Separated Siamese Twins: Intronic Small Nucleolar RNAs and Matched Host Genes May be Altered in Conjunction or Separately in Multiple Cancer Types



Abstract
:1. Introduction
2. Materials and Methods
2.1. TCGA Data
2.2. Copy Number Alterations and Gene Expression Data
2.3. Statistical Analyses
3. Results
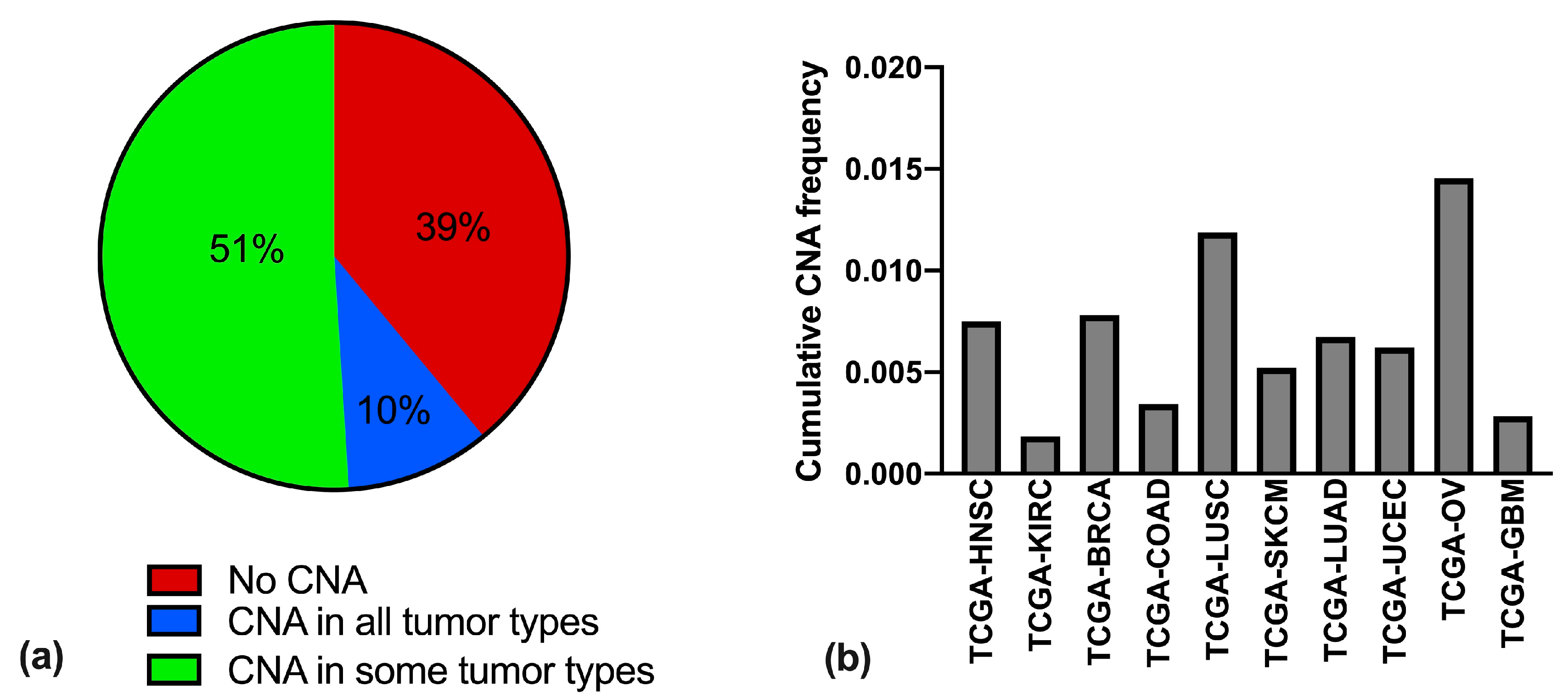
3.1. snoRNAs and snoRNA Host Genes Alterations Vary Significantly in Different Cancer Types
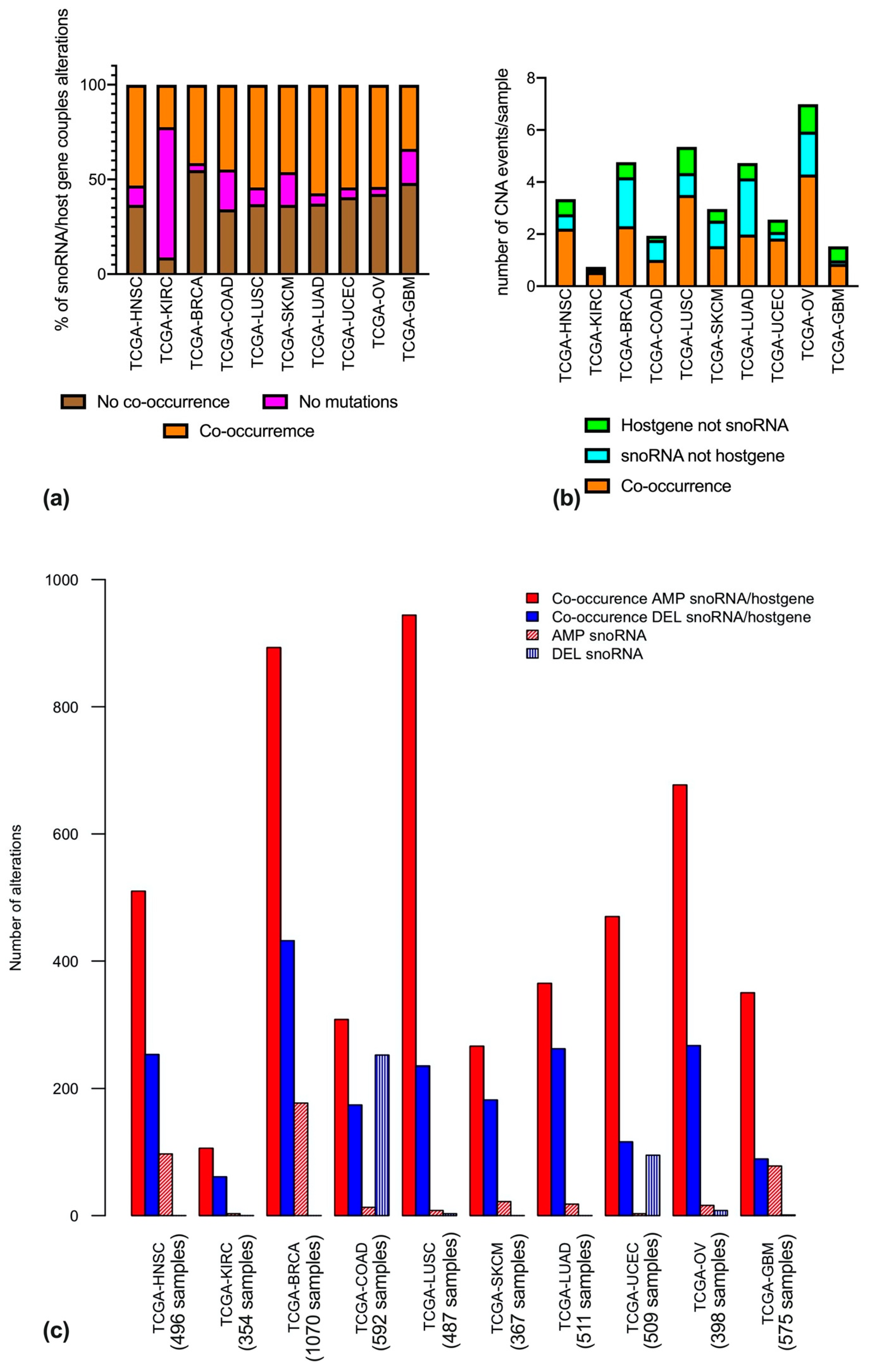
3.2. Copy Number Alterations of snoRNAs and Matched Host Genes are Not Always Coupled
3.3. Frequently Altered snoRNA-Host Gene Couples Recur in Different Cancer Types
3.4. Differential ExpressionAalterations of snoRNAs and Host Genes in Multiple Cancer Types
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Smith, C.M.; Steitz, J.A. Classification of gas5 as a Multi-Small-Nucleolar-RNA (snoRNA) Host Gene and a Member of the 5′-Terminal Oligopyrimidine Gene Family Reveals Common Features of snoRNA Host Genes. Mol. Cell. Biol. 1998, 18, 6897–6909. [Google Scholar] [CrossRef] [Green Version]
- Pelczar, P.; Filipowicz, W. The Host Gene for Intronic U17 Small Nucleolar RNAs in Mammals Has No Protein-Coding Potential and Is a Member of the 5′-Terminal Oligopyrimidine Gene Family. Mol. Cell. Biol. 1998, 18, 4509–4518. [Google Scholar] [CrossRef] [Green Version]
- Meyuhas, O.; Kahan, T. The race to decipher the top secrets of TOP mRNAs. Biochim. Biophys. Acta 2015, 1849, 801–811. [Google Scholar] [CrossRef]
- Filipowicz, W.; Pogacić, V. Biogenesis of small nucleolar ribonucleoproteins. Curr. Opin. Cell Biol. 2002, 14, 319–327. [Google Scholar] [CrossRef]
- Dieci, G.; Preti, M.; Montanini, B. Eukaryotic snoRNAs: A paradigm for gene expression flexibility. Genomics 2009, 94, 83–88. [Google Scholar] [CrossRef] [Green Version]
- Penzo, M.; Galbiati, A.; Treré, D.; Montanaro, L. The importance of being (slightly) modified: The role of rRNA editing on gene expression control and its connections with cancer. Biochim. Biophys. Acta Rev. Cancer 2016, 1866, 330–338. [Google Scholar] [CrossRef]
- Penzo, M.; Guerrieri, A.N.; Zacchini, F.; Treré, D.; Montanaro, L. RNA Pseudouridylation in Physiology and Medicine: For Better and for Worse. Genes 2017, 8, 301. [Google Scholar] [CrossRef] [Green Version]
- Lo Monaco, P.; Marcel, V.; Diaz, J.J.; Catez, F. 2′-O-methylation of ribosomal RNA: Towards an epitranscriptomic control of translation? Biomolecules 2018, 8, 106. [Google Scholar] [CrossRef] [Green Version]
- Jorjani, H.; Kehr, S.; Jedlinski, D.J.; Gumienny, R.; Hertel, J.; Stadler, P.F.; Zavolan, M.; Gruber, A.R. An updated human snoRNAome. Nucleic Acids Res. 2016, 44, 5068–5082. [Google Scholar] [CrossRef]
- Dudnakova, T.; Dunn-Davies, H.; Peters, R.; Tollervey, D. Mapping targets for small nucleolar RNAs in yeast. Wellcome Open Res. 2018, 3, 120. [Google Scholar] [CrossRef]
- Taft, R.J.; Glazov, E.A.; Lassmann, T.; Hayashizaki, Y.; Carninci, P.; Mattick, J.S. Small RNAs derived from snoRNAs. RNA 2009, 15, 1233–1240. [Google Scholar] [CrossRef] [Green Version]
- Williams, G.T.; Farzaneh, F. Are snoRNAs and snoRNA host genes new players in cancer? Nat. Rev. Cancer 2012, 12, 84–88. [Google Scholar] [CrossRef]
- Mannoor, K.; Liao, J.; Jiang, F. Small nucleolar RNAs in cancer. Biochim. Biophys. Acta 2012, 1826, 121–128. [Google Scholar] [CrossRef] [Green Version]
- McMahon, M.; Contreras, A.; Ruggero, D. Small RNAs with big implications: New insights into H/ACA snoRNA function and their role in human disease. Wiley Interdiscip. Rev. RNA 2015, 6, 173–189. [Google Scholar] [CrossRef] [Green Version]
- Bratkovič, T.; Božič, J.; Rogelj, B. Functional diversity of small nucleolar RNAs. Nucleic Acids Res. 2019. [Google Scholar] [CrossRef] [Green Version]
- Mei, Y.-P.; Liao, J.-P.; Shen, J.; Yu, L.; Liu, B.-L.; Liu, L.; Li, R.-Y.; Ji, L.; Dorsey, S.G.; Jiang, Z.-R.; et al. Small nucleolar RNA 42 acts as an oncogene in lung tumorigenesis. Oncogene 2012, 31, 2794–2804. [Google Scholar] [CrossRef]
- Soeno, Y.; Fujita, K.; Kudo, T.; Asagiri, M.; Kakuta, S.; Taya, Y.; Shimazu, Y.; Sato, K.; Tanaka-Fujita, R.; Kubo, S.; et al. Generation of a Mouse Model with Down-Regulated U50 snoRNA (SNORD50) Expression and Its Organ-Specific Phenotypic Modulation. PLoS ONE 2013, 8, e72105. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Han, L.; Wei, J.; Zhang, K.; Shi, Z.; Duan, R.; Li, S.; Zhou, X.; Pu, P.; Zhang, J.; et al. SNORD76, a box C/D snoRNA, acts as a tumor suppressor in glioblastoma. Sci. Rep. 2015, 5, 8588. [Google Scholar] [CrossRef] [Green Version]
- Gong, J.; Li, Y.; Liu, C.-J.; Xiang, Y.; Li, C.; Ye, Y.; Zhang, Z.; Hawke, D.H.; Park, P.K.; Diao, L.; et al. A Pan-cancer Analysis of the Expression and Clinical Relevance of Small Nucleolar RNAs in Human Cancer. Cell Rep. 2017, 21, 1968–1981. [Google Scholar] [CrossRef] [Green Version]
- Dong, X.-Y.; Guo, P.; Boyd, J.; Sun, X.; Li, Q.; Zhou, W.; Dong, J.-T. Implication of snoRNA U50 in human breast cancer. J. Genet. Genomics 2009, 36, 447–454. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, R.; Satoh, H.; Moriyama, M.; Satoh, K.; Morishita, Y.; Yoshida, S.; Watanabe, T.; Nakamura, Y.; Mori, S. Intronic U50 small-nucleolar-RNA (snoRNA) host gene of no protein-coding potential is mapped at the chromosome breakpoint t(3;6)(q27;q15) of human B-cell lymphoma. Genes Cells 2000, 5, 277–287. [Google Scholar] [CrossRef] [PubMed]
- Testa, J.R.; Liu, Z.; Feder, M.; Bell, D.W.; Balsara, B.; Cheng, J.Q.; Taguchi, T. Advances in the analysis of chromosome alterations in human lung carcinomas. Cancer Genet. Cytogenet. 1997, 95, 20–32. [Google Scholar] [CrossRef]
- Cerami, E.; Gao, J.; Dogrusoz, U.; Gross, B.E.; Sumer, S.O.; Aksoy, B.A.; Jacobsen, A.; Byrne, C.J.; Heuer, M.L.; Larsson, E.; et al. The cBio Cancer Genomics Portal: An Open Platform for Exploring Multidimensional Cancer Genomics Data: Figure 1. Cancer Discov. 2012, 2, 401–404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, J.; Aksoy, B.A.; Dogrusoz, U.; Dresdner, G.; Gross, B.; Sumer, S.O.; Sun, Y.; Jacobsen, A.; Sinha, R.; Larsson, E.; et al. Integrative analysis of complex cancer genomics and clinical profiles using the cBioPortal. Sci. Signal. 2013, 6, pl1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mermel, C.H.; Schumacher, S.E.; Hill, B.; Meyerson, M.L.; Beroukhim, R.; Getz, G. GISTIC2.0 facilitates sensitive and confident localization of the targets of focal somatic copy-number alteration in human cancers. Genome Biol. 2011, 12, R41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [Green Version]
- Bouchard-Bourelle, P.; Desjardins-Henri, C.; Mathurin-St-Pierre, D.; Deschamps-Francoeur, G.; Fafard-Couture, É.; Garant, J.-M.; Elela, S.A.; Scott, M.S. snoDB: An interactive database of human snoRNA sequences, abundance and interactions. Nucleic Acids Res. 2019, 48, D220–D225. [Google Scholar] [CrossRef]
- Zack, T.I.; Schumacher, S.E.; Carter, S.L.; Cherniack, A.D.; Saksena, G.; Tabak, B.; Lawrence, M.S.; Zhang, C.Z.; Wala, J.; Mermel, C.H.; et al. Pan-cancer patterns of somatic copy number alteration. Nat. Genet. 2013, 45, 1134–1140. [Google Scholar] [CrossRef] [Green Version]
- Heitzer, E.; Ulz, P.; Geigl, J.B.; Speicher, M.R. Non-invasive detection of genome-wide somatic copy number alterations by liquid biopsies. Mol. Oncol. 2016, 10, 494–502. [Google Scholar] [CrossRef] [Green Version]
- Santarius, T.; Shipley, J.; Brewer, D.; Stratton, M.R.; Cooper, C.S. A census of amplified and overexpressed human cancer genes. Nat. Rev. Cancer 2010, 10, 59–64. [Google Scholar] [CrossRef]
- Lee, J.A.; Madrid, R.E.; Sperle, K.; Ritterson, C.M.; Hobson, G.M.; Garbern, J.; Lupski, J.R.; Inoue, K. Spastic paraplegia type 2 associated with axonal neuropathy and apparentPLP1 position effect. Ann. Neurol. 2006, 59, 398–403. [Google Scholar] [CrossRef] [PubMed]
- Stranger, B.E.; Forrest, M.S.; Dunning, M.; Ingle, C.E.; Beazlsy, C.; Thorne, N.; Redon, R.; Bird, C.P.; De Grassi, A.; Lee, C.; et al. Relative impact of nucleotide and copy number variation on gene phenotypes. Science 2007, 315, 848–853. [Google Scholar] [CrossRef] [Green Version]
- Penzo, M.; Montanaro, L.; Treré, D.; Derenzini, M. The Ribosome Biogenesis—Cancer Connection. Cells 2019, 8, 55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guimaraes, J.C.; Zavolan, M. Patterns of ribosomal protein expression specify normal and malignant human cells. Genome Biol. 2016, 17, 236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hassan, M.K.; Kumar, D.; Naik, M.; Dixit, M. The expression profile and prognostic significance of eukaryotic translation elongation factors in different cancers. PLoS ONE 2018, 13. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.U.; Ur Rahman, M.S.; Jia, Z.; Jiang, C. Eukaryotic translation initiation factors and cancer. Tumor Biol. 2017, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montanaro, L. Dyskerin and cancer: More than telomerase. The defect in mRNA translation helps in explaining how a proliferative defect leads to cancer. J. Pathol. 2010, 222, 345–349. [Google Scholar] [CrossRef] [PubMed]
- Marcel, V.; Ghayad, S.E.; Belin, S.; Therizols, G.; Morel, A.-P.; Solano-Gonzàlez, E.; Vendrell, J.A.; Hacot, S.; Mertani, H.C.; Albaret, M.A.; et al. p53 acts as a safeguard of translational control by regulating fibrillarin and rRNA methylation in cancer. Cancer Cell 2013, 24, 318–330. [Google Scholar] [CrossRef] [Green Version]
- Dong, D.; Mu, Z.; Zhao, C.; Sun, M. ZFAS1: A novel tumor-related long non-coding RNA. Cancer Cell Int. 2018, 18, 125. [Google Scholar] [CrossRef]
- Ji, J.; Dai, X.; Yeung, S.C.J.; He, X. The role of long non-coding RNA GAS5 in cancers. Cancer Manag. Res. 2019, 11, 2729–2737. [Google Scholar] [CrossRef] [Green Version]
- Terashima, M.; Ishimura, A.; Wanna-udom, S.; Suzuki, T. MEG8 long noncoding RNA contributes to epigenetic progression of the epithelial-mesenchymal transition of lung and pancreatic cancer cells. J. Biol. Chem. 2018, 293, 18016–18030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tran, H.V.U.; Kiemer, A.K.; Helms, V. Copy number alterations in tumor genomes deleting antineoplastic drug targets partially compensated by complementary amplifications. Cancer Genom. Proteom. 2018, 15, 365–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weber, M.J. Mammalian small nucleolar RNAs are mobile genetic elements. PLoS Genet. 2006, 2, 1984–1997. [Google Scholar] [CrossRef] [PubMed]
- Comtesse, N.; Keller, A.; Diesinger, I.; Bauer, C.; Kayser, K.; Huwer, H.; Lenhof, H.-P.; Meese, E. Frequent overexpression of the genes FXR1, CLAPM1 and EIF4G located on amplicon 3q26-27 in squamous cell carcinoma of the lung. Int. J. Cancer 2007, 120, 2538–2544. [Google Scholar] [CrossRef] [PubMed]
- Shi, Z.Z.; Jiang, Y.Y.; Hao, J.J.; Zhang, Y.; Zhang, T.T.; Shang, L.; Liu, S.G.; Shi, F.; Wang, M.R. Identification of putative target genes for amplification within 11q13.2 and 3q27.1 in esophageal squamous cell carcinoma. Clin. Transl. Oncol. 2014, 16, 606–615. [Google Scholar] [CrossRef]
- Lin, S.-C.; Liu, C.-J.; Ko, S.-Y.; Chang, H.-C.; Liu, T.-Y.; Chang, K.-W. Copy number amplification of 3q26-27 oncogenes in microdissected oral squamous cell carcinoma and oral brushed samples from areca chewers. J. Pathol. 2005, 206, 417–422. [Google Scholar] [CrossRef]
- Karube, K.; Ying, G.; Tagawa, H.; Niino, D.; Aoki, R.; Kimura, Y.; Hashikawa, K.; Suefuji, N.; Sugita, Y.; Nomura, Y.; et al. BCL6 gene amplification/3q27 gain is associated with unique clinicopathological characteristics among follicular lymphoma without BCL2 gene translocation. Mod. Pathol. 2008, 21, 973–978. [Google Scholar] [CrossRef] [Green Version]
- Subramonia-Iyer, S.; Hrinczenko, B. A review of chromosome 3q abnormalities in lung cancer. J. Clin. Oncol. 2017. [Google Scholar] [CrossRef]
- Bulygin, K.N.; Bartuli, Y.S.; Malygin, A.A.; Graifer, D.M.; Frolova, L.Y.; Karpova, G.G. Chemical footprinting reveals conformational changes of 18S and 28S rRNAs at different steps of translation termination on the human ribosome. RNA 2016, 22, 278–289. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.-H.; Qi, J.-J.; Wu, Q.-N.; Lu, J.-H.; Liu, Z.-X.; Wang, Y.; Hu, P.-S.; Li, T.; Lin, J.-F.; Wu, X.-Y.; et al. Eukaryotic initiation factor 4A2 promotes experimental metastasis and oxaliplatin resistance in colorectal cancer. J. Exp. Clin. Cancer Res. 2019, 38, 196. [Google Scholar] [CrossRef]
- Shaoyan, X.; Juanjuan, Y.; Yalan, T.; Ping, H.; Jianzhong, L.; Qinian, W. Downregulation of EIF4A2 in non-small-cell lung cancer associates with poor prognosis. Clin. Lung Cancer 2013, 14, 658–665. [Google Scholar] [CrossRef]
- Lu, W.; Tao, X.; Fan, Y.; Tang, Y.; Xu, X.; Fan, S.; Huang, Y.; Yu, Y.; Luo, D. LINC00888 promoted tumorigenicity of melanoma via miR-126/CRK signaling axis. Onco. Targets. Ther. 2018, 11, 4431–4442. [Google Scholar] [CrossRef] [Green Version]
- Antony, A.C.; Ram, A.K.; Dutta, K.; Alone, P.V. Ribosomal mutation in helix 32 of 18S rRNA alters fidelity of eukaryotic translation start site selection. FEBS Lett. 2019, 593, 852–867. [Google Scholar] [CrossRef] [PubMed]
- Su, J.; Liao, J.; Gao, L.; Shen, J.; Guarnera, M.A.; Zhan, M.; Fang, H.B.; Stass, S.A.; Jiang, F. Analysis of small nucleolar RNAs in sputum for lung cancer diagnosis. Oncotarget 2016, 7, 5131–5142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Badura, M.; Braunstein, S.; Zavadil, J.; Schneider, R.J. DNA damage and eIF4G1 in breast cancer cells reprogram translation for survival and DNA repair mRNAs. Proc. Natl. Acad. Sci. USA 2012, 109, 18767–18772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Luo, Q.; Xie, Z.; Li, G.; Mao, C.; Liu, Y.; Wen, X.; Yin, N.; Cao, J.; Wang, J.; et al. Characterization of the Expression of the RNA Binding Protein eIF4G1 and Its Clinicopathological Correlation with Serous Ovarian Cancer. PLoS ONE 2016, 11, e0163447. [Google Scholar] [CrossRef] [Green Version]
- Cao, Y.; Wei, M.; Li, B.; Liu, Y.; Lu, Y.; Tang, Z.; Lu, T.; Yin, Y.; Qin, Z.; Xu, Z. Functional role of eukaryotic translation initiation factor 4 gamma 1 (EIF4G1) in NSCLC. Oncotarget 2016, 7, 24242–24251. [Google Scholar] [CrossRef] [Green Version]
- Goh, T.S.; Ha, M.; Lee, J.S.; Jeong, D.C.; Jung, E.S.; Han, M.E.; Kim, Y.H.; Oh, S.O. Prognostic significance of EIF4G1 in patients with pancreatic ductal adenocarcinoma. Onco. Targets. Ther. 2019, 12, 2853–2859. [Google Scholar] [CrossRef] [Green Version]
- Ying, Z.; Tian, H.; Li, Y.; Lian, R.; Li, W.; Wu, S.; Zhang, H.Z.; Wu, J.; Liu, L.; Song, J.; et al. CCT6A suppresses SMAD2 and promotes prometastatic TGF-β signaling. J. Clin. Investig. 2017, 127, 1725–1740. [Google Scholar] [CrossRef] [Green Version]
- Zeng, G.; Wang, J.; Huang, Y.; Lian, Y.; Chen, D.; Wei, H.; Lin, C.; Huang, Y. Overexpressing CCT6A contributes to cancer cell growth by affecting the G1-To-S phase transition and predicts a negative prognosis in hepatocellular carcinoma. Onco. Targets. Ther. 2019, 12, 10427–10439. [Google Scholar] [CrossRef] [Green Version]
- Curry, J.M.; Tuluc, M.; Whitaker-Menezes, D.; Ames, J.A.; Anantharaman, A.; Butera, A.; Leiby, B.; Cognetti, D.M.; Sotgia, F.; Lisanti, M.P.; et al. Cancer metabolism, stemness and tumor recurrence : MCT1 and MCT4 are functional biomarkers of metabolic symbiosis in head and neck cancer. Cell Cycle 2013, 12, 1371–1384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Curry, J.M.; Tassone, P.; Cotzia, P.; Sprandio, J.; Luginbuhl, A.; Cognetti, D.M.; Mollaee, M.; Domingo-Vidal, M.; Pribitkin, E.A.; Keane, W.M.; et al. Multicompartment metabolism in papillary thyroid cancer. Laryngoscope 2016, 126, 2410–2418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Nues, R.W.; Venema, J.; Planta, R.J.; Raué, H.A. Variable region V1 of Saccharomyces cerevisiae 18S rRNA participates in biogenesis and function of the small ribosomal subunit. Chromosoma 1997, 105, 523–531. [Google Scholar] [CrossRef] [PubMed]
- Vaarala, M.H.; Porvari, K.S.; Kyllönen, A.P.; Mustonen, M.V.; Lukkarinen, O.; Vihko, P.T. Several genes encoding ribosomal proteins are over-expressed in prostate-cancer cell lines: Confirmation of L7a and L37 over-expression in prostate-cancer tissue samples. Int. J. Cancer 1998, 78, 27–32. [Google Scholar] [CrossRef]
- Sloma, M.S.; Nygård, O. Possible interaction sites of mRNA, tRNA, translation factors and the nascent peptide in 5S, 5.8S and 28S rRNA in in vivo assembled eukaryotic ribosomal complexes. Biochim. Biophys. Acta Gene Struct. Expr. 2001, 1521, 30–38. [Google Scholar] [CrossRef]
- Penzo, M.; Ludovini, V.; Treré, D.; Siggillino, A.; Vannucci, J.; Bellezza, G.; Crinò, L.; Montanaro, L. Dyskerin and TERC expression may condition survival in lung cancer patients. Oncotarget 2015, 6, 21755–21760. [Google Scholar] [CrossRef] [PubMed]
- Montanaro, L.; Brigotti, M.; Clohessy, J.; Barbieri, S.; Ceccarelli, C.; Santini, D.; Taffurelli, M.; Calienni, M.; Teruya-Feldstein, J.; Trerè, D.; et al. Dyskerin expression influences the level of ribosomal RNA pseudo-uridylation and telomerase RNA component in human breast cancer. J. Pathol. 2006, 210, 10–18. [Google Scholar] [CrossRef]
- Penzo, M.; Rocchi, L.; Brugiere, S.; Carnicelli, D.; Onofrillo, C.; Coute, Y.; Brigotti, M.; Montanaro, L. Human ribosomes from cells with reduced dyskerin levels are intrinsically altered in translation. FASEB J. 2015, 29, 3472–3482. [Google Scholar] [CrossRef] [Green Version]
- Cheng, J.; Kellner, N.; Berninghausen, O.; Hurt, E.; Beckmann, R. 3.2-Å-resolution structure of the 90S preribosome before A1 pre-rRNA cleavage. Nat. Struct. Mol. Biol. 2017, 24, 954–964. [Google Scholar] [CrossRef]
- RNA modification. In The Enzymes; Tamanoi, F.; Chanfreau, G. (Eds.) Academic Press: Cambridge, MA, USA, 2017. [Google Scholar]
- Wang, B.; Ye, K. Nop9 binds the central pseudoknot region of 18S rRNA. Nucleic Acids Res. 2017, 45, 3559–3567. [Google Scholar] [CrossRef] [Green Version]
- Dong, J.; Nanda, J.S.; Rahman, H.; Pruitt, M.R.; Shin, B.S.; Wong, C.M.; Lorsch, J.R.; Hinnebusch, A.G. Genetic identification of yeast 18S rRNA residues required for efficient recruitment of initiator tRNAMet and AUG selection. Genes Dev. 2008, 22, 2242–2255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, Z.; Yang, Y.; Dengyan, Z.; Chunyang, Z.; Donglei, L.; Kai, W.; Song, Z. RAP1B, a DVL2 binding protein, activates Wnt/beta-catenin signaling in esophageal squamous cell carcinoma. Gene 2017, 611, 15–20. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.-H.; Wu, Q.; Lu, J.; Chen, L.; Wu, X.; Wang, Y.; Ju, H.; Lu, Y.; Chen, Y.; Wang, F.; et al. Eukaryotic initiation factor 4A2 (EIF4A2) expression in colorectal cancer and prediction of prognosis. J. Clin. Oncol. 2018, 36, 664. [Google Scholar] [CrossRef]
- Modelska, A.; Turro, E.; Russell, R.; Beaton, J.; Sbarrato, T.; Spriggs, K.; Miller, J.; Gräf, S.; Provenzano, E.; Blows, F.; et al. The malignant phenotype in breast cancer is driven by eIf4A1-mediated changes in the translational landscape. Cell Death Dis. 2015, 6, e1603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanic, N.; Brkic, G.; Dimitrijevic, B.; Dedovic-Tanic, N.; Gefen, N.; Benharroch, D.; Gopas, J. Identification of differentially expressed mRNA transcripts in drug-resistant versus parental human melanoma cell lines. Anticancer. Res. 2006, 26, 2137–2142. [Google Scholar] [PubMed]
- Wang, A.; Zhao, C.; Liu, X.; Su, W.; Duan, G.; Xie, Z.; Chu, S.; Gao, Y. Knockdown of TBRG4 affects tumorigenesis in human H1299 lung cancer cells by regulating DDIT3, CAV1 and RRM2. Oncol. Lett. 2018, 15, 121–128. [Google Scholar] [CrossRef]
- Mishra, S.; Saleembhasha, A. Long non-coding RNAs as pan-cancer master gene regulators of associated protein-coding genes: A systems biology approach. Peer J. 2019, 7, e6388. [Google Scholar]
- Xu, W.; Zhou, G.; Wang, H.; Liu, Y.; Chen, B.; Chen, W.; Lin, C.; Wu, S.; Gong, A.; Xu, M. Circulating lncRNA SNHG11 as a novel biomarker for early diagnosis and prognosis of colorectal cancer. Int. J. Cancer 2019. [Google Scholar] [CrossRef]
- Ye, G.; Guo, L.; Xing, Y.; Sun, W.; Yuan, M. Identification of prognostic biomarkers of prostate cancer with long non-coding RNA-mediated competitive endogenous RNA network. Exp. Ther. Med. 2019, 17, 3035–3040. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Q.; Fan, C. A novel risk score system for assessment of ovarian cancer based on co-expression network analysis and expression level of five lncRNAs. BMC Med. Genet. 2019, 20, 103. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tumor Acronym | Tumor Type | Sample Number |
|---|---|---|
| TCGA-HNSC | Head and Neck squamous cell carcinoma | 496 |
| TCGA-KIRC | Kidney renal clear cell carcinoma | 354 |
| TCGA-BRCA | Breast invasive carcinoma | 1070 |
| TCGA-COAD | Colorectal adenocarcinoma | 592 |
| TCGA-LUSC | Lung squamous cell carcinoma | 487 |
| TCGA-SKCM | Skin Cutaneous Melanoma | 367 |
| TCGA-LUAD | Lung adenocarcinoma | 511 |
| TCGA-UCEC | Uterine Corpus Endometrial Carcinoma | 509 |
| TCGA-OV | Ovarian serous cystadenocarcinoma | 398 |
| TCGA-GBM | Glioblastoma multiforme | 575 |
| snoRNA Host Gene | Known Target | Biological Relevance | Clinical Relevance | References |
|---|---|---|---|---|
| SNORA63 EIF4A2 | 28S rRNA U4390 (Helix 89) -- | Control of translation initiation, elongation and termination Translation initiation | None reported Inversely correlates with prognosis in COAD, NSCLC, BRCA | [49,50,51] |
| SNORA63E LINC00888 | Unknown -- | -- | None reported Increased in SKCM | [52] |
| SNORD66 EIF4G1 | 18S rRNA C1272 (Helix 32) -- | Translation initiation Translation initiation | Biomarker of NSCLC in sputum, plasma and tissue; correlates to overall survival in NSCLC Inversely correlates with prognosis in BRCA, OV, NSCLC, PDA | [53,54,55,56,57,58] |
| SNORA15 CCT6A | 18S rRNA U1367 (Helix 37) -- | Activation of TGFbeta signaling | None reported Correlates with negative prognosis in HCC | [59,60] |
| SNORA14B TOMM20 | 18S rRNA U966 (Helix 23) -- | E site/translation elongation Increases mitochondrial ATP synthesis | None reported Overexpressed in various cancer types; potential therapeutic target in COAD | [61,62] |
| SNORD72 RPL37 | 5.8S rRNA U55 (Helix 6) -- | Formation of the ribosome small subunit Activator of ribosomal stress pathway | None reported Overexpressed in PC | [33,63,64] |
| SNORA56 DKC1 | 28S rRNA U1664 (Helix 37) -- | Putative mRNA/tRNA binding rRNA pseudouridylation; regulates translational fidelity and Cap-independent translation; Telomere binding | None reported Negatively correlates with survival in BRCA, and NSCLC | [37,65,66,67,68] |
| SNORA36A DKC1 | 18S rRNA U105 (Helix 7) and U1244 (Helix 31) -- | Binding of factors to form the 90S preribosome (H7); Translation elongation (H31) See above | None reported See above | [69,70] |
| SNORA70G RAP1B | 18S rRNA U1692 (Helix 28) -- | Small ribosomal subunit maturation; translation initiation GTPase regulating cell adhesion, migration, polarity, differentiation, growth and angiogenesis | None reported Negatively correlates with prognosis in GC; up-regulated in ESCC | [71,72,73] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Penzo, M.; Clima, R.; Trerè, D.; Montanaro, L. Separated Siamese Twins: Intronic Small Nucleolar RNAs and Matched Host Genes May be Altered in Conjunction or Separately in Multiple Cancer Types. Cells 2020, 9, 387. https://doi.org/10.3390/cells9020387
Penzo M, Clima R, Trerè D, Montanaro L. Separated Siamese Twins: Intronic Small Nucleolar RNAs and Matched Host Genes May be Altered in Conjunction or Separately in Multiple Cancer Types. Cells. 2020; 9(2):387. https://doi.org/10.3390/cells9020387
Chicago/Turabian StylePenzo, Marianna, Rosanna Clima, Davide Trerè, and Lorenzo Montanaro. 2020. "Separated Siamese Twins: Intronic Small Nucleolar RNAs and Matched Host Genes May be Altered in Conjunction or Separately in Multiple Cancer Types" Cells 9, no. 2: 387. https://doi.org/10.3390/cells9020387