Constitutive Androstane Receptor: A Peripheral and a Neurovascular Stress or Environmental Sensor

Abstract
:1. Introduction: CAR Governs Detoxification Mechanisms
2. CAR Has a Particular Mechanism of Action: Direct and Indirect Activation
3. Functional Roles of CAR in Peripheral Organs
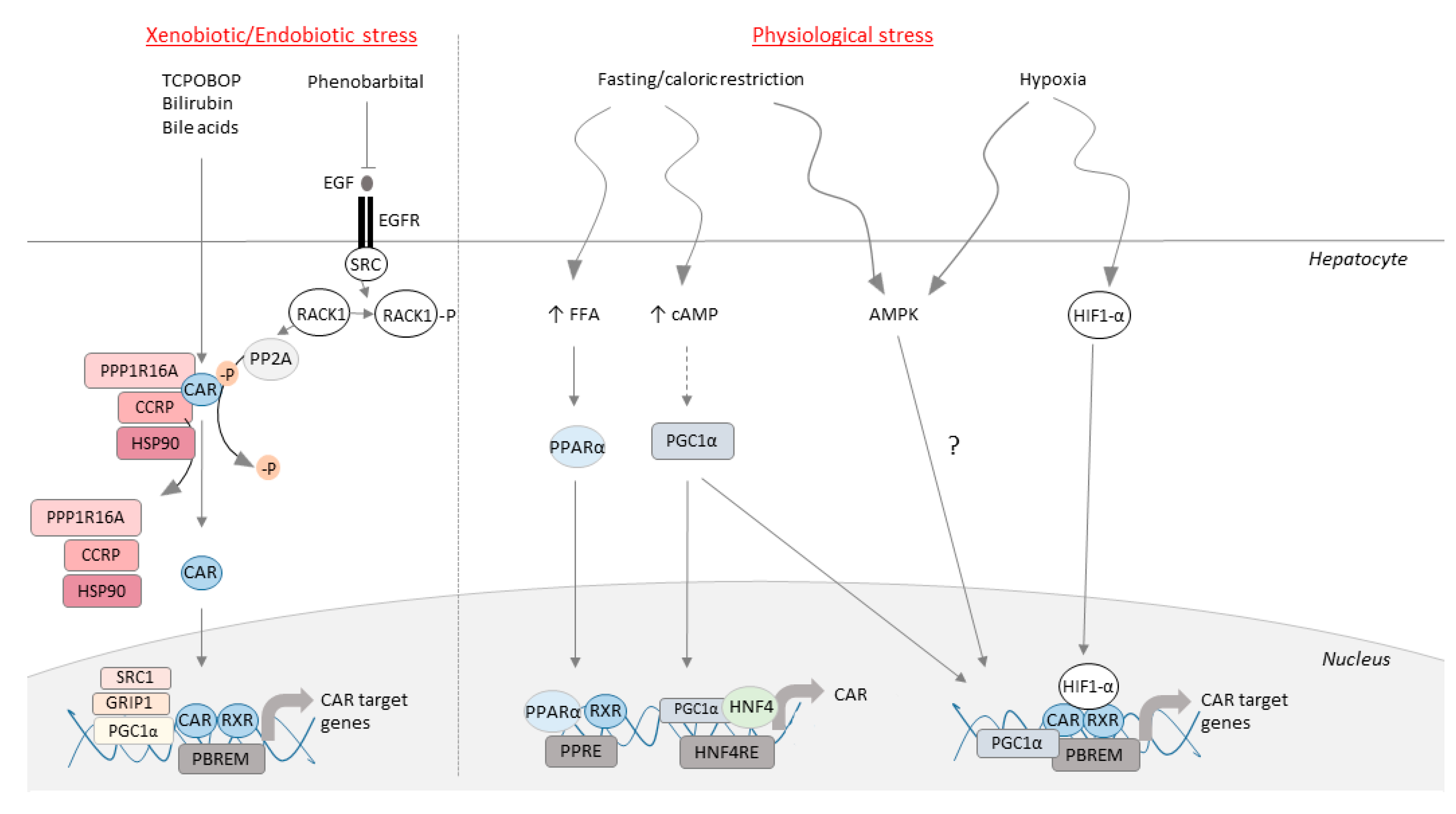
3.1. CAR as an Endobiotic Stress Sensor
3.2. CAR as a Regulator of Steroid and Thyroid Hormones
3.3. CAR as a Sensor of Fasting and Caloric Restriction
3.4. CAR as a Glucose Sensor
3.5. CAR as a Lipid Sensor
3.6. CAR as a Hypoxia Sensor
3.7. CAR Intestinal Response to Inflammatory Stress
3.8. CAR Protects from Acute Kidney Injury
4. Functional Roles of CAR in the Brain: Focus on the Neurovascular Unit
4.1. Brain Expression, Regulation and Function of CAR
4.2. CAR and Brain Disease Conditions: Initial Clinical and Experimental Clues
5. Conclusions: Can We Integrate CAR within a Peripheral–Brain Axis?
Author Contributions
Funding
Conflicts of Interest
References
- Wei, P.; Zhang, J.; Egan-Hafley, M.; Liang, S.; Moore, D.D. The nuclear receptor CAR mediates specific xenobiotic induction of drug metabolism. Nature 2000, 407, 920–923. [Google Scholar] [CrossRef] [PubMed]
- Forman, B.M.; Tzameli, I.; Choi, H.S.; Chen, J.; Simha, D.; Seol, W.; Evans, R.M.; Moore, D.D. Androstane metabolites bind to and deactivate the nuclear receptor CAR-beta. Nature 1998, 395, 612–615. [Google Scholar] [CrossRef] [PubMed]
- Kawamoto, T.; Sueyoshi, T.; Zelko, I.; Moore, R.; Washburn, K.; Negishi, M. Phenobarbital-responsive nuclear translocation of the receptor CAR in induction of the CYP2B gene. Mol. Cell. Biol. 1999, 19, 6318–6322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernandez, J.; Mota, L.; Baldwin, W. Activation of CAR and PXR by Dietary, Environmental and Occupational Chemicals Alters Drug Metabolism, Intermediary Metabolism, and Cell Proliferation. Curr. Pharm. Person. Med. 2009, 7, 81–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, D.; Mackowiak, B.; Brayman, T.G.; Mitchell, M.; Zhang, L.; Huang, S.; Wang, H.; Spring, S. Genome-wide Analysis of Human Constitutive Androstane Receptor (CAR) Transcriptome in Wild-type and CAR-knockout HepaRG cells. Biochem. Pharmacol. 2015, 98, 190–202. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Wang, H. Activation of xenobiotic receptors: Driving into the nucleus. Expert Opin. Drug Metab. Toxicol. 2010, 6, 409–426. [Google Scholar] [CrossRef] [Green Version]
- Maglich, J.M.; Stoltz, C.M.; Goodwin, B.; Hawkins-Brown, D.; Moore, J.T.; Kliewer, S.A. Nuclear pregnane X receptor and constitutive androstane receptor regulate overlapping but distinct sets of genes involved in xenobiotic detoxification. Mol. Pharmacol. 2002, 62, 638–646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saini, S.P.S.; Sonoda, J.; Xu, L.; Toma, D.; Uppal, H.; Mu, Y.; Ren, S.; Moore, D.D.; Evans, R.M.; Xie, W. A Novel Constitutive Androstane Receptor-Mediated and CYP3A-Independent Pathway of Bile Acid Detoxification. Mol. Pharmacol. 2004, 65, 292–300. [Google Scholar] [CrossRef] [Green Version]
- Sugatani, J.; Yamakawa, K.; Yoshinari, K.; Miwa, M.; Machida, T.; Takagi, H.; Mori, M.; Kakizaki, S.; Sueyoshi, T.; Negishi, M. Identification of a defect in the UGT1A1 gene promoter and its association with hyperbilirubinemia. Biochem. Biophys. Res. Commun. 2002, 292, 492–497. [Google Scholar] [CrossRef]
- Ueda, A.; Hamadeh, H.K.; Webb, H.K.; Yamamoto, Y.; Sueyoshi, T.; Afshari, C.A.; Lehmann, J.M.; Negishi, M. Diverse roles of the nuclear orphan receptor CAR in regulating hepatic genes in response to phenobarbital. Mol. Pharmacol. 2002, 61, 1–6. [Google Scholar] [CrossRef]
- Locker, J.; Tian, J.; Carver, R.; Concas, D.; Cossu, C.; Ledda-Columbano, G.M.; Columbano, A. A common set of immediate-early response genes in liver regeneration and hyperplasia. Hepatology 2003, 38, 314–325. [Google Scholar] [CrossRef] [PubMed]
- Dong, B.; Saha, P.K.; Huang, W.; Chen, W.; Abu-Elheiga, L.A.; Wakil, S.J.; Stevens, R.D.; Ilkayeva, O.; Newgard, C.B.; Chan, L.; et al. Activation of nuclear receptor CAR ameliorates diabetes and fatty liver disease. Proc. Natl. Acad. Sci. USA 2009, 106, 18831–18836. [Google Scholar] [CrossRef] [Green Version]
- Gao, J.; He, J.; Zhai, Y.; Wada, T.; Xie, W. The constitutive androstane receptor is an anti-obesity nuclear receptor that improves insulin sensitivity. J. Biol. Chem. 2009, 284, 25984–25992. [Google Scholar] [CrossRef] [Green Version]
- Ding, X.; Lichti, K.; Kim, I.; Gonzalez, F.J.; Staudinger, J.L. Regulation of constitutive androstane receptor and its target genes by fasting, cAMP, hepatocyte nuclear factor alpha, and the coactivator peroxisome proliferator-activated receptor gamma coactivator-1alpha. J. Biol. Chem. 2006, 281, 26540–26551. [Google Scholar] [CrossRef] [Green Version]
- Maglich, J.M.; Watson, J.; McMillen, P.J.; Goodwin, B.; Willson, T.M.; Moore, J.T. The nuclear receptor CAR is a regulator of thyroid hormone metabolism during caloric restriction. J. Biol. Chem. 2004, 279, 19832–19838. [Google Scholar] [CrossRef] [Green Version]
- Shizu, R.; Shindo, S.; Yoshida, T.; Numazawa, S. Cross-talk between constitutive androstane receptor and hypoxia-inducible factor in the regulation of gene expression. Toxicol. Lett. 2013, 219, 143–150. [Google Scholar] [CrossRef]
- Shindo, S.; Numazawa, S.; Yoshida, T. A physiological role of AMP-activated protein kinase in phenobarbital-mediated constitutive androstane receptor activation and CYP2B induction. Biochem. J. 2007, 401, 735–741. [Google Scholar] [CrossRef]
- Osabe, M.; Sugatani, J.; Takemura, A.; Kurosawa, M.; Yamazaki, Y.; Ikari, A.; Miwa, M. Up-regulation of CAR expression through Elk-1 in HepG2 and SW480 cells by serum starvation stress. FEBS Lett. 2009, 583, 885–889. [Google Scholar] [CrossRef] [Green Version]
- Xu, R.X.; Lambert, M.H.; Wisely, B.B.; Warren, E.N.; Weinert, E.E.; Waitt, G.M.; Williams, J.D.; Collins, J.L.; Moore, L.B.; Willson, T.M.; et al. A structural basis for constitutive activity in the human CAR/RXRα heterodimer. Mol. Cell 2004, 16, 919–928. [Google Scholar] [CrossRef]
- Shan, L.; Vincent, J.; Brunzelle, J.S.; Dussault, I.; Lin, M.; Ianculescu, I.; Sherman, M.A.; Forman, B.M.; Fernandez, E.J. Structure of the murine constitutive androstane receptor complexed to androstenol: A molecular basis for inverse agonism. Mol. Cell 2004, 16, 907–917. [Google Scholar] [CrossRef] [Green Version]
- Yoshinari, K.; Kobayashi, K.; Moore, R.; Kawamoto, T.; Negishi, M. Identification of the nuclear receptor CAR:HSP90 complex in mouse liver and recruitment of protein phosphatase 2A in response to phenobarbital. FEBS Lett. 2003, 548, 17–20. [Google Scholar] [CrossRef] [Green Version]
- Sueyoshi, T.; Moore, R.; Sugatani, J.; Matsumura, Y.; Negishi, M. PPP1R16A, the membrane subunit of protein phosphatase 1beta, signals nuclear translocation of the nuclear receptor constitutive active/androstane receptor. Mol. Pharmacol. 2008, 73, 1113–1121. [Google Scholar] [CrossRef] [Green Version]
- Tzameli, I.; Pissios, P.; Schuetz, E.G.; Moore, D.D. The xenobiotic compound 1,4-bis[2-(3,5-dichloropyridyloxy)]benzene is an agonist ligand for the nuclear receptor CAR. Mol. Cell. Biol. 2000, 20, 2951–2958. [Google Scholar] [CrossRef] [Green Version]
- Mutoh, S.; Osabe, M.; Inoue, K.; Moore, R.; Pedersen, L.; Perera, L.; Rebolloso, Y.; Sueyoshi, T.; Negishi, M. Dephosphorylation of threonine 38 Is required for nuclear translocation and activation of human xenobiotic receptor CAR (NR1I3). J. Biol. Chem. 2009, 284, 34785–34792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mäkinen, J.; Frank, C.; Jyrkkärinne, J.; Gynther, J.; Carlberg, C.; Honkakoski, P. Modulation of mouse and human phenobarbital-responsive enhancer module by nuclear receptors. Mol. Pharmacol. 2002, 62, 366–378. [Google Scholar] [CrossRef] [Green Version]
- Min, G.; Kemper, J.K.; Kemper, B. Glucocorticoid receptor-interacting protein 1 mediates ligand-independent nuclear translocation and activation of constitutive androstane receptor in vivo. J. Biol. Chem. 2002, 277, 26356–26363. [Google Scholar] [CrossRef] [Green Version]
- Shiraki, T.; Sakai, N.; Kanaya, E.; Jingami, H. Activation of orphan nuclear constitutive androstane receptor requires subnuclear targeting by peroxisome proliferator-activated receptor γ coactivator-1α: A possible link between xenobiotic response and nutritional state. J. Biol. Chem. 2003, 278, 11344–11350. [Google Scholar] [CrossRef] [Green Version]
- Wright, E.; Vincent, J.; Fernandez, E.J. Thermodynamic characterization of the interaction between CAR-RXR and SRC-1 peptide by isothermal titration calorimetry. Biochemistry 2007, 46, 862–870. [Google Scholar] [CrossRef] [Green Version]
- Baes, M.; Gulick, T.; Choi, H.S.; Martinoli, M.G.; Simha, D.; Moore, D.D. A new orphan member of the nuclear hormone receptor superfamily that interacts with a subset of retinoic acid response elements. Mol. Cell. Biol. 1994, 14, 1544–1552. [Google Scholar] [CrossRef] [Green Version]
- Zmrzljak, U.P.; Rozman, D. Circadian regulation of the hepatic endobiotic and xenobitoic detoxification pathways: The time matters. Chem. Res. Toxicol. 2012, 25, 811–824. [Google Scholar] [CrossRef]
- Moore, L.B.; Parks, D.J.; Jones, S.A.; Bledsoe, R.K.; Consler, T.G.; Stimmel, J.B.; Goodwin, B.; Liddle, C.; Blanchard, S.G.; Willson, T.M.; et al. Orphan nuclear receptors constitutive androstane receptor and pregnane X receptor share xenobiotic and steroid ligands. J. Biol. Chem. 2000, 275, 15122–15127. [Google Scholar] [CrossRef] [Green Version]
- Hernandez, J.P.; Mota, L.C.; Huang, W.; Moore, D.D.; Baldwin, W.S. Sexually dimorphic regulation and induction of P450s by the constitutive androstane receptor (CAR). Toxicology 2009, 256, 53–64. [Google Scholar] [CrossRef] [Green Version]
- Lynch, C.; Mackowiak, B.; Huang, R.; Li, L.; Heyward, S.; Srilatha, S.; Wang, H.; Xia, M. Identification of Modulators that Activate the Constitutive Androstane Receptor from the Tox21 10K Compound Library. Toxicol. Sci. 2019, 1, 282–292. [Google Scholar] [CrossRef] [PubMed]
- Fujino, C.; Watanabe, Y.; Sanoh, S.; Nakajima, H.; Uramaru, N.; Kojima, H.; Yoshinari, K.; Ohta, S.; Kitamura, S. Activation of PXR, CAR and PPARα by pyrethroid pesticides and the effect of metabolism by rat liver microsomes. Heliyon 2019, 5, e02466. [Google Scholar] [CrossRef] [Green Version]
- Abe, T.; Takahashi, M.; Kano, M.; Amaike, Y.; Ishii, C.; Maeda, K.; Kudoh, Y.; Morishita, T.; Hosaka, T.; Sasaki, T.; et al. Activation of nuclear receptor CAR by an environmental pollutant perfluorooctanoic acid. Arch. Toxicol. 2017, 91, 2365–2374. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; Klaassen, C.D. Perfluorocarboxylic acids induce cytochrome P450 enzymes in mouse liver through activation of PPAR-α and CAR transcription factors. Toxicol. Sci. 2008, 106, 29–36. [Google Scholar] [CrossRef] [Green Version]
- Oshida, K.; Vasani, N.; Jones, C.; Moore, T.; Hester, S.; Nesnow, S.; Auerbach, S.; Geter, D.R.; Aleksunes, L.M.; Thomas, R.S.; et al. Identification of chemical modulators of the constitutive activated receptor (CAR) in a gene expression compendium. Nucl. Recept. Signal. 2015, 13, e002. [Google Scholar] [CrossRef] [Green Version]
- Elcombe, C.R.; Elcombe, B.M.; Foster, J.R.; Chang, S.C.; Ehresman, D.J.; Butenhoff, J.L. Hepatocellular hypertrophy and cell proliferation in Sprague-Dawley rats from dietary exposure to potassium perfluorooctanesulfonate results from increased expression of xenosensor nuclear receptors PPARα and CAR/PXR. Toxicology 2012, 293, 16–29. [Google Scholar] [CrossRef]
- Baldwin, W.S.; Roling, J.A. A Concentration Addition Model for the Activation of the Constitutive Androstane Receptor by Xenobiotic Mixtures. Toxicol. Sci. 2009, 107, 93–105. [Google Scholar] [CrossRef] [Green Version]
- Pakharukova, M.Y.; Smetanina, M.A.; Kaledin, V.I.; Kobzev, V.F.; Romanova, I.V.; Merkulova, T.I. Activation of constitutive androstane receptor under the effect of hepatocarcinogenic aminoazo dyes in mouse and rat liver. Bull. Exp. Biol. Med. 2007, 144, 338–341. [Google Scholar] [CrossRef]
- Peffer, R.C.; Moggs, J.G.; Pastoor, T.; Currie, R.A.; Wright, J.; Milburn, G.; Waechter, F.; Rusyn, I. Mouse liver effects of cyproconazole, a triazole fungicide: Role of the constitutive androstane receptor. Toxicol. Sci. 2007, 99, 315–325. [Google Scholar] [CrossRef]
- Wyde, M.E.; Kirwan, S.E.; Zhang, F.; Laughter, A.; Hoffman, H.B.; Bartolucci-Page, E.; Gaido, K.W.; Yan, B.; You, L. Di-n-butyl phthalate activates constitutive androstane receptor and pregnane X receptor and enhances the expression of steroid-metabolizing enzymes in the liver of rat fetuses. Toxicol. Sci. 2005, 86, 281–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wyde, M.E.; Bartolucci, E.; Ueda, A.; Zhang, H.; Yan, B.; Negishi, M.; You, L. The environmental pollutant 1,1-Dichloro-2,2-bis (p-chlorophenyl)ethylene induces rat hepatic cytochrome P450 2B and 3A expression through the constitutive androstane receptor and pregnane X receptor. Mol. Pharmacol. 2003, 64, 474–481. [Google Scholar] [CrossRef]
- Laurenzana, E.M.; Coslo, D.M.; Vigilar, M.V.; Roman, A.M.; Omiecinski, C.J. Activation of the Constitutive Androstane Receptor by Monophthalates. Chem. Res. Toxicol. 2016, 29, 1651–1661. [Google Scholar] [CrossRef]
- Sueyoshi, T.; Kawamoto, T.; Zelko, I.; Honkakoski, P.; Negishi, M. The repressed nuclear receptor CAR responds to phenobarbital in activating the human CYP2B6 gene. J. Biol. Chem. 1999, 274, 6043–6046. [Google Scholar] [CrossRef] [Green Version]
- DeKeyser, J.G.; Stagliano, M.C.; Auerbach, S.S.; Prabhu, K.S.; Jones, A.D.; Omiecinski, C.J. Di(2-ethylhexyl) phthalate is a highly potent agonist for the human constitutive androstane receptor splice variant CAR2. Mol. Pharmacol. 2009, 75, 1005–1012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, P.; Zhang, J.; Dowhan, D.H.; Han, Y.; Moore, D.D. Specific and overlapping functions of the nuclear hormone receptors CAR and PXR in xenobiotic response. Pharm. J. 2002, 2, 117–126. [Google Scholar] [CrossRef]
- Savary, C.C.; Jossé, R.; Bruyère, A.; Guillet, F.; Robin, M.A.; Guillouzo, A. Interactions of endosulfan and methoxychlor involving CYP3A4 and CYP2B6 in human HepaRG cells. Drug Metab. Dispos. 2014, 42, 1234–1240. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.J.; Shi, Z.; Lyv, J.X.; He, X.; Englert, N.A.; Zhang, S.Y. Pyrene is a novel constitutive androstane receptor (CAR) activator and causes hepatotoxicity by CAR. Toxicol. Sci. 2015, 147, 436–445. [Google Scholar] [CrossRef] [Green Version]
- Blizard, D.; Sueyoshi, T.; Negishi, M.; Dehal, S.S.; Kupfer, D. Mechanism of Induction of Cytochrome P450 Enzymes by the Proestrogenic Endocrine Disruptor Pesticide-Methoxychlor: Interactions of Methoxychlor Metabolites with the Constitutive Androstane Receptor System. Drug Metab. Dispos. 2001, 29, 781–785. [Google Scholar]
- Prokopec, S.D.; Watson, J.D.; Lee, J.; Pohjanvirta, R.; Boutros, P.C. Sex-related differences in murine hepatic transcriptional and proteomic responses to TCDD. Toxicol. Appl. Pharmacol. 2015, 284, 188–196. [Google Scholar] [CrossRef]
- Mackowiak, B.; Li, L.; Lynch, C.; Ziman, A.; Heyward, S.; Wang, H.; States, U.; States, U.; States, U.; Technologies, I.V.; et al. High-content analysis of constitutive androstane receptor (CAR) translocation identifies mosapride citrate as a CAR agonist that represses gluconeogenesis. Biochem. Pharmacol. 2019, 168, 224–236. [Google Scholar] [CrossRef]
- Liang, D.; Li, L.; Lynch, C.; Diethelm-Varela, B.; Xia, M.; Xue, F.; Wang, H. DL5050, a Selective Agonist for the Human Constitutive Androstane Receptor. ACS Med. Chem. Lett. 2019, 10, 1039–1044. [Google Scholar] [CrossRef] [PubMed]
- Cerveny, L.; Svecova, L.; Anzenbacherova, E.; Vrzal, R.; Staud, F.; Dvorak, Z.; Ulrichova, J.; Anzenbacher, P.; Pavek, P. Valproic acid induces CYP3A4 and MDR1 gene expression by activation of constitutive androstane receptor and pregnane X receptor pathways. Drug Metab. Dispos. 2007, 35, 1032–1041. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Huang, W.; Chua, S.S.; Wei, P.; Moore, D.D. Modulation of acetaminophen-induced hepatotoxicity by the xenobiotic receptor CAR. Science 2002, 298, 422–424. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Huang, W.; Qatanani, M.; Evans, R.M.; Moore, D.D. The constitutive androstane receptor and pregnane X receptor function coordinately to prevent bile acid-induced hepatotoxicity. J. Biol. Chem. 2004, 279, 49517–49522. [Google Scholar] [CrossRef] [Green Version]
- Goetz, A.K.; Bao, W.; Ren, H.; Schmid, J.E.; Tully, D.B.; Wood, C.; Rockett, J.C.; Narotsky, M.G.; Sun, G.; Lambert, G.R.; et al. Gene expression profiling in the liver of CD-1 mice to characterize the hepatotoxicity of triazole fungicides. Toxicol. Appl. Pharmacol. 2006, 215, 274–284. [Google Scholar] [CrossRef]
- Burk, O.; Arnold, K.A.; Nussler, A.K.; Schaeffeler, E.; Efimova, E.; Avery, B.A.; Avery, M.A.; Fromm, M.F.; Eichelbaum, M. Antimalarial artemisinin drugs induce cytochrome P450 and MDR1 expression by activation of xenosensors pregnane X receptor and constitutive androstane receptor. Mol. Pharmacol. 2005, 67, 1954–1965. [Google Scholar] [CrossRef] [Green Version]
- Swales, K.; Negishi, M. CAR, Driving into the Future. Mol. Endocrinol. 2004, 18, 1589–1598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Chen, T.; Stanton, J.D.; Sueyoshi, T.; Negishi, M.; Wang, H. The peripheral benzodiazepine receptor ligand 1-(2-chlorophenyl-methylpropyl)-3-isoquinoline-carboxamide is a novel antagonist of human constitutive androstane receptor. Mol. Pharmacol. 2008, 74, 443–453. [Google Scholar] [CrossRef] [Green Version]
- Malaplate-Armand, C.; Ferrari, L.; Masson, C.; Visvikis-Siest, S.; Lambert, H.; Batt, A.M. Down-regulation of astroglial CYP2C, glucocorticoid receptor and constitutive androstane receptor genes in response to cocaine in human U373 MG astrocytoma cells. Toxicol. Lett. 2005, 159, 203–211. [Google Scholar] [CrossRef]
- Pascussi, J.M.; Gerbal-Chaloin, S.; Fabre, J.M.; Maurel, P.; Vilarem, M.J. Dexamethasone enhances constitutive androstane receptor expression in human hepatocytes: Consequences on cytochrome P450 gene regulation. Mol. Pharmacol. 2000, 58, 1441–1450. [Google Scholar] [CrossRef] [Green Version]
- Qatanani, M.; Wei, P.; Moore, D.D. Alterations in the distribution and orexigenic effects of dexamethasone in CAR-null mice. Pharmacol. Biochem. Behav. 2004, 78, 285–291. [Google Scholar] [CrossRef]
- Duret, C.; Daujat-Chavanieu, M.; Pascussi, J.M.; Pichard-Garcia, L.; Balaguer, P.; Fabre, J.M.; Vilarem, M.J.; Maurel, P.; Gerbal-Chaloin, S. Ketoconazole and miconazole are antagonists of the human glucocorticoid receptor: Consequences on the expression and function of the constitutive androstane receptor and the pregnane X receptor. Mol. Pharmacol. 2006, 70, 329–339. [Google Scholar] [CrossRef] [Green Version]
- Huang, W.; Zhang, J.; Wei, P.; Schrader, W.T.; Moore, D.D. Meclizine is an agonist ligand for mouse constitutive androstane receptor (CAR) and an inverse agonist for human CAR. Mol. Endocrinol. 2004, 18, 2402–2408. [Google Scholar] [CrossRef] [Green Version]
- Saussele, T.; Burk, O.; Blievernicht, J.K.; Klein, K.; Nussler, A.; Nussler, N.; Hengstler, J.G.; Eichelbaum, M.; Schwab, M.; Zanger, U.M. Selective induction of human hepatic cytochromes P450 2B6 and 3A4 by metamizole. Clin. Pharmacol. Ther. 2007, 82, 265–274. [Google Scholar] [CrossRef]
- Chen, X.; Maiti, S.; Zhang, J.; Chen, G. Nuclear receptor interactions in methotrexate induction of human dehydroepiandrosterone sulfotransferase (hSULT2A1). J. Biochem. Mol. Toxicol. 2006, 20, 309–317. [Google Scholar] [CrossRef]
- Shibayama, Y.; Ushinohama, K.; Ikeda, R.; Yoshikawa, Y.; Motoya, T.; Takeda, Y.; Yamada, K. Effect of methotrexate treatment on expression levels of multidrug resistance protein 2, breast cancer resistance protein and organic anion transporters Oat1, Oat2 and Oat3 in rats. Cancer Sci. 2006, 97, 1260–1266. [Google Scholar] [CrossRef]
- Murray, M.; Fiala-Beer, E.; Sutton, D. Upregulation of cytochromes P450 2B in rat liver by orphenadrine. Br. J. Pharmacol. 2003, 139, 787–796. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Bao, X.; Zhang, Q.Y.; Negishi, M.; Ding, X. Role of Cyp2b in phenobarbital-induced hepatocyte proliferation in mice. Drug Metab. Dispos. 2017, 45, 977–981. [Google Scholar] [CrossRef] [Green Version]
- Currie, R.A.; Peffer, R.C.; Goetz, A.K.; Omiecinski, C.J.; Goodman, J.I. Phenobarbital and propiconazole toxicogenomic profiles in mice show major similarities consistent with the key role that constitutive androstane receptor (CAR) activation plays in their mode of action. Toxicology 2014, 321, 80–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jackson, J.P.; Ferguson, S.S.; Moore, R.; Negishi, M.; Goldstein, J.A. The constitutive active/androstane receptor regulates phenytoin induction of Cyp2c29. Mol. Pharmacol. 2004, 65, 1397–1404. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Faucette, S.; Moore, R.; Sueyoshi, T.; Negishi, M.; LeCluyse, E. Human constitutive androstane receptor mediates induction of CYP2B6 gene expression by phenytoin. J. Biol. Chem. 2004, 279, 29295–29301. [Google Scholar] [CrossRef] [Green Version]
- Jackson, J.P.; Ferguson, S.S.; Negishi, M.; Goldstein, J.A. Phenytoin Induction of the Cyp2c37 Gene is Mediated by the Constitutive Androstane Receptor. Drug Metab. Dispos. 2006, 34, 2003–2010. [Google Scholar] [CrossRef] [Green Version]
- Howe, K.; Sanat, F.; Thumser, A.E.; Coleman, T.; Plant, N. The statin class of HMG-CoA reductase inhibitors demonstrate differential activation of the nuclear receptors PXR, CAR and FXR, as well as their downstream target genes. Xenobiotica 2011, 41, 519–529. [Google Scholar] [CrossRef] [Green Version]
- Režen, T.; Hafner, M.; Kortagere, S.; Ekins, S.; Hodnik, V.; Rozman, D. Rosuvastatin and atorvastatin are ligands of the human constitutive androstane receptor/retinoid X receptor α complex. Drug Metab. Dispos. 2017, 45, 974–976. [Google Scholar] [CrossRef] [PubMed]
- Sugatani, J.; Kojima, H.; Ueda, A.; Kakizaki, S.; Yoshinari, K.; Gong, Q.H.; Owens, I.S.; Negishi, M.; Sueyoshi, T. The phenobarbital response enhancer module in the human bilirubin UDP-glucuronosyltransferase UGT1A1 gene and regulation by the nuclear receptor CAR. Hepatology 2001, 33, 1232–1238. [Google Scholar] [CrossRef]
- Guo, G.L.; Choudhuri, S.; Klaassen, C.D. Induction profile of rat organic anion transporting polypeptide 2 (oatp2) by prototypical drug-metabolizing enzyme inducers that activate gene expression through ligand-activated transcription factor pathways. J. Pharmacol. Exp. Ther. 2002, 300, 206–212. [Google Scholar] [CrossRef] [Green Version]
- Ridlon, J.M.; Kang, D.J.; Hylemon, P.B. Bile salt biotransformations by human intestinal bacteria. J. Lipid Res. 2006, 47, 241–259. [Google Scholar] [CrossRef] [Green Version]
- Beilke, L.D.; Aleksunes, L.M.; Holland, R.D.; Besselsen, D.G.; Beger, R.D.; Klaassen, C.D.; Cherrington, N.J. Constitutive androstane receptor-mediated changes in bile acid composition contributes to hepatoprotection from lithocholic acid-Induced liver injury in mice. Drug Metab. Dispos. 2009, 37, 1035–1045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schenkman, J.B. Steroid metabolism by constitutive cytochromes P450. J. Steroid Biochem. Mol. Biol. 1992, 43, 1023–1030. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Moore, R.; Hess, H.A.; Guo, G.L.; Gonzalez, F.J.; Korach, K.S.; Maronpot, R.R.; Negishi, M. Estrogen receptor α mediates 17α-ethynylestradiol causing hepatotoxicity. J. Biol. Chem. 2006, 281, 16625–16631. [Google Scholar] [CrossRef] [Green Version]
- Sueyoshi, T.; Green, W.D.; Vinal, K.; Woodrum, T.S.; Moore, R.; Negishi, M. Garlic Extract Diallyl Sulfide (DAS) Activates Nuclear Receptor CAR to Induce the Sult1e1 Gene in Mouse Liver. PLoS ONE 2011, 6, e21229. [Google Scholar] [CrossRef] [Green Version]
- Curran, P.G.; DeGroot, L.J. The effect of hepatic enzyme-inducing drugs on thyroid hormones and the thyroid gland. Endocr. Rev. 1991, 12, 135–150. [Google Scholar] [CrossRef]
- Qatanani, M.; Zhang, J.; Moore, D.D. Role of the Constitutive Androstane Receptor in Xenobiotic-Induced Thyroid Hormone Metabolism. Endocrinology 2005, 146, 995–1002. [Google Scholar] [CrossRef]
- Visser, T.J.; Kaptein, E.; Glatt, H.; Bartsch, I.; Hagen, M.; Coughtrie, M.W.H. Characterization of thyroid hormone sulfotransferases. Chem. Biol. Interact. 1998, 109, 279–291. [Google Scholar] [CrossRef]
- Lukowicz, C.; Ellero-Simatos, S.; Régnier, M.; Oliviero, F.; Lasserre, F.; Polizzi, A.; Montagner, A.; Smati, S.; Boudou, F.; Lenfant, F.; et al. Dimorphic metabolic and endocrine disorders in mice lacking the constitutive androstane receptor. Sci. Rep. 2019, 9, 20169. [Google Scholar] [CrossRef]
- Wieneke, N.; Hirsch-Ernst, K.I.; Kuna, M.; Kersten, S.; Püschel, G.P. PPARα-dependent induction of the energy homeostasis-regulating nuclear receptor NR1i3 (CAR) in rat hepatocytes: Potential role in starvation adaptation. FEBS Lett. 2007, 581, 5617–5626. [Google Scholar] [CrossRef] [Green Version]
- Pascussi, J.M.; Busson-Le Coniat, M.; Maurel, P.; Vilarem, M.J. Transcriptional analysis of the orphan nuclear receptor constitutive androstane receptor (NR1I3) gene promoter: Identification of a distal glucocorticoid response element. Mol. Endocrinol. 2003, 17, 42–55. [Google Scholar] [CrossRef] [Green Version]
- Lahtela, J.T.; Arranto, A.J.; Sotaniemi, E.A. Enzyme inducers improve insulin sensitivity in non-insulin-dependent diabetic subjects. Diabetes 1985, 34, 911–916. [Google Scholar] [CrossRef]
- Sotaniemi, E.A.; Arranto, A.J.; Sutinen, S.; Stengård, J.H.; Sutinen, S. Treatment of noninsulin-dependent diabetes mellitus with enzyme inducers. Clin. Pharmacol. Ther. 1983, 33, 826–835. [Google Scholar] [CrossRef]
- Kodama, S.; Koike, C.; Negishi, M.; Yamamoto, Y. Nuclear receptors CAR and PXR cross talk with FOXO1 to regulate genes that encode drug-metabolizing and gluconeogenic enzymes. Mol. Cell. Biol. 2004, 24, 7931–7940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miao, J.; Fang, S.; Bae, Y.; Kemper, J.K. Functional inhibitory cross-talk between constitutive androstane receptor and hepatic nuclear factor-4 in hepatic lipid/glucose metabolism is mediated by competition for binding to the DR1 motif and to the common coactivators, GRIP-1 and PGC-1alpha. J. Biol. Chem. 2006, 281, 14537–14546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, J.; Yan, J.; Xu, M.; Ren, S.; Xie, W. CAR Suppresses Hepatic Gluconeogenesis by Facilitating the Ubiquitination and Degradation of PGC1α. Mol. Endocrinol. 2015, 29, 1558–1570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, X.; Cheng, Q.; Xu, L.; Yan, J.; Jiang, M.; He, J.; Xu, M.; Stefanovic-Racic, M.; Sipula, I.; O’Doherty, R.M.; et al. Cholesterol Sulfate and Cholesterol Sulfotransferase Inhibit Gluconeogenesis by Targeting Hepatocyte Nuclear Factor 4. Mol. Cell. Biol. 2014, 34, 485–497. [Google Scholar] [CrossRef] [Green Version]
- Lynch, C.; Pan, Y.; Li, L.; Heyward, S.; Moeller, T.; Swaan, P.W.; Wang, H. Activation of the constitutive androstane receptor inhibits gluconeogenesis without affecting lipogenesis or fatty acid synthesis in human hepatocytes. Toxicol. Appl. Pharmacol. 2014, 279, 33–42. [Google Scholar] [CrossRef] [Green Version]
- Breuker, C.; Moreau, A.; Lakhal, L.; Tamasi, V.; Parmentier, Y.; Meyer, U.; Maurel, P.; Lumbroso, S.; Vilarem, M.J.; Pascussi, J.M. Hepatic expression of thyroid hormone-responsive spot 14 protein is regulated by constitutive androstane receptor (NR1I3). Endocrinology 2010, 151, 1653–1661. [Google Scholar] [CrossRef] [Green Version]
- Marmugi, A.; Lukowicz, C.; Lasserre, F.; Montagner, A.; Polizzi, A.; Ducheix, S.; Goron, A.; Gamet-Payrastre, L.; Gerbal-Chaloin, S.; Pascussi, J.M.; et al. Activation of the Constitutive Androstane Receptor induces hepatic lipogenesis and regulates Pnpla3 gene expression in a LXR-independent way. Toxicol. Appl. Pharmacol. 2016, 303, 90–100. [Google Scholar] [CrossRef]
- La Vecchia, C.; Negri, E. A review of epidemiological data on epilepsy, phenobarbital, and risk of liver cancer. Eur. J. Cancer Prev. 2014, 23, 1–7. [Google Scholar]
- Verrotti, A.; Agostinelli, S.; Parisi, P.; Chiarelli, F.; Coppola, G. Nonalcoholic fatty liver disease in adolescents receiving valproic acid. Epilepsy Behav. 2011, 20, 382–385. [Google Scholar] [CrossRef]
- Kassam, A.; Winrow, C.J.; Fernandez-Rachubinski, F.; Capone, J.P.; Rachubinski, R.A. The peroxisome proliferator response element of the gene encoding the peroxisomal beta-oxidation enzyme enoyl-CoA hydratase/3-hydroxyacyl-CoA dehydrogenase is a target for constitutive androstane receptor beta/9-cis-retinoic acid receptor-mediated transactivation. J. Biol. Chem. 2000, 275, 4345–4350. [Google Scholar]
- Chen, W.; Chen, G.; Head, D.L.; Mangelsdorf, D.J.; Russell, D.W. Enzymatic Reduction of Oxysterols Impairs LXR Signaling in Cultured Cells and the Livers of Mice. Cell Metab. 2007, 5, 73–79. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.; Hwang, J.T.; Lee, H.J.; Jung, S.N.; Kang, I.; Chi, S.G.; Kim, S.S.; Ha, J. AMP-activated protein kinase activity is critical for hypoxia-inducible factor-1 transcriptional activity and its target gene expression under hypoxic conditions in DU145 cells. J. Biol. Chem. 2003, 278, 39653–39661. [Google Scholar] [CrossRef] [Green Version]
- Hudson, G.M.; Flannigan, K.L.; Erickson, S.L.; Vicentini, F.A.; Zamponi, A.; Hirota, C.L.; Alston, L.; Altier, C.; Ghosh, S.; Rioux, K.P.; et al. Constitutive androstane receptor regulates the intestinal mucosal response to injury. Br. J. Pharmacol. 2017, 174, 1857–1871. [Google Scholar] [CrossRef]
- Dempsey, J.L.; Wang, D.; Siginir, G.; Fei, Q.; Raftery, D.; Gu, H.; Yue Cui, J. Pharmacological Activation of PXR and CAR Downregulates Distinct Bile Acid-Metabolizing Intestinal Bacteria and Alters Bile Acid Homeostasis. Toxicol. Sci. 2019, 168, 40–60. [Google Scholar] [CrossRef]
- Little, M.; Dutta, M.; Li, H.; Matson, A.; Shi, X.; Gu, H.; Mani, S.; Cui, J.Y. Understanding the Physiological Functions of the Host Xenobiotic-Sensing Nuclear Receptors PXR and CAR on the Gut Microbiome Using Genetically Modified Mice. Master’s Thesis, University of Washington, Seattle, WA, USA, 2019. [Google Scholar]
- Choi, Y.J.; Zhou, D.; Barbosa, A.C.S.; Niu, Y.; Guan, X.; Xu, M.; Ren, S.; Nolin, T.D.; Liu, Y.; Xie, W. Activation of constitutive androstane receptor ameliorates renal ischemia-reperfusion-induced kidney and liver injury. Mol. Pharmacol. 2018, 93, 239–250. [Google Scholar] [CrossRef]
- Oh, E.; Kim, M.S. Brain regulation of energy metabolism. Endocrinol. Metab. 2016, 31, 519–524. [Google Scholar]
- Giannoni, P.; Badaut, J.; Dargazanli, C.; De Maudave, A.F.H.; Klement, W.; Costalat, V.; Marchi, N. The pericyte-glia interface at the blood-brain barrier. Clin. Sci. 2018, 132, 361–374. [Google Scholar] [CrossRef] [Green Version]
- Giannoni, P.; Claeysen, S.; Noe, F.; Marchi, N. Peripheral Routes to Neurodegeneration: Passing Through the Blood–Brain Barrier. Front. Aging Neurosci. 2020, 12, 3. [Google Scholar] [CrossRef] [Green Version]
- Sweeney, M.D.; Zhao, Z.; Montagne, A.; Nelson, A.R.; Zlokovic, B.V. Blood-brain barrier: From physiology to disease and back. Physiol. Rev. 2019, 99, 21–78. [Google Scholar] [CrossRef]
- Abbott, N.J.; Patabendige, A.A.K.; Dolman, D.E.M.; Yusof, S.R.; Begley, D.J. Structure and function of the blood-brain barrier. Neurobiol. Dis. 2010, 37, 13–25. [Google Scholar] [CrossRef]
- Nishimura, M.; Naito, S.; Yokoi, T. Tissue-specific mRNA expression profiles of human nuclear receptor subfamilies. Drug Metab. Pharmacokinet. 2004, 19, 135–149. [Google Scholar] [CrossRef]
- Lamba, J.K.; Lamba, V.; Yasuda, K.; Lin, Y.S.; Assem, M.; Thompson, E.; Strom, S.; Schuetz, E.G. Expression of constitutive androstane receptor splice variants in human tissues and their functional consequences. J. Pharmacol. Exp. Ther. 2004, 311, 811–821. [Google Scholar] [CrossRef] [Green Version]
- Dutheil, F.; Dauchy, S.; Diry, M.; Sazdovitch, V.; Cloarec, O.; Mellottée, L.; Bièche, I.; Ingelman-Sundberg, M.; Flinois, J.P.; De Waziers, I.; et al. Xenobiotic-metabolizing enzymes and transporters in the normal human brain: Regional and cellular mapping as a basis for putative roles in cerebral function. Drug Metab. Dispos. 2009, 37, 1528–1538. [Google Scholar] [CrossRef] [Green Version]
- Souidi, M.; Gueguen, Y.; Linard, C.; Dudoignon, N.; Grison, S.; Baudelin, C.; Marquette, C.; Gourmelon, P.; Aigueperse, J.; Dublineau, I. In vivo effects of chronic contamination with depleted uranium on CYP3A and associated nuclear receptors PXR and CAR in the rat. Toxicology 2005, 214, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Marini, S.; Nannelli, A.; Sodini, D.; Dragoni, S.; Valoti, M.; Longo, V.; Gervasi, P.G. Expression, microsomal and mitochondrial activities of cytochrome P450 enzymes in brain regions from control and phenobarbital-treated rabbits. Life Sci. 2007, 80, 910–917. [Google Scholar] [CrossRef]
- Kajta, M.; Wnuk, A.; Rzemieniec, J.; Lason, W.; Mackowiak, M.; Chwastek, E.; Staniszewska, M.; Nehring, I.; Wojtowicz, A.K. Triclocarban Disrupts the Epigenetic Status of Neuronal Cells and Induces AHR/CAR-Mediated Apoptosis. Mol. Neurobiol. 2019, 56, 3113–3131. [Google Scholar] [CrossRef] [PubMed]
- Litwa, E.; Rzemieniec, J.; Wnuk, A.; Lason, W.; Krzeptowski, W.; Kajta, M. RXRα, PXR and CAR xenobiotic receptors mediate the apoptotic and neurotoxic actions of nonylphenol in mouse hippocampal cells. J. Steroid Biochem. Mol. Biol. 2016, 156, 43–52. [Google Scholar] [CrossRef]
- The Human Protein Atlas. Available online: https://www.proteinatlas.org/ENSG00000143257-NR1I3/brain (accessed on 1 September 2020).
- Weksler, B.; Romero, I.A.; Couraud, P.O. The hCMEC/D3 cell line as a model of the human blood brain barrier. Fluids Barriers CNS 2013, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; LeCluyse, E.L. Role of Orphan Nuclear Receptors in the Regulation of Drug-Metabolising Enzymes. Clin. Pharmacokinet. 2003, 42, 1331–1357. [Google Scholar] [CrossRef]
- Miller, D.S. Regulation of ABC Transporters Blood-Brain Barrier. The Good, the Bad, and the Ugly. In Advances in Cancer Research; Academic Press Inc.: Cambridge, MA, USA, 2015; Volume 125, pp. 43–70. [Google Scholar]
- Chan, G.N.Y.; Hoque, M.T.; Cummins, C.L.; Bendayan, R. Regulation of P-glycoprotein by orphan nuclear receptors in human brain microvessel endothelial cells. J. Neurochem. 2011, 118, 163–175. [Google Scholar] [CrossRef]
- Wang, X.; Sykes, D.B.; Miller, D.S. Constitutive androstane receptor-mediated up-regulation of ATP-driven xenobiotic efflux transporters at the blood-brain barrier. Mol. Pharmacol. 2010, 78, 376–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slosky, L.M.; Thompson, B.J.; Sanchez-Covarrubias, L.; Zhang, Y.; Laracuente, M.L.; Vanderah, T.W.; Ronaldson, P.T.; Davis, T.P. Acetaminophen modulates P-glycoprotein functional expression at the blood-brain barrier by a constitutive androstane receptor-dependent mechanism. Mol. Pharmacol. 2013, 84, 774–786. [Google Scholar] [CrossRef] [Green Version]
- Tripathi, V.K.; Kumar, V.; Pandey, A.; Vatsa, P.; Dhasmana, A.; Singh, R.P.; Appikonda, S.H.C.; Hwang, I.; Lohani, M. Monocrotophos Induces the Expression of Xenobiotic Metabolizing Cytochrome P450s (CYP2C8 and CYP3A4) and Neurotoxicity in Human Brain Cells. Mol. Neurobiol. 2017, 54, 3633–3651. [Google Scholar] [CrossRef]
- Ghosh, C.; Puvenna, V.; Gonzalez-Martinez, J.; Janigro, D.; Marchi, N. Blood-Brain Barrier P450 Enzymes and Multidrug Transporters in Drug Resistance: A Synergistic Role in Neurological Diseases. Curr. Drug Metab. 2011, 12, 742–749. [Google Scholar] [CrossRef] [Green Version]
- Chakraborty, S.; Kanakasabai, S.; Bright, J.J. Constitutive androstane receptor agonist CITCO inhibits growth and expansion of brain tumour stem cells. Br. J. Cancer 2011, 104, 448–459. [Google Scholar] [CrossRef] [PubMed]
- Maglich, J.M.; Parks, D.J.; Moore, L.B.; Collins, J.L.; Goodwin, B.; Billin, A.N.; Stoltz, C.A.; Kliewer, S.A.; Lambert, M.H.; Willson, T.M.; et al. Identification of a Novel Human Constitutive Androstane Receptor (CAR) Agonist and Its Use in the Identification of CAR Target Genes. J. Biol. Chem. 2003, 278, 17277–17283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lemmen, J.; Tozakidis, I.E.P.; Bele, P.; Galla, H.J. Constitutive androstane receptor upregulates Abcb1 and Abcg2 at the blood-brain barrier after CITCO activation. Brain Res. 2013, 1501, 68–80. [Google Scholar] [CrossRef]
- Koemans, T.S.; Kleefstra, T.; Chubak, M.C.; Stone, M.H.; Reijnders, M.R.F.; de Munnik, S.; Willemsen, M.H.; Fenckova, M.; Stumpel, C.T.R.M.; Bok, L.A.; et al. Functional convergence of histone methyltransferases EHMT1 and KMT2C involved in intellectual disability and autism spectrum disorder. PLoS Genet. 2017, 13. [Google Scholar] [CrossRef]
- Boussadia, B.; Gangarossa, G.; Mselli-Lakhal, L.; Rousset, M.-C.; de Bock, F.; Lassere, F.; Ghosh, C.; Pascussi, J.-M.; Janigro, D.; Marchi, N. Lack of CAR impacts neuronal function and cerebrovascular integrity in vivo. Exp. Neurol. 2016, 283, 39–48. [Google Scholar] [CrossRef]
- Torres-Vergara, P.; Ho, Y.S.; Espinoza, F.; Nualart, F.; Escudero, C.; Penny, J. The constitutive androstane receptor and pregnane X receptor in the brain. Br. J. Pharmacol. 2020, 177, 2666–2682. [Google Scholar] [CrossRef] [PubMed]
- Davidson, T.L.; Monnot, A.; Neal, A.U.; Martin, A.A.; Horton, J.J.; Zheng, W. The effects of a high-energy diet on hippocampal-dependent discrimination performance and blood-brain barrier integrity differ for diet-induced obese and diet-resistant rats. Physiol. Behav. 2012, 107, 26–33. [Google Scholar] [CrossRef] [Green Version]
- Ferro, J.M.; Oliveira, S.N.; Correia, L. Neurologic manifestations of inflammatory bowel diseases. In Handbook of Clinical Neurology; Elsevier B.V.: Amsterdam, The Netherlands, 2014; Volume 120, pp. 595–605. [Google Scholar]
- Casella, G.; Tontini, G.E.; Bassotti, G.; Pastorelli, L.; Villanacci, V.; Spina, L.; Baldini, V.; Vecchi, M. Neurological disorders and inflammatory bowel diseases. World J. Gastroenterol. 2014, 20, 8764–8782. [Google Scholar]
- Miehsler, W.; Reinisch, W.; Valic, E.; Osterode, W.; Tillinger, W.; Feichtenschlager, T.; Grisar, J.; Machold, K.; Scholz, S.; Vogelsang, H.; et al. Is inflammatory bowel disease an independent and disease specific risk factor for thromboembolism? Gut 2004, 53, 542–548. [Google Scholar] [CrossRef] [Green Version]
- Varatharaj, A.; Galea, I. The blood-brain barrier in systemic inflammation. Brain. Behav. Immun. 2017, 60, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duszka, K.; Wahli, W. Enteric Microbiota–Gut–Brain Axis from the Perspective of Nuclear Receptors. Int. J. Mol. Sci. 2018, 19, 2210. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
| Mice | Human | |
|---|---|---|
| Phase I | Cyp1a1, Cyp1a2, Cyp2a4, Cyp2b10, Cyp2c29, Cyp2c37, Cyp2c55, Cyp3a11, Nqo1, Aldh1a1, Aldh1a7, Akr1b7, Ces6 | CYP1A1, CYP1A2, CYP2B6, CYP2C8, CYP2C9, CYP2C19, CYP3A4, CYP3A5 |
| Phase II | Ugt1a1, Ugt1a9, Ugt2b34, Ugt2b35, Ugt2b36, Sult1e1, Sult2a1, Sult2a2, Sult3a1, Sult5a1, Gsta1, Gsta4, Gstm1, Gstm2, Gstm3, Gstm4, Gstp, Gstt1 | UGT1A1, SULT2A1 |
| Transporters | Mrp2, Mrp3, Mrp4, Oatp1a4 | MDR1, PTGS2, GCK, PTPRN, ATP2B2 |
| Contaminants | Species | References |
|---|---|---|
| Diphenamid (Pesticide) | Human | [33] |
| Phenothrin (Pesticide) | Human | [33] |
| Permethrin (Pesticide) | Rat | [34] |
| Perfluorocarboxylic acid, PFCA (Detergent) | Mice | [35,36] |
| Perfluorooctanoic acid, PFOA (Detergent) | Mice | [3,37] |
| Perfluorooctanesulfonic acid PFOS (Detergent) | Rat | [38] |
| Alachlor (Pesticide) | Mice | [39] |
| Arsenite (Chemical) | Mice | [39] |
| Azo dyes (Paint) | Mice/Rat | [40] |
| Bisphenol A (Chemical) | Mice | [39] |
| Butylate (Pesticide) | Mice | [39] |
| Chlorpropham (Pesticide) | Mice | [39] |
| Chlorpyrifos (Pesticide) | Mice | [39] |
| Cypermythrin (Pesticide) | Mice | [39] |
| Cyproconazole (Pesticide) | Mice | [41] |
| DBP, Di-n-butylphtalate (Plasticizer) | Rat | [42] |
| DDE, Dichlorodiphenyldichloroethylene (Pesticide) | Rat | [43] |
| Di-isononyl phthalate (DiBP) (Plasticizer) | Human | [44] |
| O,p-DDT,1,1,1-Tichloro-2-(2-chlorophenyl)2-(4-chlorophenyl)ethane (Pesticide) | Mice/Rat | [43,45] |
| DEHP (Plasticizer) | Mice/Human | [39,46] |
| Dieldrine (Pesticide) | Mice | [47] |
| Endosulfan (Pesticide) | Mice/Human | [39,48] |
| Fernitrothion (Pesticide) | Mice | [39] |
| Polycyclic aromatic hydrocarbons | Mice | [49] |
| Imazalil (Pesticide) | Mice | [39] |
| Kepone (Pesticide) | Mice | [39] |
| MEHP (Plasticizer) | Mice | [39] |
| Metolachlor (Pesticide) | Mice | [39] |
| Methoxychlor (Pesticide) and metabolites | Mice/Rat/Human | [39,48,50] |
| Monosodium methane arsenate | Mice | [39] |
| Nonylphenol (Plasticizer) | Human | [4] |
| Parathion (Pesticide) | Mice | [4] |
| PCB Polychlorobiphenyles (Chemical derivatives) | Mice | [45] |
| Propachlor (Pesticide) | Mice | [39] |
| 2,3,7,8-Tetrachlorodibenzo-p-dioxin (TCDD) | Mice | [51] |
| SSS-Tributylphosphorotithioate (Pesticide) | Mice | [39] |
| Triclopyr (Pesticide) | Mice | [39] |
| Drugs | Species | Action | References |
|---|---|---|---|
| Neticonazole | Human | Activator | [33] |
| Rimcazole | Human | Activator | [33] |
| Sorafenib | Human | Inhibitor | [52] |
| Rimonabant | Human | Inhibitor | [52] |
| DL5050 | Human | Activator | [53] |
| Valproic Acid | Human | Activator | [54] |
| Acetaminophen | Mice | Activator | [55,56] |
| Triazole Antifungals | Mice | Activator | [41,57] |
| Artemisinin | Mice/Human | Activator | [58,59] |
| Benzodiazepines | Human | Inhibitor | [60] |
| Clotrimoxazole | Human | Inhibitor | [31] |
| Cocaine | Human | Inhibitor | [61] |
| Dexamethasone | Human | Activator | [62,63] |
| Ketoconazole | Human | Inhibitor | [64] |
| Meclizine | Mice/Human | Activator (Mice) Inhibitor (Human) | [65] |
| Metamizole | Human | Activator | [66] |
| Methotrexate | Mice | Inhibitor | [67,68] |
| Orphenadrine | Rat | Activator | [69] |
| Phenobarbital | Mice, Rat | Activator | [70,71] |
| Phenytoin | Human | Activator | [72,73,74] |
| Statins | Human | Activator | [75,76] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oliviero, F.; Lukowicz, C.; Boussadia, B.; Forner-Piquer, I.; Pascussi, J.-M.; Marchi, N.; Mselli-Lakhal, L. Constitutive Androstane Receptor: A Peripheral and a Neurovascular Stress or Environmental Sensor. Cells 2020, 9, 2426. https://doi.org/10.3390/cells9112426
Oliviero F, Lukowicz C, Boussadia B, Forner-Piquer I, Pascussi J-M, Marchi N, Mselli-Lakhal L. Constitutive Androstane Receptor: A Peripheral and a Neurovascular Stress or Environmental Sensor. Cells. 2020; 9(11):2426. https://doi.org/10.3390/cells9112426
Chicago/Turabian StyleOliviero, Fabiana, Céline Lukowicz, Badreddine Boussadia, Isabel Forner-Piquer, Jean-Marc Pascussi, Nicola Marchi, and Laila Mselli-Lakhal. 2020. "Constitutive Androstane Receptor: A Peripheral and a Neurovascular Stress or Environmental Sensor" Cells 9, no. 11: 2426. https://doi.org/10.3390/cells9112426