
The Productivity and Carbon Exchange of an Intensively Managed Pasture in Central Kentucky



School of Agriculture and Natural Resources, Kentucky State University, 400 East Main Street, Frankfort, KY 40601, USA
*
Author to whom correspondence should be addressed.
Atmosphere 2024, 15(3), 348; https://doi.org/10.3390/atmos15030348
Submission received: 11 February 2024
/
Revised: 4 March 2024
/
Accepted: 9 March 2024
/
Published: 12 March 2024
(This article belongs to the Special Issue Climate Change and Agriculture: Impacts and Adaptation)
Abstract
:Intensive pasture management that aims at providing season-long forage while minimizing soil degradation is increasingly becoming an important grazing strategy in Kentucky. Typically, it involves the use of high-yielding warm and cool season forage species that are well suited to local soil and climate conditions, meeting the dual-purpose provision of high nutritional value while remaining resilient to grazing pressure and changing climate. Monitoring carbon exchange is a crucial component for effective pasture management to promote sustainable pastureland management practices. We hypothesized that pasturelands, when intensively managed, would exhibit a small but important CO2 cumulative uptake year-round. We used the Eddy covariance method to measure the net ecosystem exchange of CO2 (NEE) and productivity of an intensively managed pastureland at Kentucky State University Research and Demonstration station from 2015 to 2020. The study has two objectives: to quantify interannual variability in net ecosystem exchange, and examine the controlling environmental factors, in particular temperature, sunlight, and precipitation of NEE. Diurnal and seasonal fluctuations followed typical patterns of carbon uptake and release. Overall, the pasture site consistently was carbon sink except for 2016, in part due to a warmer winter season than usual, sequestering 1394 gCm−2 over the study period. Precipitation and temperature were critical environmental factors underpinning seasonal CO2 uptake and release. Of critical importance was the net carbon uptake during the non-growing season.
1. Introduction
Animal agriculture is a carbon-intensive sector emitting 75–80 percent of the total agricultural greenhouse gases (GHG) [1]. The sector’s activities take up a massive 40 million km2 (77 percent) of the land available globally for agriculture (51 million km2) [2,3]. This comes as no surprise, as the sector is a leading source of protein products, including dairy, eggs, meat, and poultry, with demand growing rapidly in developing economies [4]. Kentucky’s mild climate, optimum precipitation, and productive soil are conducive for establishing wider use and adoption of intensive pasture management practices. With a land mass of 1.65 million hectares [5], Kentucky’s pastures are predominantly planted with high-yield grass–legume mixtures as key components of rotational grazing. However, their intensive management and rotational grazing practices make them sensitive to carbon losses, distinguishing them from permanent grasslands. Improved pasture management practices to increase productivity and increase soil C sequestration include rotational grazing systems (rotational grazing), amendments to boost pasture productivity, and market and financial incentives for early adopters and ranchers. The increased demand for beef and dairy products has intensified grazing, and today 40 percent of the US’s total methane (CH4) emission, is mostly from livestock production [6].
The intention of managing pastures intensively is to stimulate intermittent above-ground biomass accumulation via photosynthesis while allowing recovering soils to slowly accumulate soil organic carbon over time [7]. How land for future grazing and animal feed production is sustainably managed with increasing demands for animal protein is a question that continues to evolve [8,9]. While the biological and ecological damages of extensive grazing on ecosystems, such as overgrazing and vegetation degradation, extensive damage to water quality, soil compaction and erosion increase, elevated risk to threatened species, and other ecosystem disruptions, are well documented and have been extensively explored [7,10,11], intensively managed pastures remain largely unknown. This is despite the fact that over the past three decades, intensive pasture farming has evolved substantially.
Of the 5.1 billion hectares of land agriculture occupies, 78 percent (~4 billion hectares of land) is under grazing or used for livestock production [12], making it the largest land use devoted to human use by area [13,14]. This is gaining increasing importance and increased emphasis in the C sequestration options considering that it accounts for 10–30 percent of the global SOC [15,16]. According to recent data, of the total land area of 370 million hectares in the U.S.A., about 40% was farmland, of which 45.4% was permanent pasture [17]. The deployment cost of improved grazing could be higher than conventional grazing systems.
The traditional intensive or mob grazing system is often blamed for soil degradation and the crisis has forced ranchers and producers in the livestock sector to accelerate the use and adoption of rotational grazing practices. In rotational grazing, animals (mainly cows or sheep) are on a regular schedule to move from one paddock to another. It may also be seen as moving animal herd assets from one sector to another. This practice in particular buys time while another recovers. This process is used to capture carbon gains from cycles and diversify holdings. The strategy used for animal rotation is quite tricky and somewhat risky, as the costs associated with the operation are high. In the United States, large-scale use and implementation of managed pasturing have recently become a particularly popular C sequestration strategy, with multiple carbon and other soil benefits, both on and off sites. In contrast with intensive grazing (e.g., mob grazing), rotational grazing has been proposed as the scenario where soil and recovery grass species would profit from the decaying animal feces (cow dung), potentially adding organic carbon. This pathway is thought of as a contribution to the long-term carbon livestock decarbonization.
Effectively managing pastures can enhance the significant accumulation of carbon [18]. Most of the strategies for raising pasture yield revolve around encouraging soil-based carbon resources and targeting ground carbon storage, while maintaining moderate stocking rates [19,20,21,22]. Accordingly pasture areas have the potential to trap a net 295 Tg CO2 yr−1 globally (1 Tg = 1012 g), and when properly managed could store 50% more annually [23]. Some studies have demonstrated that grazed grasslands are both sources and sinks of carbon [24], while others have shown them to be carbon neutral [25]. Regarding the effect of grazing on the amount of soil carbon, it has been established that it can enhance and reduce it, largely influenced by intensity [26,27].
The carbon status of an ecosystem serves as an indicator of its ecological productivity [28]. Gross Primary productivity (GPP) is important in that it is quantified as the carbon fixed during photosynthesis over a given period and is driven by biophysical drivers such as air temperature, vapor pressure deficit (VPD), soil water content (SWC), incoming radiation, vegetation type, and canopy characteristics [29,30].
NEE is driven by many biophysical factors and processes, such as soil moisture [31,32,33]. Constraining the global C balance requires an understanding of the diurnal, monthly, and yearly variations in ecosystem exchange, as well as the micrometeorological and biophysical drivers of NEE [34,35]. Therefore, this study aims to measure the pasture’s NEE at temporal scales (diurnal and seasonal). We employed a 4-m tall Eddy Covariance (flux) station. Intensive pasture production practices that aim at maximizing the yield and quality of forage while minimizing soil degradation are increasingly becoming an important management strategy for producers. Typically, it involves the alternative use of high-yielding warm and cool season forage species that are well-suited to local soil and climate conditions, providing high nutritional value to grazing animals and are resilient to grazing pressure. Measurements that monitor soil carbon loss are also a crucial part, as they provide insights into how different land management practices, such as rotational grazing, fertilization, and soil conservation, impact carbon dynamics in pasture ecosystems. To address this measurement gap and help understand the soil organic carbon sequestration potential, we studied the net ecosystem exchange (NEE) of a pastureland at a demonstration site located at Kentucky State University. We hypothesized that pasturelands, when intensively managed, would exhibit a small but important CO2 uptake throughout the seasons. The goal was to quantify interannual variability in NEE and to investigate the environmental factors influencing carbon exchange, particularly temperature, radiation, and precipitation.
This paper was structured as follows: First, we give a detailed introduction of the study covering the key research questions and the objective we intend to satisfy, and then present the materials and methods inclusive of site description, Eddy Covariance principle, data collection, and processing. The next sections present the results, the discussion, and then the conclusion session.
2. Materials and Methods
2.1. Site Characterization
The study was carried out at the 40-ha Kentucky State University Research Farm in Frankfort, Kentucky, which is located at 38° 6′56.71″ N, 84°53′22.91″ W (Figure 1). The site has a humid temperate climate with a mean yearly temperature of 15.8 °C and 1460 mm of rainfall. The instrumentation for the flux site is listed in Table 1.
The soil at the station is mainly composed of well-drained McAfee silt loam and Lowell-Sandview silt loam. Organic matter is around 2%, and bulk density is 1.4 g/cm3, categorizing it as Hydrologic soil group C, indicating a slow infiltration rate. Moderate frost action occurs, with 182–191 frost-free days. The climate is humid subtropical with abundant rainfall, while the EC tower aligns with the prevailing southwest wind. The parent material is clayey residuum from phosphatic limestone/limestone and shale. Cation exchange capacity is 16–18 milliequivalents per 100 g, and soil pH is 6.6, with a 6% to 20% slope. Meat goats undergo four weeks of rotational grazing in a pasture of mixed grasses.
2.2. Principles of Eddy Covariance
Eddy covariance is a micrometeorological approach that measures turbulent gas flux and wind at the boundary layer [36], by focusing solely on the turbulent, non-deterministic portion of the layer [37,38]. This methodology has allowed for the quantification of trace gas exchange over any canopy and ensures that these observations can be conducted over long periods compared to destructive approaches that are often laborious to conduct and largely unscalable [38,39]. Through this methodology, we were able to determine the patterns of gas emissions across temporal scales and therefore have a better understanding of how they relate to environmental variables [25,39,40,41].
The Eddy covariance technique is premised on the law of mass conservation. Considering a plant canopy acting as a uniform source (sink) of gas (e.g., CO2), we wish to determine the exchange of C between the canopy and the layer of air above it. For ease of understanding, micrometeorologists use an idealized notional control volume (CV) placed above a plant canopy. The EC calculates flux as the cross product (covariance) of the instantaneous turbulent vertical wind velocity () and concentration of a scalar quantity (density, ) of interest averaged over a sufficiently long time, crossing a horizontal plane at the measurement height (where ) in the Eulerian frame of reference.
The principle of Reynolds averaging guides Eddy Covariance. As a result of the changing and random character of turbulent motion, meteorological factors recorded at a given location in space show considerable changes around the mean over time. Fluxes in the unstable atmosphere are random, necessitating statistical analysis of their observations [38]. Following Reynolds (1895) [42], it is a usual practice to divide flow variables into mean and fluctuation components; to do this, we express the two field variables (CO2) as a scalar amount and upward wind speed, respectively. W is taken to be normal to the ground. The Reynolds averaging rule helps to partition into slowly shifting mean values and relatively fluctuating unstable parts:
where the mean values are indicated by overbars and the fluctuating components by primes. Based on this rule, the average of a fluctuating variable is zero, and the average of a product between a mean and fluctuating variable:
However, the product of two co-varying variables is non-zero. The average of the product of two variables is equal to the product of the averages of the means () plus the product of the averages of the fluctuating components (). This is represented below.
The measured and computed parameters (variance, covariance, and flux) are also regarded as random variables as the materials that traverse the interface are carried by random turbulent movements. Other estimated flow values and statistical variables for the second instant are also random values.
Certain simplifying assumptions were made for the Eddy Covariance principle to work. Under steady flow, scalar concentrations and wind velocities remain constant over time. Also, when the surface is flat, uniform surfaces eliminate horizontal advection, focusing only on vertical mass flux. Moreso, advection occurs when the assumption of surface homogeneity fails [43,44]. At night, thermal stratification develops due to cooler soil surfaces and warmer atmospheric air, due to accompanying long-wave radiative heat loss to the atmosphere. The steady-state conditions assume materials leaving the surface reach reference height within the averaging time [45]. Calm nights or weak turbulence hinder material transport to measuring height, affecting flux measurements. However, Eddy covariance techniques may underestimate nighttime fluxes due to weak turbulence.
2.3. Data Collection
In 2015, an EC flux tower equipped with a closed-path Li-COR (Li-7200) Infrared Gas (CO2/H2O) Analyzer (IRGA) and 3D sonic anemometer began collecting high-frequency gas, wind and Biomet data established over a grass pasture (Table 1). The site has a gentle slope and 100–150 m fetch. Standard micrometeorological instruments are used at the LI-7200RS supporting site. These include precipitation, air temperature (Ta), and relative humidity (RH), which are monitored using a humidity and temperature probe, Vaisala, Helsinki area (Vantaa), Finland. Photosynthetic Photon Flux Density and a CNR-1 4-way component radiometer were used for radiometric measurements. The LI-7550 interface unit was used to gather data, which were then stored in GHG file format. The raw data were processed using EddyPro® v6.2.2 as seen in Figure 2. Time series data were checked for temporal discontinuities, outliers, and missing observations (Not a Number, or NANs), and noted for gap filling.
NEE is determined by calculating the covariance between instantaneous variances of the trace gas (c′) and the vertical wind velocity (w′), as represented by Equation (4).
The covariates are averaged over half-hour intervals. The micrometeorological sign standard was used in this analysis, where negative NEE numbers imply the gain of CO2 into the reference ecosystem.
2.4. Data Filtering, Gap Filling, and Flux Correction
Before completing the gap-filling process, low-quality data were eliminated from the requisite EddyPro® output file. Data entries in the output files are flagged for quality following [46,47], and despiking algorithms are executed within EddyPro® v6.2.2 to generate flux calculations. In consideration of the biological exchange capacity inherent in grasslands, CO2 flux values exceeding 40 μmolm−2s−1 and falling below −40 μmolm−2s−1 were also removed for subsequent gap-filling. Additionally, outliers were excluded (Table 2). For gap-filling, the REddyProc software (1.3.2) was used [48,49]. In both day and nighttime conditions, when data gaps were smaller than one hour, the 30 min averages of the preceding and following time steps were averaged (i.e., interpolation) and used to fill in the missing values. Under these assumptions, daytime Re is temperature dependent as well, and the time Re vs. Ts relationship can be extended to daytime conditions (i.e., nighttime NEE (NEEnight) = Re). Finally, GPP was calculated by subtracting NEE from Re (GPP = Re − NEE). Percentages of data gap filled for the years were: 2015 (66.33%), 2016 (38.85%), 2017 (37.65%), 2018 (47.78%), 2019 (33.70%), and 2020 (61.03%). In the years 2015 and 2020, due to instrument failure and calibration challenges, 151 and 127 days of observations were not recorded.
3. Results
3.1. Meteorological Conditions in 2015–2020
The study area’s air temperature varied significantly with the seasons; in July, it was the hottest at 24.57 ± 0.35 °C, and in January, it was lowest at 1.19 ± 1.23 °C (Figure 3). Every year was hotter than the historical average (13.21 °C) from 2008 to 2022, with 2016 being the hottest at 13.95 °C (21.9%) (Figure 4). January, February, and December saw considerable shifts in temperature from the norm (1.66 °C, 1.78 °C, and 1.41 °C, respectively). All the months were hotter than the historical average except for March, April, and June (−0.37 °C, −0.16 °C, and −0.35 °C, respectively). Monthly precipitation followed clear historical patterns for the site (Figure 5). February, March, June, August, October, and November had higher average precipitation than other months over the study period. The year 2018 had the highest amount of precipitation (1828.3 ± 0.61 mm) (Figure 6).
3.2. Light Response Curve
The Michaelis–Menten (light response) model was used to assess the response of Net Ecosystem Exchange (NEE) to light intensity (PPFD) for a pasture typical year (2016) in the study area. The model was fitted according to this equation:
where Amax is the maximum ecosystem C uptake rate (g C m−2 s−1) at saturated light intensity, Re is the daytime ecosystem respiration (g m−2 s−1), PPFD is photosynthetic photon flux density PPFD (μmol m−2 s−1), and ∝ is the apparent quantum efficiency (g C m−2 s−1 PPFD−1).
A typical year, 2016, was chosen from the dataset for the modeling (Table 2). The Michaelis–Menten rectangular hyperbolae model (Equation (5)) was used to determine the effect of light intensity, proxied using photosynthetically active photon flux density (PPFD), on net ecosystem exchange (Figure 7).
Amax was lowest in the winter months, particularly lowest in January (0.18 g C m−2 s−1), and peaks in the growing season, June (2.36 g C m−2 s−1). This indicates that despite the summer months having the highest optimal carbon uptake rate at saturated lights, the pasture had substantial carbon uptake even in the cold winter months. The non-zero values of this Michalis Menten constant reveal that photosynthesis and carbon storage occur year-round, even at low PPFD (4.4 KW/m2). The efficiency at which pasture converts photons of light to stored carbon follows a similar trend, as α was lowest in the winter months (0.0003–0.0008 g C m−2 s−1 PPFD−1) and highest in June (2.36 g C m−2 s−1 PPFD−1). July and August had relatively lower light use efficiency, which may be attributable to the oversaturation of light making the pasture highly inefficient. Likewise, respiration was highest in June (5.36 g C m−2 s−1).
3.3. Daily, Diurnal and Seasonal Variations
The amount of carbon exchanged within an ecosystem, or net ecosystem exchange, is a key indicator of its overall productivity, which explains the relationship concerning respiration and photosynthesis as well as the flow of carbon dioxide between land and above ecosystems [50,51,52], and is better understood when examined daily, diurnally, and seasonally.
3.3.1. Daily and Diurnal NEE
Diurnal NEE patterns were a clear U-shape (Figure 8d,f). Typically, the pasture begins to gain carbon from 8 am and increases towards noon as more light is received for photosynthesis. This typically peaks from 12 noon to 1 pm and diminishes as the day passes by. NEE was positive (carbon loss) typically at hours before 8 am and beyond 8 pm. This shows a distinct diurnal trend that rules daytime and nighttime carbon exchange. A slightly similar but reversed pattern was observed for GPP. The observed pattern for NEE varies by month and season with a higher carbon uptake rate (-ve NEE) observed in the spring and summer months. Due to the lower winter air temperatures, diurnal NEE patterns were diminished in all of the years under investigation during the nongrowing seasons, which ran from January through March, and October through December (Figure 8d–f). January–March had the lowest NEE, GPP, and Re. The daily rate of carbon uptake was not constant all year round. The average diurnal uptake rate of carbon (0.163 gCm−2day−1) by the pasture vegetation from April to September was higher than the Re (0.139 gCm−2day−1), and clearly for the growing season for DOY-135 and 26 October 2016 (Figure 8a,c) thereby acting as storage of carbon.
3.3.2. Seasonal NEE
Significant seasonal fluctuations were also observed for carbon exchange; more carbon was lost from January to March (Figure 9). Carbon assimilation started in March, at the time GPP was higher than respiration, and NEE peaked in April which gradually reduced towards the end of the year. After that, there was a consistent change in both photosynthetic and respiratory processes through July (Figure 9). Across the years, the monthly total of GPP, NEE, and Re ranged from 1488.58 gCm−2, −400.95 gCm−2, and 1271.70 gCm−2 in July, April, and July, respectively, to 135.25 gCm−2, 63.19 gCm−2, and 198.44 gCm−2 all in January.
Cumulative carbon for the pasture site for the 6–year period (2015–2020) was calculated by aggregating the half-hourly data, totaling −1394 ± 146 gCm−2, indicating carbon uptake of −1242 ± 151 gCm−2 during the growing season and −152 ± 91 gCm−2 outside of the growing season, indicative of its sink status (Figure 10, Table 3). Carbon assimilation was higher in the growing seasons than in other periods. This indicates that the pasture acted as a carbon sink all year long (Table 3). For example, the non-growing seasons of 2018 and 2019 had lower carbon uptake of 44.31 gCm−2yr−1 and 31.37 gCm−2yr−1, respectively, due to lower precipitations. Overall, both the growing (−1242 ± 151 gCm−2) and the non-growing (−152 ± 91 gCm−2) seasons of the pasture were net carbon sinks. The non-growing season contributed about 11% of the total carbon sink in the pasture.
When comparing the ratio of GPP to Re, the growing season of year 2016 had the highest ratio of 1.07, indicating that 107% of the plant carbon assimilation was excessively consumed by pasture or to support the activities of heterotrophs in the soil. The lowest ratio for the growing season was in the year 2020 with an average value of 0.56. For the non-growing season, the year 2018 had a higher ratio of 1.2 while 2020 also had the lowest (0.63). When both seasons were pooled together for each year, 2016 had the highest ratio, 1.01, while 56.8% of the photosynthesis assimilation of pasture in 2020 was used for plant support, structural maintenance, and other soil activities. The values fell within the range of 0.55–1.02 presented by [39]. Pasture maintains a critical balance of photosynthesis and respiration (carbon storage and usage) efficiently in both the growing and non-growing seasons which supports why they sequester carbon year-long.
4. Discussion
The primary focus of this study is to examine how bioclimatic factors and a moderately grazed pasture in Kentucky, which proxy the roles of management, interact to control the exchange of carbon dioxide (CO2) in pastureland at temporal scales. Our investigation into the carbon exchange dynamics reveals consistent patterns in line with established vegetation ecosystem behaviors and prior research [53]. Diurnal variations in Net Ecosystem Exchange (NEE) are observed, with negative values during daylight hours indicating heightened photosynthesis, while respiration processes dominate during the nighttime, resulting in near-zero or positive NEE values. This diurnal pattern corresponds to normal carbon dynamics in central Kentucky throughout the growing cycle [54].
This study emphasizes the significance of temporal scaling in understanding carbon dynamics, particularly about precipitation and air temperatures. These factors significantly influence surface–atmosphere CO2 exchange at temporal scales ranging from 30 min to interannual. Seasonal Net Ecosystem Exchange (NEE) mirrors pasture phenology, with precipitation playing a pivotal role in determining net productivity. Natural responses regulate the quantity of NEE rates [55]. These reactions also adjust respiration to respond to changes in temperature and moisture [56], which in turn affects the timing and rate of CO2 sequestration [57].
The non-growing season of 2015 displayed unique favorable conditions for carbon assimilation, characterized by a mean temperature of 4.2 °C and substantial total rainfall of 566.7 mm. A favorable carbon balance resulted from lower temperatures suppressing respiration and microbial breakdown. GPP was more than RE, leading to carbon uptake. Abundant precipitation provided the moisture necessary for plant health and photosynthetic activity, further promoting carbon uptake. The presence of evergreen or cold-tolerant species sustained active photosynthesis during the non-growing season, contributing to carbon sequestration. Furthermore, the carbon storage benefit was strengthened by carbon accumulated from the previous growing period as with forest ecosystems [58].
In contrast, the non-growing months of 2018 and 2019 exhibited net carbon losses. Elevated precipitation (712.216 mm) and lower air temperatures (3.99 °C) resulted in GPP (220.29 gCm−2) falling behind RE (264.59 gCm−2). Colder conditions inhibited plant growth, photosynthesis, and microbial activity, causing a net release of carbon in the form of CO2. Excessive precipitation led to waterlogged soils, reducing oxygen availability, and limiting root respiration and microbial activity. Plant senescence during the non-growing season further contributed to reduced photosynthetic activity, exacerbating the carbon imbalance. Carbon allocation and potential decomposition further accentuated carbon losses.
The cumulative carbon budget, evaluated over a six-year research period, underscores the consistent role of the pasture as a carbon sink. Our observations align with the work of Henderson et al. (2015), documenting carbon sequestration and exchange dynamics. The net carbon exchange for the study was estimated at −1394 ± 146 gCm−2, comprising a carbon uptake of −1242 ± 151 gCm−2 during the growing season and a net carbon absorption of −152 ± 91 gCm−2 during the non-growing season. This sustained carbon sequestration highlights the pasture’s role as a year-round carbon sink.
In this study, the non-growing season’s contribution to the total carbon uptake, approximately 11%, underscores the importance of pasture ecosystems throughout the year. Moreover, when considering all seasons combined, an average of 14.5% (−1393.7 gCm−2) of the photosynthetic assimilation generated was stored within the pasture systems (Table 4).
We compared our findings with other values obtained in the reviewed literature in Table 4. Several studies have estimated the amount of NEE derived from pastures across the world, which ranged from −408 to 261 gC m−2 yr−1 (Table 4). Our pasture site was intensively managed with moderate grazing allowed for one month only in a year, which led to an annual average of −232.3 gC m−2. This was within the range of values observed at the experimentation station in Macaíba District, Brazil, South America [77] where a rotationally grazed pasture yielded −215 ± 22 gC m−2 yr−1, Asia (−116 to −41 gC m−2 yr−1), Europe (−245 to 1.32 gC m−2 yr−1), and Oceania (−408 to 190 gC m−2 yr−1).
In North America, NEE values ranged from −59 to 261 gC m−2 yr−1. This showed some disparities in the values, which were due to the type and intensity of grazing, soil characteristics, and other variables. Upon close examination, the finding by [60], which was 261 gC m−2 yr−1 where pasture was a net source, was reported to have been influenced by the severe drought and the uncontrolled grazing at the Sardinilla site. Also, the carbon source study [74] at the Waikato dairy deep peat farm in Oceania was a result of an intense, year-round rotational grazing coupled with the use of a mobile Eddy covariance setup, and carbon loss was determined using Phytomass Index approach, which estimates all carbon lost within the farm.
Based on our findings in Kentucky (232.3 g Cm−2yr−1 or 0.04 tCO2Ha−1yr−1), of the available 1.7 million hectares of pasture lands in Kentucky, there is a potential to annually sequester about 0.07 MtCO2 and 0.22 MtCO2 when projected over the available farmland of 5.2 million hectares
When scaled for the entire U.S., the available pasture lands of 51 million hectares save 2.2 MtCO2 annually while the entire farmland of 362 million hectares could help reduce emissions by 15.4 MtCO2 in the same referenced timeframe. This more than doubles the US projection of 6.6 Mt CO2yr−1 Follett, Kimble and Lal (2001). This may be insignificant but highlights the potential role of pasture establishment in carbon sequestration and the global reduction of CO2 emissions.
Highlighting the limitations of this study is very important as to the magnitude and the interpretation of the results, as well as the Eddy covariance technique scalability. Micrometeorology tools might not be ideal to capture pasture carbon exchange, as animals are constantly moving about and their contribution may not be properly captured. Another limitation is the fact that pasture grows all year round, and the simplistic models employed by the Eddy covariance gap-filling techniques pose a challenge, as the minute changes might be under- or overestimated. Also, the assumptions of turbulence and horizontal fluxes not factored into total flux calculations pose a great risk of underestimation, although most of these are factored in the model calibrations.
5. Conclusions
This study focused on the carbon exchange over a moderately graced pasture in Kentucky from 2015 to 2020 using the Eddy Covariance technique. In all the analyzed years, our study revealed that the pasture site was able to sequester carbon significantly. This six-year study revealed that the pasture ecosystem’s ability to sequester carbon is high and is modulated greatly by precipitation and temperature, becoming a carbon sink in years with high precipitation (wet) and source/neutral in years with higher levels of air temperature (dry).
Our analysis also revealed that towards noon, the pasture begins to sequester carbon; it also stores more carbon in the spring months in particular when air temperature is moderate. Also, we observed a clear diurnal NEE pattern which is limited by photosynthetically active radiation, and seasonal trends consistent with pasture phenology and precipitation, marking the growing season. Despite the harsh conditions of the non-growing seasons in some of the years under study, the pasture was able to sequester carbon, indicative of its potential when properly managed.
Assessing the pasture carbon exchange cumulatively revealed that year-round, the pasture ecosystem is a carbon sink. Coupling this with the relative ease of establishing, growing, and managing pastures indicates that it is a potential game-changer CO2 emission reduction strategy if developed, in both the short and long term. The findings of this study fill the knowledge gap on how a managed pasture in a typical humid temperature climate exchanges carbon diurnally and seasonally, revealing a clear picture of year-round carbon storage potential, and placing the same finding within the range of the existing literature. As the world keeps searching for sustainable options to reduce global warming, recognizing the pasture ecosystem’s all-season importance in carbon sequestration may be key to actualizing this.
Author Contributions
All authors contributed to the study. M.G. and B.G.: research conception, supervision, and funding; M.G., I.R. and J.B.: data collection, and first draft for 2015–2018; I.F. and M.G.: revised draft for 2015–2020, manuscript preparation and editing; I.F.: software, data management, data analysis, visualization and validation. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the intramural research program of the U.S. Department of Agriculture, National Institute of Food and Agriculture, Evans-Allen Project #1020560.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data will be made available upon request. The data are not publicly available due to privacy.
Conflicts of Interest
The authors declare that there are no known conflicts of interest or personal ties that would be thought to have an impact on the findings of the study that is included in this paper.
References
- FAO. Emissions from Agriculture and Forest Land: Global, Regional and Country Trends 1990–2019; FAO: Rome, Italy, 2021. [Google Scholar]
- Ritchie, H. How Much of the World’s Land Would We Need in Order to Feed the Global Population with the Average Diet of a Given Country; Our World in Data: Oxford, UK, 2017. [Google Scholar]
- FAO. The State of World Fisheries and Aquaculture 2020. In Sustainability in Action; FAO: Rome, Italy, 2020. [Google Scholar] [CrossRef]
- Willett, W.; Rockström, J.; Loken, B.; Springmann, M.; Lang, T.; Vermeulen, S.; Garnett, T.; Tilman, D.; DeClerck, F.; Wood, A.; et al. Food in the Anthropocene: The EAT–Lancet Commission on Healthy Diets from Sustainable Food Systems. Lancet 2019, 393, 447–492. [Google Scholar] [CrossRef]
- Farmland Innovation Center. Kentucky Data and Statistics. Available online: https://farmlandinfo.org/statistics/kentucky-statistics/ (accessed on 9 February 2024).
- Searchinger, T.; Herrero, M.; Yan, X.; Wang, J.; Beauchemin, K.; Kebreab, E. Opportunities to Reduce Methane Emissions from Global Agriculture; Faculty of Cornell University: Ithaca, NY, USA; Faculty of Princeton University: Princeton, NJ, USA, 2021. [Google Scholar]
- Jordon, M.W.; Buffet, J.C.; Dungait, J.A.J.; Galdos, M.V.; Garnett, T.; Lee, M.R.F.; Lynch, J.; Röös, E.; Searchinger, T.D.; Smith, P.; et al. A Restatement of the Natural Science Evidence Base Concerning Grassland Management, Grazing Livestock and Soil Carbon Storage. Proc. R. Soc. B 2024, 291, 20232669. [Google Scholar] [CrossRef] [PubMed]
- Steinfeld, H.; Gerber, P. Livestock Production and the Global Environment: Consume Less or Produce Better? Proc. Natl. Acad. Sci. USA 2010, 107, 18237–18238. [Google Scholar] [CrossRef] [PubMed]
- Henchion, M.; Moloney, A.P.; Hyland, J.; Zimmermann, J.; McCarthy, S. Review: Trends for Meat, Milk and Egg Consumption for the next Decades and the Role Played by Livestock Systems in the Global Production of Proteins. Animal 2021, 15, 100287. [Google Scholar] [CrossRef] [PubMed]
- Teague, R.; Kreuter, U. Managing Grazing to Restore Soil Health, Ecosystem Function, and Ecosystem Services. Front. Sustain. Food Syst. 2020, 4, 534187. [Google Scholar] [CrossRef]
- Baggio, R.; Overbeck, G.E.; Durigan, G.; Pillar, V.D. To Graze or Not to Graze: A Core Question for Conservation and Sustainable Use of Grassy Ecosystems in Brazil. Perspect. Ecol. Conserv. 2021, 19, 256–266. [Google Scholar] [CrossRef]
- Kgosikoma, E. Grazing Management Systems and Their Effects on Savanna Ecosystem Dynamics: A Review. J. Ecol. Nat. Environ. 2013, 5, 88–94. [Google Scholar] [CrossRef]
- Asner, G.P.; Elmore, A.J.; Olander, L.P.; Martin, R.E.; Harris, T. Grazing Systems, Ecosystem Responses, and Global Change. Annu. Rev. Environ. Resour. 2004, 29, 261–299. [Google Scholar] [CrossRef]
- Liebig, M.A.; Gross, J.R.; Kronberg, S.L.; Hanson, J.D.; Frank, A.B.; Phillips, R.L. Soil Response to Long-Term Grazing in the Northern Great Plains of North America. Agric. Ecosyst. Environ. 2006, 115, 270–276. [Google Scholar] [CrossRef]
- Post, W.M.; Emanuel, W.R.; Zinke, P.J.; Stangenberger, A.G. Soil Carbon Pools and World Life Zones. Nature 1982, 298, 156–159. [Google Scholar] [CrossRef]
- Eswaran, H.; Van Den Berg, E.; Reich, P. Organic Carbon in Soils of the World. Soil Sci. Soc. Am. J. 1993, 57, 192–194. [Google Scholar] [CrossRef]
- USDA. Crop Production 2012 Summary; No. January; National Agricultural Statistics Service: Washington, DC, USA, 2013.
- Silveira, M.; Hanlon, E.; Azenha, M.; Da Silva, H.M. Carbon Sequestration in Grazing Land Ecosystems. EDIS 2012, 2012, 1–4. [Google Scholar] [CrossRef]
- Conant, R.T.; Paustian, K.; Elliott, E.T. Grassland Management and Conversion into Grassland: Effects on Soil Carbon. Ecol. Appl. 2001, 11, 343–355. [Google Scholar] [CrossRef]
- Conant, R.T. Grassland Soil Organic Carbon Stocks: Status, Opportunities, Vulnerability. In Recarbonization of the Biosphere: Ecosystems and the Global Carbon Cycle; Springer: Berlin/Heidelberg, Germany, 2012. [Google Scholar] [CrossRef]
- Follett, R.F.; Kimble, J.M. The Potential of US Grazing Lands to Sequester Carbon and Mitigate the Greenhouse Effect; CRC Press: Boca Raton, FL, USA, 2000. [Google Scholar]
- Teague, W.R.; Apfelbaum, S.; Lal, R.; Kreuter, U.P.; Rowntree, J.; Davies, C.A.; Conser, R.; Rasmussen, M.; Hatfeld, J.; Wang, T.; et al. The Role of Ruminants in Reducing Agriculture’s Carbon Footprint in North America. J. Soil Water Conserv. 2016, 71, 156–164. [Google Scholar] [CrossRef]
- Henderson, B.B.; Gerber, P.J.; Hilinski, T.E.; Falcucci, A.; Ojima, D.S.; Salvatore, M.; Conant, R.T. Greenhouse Gas Mitigation Potential of the World’s Grazing Lands: Modeling Soil Carbon and Nitrogen Fluxes of Mitigation Practices. Agric. Ecosyst. Environ. 2015, 207, 91–100. [Google Scholar] [CrossRef]
- Gilmanov, T.G.; Verma, S.B.; Sims, P.L.; Meyers, T.P.; Bradford, J.A.; Burba, G.G.; Suyker, A.E. Gross Primary Production and Light Response Parameters of Four Southern Plains Ecosystems Estimated Using Long-Term CO2-Flux Tower Measurements. Glob. Biogeochem. Cycles 2003, 17, 1071. [Google Scholar] [CrossRef]
- Flanagan, L.B.; Wever, L.A.; Carlson, P.J. Seasonal and Interannual Variation in Carbon Dioxide Exchange and Carbon Balance in a Northern Temperate Grassland. Glob. Chang. Biol. 2002, 8, 599–615. [Google Scholar] [CrossRef]
- Dormaar, J.F.; Adams, B.W.; Willms, W.D. Impacts of Rotational Grazing on Mixed Prairie Soils and Vegetation. J. Range Manag. 1997, 50, 647. [Google Scholar] [CrossRef]
- Pineiro, G.; Paruelo, J.M.; Oesterheld, M.; Jobbágy, E.G. Pathways of Grazing Effects on Soil Organic Carbon and Nitrogen. Rangel. Ecol. Manag. 2010, 63, 109–119. [Google Scholar] [CrossRef]
- Reichstein, M.; Bahn, M.; Ciais, P.; Frank, D.; Mahecha, M.D.; Seneviratne, S.I.; Zscheischler, J.; Beer, C.; Buchmann, N.; Frank, D.C.; et al. Climate Extremes and the Carbon Cycle. Nature 2013, 500, 287–295. [Google Scholar] [CrossRef]
- Reverter, B.R.; Sánchez-Cañete, E.P.; Resco, V.; Serrano-Ortiz, P.; Oyonarte, C.; Kowalski, A.S. Analyzing the Major Drivers of NEE in a Mediterranean Alpine Shrubland. Biogeosciences 2010, 7, 2601–2611. [Google Scholar] [CrossRef]
- Jiao, L.; Lu, N.; Sun, G.; Ward, E.J.; Fu, B. Biophysical Controls on Canopy Transpiration in a Black Locust (Robinia Pseudoacacia) Plantation on the Semi-Arid Loess Plateau, China. Ecohydrology 2015, 9, 1068–1081. [Google Scholar] [CrossRef]
- Flanagan, L.B.; Johnson, B.G. Interacting Effects of Temperature, Soil Moisture and Plant Biomass Production on Ecosystem Respiration in a Northern Temperate Grassland. Agric. For. Meteorol. 2005, 130, 237–253. [Google Scholar] [CrossRef]
- Jones, L.-A.; Madani, N.; Kimball, J.-S.; Reichle, R.-H.; Colliander, A. Effects of Recent Regional Soil Moisture Variability on Global Net Ecosystem CO2 Exchange. In AGU Fall Meeting Abstracts; American Geophysical Union: Washington, DC, USA, 2017; Volume 2017, p. H21I-1586. [Google Scholar]
- Jung, M.; Reichstein, M.; Schwalm, C.R.; Huntingford, C.; Sitch, S.; Ahlström, A.; Arneth, A.; Camps-Valls, G.; Ciais, P.; Friedlingstein, P.; et al. Compensatory Water Effects Link Yearly Global Land CO2 Sink Changes to Temperature. Nature 2017, 541, 516–520. [Google Scholar] [CrossRef]
- Lu, Y.; Murase, J.; Watanabe, A.; Sugimoto, A.; Kimura, M. Linking Microbial Community Dynamics to Rhizosphere Carbon Flow in a Wetland Rice Soil. FEMS Microbiol. Ecol. 2004, 48, 179–186. [Google Scholar] [CrossRef]
- Chakravarthy, D.; Raghavendra, H.V.; Ratnam, J.; Sankaran, M. Soil Respiration Is Correlated with Rainfall and Soil Moisture at Multiple Temporal Scales in a Seasonal Wet Tropical Forest. bioRxiv 2024. [Google Scholar] [CrossRef]
- Baldocchi, D.D.; Hicks, B.B.; Meyers, T.P. Measuring Biosphere-Atmosphere Exchanges of Biologically Related Gases with Micrometeorological Methods. Ecology 1988, 69, 1331–1340. [Google Scholar] [CrossRef]
- Moncrieff, J.B.; Massheder, J.M.; De Bruin, H.; Elbers, J.; Friborg, T.; Heusinkveld, B.; Kabat, P.; Scott, S.; Soegaard, H.; Verhoef, A. A System to Measure Surface Fluxes of Momentum, Sensible Heat, Water Vapour and Carbon Dioxide. J. Hydrol. 1997, 188–189, 589–611. [Google Scholar] [CrossRef]
- Baldocchi, D.D. Assessing the Eddy Covariance Technique for Evaluating Carbon Dioxide Exchange Rates of Ecosystems: Past, Present and Future. Glob. Chang. Biol. 2003, 9, 479–492. [Google Scholar] [CrossRef]
- Law, B.E.; Falge, E.; Gu, L.; Baldocchi, D.D.; Bakwin, P.; Berbigier, P.; Davis, K.; Dolman, A.J.; Falk, M.; Fuentes, J.D.; et al. Environmental Controls over Carbon Dioxide and Water Vapor Exchange of Terrestrial Vegetation. Agric. For. Meteorol. 2002, 113, 97–120. [Google Scholar] [CrossRef]
- Verma, S.B.; Dobermann, A.; Cassman, K.G.; Walters, D.T.; Knops, J.M.; Arkebauer, T.J.; Suyker, A.E.; Burba, G.G.; Amos, B.; Yang, H.; et al. Annual Carbon Dioxide Exchange in Irrigated and Rainfed Maize-Based Agroecosystems. Agric. For. Meteorol. 2005, 131, 77–96. [Google Scholar] [CrossRef]
- Suyker, A.E.; Verma, S.B. Coupling of Carbon Dioxide and Water Vapor Exchanges of Irrigated and Rainfed Maize-Soybean Cropping Systems and Water Productivity. Agric. For. Meteorol. 2010, 150, 553–563. [Google Scholar] [CrossRef]
- Reynolds, O., IV. On the Dynamical Theory of Incompressible Viscous Fluids and the Determination of the Criterion. Philos. Trans. R. Soc. Lond. A 1895, 186, 123–164. [Google Scholar] [CrossRef]
- Derbyshire, S.H. Atmospheric Boundary Layer Flows—Their Structure and Measurement. by J. C. Kaimal and J. J. Finnigan. Oxford University Press. 1994. Pp. 289. Price £50.00 (Hardback). Isbn 0 19 506239 6. Q. J. R. Meteorol. Soc. 1995, 121, 1177. [Google Scholar] [CrossRef]
- Raupach, M.R.; Finnigan, J.J.; Brunet, Y. Coherent Eddies and Turbulence in Vegetation Canopies: The Mixing-Layer Analogy. Boundary-Layer Meteorol 1996, 78, 351–382. [Google Scholar] [CrossRef]
- Goulden, M.L.; Daube, B.C.; Fan, S.M.; Sutton, D.J.; Bazzaz, A.; Munger, J.W.; Wofsy, S.C. Physiological Responses of a Black Spruce Forest to Weather. J. Geophys. Res. Atmos. 1997, 102, 28987–28996. [Google Scholar] [CrossRef]
- Mauder, M.; Foken, T. Documentation and Instruction Manual of the Eddy-Covariance Software Package TK3 (Update); University of Bayreuth: Bayreuth, Germany, 2015. [Google Scholar]
- Lee, X.; Massman, W.; Law, B. Handbook of Micrometeorology A Guide for Surface Flux Measurement and Analysis; Springer: Berlin/Heidelberg, Germany, 2005. [Google Scholar]
- Falge, E.; Baldocchi, D.; Olson, R.; Anthoni, P.; Aubinet, M.; Bernhofer, C.; Burba, G.; Ceulemans, R.; Clement, R.; Dolman, H.; et al. Gap Filling Strategies for Defensible Annual Sums of Net Ecosystem Exchange. Agric. For. Meteorol. 2001, 107, 43–69. [Google Scholar] [CrossRef]
- Reichstein, M.; Falge, E.; Baldocchi, D.; Papale, D.; Aubinet, M.; Berbigier, P.; Bernhofer, C.; Buchmann, N.; Gilmanov, T.; Granier, A.; et al. On the Separation of Net Ecosystem Exchange into Assimilation and Ecosystem Respiration: Review and Improved Algorithm. Glob. Chang. Biol. 2005, 11, 1424–1439. [Google Scholar] [CrossRef]
- Schulze, E.D.; Lange, O.L.; Ziegler, H.; Gebauer, G. Carbon and Nitrogen Isotope Ratios of Mistletoes Growing on Nitrogen and Non-Nitrogen Fixing Hosts and on CAM Plants in the Namib Desert Confirm Partial Heterotrophy. Oecologia 1991, 88, 457–462. [Google Scholar] [CrossRef]
- Braswell, B.H.; Sacks, W.J.; Linder, E.; Schimel, D.S. Estimating Diurnal to Annual Ecosystem Parameters by Synthesis of a Carbon Flux Model with Eddy Covariance Net Ecosystem Exchange Observations. Glob. Chang. Biol. 2005, 11, 335–355. [Google Scholar] [CrossRef]
- Zhou, G.; Zhou, X.; He, Y.; Shao, J.; Hu, Z.; Liu, R.; Zhou, H.; Hosseinibai, S. Grazing Intensity Significantly Affects Belowground Carbon and Nitrogen Cycling in Grassland Ecosystems: A Meta-Analysis. Glob. Chang. Biol. 2016, 23, 1167–1179. [Google Scholar] [CrossRef] [PubMed]
- Von Randow, G. When the Centre Becomes Radical. JASSS 2003, 6, 28. [Google Scholar]
- Gaumont-Guay, D.; Black, T.A.; Griffis, T.J.; Barr, A.G.; Jassal, R.S.; Nesic, Z. Interpreting the Dependence of Soil Respiration on Soil Temperature and Water Content in a Boreal Aspen Stand. Agric. For. Meteorol. 2006, 140, 220–235. [Google Scholar] [CrossRef]
- Gebremedhin, M.T.; Loescher, H.W.; Tsegaye, T.D. Carbon Balance of No-till Soybean with Winter Wheat Cover Crop in the Southeastern United States. Agron. J. 2012, 104, 1321–1335. [Google Scholar] [CrossRef]
- Wen, X.F.; Yu, G.R.; Sun, X.M.; Li, Q.K.; Liu, Y.F.; Zhang, L.M.; Ren, C.Y.; Fu, Y.L.; Li, Z.Q. Soil Moisture Effect on the Temperature Dependence of Ecosystem Respiration in a Subtropical Pinus Plantation of Southeastern China. Agric. For. Meteorol. 2006, 137, 166–175. [Google Scholar] [CrossRef]
- Omidiran, Y.A. Net Ecosystem Exchange and Air Pollution in North Alabama. Ph.D. Thesis, Alabama Agricultural and Mechanical University, Huntsville, AL, USA, 2015. [Google Scholar]
- Joetzjer, E.; Maignan, F.; Chave, J.; Goll, D.; Poulter, B.; Barichivich, J.; Maréchaux, I.; Luyssaert, S.; Guimberteau, M.; Naudts, K.; et al. Effect of Tree Demography and Flexible Root Water Uptake for Modeling the Carbon and Water Cycles of Amazonia. Ecol. Model. 2022, 469, 109969. [Google Scholar] [CrossRef]
- Gilmanov, T.G.; Morgan, J.A.; Hanan, N.P.; Wylie, B.K.; Rajan, N.; Smith, D.P.; Howard, D.M. Productivity and CO 2 Exchange of Great Plains Ecoregions. I. Shortgrass Steppe: Flux Tower Estimates. Rangel. Ecol. Manag. 2017, 70, 700–717. [Google Scholar] [CrossRef]
- Wolf, S.; Eugster, W.; Potvin, C.; Turner, B.L.; Buchmann, N. Carbon Sequestration Potential of Tropical Pasture Compared with Afforestation in Panama. Glob. Chang. Biol. 2011, 17, 2763–2780. [Google Scholar] [CrossRef]
- Skinner, R.H. High Biomass Removal Limits Carbon Sequestration Potential of Mature Temperate Pastures. J. Environ. Qual. 2008, 37, 1319–1326. [Google Scholar] [CrossRef]
- Zhang, K.; Wang, Y.; Mamtimin, A.; Liu, Y. Carbon dioxide fluxes over grassland ecosystems in the Middle Tianshan region of China with eddy covariance method. Res. Sq. 2023. [Google Scholar] [CrossRef]
- Sun, J.; Zhou, T.C.; Liu, M.; Chen, Y.C.; Liu, G.H.; Xu, M.; Shi, P.L.; Peng, F.; Tsunekawa, A.; Liu, Y.; et al. Water and Heat Availability Are Drivers of the Aboveground Plant Carbon Accumulation Rate in Alpine Grasslands on the Tibetan Plateau. Glob. Ecol. Biogeogr. 2020, 29, 50–64. [Google Scholar] [CrossRef]
- Niu, Y.Y.; Li, Y.Q.; Wang, X.Y.; Gong, X.W.; Luo, Y.Q.; Tian, D.Y. Characteristics of Annual Variation in Net Carbon Dioxide Flux in a Sandy Grassland Ecosystem during Dry Years. Acta Prataculturae Sin. 2018, 27, 215–221. [Google Scholar] [CrossRef]
- Li, S.-G.; Asanuma, J.; Eugster, W.; Kotani, A.; Liu, J.-J.; Urano, T.; Oikawa, T.; Davaa, G.; Oyunbaatar, D.; Sugita, M. Net Ecosystem Carbon Dioxide Exchange over Grazed Steppe in Central Mongolia. Glob. Chang. Biol. 2005, 11, 1941–1955. [Google Scholar] [CrossRef]
- De la Motte, L.G.; Jérôme, E.; Mamadou, O.; Beckers, Y.; Bodson, B.; Heinesch, B.; Aubinet, M. Carbon Balance of an Intensively Grazed Permanent Grassland in Southern Belgium. Agric. For. Meteorol. 2016, 228–229, 370–383. [Google Scholar] [CrossRef]
- Hirata, R.; Miyata, A.; Mano, M.; Shimizu, M.; Arita, T.; Kouda, Y.; Matsuura, S.; Niimi, M.; Saigusa, T.; Mori, A.; et al. Carbon Dioxide Exchange at Four Intensively Managed Grassland Sites across Different Climate Zones of Japan and the Influence of Manure Application on Ecosystem Carbon and Greenhouse Gas Budgets. Agric. For. Meteorol. 2013, 177, 57–68. [Google Scholar] [CrossRef]
- Allard, V.; Soussana, J.F.; Falcimagne, R.; Berbigier, P.; Bonnefond, J.M.; Ceschia, E.; D’hour, P.; Hénault, C.; Laville, P.; Martin, C.; et al. The Role of Grazing Management for the Net Biome Productivity and Greenhouse Gas Budget (CO2, N2O and CH4) of Semi-Natural Grassland. Agric. Ecosyst. Environ. 2007, 121, 47–58. [Google Scholar] [CrossRef]
- Gilmanov, T.G.; Soussana, J.F.; Aires, L.; Allard, V.; Ammann, C.; Balzarolo, M.; Barcza, Z.; Bernhofer, C.; Campbell, C.L.; Cernusca, A.; et al. Partitioning European Grassland Net Ecosystem CO2 Exchange into Gross Primary Productivity and Ecosystem Respiration Using Light Response Function Analysis. Agric. Ecosyst. Environ. 2007, 121, 93–120. [Google Scholar] [CrossRef]
- Klumpp, K.; Bloor, J.M.G.; Ambus, P.; Soussana, J.-F. Effects of Clover Density on N2O Emissions and Plant-Soil N Transfers in a Fertilised Upland Pasture. Plant Soil 2011, 343, 97–107. [Google Scholar] [CrossRef]
- Wall, A.M.; Campbell, D.I.; Morcom, C.P.; Mudge, P.L.; Schipper, L.A. Quantifying Carbon Losses from Periodic Maize Silage Cropping of Permanent Temperate Pastures. Agric. Ecosyst. Environ. 2020, 301, 107048. [Google Scholar] [CrossRef]
- Rutledge, S.; Wall, A.M.; Mudge, P.L.; Troughton, B.; Campbell, D.I.; Pronger, J.; Joshi, C.; Schipper, L.A. The Carbon Balance of Temperate Grasslands Part II: The Impact of Pasture Renewal via Direct Drilling. Agric. Ecosyst. Environ. 2017, 239, 132–142. [Google Scholar] [CrossRef]
- Hunt, J.E.; Laubach, J.; Barthel, M.; Fraser, A.; Phillips, R.L. Carbon Budgets for an Irrigated Intensively Grazed Dairy Pasture and an Unirrigated Winter-Grazed Pasture. Biogeosciences 2016, 13, 2927–2944. [Google Scholar] [CrossRef]
- Campbell, D.I.; Wall, A.M.; Nieveen, J.P.; Schipper, L.A. Variations in CO2 Exchange for Dairy Farms with Year-Round Rotational Grazing on Drained Peatlands. Agric. Ecosyst. Environ. 2015, 202, 68–78. [Google Scholar] [CrossRef]
- Mudge, P.L.; Wallace, D.F.; Rutledge, S.; Campbell, D.I.; Schipper, L.A.; Hosking, C.L. Carbon Balance of an Intensively Grazed Temperate Pasture in Two Climatically: Contrasting Years. Agric. Ecosyst. Environ. 2011, 144, 271–280. [Google Scholar] [CrossRef]
- Nieveen, J.P.; Campell, D.I.; Schipper, L.A.; Blair, I.J. Carbon Exchange of Grazed Pasture on Drained Peat Soil. Glob. Chang. Biol. 2005, 11, 607–618. [Google Scholar] [CrossRef]
- Bezerra, B.G.; Santos e Silva, C.M.; Mendes, K.R.; Mutti, P.R.; Fernandes, L.S.; Marques, T.V.; Câmara e Silva, C.L.; Campos, S.; de Lima Vieira, M.M.; Urbano, S.A.; et al. CO2 Exchanges and Evapotranspiration of a Grazed Pasture under Tropical Climate Conditions. Agric. For. Meteorol. 2022, 323, 109088. [Google Scholar] [CrossRef]
Figure 1.
Location of the Flux station (38°6′56.71″ N, 84°53′22.91″ W), Franklin County, Kentucky. Inset map showing Kentucky State with the arrow pointing to the study site.
Figure 1.
Location of the Flux station (38°6′56.71″ N, 84°53′22.91″ W), Franklin County, Kentucky. Inset map showing Kentucky State with the arrow pointing to the study site.
Figure 2.
Schematic representation of the EC data flow and analysis performed at the study site for the years under study.
Figure 2.
Schematic representation of the EC data flow and analysis performed at the study site for the years under study.
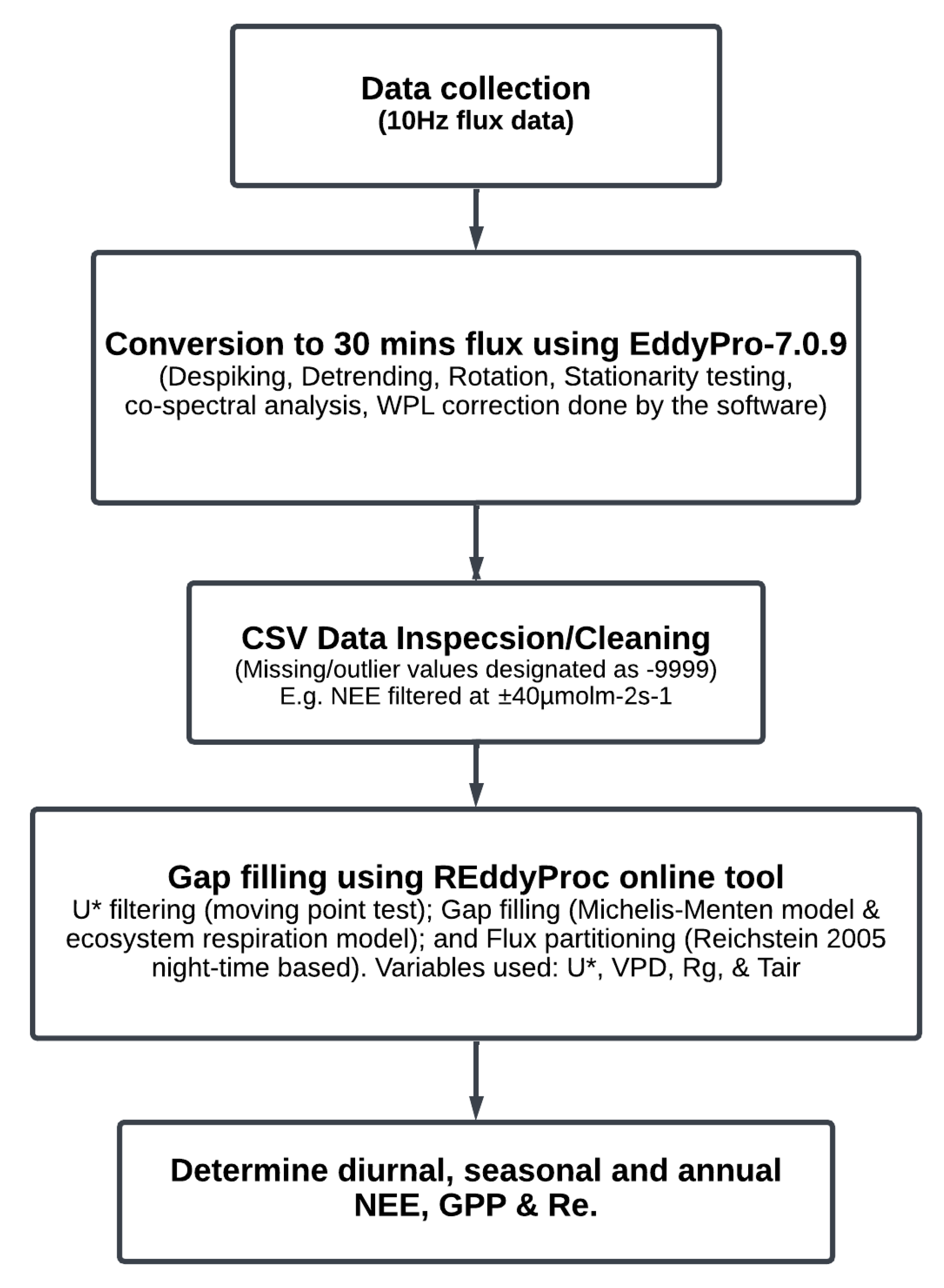
Figure 3.
Average monthly temperature (°C) under study (2015–2020) and the historical (2008–2022) monthly average.
Figure 3.
Average monthly temperature (°C) under study (2015–2020) and the historical (2008–2022) monthly average.

Figure 4.
Air temperature (°C) for all years under study (2015–2020) and the historical (2008–2022) yearly average.
Figure 4.
Air temperature (°C) for all years under study (2015–2020) and the historical (2008–2022) yearly average.
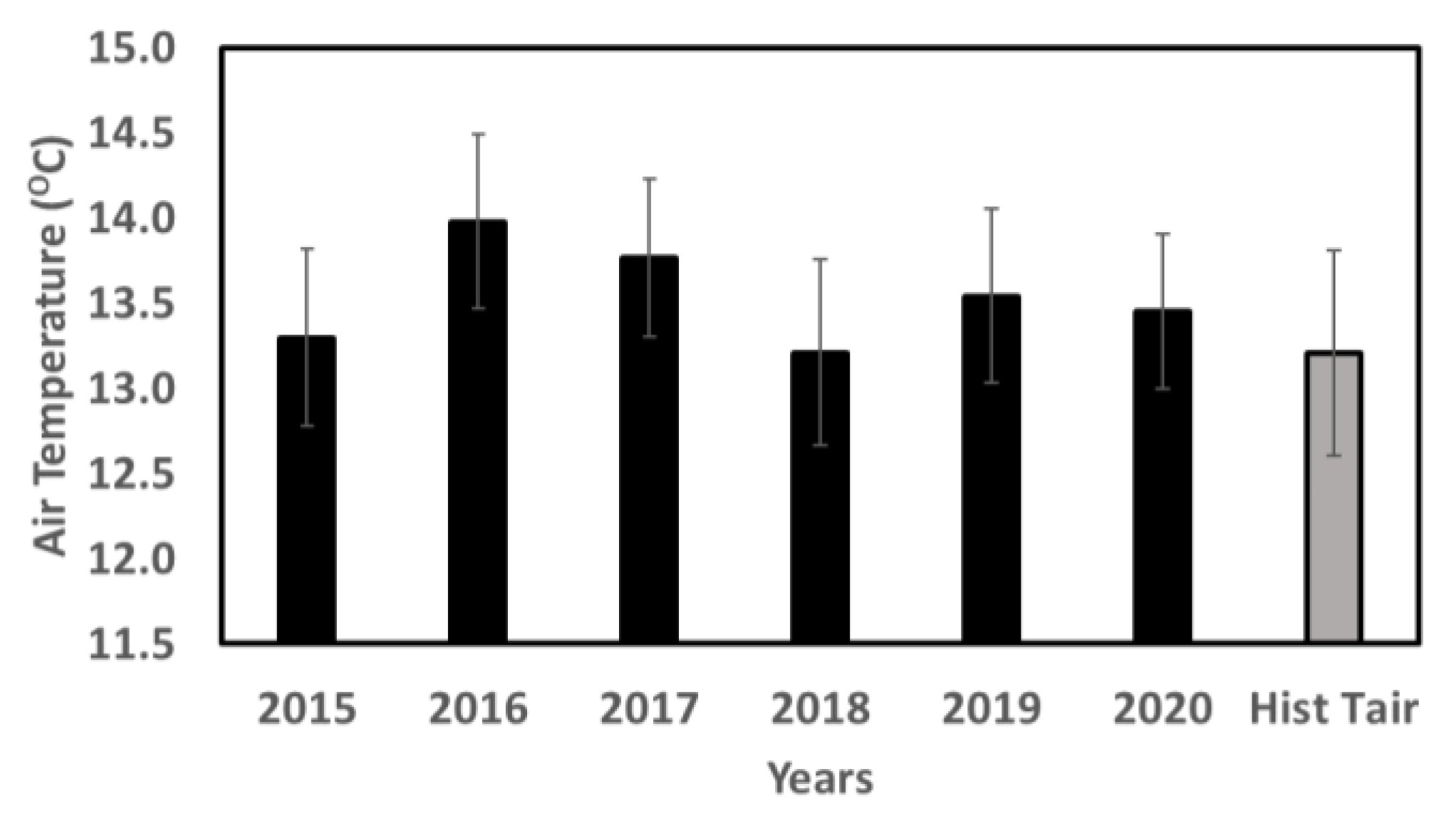
Figure 5.
Average monthly precipitation (mm) under study (2015–2020) and the historical (2008–2022) monthly average.
Figure 5.
Average monthly precipitation (mm) under study (2015–2020) and the historical (2008–2022) monthly average.

Figure 6.
Total precipitation (mm) for all years under study (2015–2020) shown in black bars and the historical (2008–2022) yearly average shown in grey-colored bar.
Figure 6.
Total precipitation (mm) for all years under study (2015–2020) shown in black bars and the historical (2008–2022) yearly average shown in grey-colored bar.

Figure 7.
Monthly response of Net Ecosystem Exchange (NEE) to photosynthetic photon flux density (PPFD) for the year 2016. Note that negative NEE indicates Carbon uptake by the pasture ecosystem. The empty black circles represent the observed NEE (g Cm−2) while black lines stand for the fitted curves of NEE to PPFD.
Figure 7.
Monthly response of Net Ecosystem Exchange (NEE) to photosynthetic photon flux density (PPFD) for the year 2016. Note that negative NEE indicates Carbon uptake by the pasture ecosystem. The empty black circles represent the observed NEE (g Cm−2) while black lines stand for the fitted curves of NEE to PPFD.
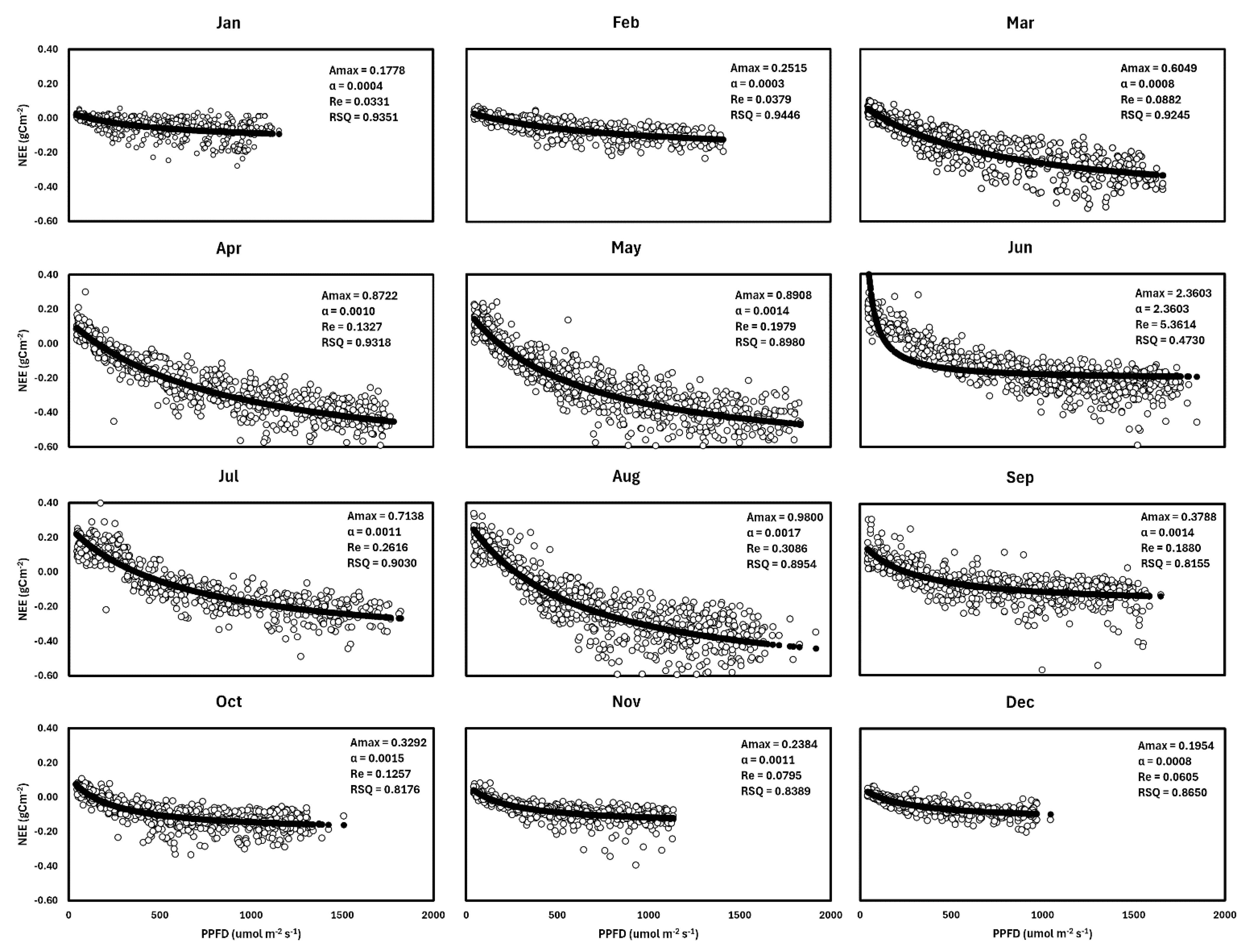
Figure 8.
Diurnal courses of net ecosystem exchange of C (NEE), ecosystem respiration (Re), and gross primary productivity (GPP) for all seasons (a–c) for DOY–43, the non-growing season; DOY–135, the early growing season; DOY–200, the peak growing season, and DOY–300, late growing season) in 2016; and the average diurnal fluxes for all years measured (d–f).
Figure 8.
Diurnal courses of net ecosystem exchange of C (NEE), ecosystem respiration (Re), and gross primary productivity (GPP) for all seasons (a–c) for DOY–43, the non-growing season; DOY–135, the early growing season; DOY–200, the peak growing season, and DOY–300, late growing season) in 2016; and the average diurnal fluxes for all years measured (d–f).

Figure 9.
Monthly values of GPP, NEE, and RE from 2015 to 2020.
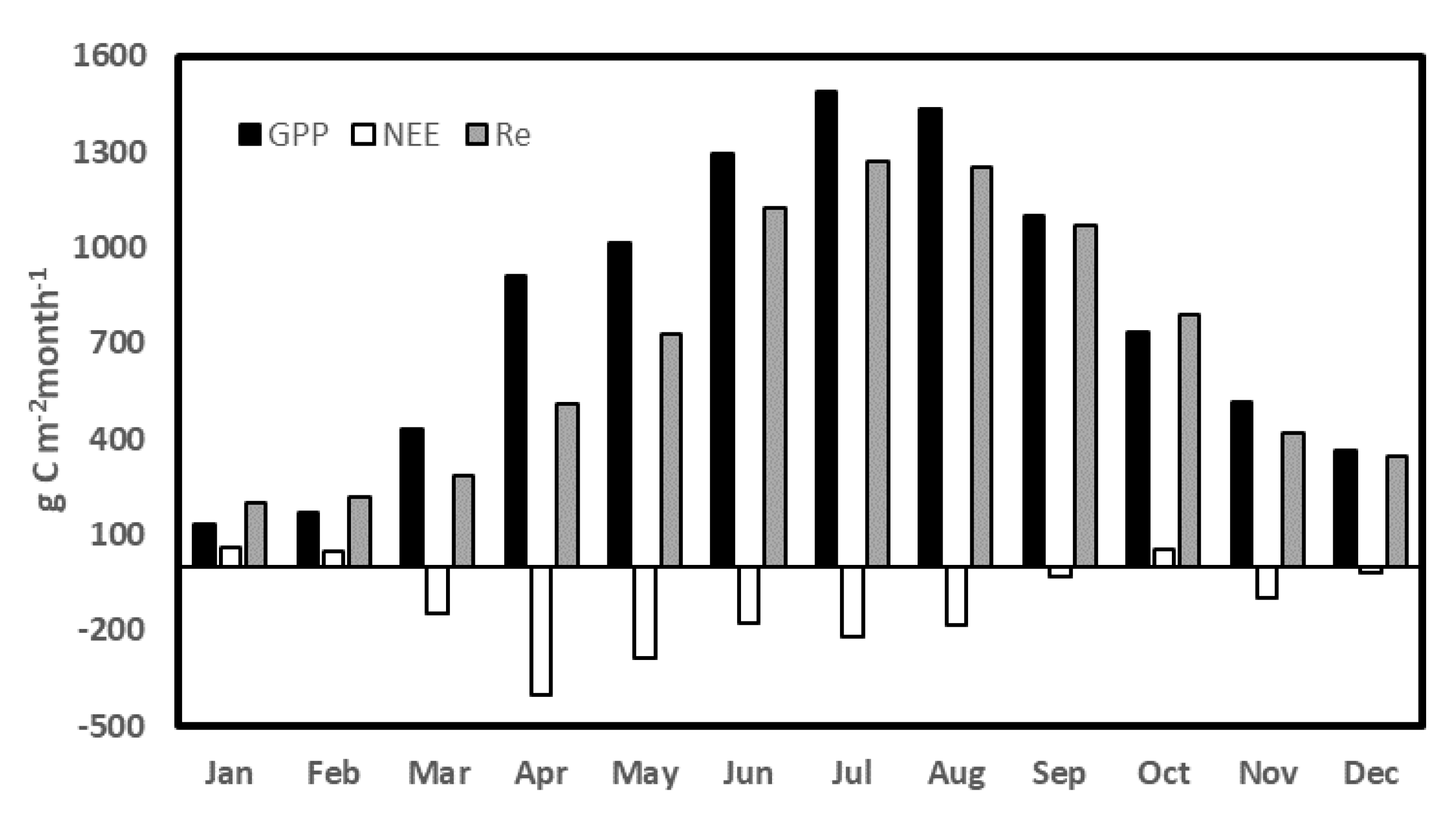
Figure 10.
Yearly total values of GPP, NEE, and RE from 2015 to 2020.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Micrometeorological Instrumentation at the Kentucky State University Research Farm.
| Measurement Parameter | Instrumentation | Model |
|---|---|---|
| H2O/CO2 EC-Flux, 10 Hz | Infrared Gas Analyzer | LI-7200, Closed Path (LI-COR Environmental GmbH, Bad Homburg, Germany) |
| Precipitation, total | Tipping bucket rain gauge | Texas Electronics TE525 (Texas Electronics, Dallas, TX, USA) |
| Photosynthetic Photon Flux Density (PPFD) | Quantum Sensor | LI-190R Quantum Sensor (LI-COR Environmental GmbH, Bad Homburg, Germany) |
| Soil, heat flux | Soil heat flux plate | REBS HFT (Radiation and Energy Balance Systems, Inc., Seattle, WA, USA) |
| Temperature, air | Temperature/humidity probe | Vaisala HMP 35C (Vaisala Oyj, Vantaa, Finland) |
| 3D Wind | 3D Sonic Anemometer | Gill (Gill Instruments Limited, Lymington, Hampshire, UK) |
Table 2.
Michaelis–Menten model curve-fitted parameters of the maximum ecosystem C uptake rate—Amax (g C m−2 s−1) at saturated light intensity, daytime ecosystem respiration—Re (g m−2 s−1), cumulative photosynthetic photon flux density—PPFD (kW m−2 month−1) and apparent quantum efficiency—α (g C m−2 s−1 PPFD−1).
Table 2.
Michaelis–Menten model curve-fitted parameters of the maximum ecosystem C uptake rate—Amax (g C m−2 s−1) at saturated light intensity, daytime ecosystem respiration—Re (g m−2 s−1), cumulative photosynthetic photon flux density—PPFD (kW m−2 month−1) and apparent quantum efficiency—α (g C m−2 s−1 PPFD−1).
| Amax | α | Re (gCm−2) | n | R2 | PPFD (KW/m2/Month) | |
|---|---|---|---|---|---|---|
| January | 0.1778 | 0.0004 | 0.0331 | 551 | 0.9351 | 5.80 |
| February | 0.2515 | 0.0003 | 0.0379 | 563 | 0.9446 | 6.44 |
| March | 0.6049 | 0.0008 | 0.0882 | 684 | 0.9245 | 9.49 |
| April | 0.8722 | 0.0010 | 0.1327 | 743 | 0.9318 | 13.24 |
| May | 0.8908 | 0.0014 | 0.1979 | 810 | 0.8980 | 12.85 |
| June | 2.3603 | 2.3603 | 5.3614 | 820 | 0.4730 | 16.46 |
| July | 0.7138 | 0.0011 | 0.2616 | 541 | 0.9030 | 8.97 |
| August | 0.9800 | 0.0017 | 0.3086 | 785 | 0.8954 | 13.26 |
| September | 0.3788 | 0.0014 | 0.1880 | 697 | 0.8155 | 12.01 |
| October | 0.3292 | 0.0015 | 0.1257 | 644 | 0.8176 | 9.23 |
| November | 0.2384 | 0.0011 | 0.0795 | 566 | 0.8389 | 6.96 |
| December | 0.1954 | 0.0008 | 0.0605 | 510 | 0.8650 | 4.40 |
Table 3.
GPP, RE, and NEE by year and season.
| GPP (gCm−2yr−1) | RE (gCm−2yr−1) | NEE (gCm−2yr−1) | ||||||
|---|---|---|---|---|---|---|---|---|
| Year | GS | NGS | GS | NGS | GS | NGS | NEE | Status |
| 2015 | 1411.92 | 333.87 | 1261.98 | 239.62 | −149.94 | −94.25 | −244.19 | Sink |
| 2016 | 1008.99 | 330.33 | 1079.65 | 276.52 | 70.65 | −53.80 | 16.85 | Neutral |
| 2017 | 1651.25 | 340.42 | 1498.81 | 315.99 | −152.44 | −24.43 | −176.88 | Sink |
| 2018 | 1343.22 | 220.29 | 763.50 | 264.59 | −579.72 | 44.31 | −535.41 | Sink |
| 2019 | 1731.87 | 246.13 | 1672.11 | 277.50 | −59.75 | 31.37 | −28.38 | Sink |
| 2020 | 836.14 | 148.43 | 465.32 | 93.56 | −370.83 | −54.87 | −425.69 | Sink |
| Sub Total | 7983 ± 279 | 1619 ± 165 | 6741 ± 291 | 1468 ± 90 | −1242 ± 151 | −152 ± 91 | ||
| Cumulative | 9603 ± 479 | 8209 ± 410 | −1394 ± 146 | Sink | ||||
GS = Growing Season (April–October) and NGS = Non-Growing Season (January–March, November–December).
Table 4.
Mean annual temperature, precipitation and NEE across the world.
| Continent | Station | Cropping Management | Temp. (°C) | Precip. (mm) | NEE (g C yr−1) | Study Years | Location | Reference | |
|---|---|---|---|---|---|---|---|---|---|
| N. America | i. | Harold Benson Research Farm, Frankfort, KY | Managed Pasture | 13.54 | 1369.95 | −232.3 | 2015–2020 | 38.11° N, 84.88° W | Present Study |
| ii. | SGS ecoregion of the North American Great Plains, Colorado, USA | Heavy/moderate continuous grazing | −23/−59 | 2018 | [59] | ||||
| iii. | Sardinilla, N.E. Barro Colorado Island, Panama | 261 | 2008–2009 | 9°19′ N, 79°38′ W | [60] | ||||
| iv | Pennsylvania State Uni. Haller Research Farm | 9.7 | 1014 | 14/−5 | 2008 | 40.9° N; 77.8° W | [61] | ||
| Asia | i. | Ulastai station, Tien Shan Mountains, Xinjiang Uygur Autonomous Region. | Middle Tien Shan grassland ecosystem | 7.26 | 257.61 | −89.95 | 2018 | 43°28′ N, 87°12′ E | [62] |
| ii. | Yunnan-Guizhou Plateau, China | Grassland ecosystem | −116.06 | 2017–2018 | 27°46′ N, 107°28′ E | [63] | |||
| iii. | Horqin District, Inner Mongolia, China | Grassland ecosystem | −32.91 | 2015–2018 | 42°55′ N, 12°42′ E | [64] | |||
| iv. | Kherlenbayan-Ulaan, Hentiy province, central Mongolia | −41 | 2005 | [65] | |||||
| Europe | i. | Condroz region, Belgium | −141 | 2016 | [66] | ||||
| ii. | Grassland at Nakashibetsu, Shizunai, Nasushiobara, and Kobayashi., Japan | −245/−158 | 2013 | [67] | |||||
| iii. | Laqueuille, Puy de Döme, France | 1.32 | 2007 | [68] | |||||
| iv. | European grazed grassland | −32 | 2007 | [69] | |||||
| v. | Seminatural grassland site in Auvergne, ‘Laqueuille’, France | Extensively managed paddock | 7.07 | 997.5 | −225 | 2003–2008 | 45°38′ N, 2°44′ E | [70] | |
| vi. | “ | Intensively managed paddock | 7.07 | 997.5 | −209 | 2003–2008 | 45°38′ N, 2°44′ E | ‘’ | |
| Oceania | i. | Waikato region, New Zealand | Rotationally grazed pasture | 13.3 | 1249 | −114 | 2018 | 37°45.6′ S, 175°47.2′ E | [71] |
| ii. | “ | “ | 13.3 | 1249 | 11 | 2017 | 37°45.6′ S, 175°47.2′ E | ‘’ | |
| iii. | Troughton Farm, Waikato region, North Island, New Zealand | old pasture/renewed pasture | −187/−148 | 2017 | 37°45.6′ S, 175°47.2′ E | [72] | |||
| iv. | Canterbury, New Zealand | dryland/irrigated | −20/−408 | 2016 | 43.7542° S, 171.1637° E | [73] | |||
| v. | Hamilton Basin, Waikato region, North Island/Te Ika-a-Maui, New Zealand | Intense, year-round rotational grazing | 190 | 2015 | [74] | ||||
| vi. | Scott Farm, Waikato region, North Island, New Zealand | −195 | 2011 | [75] | |||||
| vii. | Rukuhia peatland, Waikato region, New Zealand | 4 | 2005 | [76] | |||||
| S. America | i. | Experimental station, GEFORP -EAJ-UFRN, Northeast coast of Brazil, Macaíba district, Brazil | Rotational grazing | 25.5 | 1100 | −215 ± 22 | 2015–2017 | 5◦53′34″ S. 35◦21′50″ W | [77] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Familusi, I.; Gebremedhin, M.; Ries, I.; Brown, J.; Gyawali, B. The Productivity and Carbon Exchange of an Intensively Managed Pasture in Central Kentucky. Atmosphere 2024, 15, 348. https://doi.org/10.3390/atmos15030348
AMA Style
Familusi I, Gebremedhin M, Ries I, Brown J, Gyawali B. The Productivity and Carbon Exchange of an Intensively Managed Pasture in Central Kentucky. Atmosphere. 2024; 15(3):348. https://doi.org/10.3390/atmos15030348
Chicago/Turabian StyleFamilusi, Ife, Maheteme Gebremedhin, Ian Ries, Jacob Brown, and Buddhi Gyawali. 2024. "The Productivity and Carbon Exchange of an Intensively Managed Pasture in Central Kentucky" Atmosphere 15, no. 3: 348. https://doi.org/10.3390/atmos15030348
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.