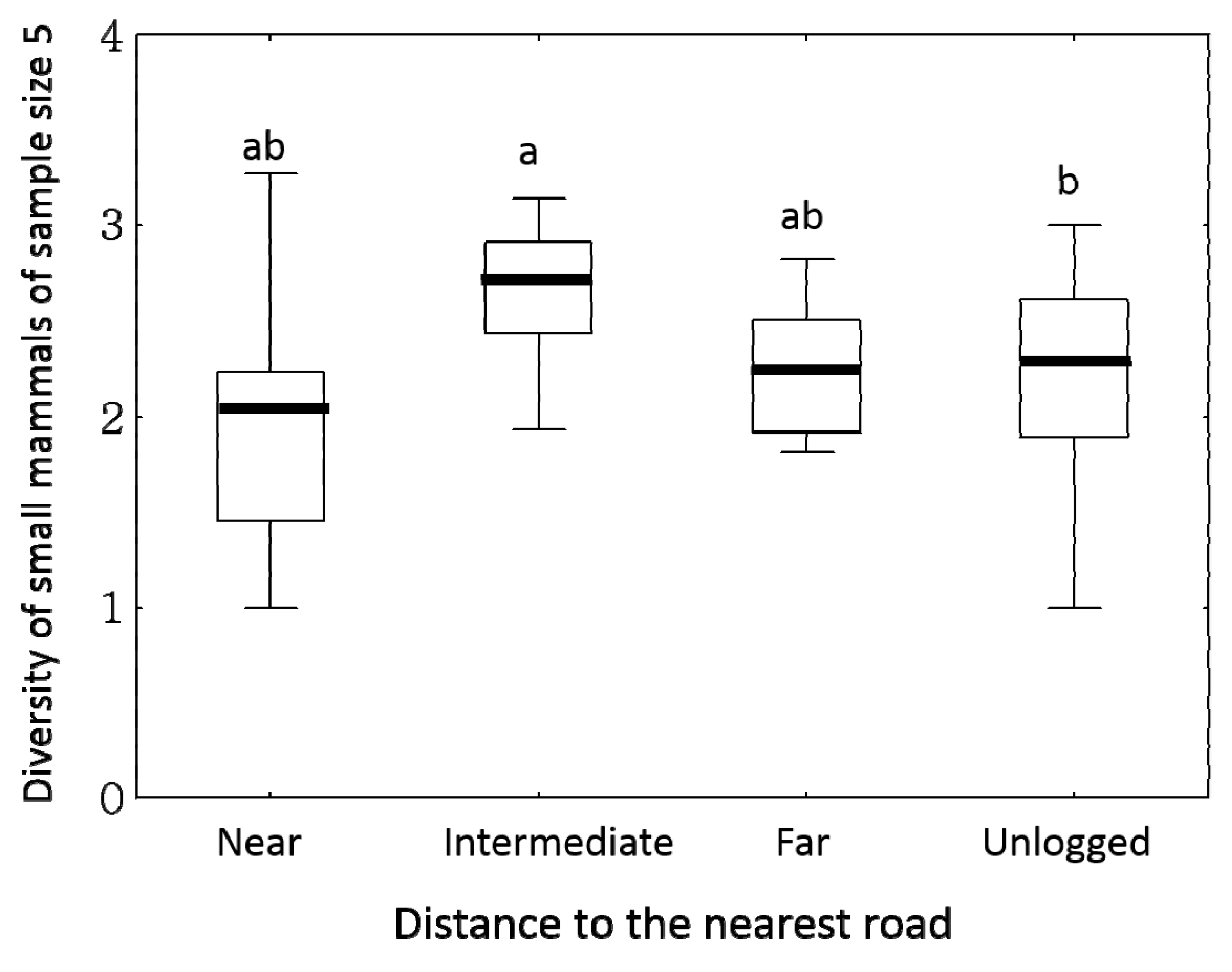
3.2. Small Mammals
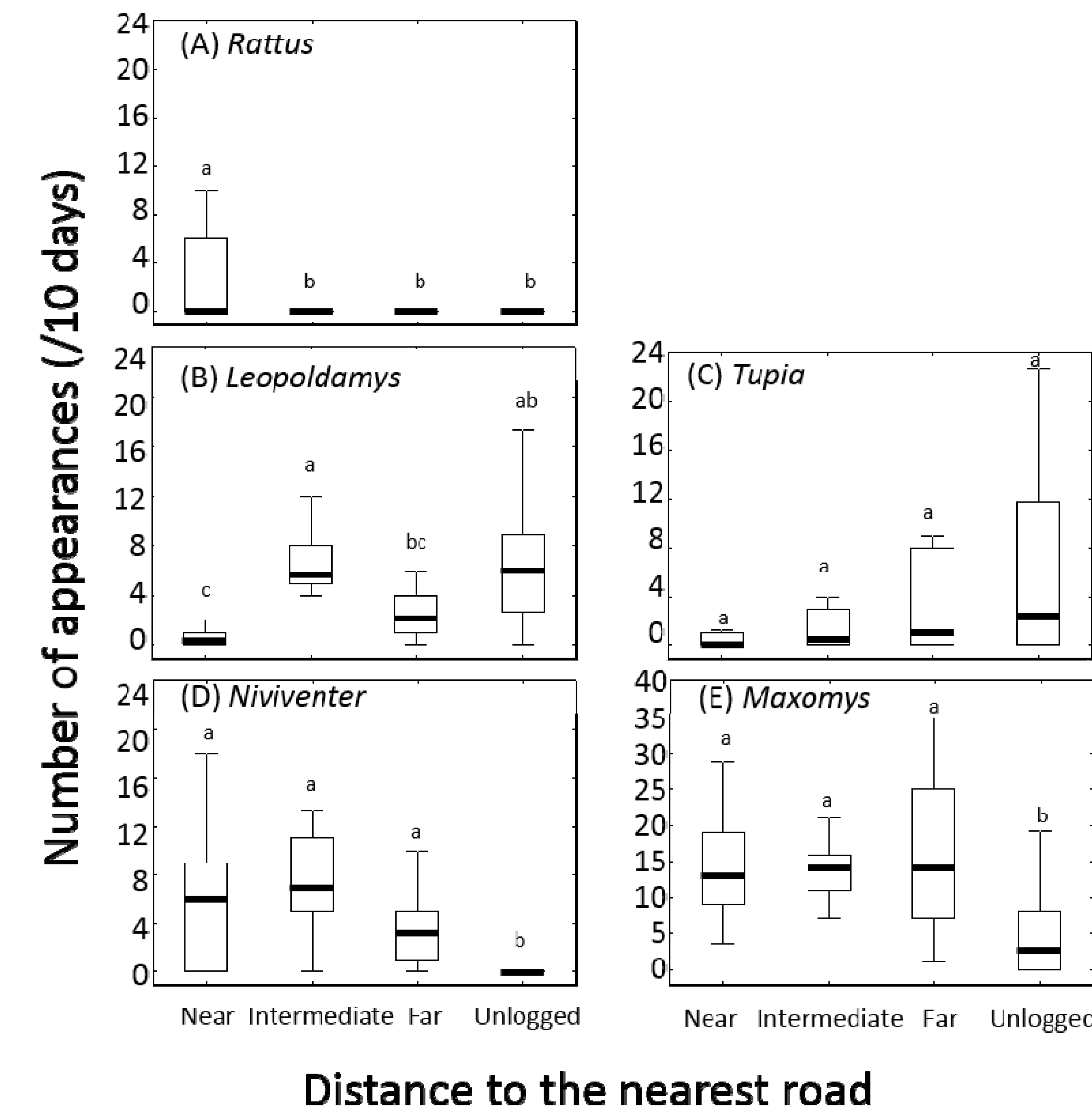

4. Discussion
4.1. Impacts of Logging Roads on Dung Beetles and Small Mammals
4.2. A Guideline to Safeguarding Biodiversity in Malaysian Production Forests
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Sodhi, N.S.; Koh, L.P.; Brook, B.W.; Ng, P.K.L. Southeast Asian biodiversity: An impending disaster. Trends Ecol. Evol. 2004, 19, 654–660. [Google Scholar] [CrossRef]
- Sist, P.; Garcia-Fernandez, C.; Fredericksen, D. Moving beyond reduced-impact logging towards a more holistic management of tropical forests. For. Ecol. Manage. 2008, 256, 7–11. [Google Scholar]
- Didham, R.K. Life after logging: Strategic withdrawal from the Garden of Eden or tactical error for wilderness conservation? Biotropica 2011, 43, 393–395. [Google Scholar] [CrossRef]
- Food and Agriculture Organization-United Nations (FAO). Global Forest Resources Assessment 2010; Main Report-FAO Forestry Paper 163 (FRA2010); FAO: Rome, Italy, 2010. [Google Scholar]
- Blaser, J.; Sarre, A.; Poore, D.; Johnson, S. Status of Tropical Forest Management; ITTO Technical Series No. 38; International Tropical Timber Organization: Yokohama, Japan, 2011. [Google Scholar]
- Edwards, D.P.; Larsen, T.H.; Docherty, T.D.S.; Ansell, F.A.; Hsu, W.W.; Derhé, A.M.; Hamer, K.C.; Wilcove, D.S. Degraded lands worth protecting: The biological importance of Southeast Asia’s repeatedly logged forests. Proc. R. Soc. B 2011, 278, 82–90. [Google Scholar] [CrossRef]
- Bischoff, W.; Newbery, D.M.; Lingenfelder, M.; Schnaeckel, R.; Hubert Petol, G.; Madani, L.; Colin, E.R. Secondary succession and dipterocarp recruitment in Bornean rain forest after logging. For. Ecol. Manag. 2005, 218, 174–192. [Google Scholar] [CrossRef]
- Okuda, T.; Suzuki, M.; Adachi, N.; Quah, E.S.; Hussein, N.A.; Manokaran, N. Effect of selective logging on canopy and stand structure and tree species composition in a lowland dipterocarp forest in Peninsular Malaysia. For. Ecol. Manag. 2003, 175, 297–320. [Google Scholar] [CrossRef]
- Sist, P.; Gourlet-Fleury, S.; Putz, F.E. The impacts of selective logging: Questionable conclusions. BioScience 2012, 62, 786. [Google Scholar]
- Zimmerman, B.L.; Kormos, C.F. Prospects for sustainable logging in tropical forests. BioScience 2012, 62, 479–487. [Google Scholar] [CrossRef]
- Pimm, S.L.; Raven, P. Biodiversity: Extinction by numbers. Nature 2000, 403, 843–845. [Google Scholar] [CrossRef]
- Gibson, L.; Lee, T.M.; Koh, L.P.; Brook, B.W.; Gardner, T.A.; Barlow, J.; Peres, C.A.; Bradshaw, C.J.A.; Laurance, W.F.; Lovejoy, T.E.; et al. Primary forests are irreplaceable for sustaining tropical biodiversity. Nature 2011, 478, 378–383. [Google Scholar] [CrossRef]
- Putz, F.E.; Putz, F.E.; Zuidema, P.A.; Synnott, T.; Peña-Claros, M.; Pinard, M.A.; Sheil, D.; Vanclay, J.K.; Sist, P.; Gourlet-Fleury, S.; et al. Sustaining conservation values in selectively logged tropical forests: The attained and the attainable. Conserv. Lett. 2012, 5, 296–303. [Google Scholar] [CrossRef]
- Slade, E.M.; Mann, D.J.; Lewis, O.T. Biodiversity and ecosystem function of tropical forest dung beetles under contrasting logging regimes. Conserv. Biol. 2011, 144, 166–174. [Google Scholar] [CrossRef]
- Oxley, D.J.; Fenton, M.B.; Carmody, G.R. The effects of roads on populations of small mammals. J. Appl. Ecol. 1974, 1, 51–59. [Google Scholar]
- Adams, L.W.; Geis, A.D. Effects of roads on small mammals. J. Appl. Ecol. 1983, 20, 403–415. [Google Scholar] [CrossRef]
- Forman, R.T.T; Deblinger, R.D. The ecological road-effect zone of a Massachusetts (USA) suburban highway. Conserv. Biol. 2000, 14, 36–46. [Google Scholar] [CrossRef]
- Gullison, R.E.; Hardner, J.J. The effects of road design and harvest intensity on forest damage caused by selective logging: Empirical results and a simulation model from the Bosque Chimanes, Bolivia. For. Ecol. Manag. 1993, 59, 1–14. [Google Scholar]
- Malcolm, J.R.; Ray, J.C. Influence of timber extraction routes on central African small-mammal communities, forest structure, and tree diversity. Conserv. Biol. 2000, 14, 1623–1638. [Google Scholar] [CrossRef]
- Jackson, S.M.; Frederichsen, T.S.; Malcolm, J.R. Area disturbed and residual stand damage following logging in a Bolivian tropical forest. For. Ecol. Manag. 2002, 166, 271–283. [Google Scholar] [CrossRef]
- Putz, F.E.; Sist, P.; Fredericksen, T.; Dykstra, D. Reduced-impact logging: Challenges and opportunities. For. Ecol. Manag. 2008, 256, 1427–1433. [Google Scholar] [CrossRef]
- Pinard, M.A.; Barker, M.G.; Tay, J. Soil disturbance and post-logging forest recovery on bulldozer paths in Sabah, Malaysia. For. Ecol. Manag. 2000, 130, 213–225. [Google Scholar] [CrossRef]
- Uhl, C.; Vieira, I.C.G. Impacts of selective logging in the Brazilian Amazon: A case study from the Paragominas region of the state of Para. Biotropica 1989, 21, 98–106. [Google Scholar] [CrossRef]
- UN-REDD Programme. UN-REDD programme social and environmental principles and criteria. In Proceedings of the UN-REDD Programme Sixth Policy Board Meeting, Asunción, Paraguay, 25–26 March 2012.
- International Secretariat of the REDD+ SES Initiative. REDD+ Social & Environmental Standards (REDD+ SES), version 2 (10 September 2012); Available online: http://www.redd-standards.org/files/REDDSES_Version_2/REDDSES_Version_2_-_10_September_2012.pdf (accessed on 14 February 2014).
- The Climate, Community & Biodiversity Alliance (CCBA). Climate, Community & Biodiversity Standards, 3rd ed.; CCBA: Arlington, VA, USA, 2013. [Google Scholar]
- Thang, H.C.; Chapell, N.A. Minimizing the hydrological impact of forest harvesting in Malaysia’s rain forests. In Forests, Water and People in the Humid Tropics; Bonell, M., Bruijnzeel, L.A., Eds.; Cambridge University Press: Cambridge, MA, USA, 2005; pp. 852–865. [Google Scholar]
- Gardner, T.A.; Barlow, J.; Araujo, I.S.; Avila-Piresm, T.C.; Bonaldo, A.B.; Costa, J.E.; Esposito, M.C.; Ferreira, L.V.; Hawes, J.; Hernandez, M.I.; et al. The cost-effectiveness of biodiversity surveys in tropical forests. Ecol. Lett. 2008, 11, 139–150. [Google Scholar] [CrossRef]
- Nichols, E.; Spector, S.; Louzada, J.; Larsen, T.; Amezquita, S.; Favila, M.E. The Scarabaeinae Research Network—Ecological functions and ecosystem services provided by Scarabaeinae dung beetles. Biol. Conserv. 2008, 141, 1461–1474. [Google Scholar] [CrossRef]
- Andresen, E.; Feer, F. The role of dung beetles as secondary seed dispersers and their effect on plant regeneration in tropical rainforests. In Seed Fate: Predation, Dispersal and Seedling Establishment; Forget, P.M., Lambert, J.E., Hulme, P.E., vander Wall, S.B., Eds.; CAB International: Wallingford, UK, 2005; pp. 331–349. [Google Scholar]
- Jordano, P. Fruits and frugivory. In Seeds: The Ecology of Regeneration in Plant Communities; Fenner, M., Ed.; CAB International: Oxon, UK, 2000; pp. 125–165. [Google Scholar]
- Andresen, E. Seed dispersal by monkeys and the fate of dispersed seeds in a Peruvian rain forest. Biotropica 1999, 311, 45–158. [Google Scholar]
- Andresen, E. Effects of dung presence, dung amount and secondary seed dispersal by dung beetles on the fate of Micropholis guyanensis (Sapotaceae) seeds in Central Amazonia. J. Trop. Ecol. 2001, 17, 61–78. [Google Scholar] [CrossRef]
- Andresen, E.; Levey, D.J. Effects of dung and seed size on secondary seed dispersal, seed predation, and seedling establishment of rain forest trees. Oecologia 2004, 139, 45–54. [Google Scholar] [CrossRef]
- Beaune, D.; Bollache, L.; Bretagnolle, F.; Fruth, B. Dung beetles are critical in preventing post-dispersal seed removal by rodents in Congo rain forest. J. Trop. Ecol. 2012, 28, 507–510. [Google Scholar] [CrossRef]
- Estrada, A.; Coates-Estrada, R. Howler monkeys (Alouatta palliata), dung beetles (Scarabaeidae) and seed dispersal: Ecological interactions in the tropical rain forest of Los Tuxtlas, Mexico. J. Trop. Ecol. 1991, 7, 459–474. [Google Scholar] [CrossRef]
- Shepherd, V.E.; Chapman, C.A. Dung beetles as secondary seed dispersers: Impact on seed predation and germination. J. Trop. Ecol. 1998, 14, 199–215. [Google Scholar] [CrossRef]
- Ratnam, L.; Lim, B.L.; Hussein, N.A. Mammals of the Sungai Singgor area in Temengor Forest Reserve, Hulu Perak, Malaysia. Mal. Nat. J. 1995, 48, 409–423. [Google Scholar]
- Ghazoul, J.; Sheil, D. Tropical Rain Forest Ecology, Diversity, and Conservation; Oxford University Press: Oxford, UK, 2010. [Google Scholar]
- Forget, P.-M.; vander Wall, S.B. Scatter-hoarding rodents and marsupials: Convergent evolution on diverging continents. Trends Ecol. Evol. 2001, 16, 65–67. [Google Scholar] [CrossRef]
- Carpio, C.; Donoso, D.A.; Ramon, G.; Dangles, O. Short term response of dung beetle communities to disturbance by road construction in the Ecuadorian Amazon. Ann. Soc. Entomol. Fr. 2009, 45, 455–469. [Google Scholar]
- The Perak Integrated Timber Complex (PITC). Second Forest Management Plan Perak ITC Concession Area Temengor Forest Reserve 2010–2020; PITC: Perak, Malaysia, 2010. Available online: http://www.perakitc.com.my/index.php?ch=7&pg=18&ac=2&bb=50 (accessed on 15 July 2012).
- Kaur, R.; Ong, T.; Lim, K.C.; Yeap, C.A. A survey on mass movements of the vulnerable plain-pouched hornbill in the Belum-Temengor forest complex, peninsular Malaysia. Raffles Bull. Zool. 2011, 24, 171–176. [Google Scholar]
- Hosaka, T.; Niino, M.; Kon, M.; Okuda, T. Effects of logging road networks on the ecological functions of dung beetles in Peninsular Malaysia. For. Ecol. Manag. 2014, 326, 18–24. [Google Scholar] [CrossRef]
- Larsen, T.H.; Forsyth, A. Trap spacing and transect design for dung beetle biodiversity studies. Biotropica 2005, 37, 322–325. [Google Scholar] [CrossRef]
- Chao, A. Species richness estimation. In Encyclopedia of Statistical Sciences; Balakrishnan, N., Read, C.B., Vidakovic, B., Eds.; Wiley: New York, NY, USA, 2005; pp. 7909–7916. [Google Scholar]
- Colwell, R.K. EstimateS: Statistical Estimation of Species Richness and Shared Species from Samples. User’s Guide and Application, Version 8.2. Available online: http://purl.oclc.org/estimates (accessed on 1 November 2011).
- Karanth, K.U. Estimating tiger Panthera tigris populations from camera-trap data using capturerecapture models. Biol. Conserv. 1995, 71, 139–151. [Google Scholar] [CrossRef]
- Hayes, R.A.; Nahrung, H.F.; Wilson, J.C. The response of native Australian rodents to predator odours varies seasonally: A by-product of life history variation? For. Ecol. Manag. 2006, 71, 1307–1314. [Google Scholar]
- Yasuda, M. Monitoring diversity and abundance of mammals with camera traps: A case study on Mount Tsukuba, central. Japan. Mammal Study 2004, 29, 37–46. [Google Scholar] [CrossRef]
- Francis, C.M. A Field Guide to the Mammals of South-East Asia; New Holland Publishers: London, UK, 2008. [Google Scholar]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2012. [Google Scholar]
- Holm, S. A simple sequentially rejective multiple test procedure. Scand. J. Stat. 1979, 6, 65–70. [Google Scholar]
- Hurlbert, S.H. The nonconcept of species diversity: A critique and alternative parameters. Ecology 1971, 52, 577–586. [Google Scholar] [CrossRef]
- Heck, K.L.; van Belle, G.; Simberloff, D. Explicit calculation of the rarefaction diversity measurement and the determination of sufficient sample size. Ecology 1975, 56, 1459–1461. [Google Scholar] [CrossRef]
- Hosaka, T.; Tokyo Metoripolitan University, Hachioji, Japan. A generalized linear model evaluating the effects of environmental variables on the abundance, species number of dung beetles showed that canopy openness was the leading environmental factor affecting them. 2014. [Google Scholar]
- Laurance, W.F.; Goosem, M.; Laurance, S.G.W. Impacts of roads and linear clearings on tropical forests. Trends Ecol. Evol. 2009, 24, 659–669. [Google Scholar]
- Navarrete, D.; Halffter, G. Dung beetle (Coleoptera: Scarabaeidae: Scarabaeinae) diversity in continuous forest, forest fragments and cattle pastures in a landscape of Chiapas, Mexico: The effects of anthropogenic changes. Biodivers. Conserv. 2008, 17, 2869–2898. [Google Scholar] [CrossRef]
- Peyras, M.; Vespa, N.I.; Bellocq, M.I.; Zurita, G.A. Quantifying edge effects: The role of habitat contrast and species specialization. J. Insect Conserv. 2013, 17, 807–820. [Google Scholar] [CrossRef]
- Kudavidanage, E.P.; Qie, L.; Lee, J.S.H. Linking biodiversity and ecosystem functioning of dung beetles in South and Southeast Asian tropical rainforests. Raffles B. Zool. 2012, Supplement 25, 141–154. [Google Scholar]
- Wolf, M.; Batzil, G.O. Effects of forest edge on populations of white-footed mice (Peromyscus leucopus). Ecology 2002, 25, 193–199. [Google Scholar]
- Gibson, L.; Lynam, A.J.; Bradshaw, C.J.A.; He, F.; Bickford, D.P.; Woodruff, D.S.; Bumrungsri, S.; Laurance, W.F. Near-complete extinction of native small mammal fauna 25 years after forest fragmentation. Science 2013, 341, 1508–1510. [Google Scholar] [CrossRef]
- Laurance, W.F.; Lovejoy, T.E.; Vasconcelos, H.L.; Bruna, E.M.; Didham, R.K.; Stouffer, P.C.; Gascon, C.; Bierregaard, R.O.; Laurance, S.G.; Sampaio, E. Ecosystem decay of Amazonian forest fragments: A 22-yr investigation. Conserv. Biol. 2002, 16, 605–618. [Google Scholar] [CrossRef]
- Laurance, W.F.; Croes, B.M.; Tchignoumba, L.; Lahm, S.A.; Alonso, A.; Lee, M.E.; Campbell, P.; Ondzeano, C. Impacts of roads and hunting on central African rainforest mammals. Conserv. Biol. 2006, 20, 1251–1261. [Google Scholar] [CrossRef]
- Pinard, M.A.; Putz, F.E.; Tay, J.; Sullivan, T.E. Creating timber harvest guidelines for a reduced-impact logging project in Malaysia. J. For. 1995, 93, 41–45. [Google Scholar]
- Miller, J.H.; Sirois, D.L. Soil disturbance by skyline yarding vs. skidding in a loamy hill forest. Soil Sci. Soc. Am. J. 1986, 50, 1579–1583. [Google Scholar] [CrossRef]
- Blakeney, K.J. Environmentally friendly helicopter logging in Papua New Guinea. In Proceedings of the Symposium on Harvesting and Silviculture for Sustainable Forestry in the Tropics, Kuala Lumpur, Malaysia, 5–9 October 1992; Mohd, W.R.W., Ibrahim, S., Appanah, S., Rashid, M.F.A., Eds.; Forest Research Institute Malaysia: Kepong, Malaysia, 1992; pp. 145–150. [Google Scholar]
- Hirata, Y.; Takao, G.; Sato, T.; Toriyama, J. REDD-plus Cookbook; REDD Research and Development Center, Forestry and Forest Products Research Institute: Tsukuba, Japan, 2012. [Google Scholar]
- Jusoff, K.; D’Souza, G. Quantifying disturbed hill dipterocarp forest lands in Ulu Tembeling, Malaysia with HRV/SPOT images. J. Photogramm. Remote Sens. 1996, 51, 39–48. [Google Scholar]
- De Wasseige, C.; Defourny, P. Remote sensing of selective logging impact for tropical forest management. For. Ecol. Manag. 2004, 188, 161–173. [Google Scholar] [CrossRef]
- Nilsson, M. Estimation of tree heights and stand volume using an airborne lidar system. Remote Sens. Environ. 1996, 56, 1–7. [Google Scholar] [CrossRef]
- Means, J.E.; Acker, S.A.; Harding, D.J.; Blair, J.B.; Lefsky, M.A.; Cohen, W.B.; Harmon, M.E.; McKee, W.A. Use of large-footprint scanning airborne lidar to estimate forest stand characteristics in the Western Cascades of Oregon. Remote Sens. Environ. 1999, 67, 298–308. [Google Scholar] [CrossRef]
- Zhang, K.; Chen, S.-C.; Whitman, D.; Shyu, M.-L.; Yan, J.; Zhang, C. A progressive morphological filter for removing nonground measurements from airborne LIDAR Data. IEEE Trans. Geosci. Remote Sens. 2003, 41, 872–881. [Google Scholar] [CrossRef]
- Okuda, T.; Hiroshima University, Higashi Hiroshima, Japan. Logging road networks were clearly visible on an airborne LiDAR image of a selectively logged tropical rain forest. 2014. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Yamada, T.; Niino, M.; Yoshida, S.; Hosaka, T.; Okuda, T. Impacts of Logging Road Networks on Dung Beetles and Small Mammals in a Malaysian Production Forest: Implications for Biodiversity Safeguards. Land 2014, 3, 639-657. https://doi.org/10.3390/land3030639
Yamada T, Niino M, Yoshida S, Hosaka T, Okuda T. Impacts of Logging Road Networks on Dung Beetles and Small Mammals in a Malaysian Production Forest: Implications for Biodiversity Safeguards. Land. 2014; 3(3):639-657. https://doi.org/10.3390/land3030639
Chicago/Turabian StyleYamada, Toshihiro, Masahiro Niino, Satoru Yoshida, Tetsuro Hosaka, and Toshinori Okuda. 2014. "Impacts of Logging Road Networks on Dung Beetles and Small Mammals in a Malaysian Production Forest: Implications for Biodiversity Safeguards" Land 3, no. 3: 639-657. https://doi.org/10.3390/land3030639