Intraarticular Ligament Degeneration Is Interrelated with Cartilage and Bone Destruction in Osteoarthritis


Abstract
1. Introduction
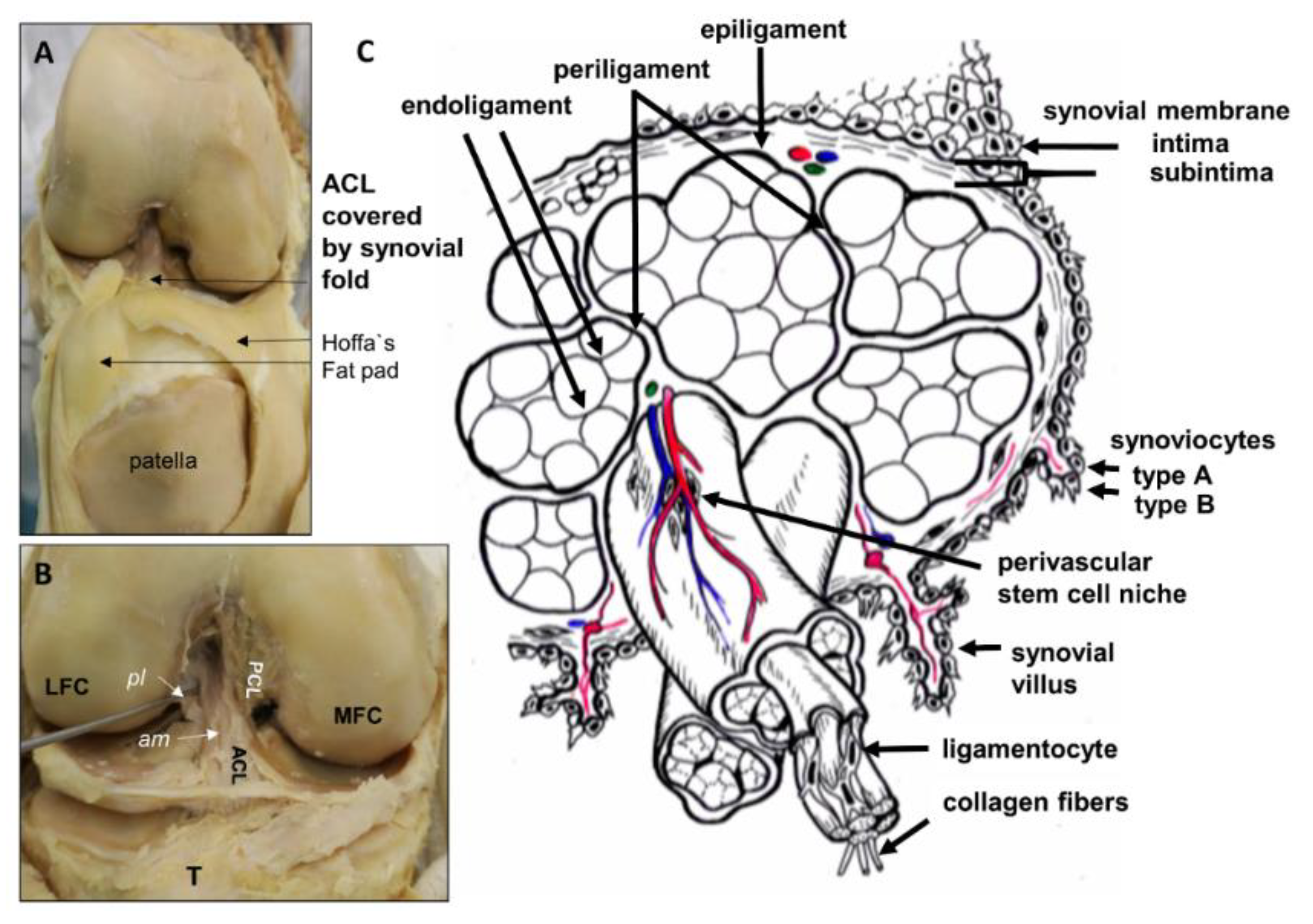
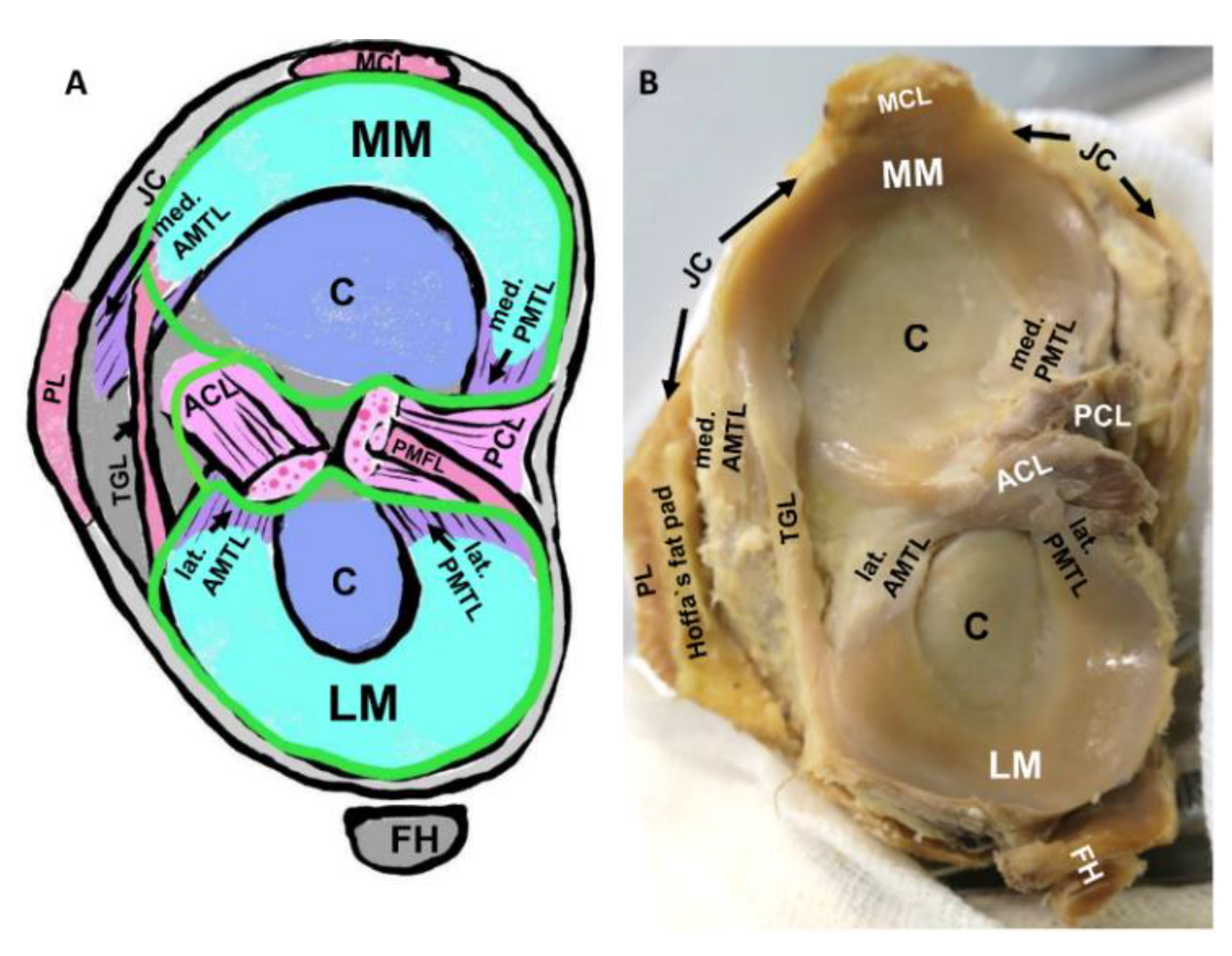
2. Anatomy of Intraarticular Ligaments of the Knee
2.1. Anterior Cruciate Ligament (ACL)
2.2. Meniscotibial Ligaments
2.3. Blood Supply of the ACL and Meniscotibial Ligaments
3. Role of Intrinsic Stem Cells in Ligaments and the ACL
Inflammation and Stem Cells Commitment
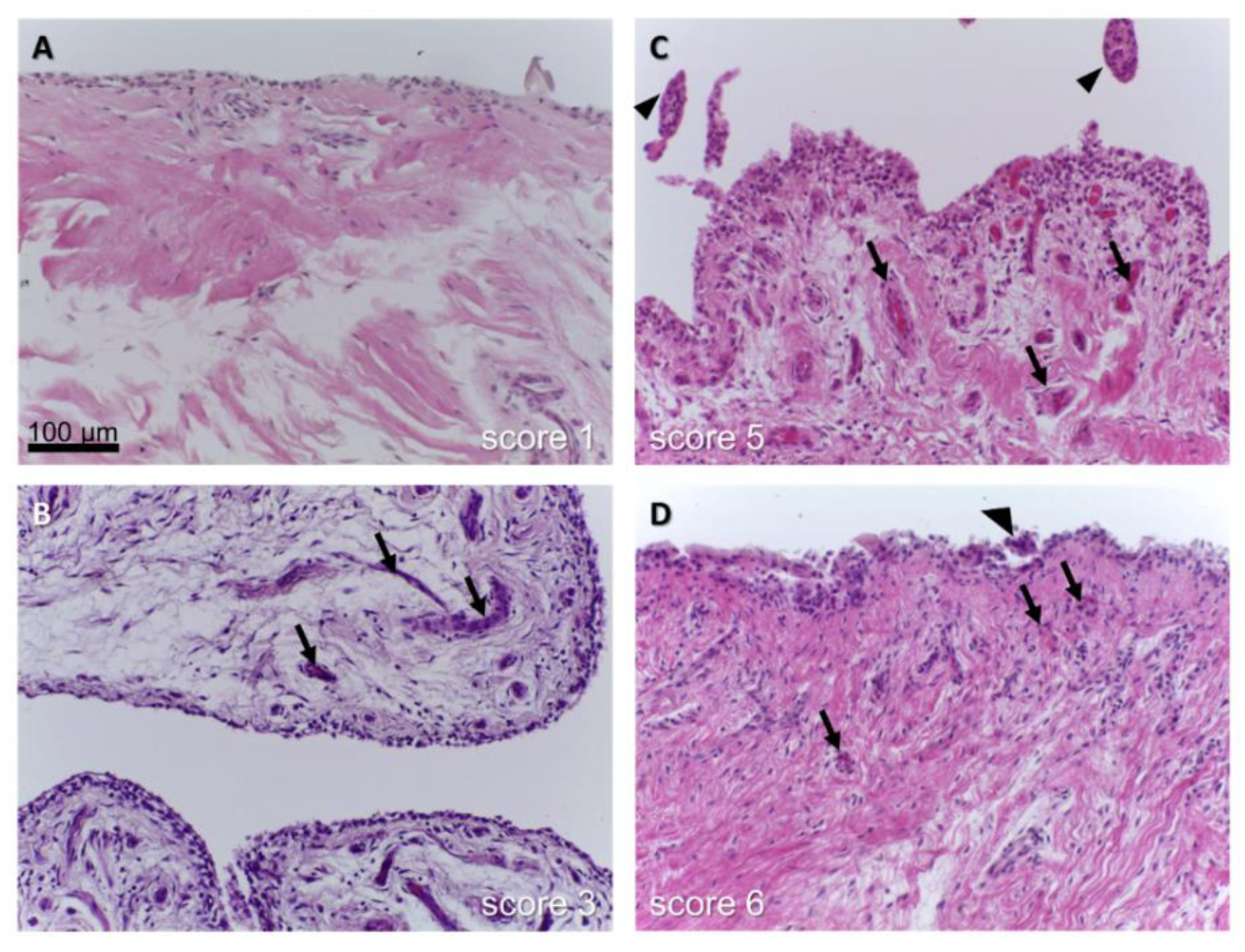
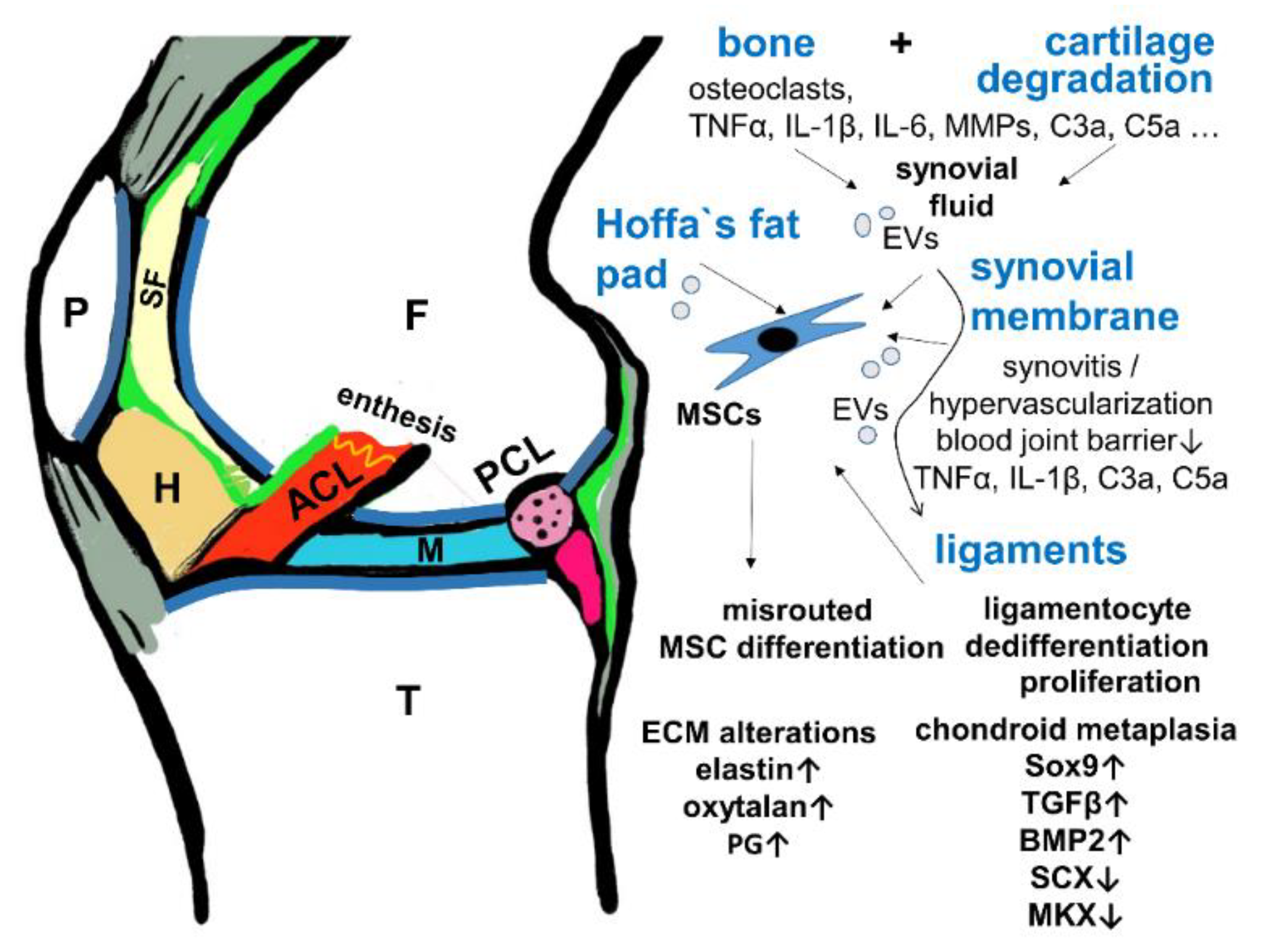
4. Contribution of Synovial Fluid and Synovitis to Ligament Degeneration
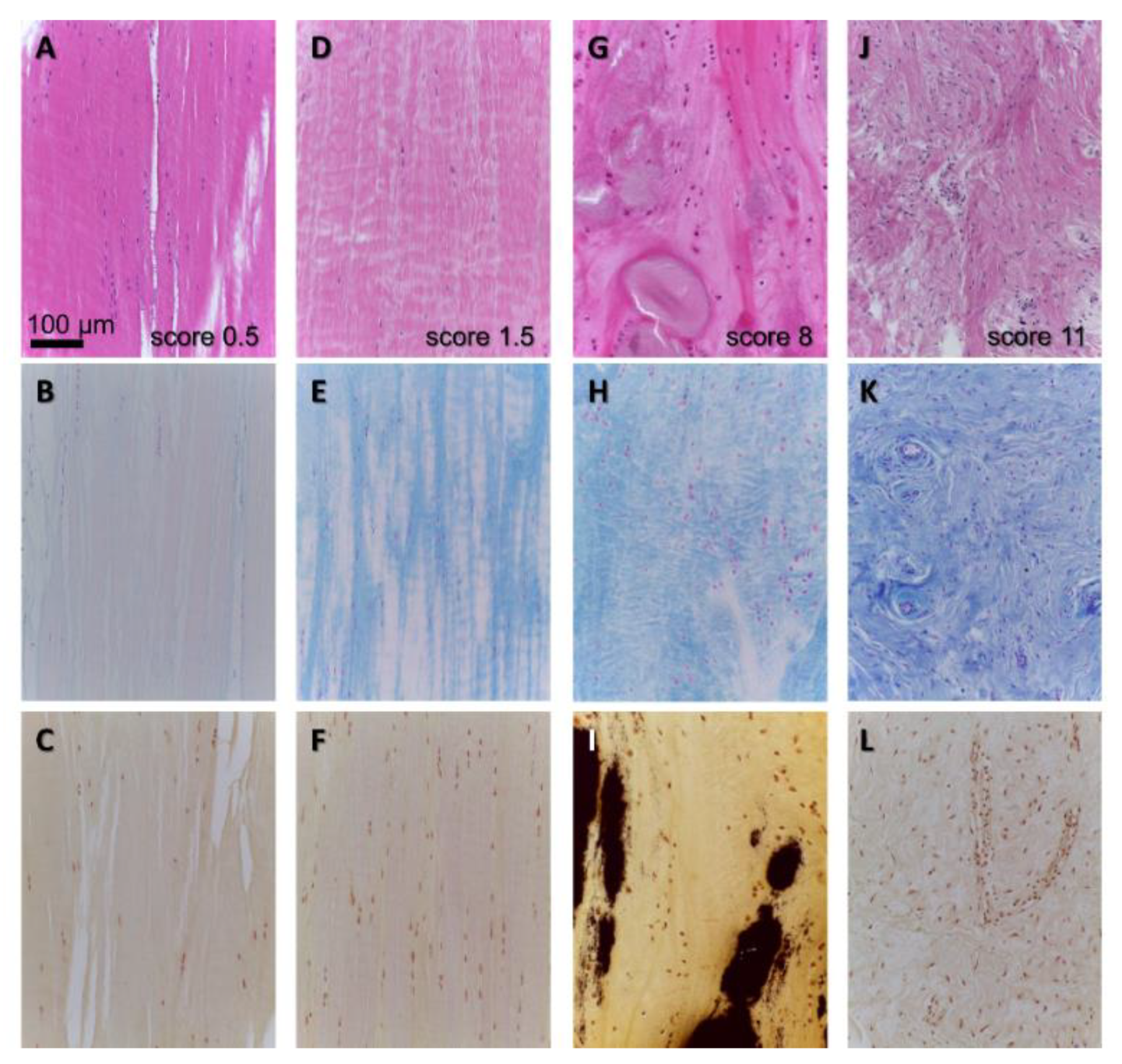
5. Ligament Histopathology in OA and Aging
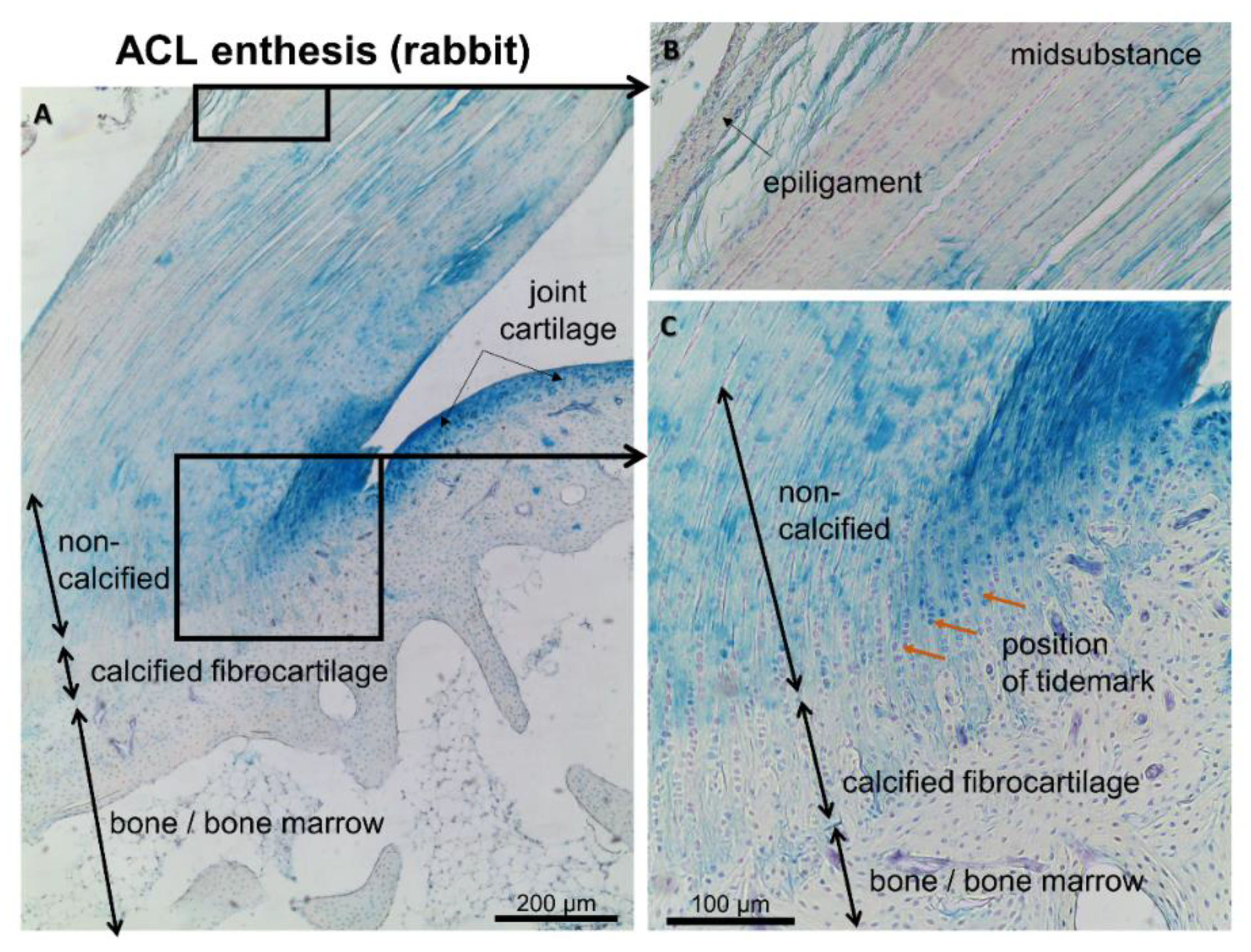
6. Involvement of the Ligament Enthesis in the OA Pathogenesis?
7. Candidate Signaling Pathways Involved in Ligament Degeneration
7.1. Transforming Growth Factor Superfamily and Ligament Degeneration
7.2. Complement Cascade
8. Materials and Methods
9. Conclusions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ACL | anterior cruciate ligament |
| ACL-MSC | ACL-derived mesenchymal stromal cells |
| AMTL | anterior meniscotibial ligament |
| ASC | adipose tissue-derived mesenchymal stromal cells |
| α-SMA | α-smooth muscle actin |
| BM-MSC | bone marrow derived mesenchymal stromal cells |
| BMP | Bone morphogenetic protein |
| CaCL | caudal cruciate ligament (ACL in quadrupedes) |
| CrCL | cranial cruciate ligament (ACL in quadrupedes) |
| CRP | complement regulatory protein |
| ECM | extracellular matrix |
| HE | hematoxylin Eosin |
| HIF1α | hypoxia inducible factor |
| IL | interleukin |
| LSPC | ligament stem / progenitor cells |
| MKX | Mohawk |
| MMP | matrix metalloproteinases |
| MRI | magnetic resonance imaging |
| MSCs | mesenchymal stromal cells |
| OA | osteoarthritis |
| Oct | octamer binding protein |
| PCL | posterior cruciate ligament |
| PG | proteoglycan |
| PMTL | posterior meniscotibial ligament |
| PTS | posterior tibial slope |
| SCX | scleraxis |
| Sox | sex determining region Y-box |
| SSEA | stage specific antigen |
| STRO-1 | stroma 1 mesenchymal stroma cell marker |
| TGFβ | Transforming growth factor β |
| TNFα | tumor necrosis factor α |
| TSPCs | tendon stem/progenitor cells |
References
- Donell, S. Subchondral bone remodelling in osteoarthritis. EFORT Open Rev. 2019, 4, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Amini, M.; Nazemi, S.M.; Lanovaz, J.L.; Kontulainen, S.; Masri, B.A.; Wilson, D.R.; Szyszkowski, W.; Johnston, J.D. Individual and combined effects of OA-related subchondral bone alterations on proximal tibial surface stiffness: A parametric finite element modeling study. Med. Eng. Phys. 2015, 37, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Ding, M.; Dalstra, M.; Linde, F.; Hvid, I. Changes in the stiffness of the human tibial cartilage-bone complex in early-stage osteoarthrosis. Acta Orthop. Scand. 1998, 69, 358–362. [Google Scholar] [CrossRef] [PubMed]
- Nagelli, C.V.; Cook, J.L.; Kuroki, K.; Bozynski, C.; Ma, R.; Hewett, T.E. Does Anterior Cruciate Ligament Innervation Matter for Joint Function and Development of Osteoarthritis? J. Knee Surg. 2017, 30, 364–371. [Google Scholar] [CrossRef]
- Maletius, W.; Messner, K. Eighteen- to twenty-four-year follow-up after complete rupture of the anterior cruciate ligament. Am. J. Sports Med. 1999, 27, 711–717. [Google Scholar] [CrossRef] [PubMed]
- Mabey, T.; Honsawek, S. Cytokines as biochemical markers for knee osteoarthritis. World J. Orthop. 2015, 6, 95–105. [Google Scholar] [CrossRef] [PubMed]
- Maldonado, M.; Nam, J. The role of changes in extracellular matrix of cartilage in the presence of inflammation on the pathology of osteoarthritis. Biomed. Res. Int. 2013, 2013, 284873. [Google Scholar] [CrossRef] [PubMed]
- Silawal, S.; Triebel, J.; Bertsch, T.; Schulze-Tanzil, G. Osteoarthritis and the Complement Cascade. Clin. Med. Insights Arthritis Musculoskelet. Disord. 2018, 11, 1179544117751430. [Google Scholar] [CrossRef]
- Wang, Q.; Rozelle, A.L.; Lepus, C.M.; Scanzello, C.R.; Song, J.J.; Larsen, D.M.; Crish, J.F.; Bebek, G.; Ritter, S.Y.; Lindstrom, T.M.; et al. Identification of a central role for complement in osteoarthritis. Nat. Med. 2011, 17, 1674–1679. [Google Scholar] [CrossRef]
- De Kroon, L.M.G.; van den Akker, G.G.H.; Brachvogel, B.; Narcisi, R.; Belluoccio, D.; Jenner, F.; Bateman, J.F.; Little, C.B.; Brama, P.A.J.; Blaney Davidson, E.N.; et al. Identification of TGFbeta-related genes regulated in murine osteoarthritis and chondrocyte hypertrophy by comparison of multiple microarray datasets. Bone 2018, 116, 67–77. [Google Scholar] [CrossRef]
- Hasegawa, A.; Otsuki, S.; Pauli, C.; Miyaki, S.; Patil, S.; Steklov, N.; Kinoshita, M.; Koziol, J.; D’Lima, D.D.; Lotz, M.K. Anterior cruciate ligament changes in the human knee joint in aging and osteoarthritis. Arthritis Rheum. 2012, 64, 696–704. [Google Scholar] [CrossRef]
- Hasegawa, A.; Nakahara, H.; Kinoshita, M.; Asahara, H.; Koziol, J.; Lotz, M.K. Cellular and extracellular matrix changes in anterior cruciate ligaments during human knee aging and osteoarthritis. Arthritis Res. Ther. 2013, 15, R29. [Google Scholar] [CrossRef]
- Ruschke, K.; Meier, C.; Ullah, M.; Krebs, A.C.; Silberreis, K.; Kohl, B.; Knaus, P.; Jagielski, M.; Arens, S.; Schulze-Tanzil, G. Bone morphogenetic protein 2/SMAD signalling in human ligamentocytes of degenerated and aged anterior cruciate ligaments. Osteoarthr. Cartil. 2016, 24, 1816–1825. [Google Scholar] [CrossRef]
- Abraham, A.C.; Donahue, T.L. From meniscus to bone: A quantitative evaluation of structure and function of the human meniscal attachments. Acta Biomater. 2013, 9, 6322–6329. [Google Scholar] [CrossRef]
- Loeser, R.F.; Goldring, S.R.; Scanzello, C.R.; Goldring, M.B. Osteoarthritis: A disease of the joint as an organ. Arthritis Rheum. 2012, 64, 1697–1707. [Google Scholar] [CrossRef]
- Chen, D.; Shen, J.; Zhao, W.; Wang, T.; Han, L.; Hamilton, J.L.; Im, H.J. Osteoarthritis: Toward a comprehensive understanding of pathological mechanism. Bone Res. 2017, 5, 16044. [Google Scholar] [CrossRef]
- Aspden, R.M.; Saunders, F.R. Osteoarthritis as an organ disease: From the cradle to the grave. Eur. Cell Mater. 2019, 37, 74–87. [Google Scholar] [CrossRef]
- Link, T.M.; Li, X. Bone marrow changes in osteoarthritis. Semin. Musculoskelet. Radiol. 2011, 15, 238–246. [Google Scholar] [CrossRef]
- Xu, L.; Hayashi, D.; Roemer, F.W.; Felson, D.T.; Guermazi, A. Magnetic resonance imaging of subchondral bone marrow lesions in association with osteoarthritis. Semin. Arthritis Rheum. 2012, 42, 105–118. [Google Scholar] [CrossRef]
- Muratovic, D.; Cicuttini, F.; Wluka, A.; Findlay, D.; Wang, Y.; Otto, S.; Taylor, D.; Humphries, J.; Lee, Y.; Labrinidis, A.; et al. Bone marrow lesions detected by specific combination of MRI sequences are associated with severity of osteochondral degeneration. Arthritis Res. Ther. 2016, 18, 54. [Google Scholar] [CrossRef]
- Li, G.; Yin, J.; Gao, J.; Cheng, T.S.; Pavlos, N.J.; Zhang, C.; Zheng, M.H. Subchondral bone in osteoarthritis: Insight into risk factors and microstructural changes. Arthritis Res. Ther. 2013, 15, 223. [Google Scholar] [CrossRef]
- Scanzello, C.R.; Goldring, S.R. The role of synovitis in osteoarthritis pathogenesis. Bone 2012, 51, 249–257. [Google Scholar] [CrossRef]
- Clockaerts, S.; Bastiaansen-Jenniskens, Y.M.; Runhaar, J.; Van Osch, G.J.; Van Offel, J.F.; Verhaar, J.A.; De Clerck, L.S.; Somville, J. The infrapatellar fat pad should be considered as an active osteoarthritic joint tissue: A narrative review. Osteoarthr. Cartil. 2010, 18, 876–882. [Google Scholar] [CrossRef]
- McGonagle, D.; Tan, A.L.; Grainger, A.J.; Benjamin, M. Heberden’s nodes and what Heberden could not see: The pivotal role of ligaments in the pathogenesis of early nodal osteoarthritis and beyond. Rheumatology (Oxford) 2008, 47, 1278–1285. [Google Scholar] [CrossRef]
- McGonagle, D.; Aydin, S.Z.; Tan, A.L. The synovio-entheseal complex and its role in tendon and capsular associated inflammation. J. Rheumatol. Suppl. 2012, 89, 11–14. [Google Scholar] [CrossRef]
- Salaffi, F.; Ciapetti, A.; Carotti, M. The sources of pain in osteoarthritis: A pathophysiological review. Reumatismo 2014, 66, 57–71. [Google Scholar] [CrossRef]
- Rumian, A.P.; Wallace, A.L.; Birch, H.L. Tendons and ligaments are anatomically distinct but overlap in molecular and morphological features--a comparative study in an ovine model. J. Orthop. Res. 2007, 25, 458–464. [Google Scholar] [CrossRef]
- Li, B.; Luo, X.D.; Wen, Y. Changes in mechanoreceptors in rabbits’ anterior cruciate ligaments with age. J. Mol. Histol. 2019, 50, 229–237. [Google Scholar] [CrossRef]
- Nyland, J.; Gamble, C.; Franklin, T.; Caborn, D.N.M. Permanent knee sensorimotor system changes following ACL injury and surgery. Knee Surg. Sports Traumatol. Arthrosc. 2017, 25, 1461–1474. [Google Scholar] [CrossRef]
- Dyment, N.A.; Galloway, J.L. Regenerative biology of tendon: Mechanisms for renewal and repair. Curr. Mol. Biol. Rep. 2015, 1, 124–131. [Google Scholar] [CrossRef]
- Chen, B.; Zhang, J.; Nie, D.; Zhao, G.; Fu, F.H.; Wang, J.H. Characterization of the structure of rabbit anterior cruciate ligament and its stem/progenitor cells. J. Cell. Biochem. 2018. [Google Scholar] [CrossRef]
- Park, I.S.; Kim, S.J. Agenesis of the posterolateral bundle of anterior cruciate ligament. Knee Surg. Sports Traumatol. Arthrosc. 2013, 21, 596–598. [Google Scholar] [CrossRef]
- Ellison, A.E.; Berg, E.E. Embryology, anatomy, and function of the anterior cruciate ligament. Orthop. Clin. North Am. 1985, 16, 3–14. [Google Scholar]
- De Rooster, H.; de Bruin, T.; van Bree, H. Morphologic and functional features of the canine cruciate ligaments. Vet. Surg. 2006, 35, 769–780. [Google Scholar] [CrossRef]
- Haus, J.; Refior, H.J. A study of the synovial and ligamentous structure of the anterior cruciate ligament. Int. Orthop. 1987, 11, 117–124. [Google Scholar] [CrossRef]
- Georgiev, G.P.; Kotov, G.; Iliev, A.; Slavchev, S.; Ovtscharoff, W.; Landzhov, B. A comparative study of the epiligament of the medial collateral and the anterior cruciate ligament in the human knee. Immunohistochemical analysis of collagen type I and V and procollagen type III. Ann. Anat. 2019, 224, 88–96. [Google Scholar] [CrossRef]
- Kharaz, Y.A.; Canty-Laird, E.G.; Tew, S.R.; Comerford, E.J. Variations in internal structure, composition and protein distribution between intra- and extra-articular knee ligaments and tendons. J. Anat. 2018, 232, 943–955. [Google Scholar] [CrossRef]
- Lorda-Diez, C.I.; Canga-Villegas, A.; Cerezal, L.; Plaza, S.; Hurle, J.M.; Garcia-Porrero, J.A.; Montero, J.A. Comparative transcriptional analysis of three human ligaments with distinct biomechanical properties. J. Anat. 2013, 223, 593–602. [Google Scholar] [CrossRef]
- McGonagle, D.; Tan, A.L.; Carey, J.; Benjamin, M. The anatomical basis for a novel classification of osteoarthritis and allied disorders. J. Anat. 2010, 216, 279–291. [Google Scholar] [CrossRef]
- Choi, J.Y.; Chang, E.Y.; Cunha, G.M.; Tafur, M.; Statum, S.; Chung, C.B. Posterior medial meniscus root ligament lesions: MRI classification and associated findings. AJR Am. J. Roentgenol. 2014, 203, 1286–1292. [Google Scholar] [CrossRef]
- Seitz, H.; Hausner, T.; Schlenz, I.; Lang, S.; Eschberger, J. Vascular anatomy of the ovine anterior cruciate ligament. A macroscopic, histological and radiographic study. Arch. Orthop. Trauma Surg. 1997, 116, 19–21. [Google Scholar] [CrossRef]
- Amiel, D.; Abel, M.F.; Kleiner, J.B.; Lieber, R.L.; Akeson, W.H. Synovial fluid nutrient delivery in the diathrial joint: An analysis of rabbit knee ligaments. J. Orthop. Res. 1986, 4, 90–95. [Google Scholar] [CrossRef]
- Whiteside, L.A.; Sweeney, R.E., Jr. Nutrient pathways of the cruciate ligaments. An experimental study using the hydrogen wash-out technique. J. Bone Joint Surg. Am. 1980, 62, 1176–1180. [Google Scholar] [CrossRef]
- Skyhar, M.J.; Danzig, L.A.; Hargens, A.R.; Akeson, W.H. Nutrition of the anterior cruciate ligament. Effects of continuous passive motion. Am. J. Sports Med. 1985, 13, 415–418. [Google Scholar] [CrossRef]
- Paulsen, F.; Sobotta, W.J. Atlas der Anatomie des Menschens, 23 Auflage; Urban und Fischer: Munich, Germany, 2018. [Google Scholar]
- Wang, Y.; Xu, J.; Zhang, X.; Wang, C.; Huang, Y.; Dai, K.; Zhang, X. TNF-alpha-induced LRG1 promotes angiogenesis and mesenchymal stem cell migration in the subchondral bone during osteoarthritis. Cell Death Dis. 2017, 8, e2715. [Google Scholar] [CrossRef]
- Henrotin, Y.; Pesesse, L.; Lambert, C. Targeting the synovial angiogenesis as a novel treatment approach to osteoarthritis. Ther. Adv. Musculoskelet. Dis. 2014, 6, 20–34. [Google Scholar] [CrossRef]
- Mapp, P.I.; Walsh, D.A. Mechanisms and targets of angiogenesis and nerve growth in osteoarthritis. Nat. Rev. Rheumatol. 2012, 8, 390–398. [Google Scholar] [CrossRef]
- Korchi, A.M.; Cengarle-Samak, A.; Okuno, Y.; Martel-Pelletier, J.; Pelletier, J.P.; Boesen, M.; Doyon, J.; Bodson-Clermont, P.; Lussier, B.; Heon, H.; et al. Inflammation and Hypervascularization in a Large Animal Model of Knee Osteoarthritis: Imaging with Pathohistologic Correlation. J. Vasc. Interv. Radiol. 2019, 30, 1116–1127. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, J.H. Production of PGE(2) increases in tendons subjected to repetitive mechanical loading and induces differentiation of tendon stem cells into non-tenocytes. J. Orthop. Res. 2010, 28, 198–203. [Google Scholar]
- Burssens, A.; Forsyth, R.; Bongaerts, W.; Jagodzinski, M.; Mahieu, N.; Praet, M.; Victor, J. Arguments for an increasing differentiation towards fibrocartilaginous components in midportion Achilles tendinopathy. Knee Surg. Sports Traumatol. Arthrosc. 2013, 21, 1459–1467. [Google Scholar] [CrossRef]
- Lin, D.; Alberton, P.; Caceres, M.D.; Volkmer, E.; Schieker, M.; Docheva, D. Tenomodulin is essential for prevention of adipocyte accumulation and fibrovascular scar formation during early tendon healing. Cell Death Dis. 2017, 8, e3116. [Google Scholar] [CrossRef]
- Tempfer, H.; Wagner, A.; Gehwolf, R.; Lehner, C.; Tauber, M.; Resch, H.; Bauer, H.C. Perivascular cells of the supraspinatus tendon express both tendon- and stem cell-related markers. Histochem. Cell Biol. 2009, 131, 733–741. [Google Scholar] [CrossRef]
- Mienaltowski, M.J.; Adams, S.M.; Birk, D.E. Regional differences in stem cell/progenitor cell populations from the mouse achilles tendon. Tissue Eng. Part A. 2013, 19, 199–210. [Google Scholar] [CrossRef]
- Schneider, M.; Docheva, D. Mysteries behind the Cellular Content of Tendon Tissues. J. Am. Acad. Orthop. Surg. 2017, 25, e289–e290. [Google Scholar] [CrossRef]
- Mifune, Y.; Matsumoto, T.; Ota, S.; Nishimori, M.; Usas, A.; Kopf, S.; Kuroda, R.; Kurosaka, M.; Fu, F.H.; Huard, J. Therapeutic potential of anterior cruciate ligament-derived stem cells for anterior cruciate ligament reconstruction. Cell Transplant. 2012, 21, 1651–1665. [Google Scholar] [CrossRef]
- Bi, Y.; Ehirchiou, D.; Kilts, T.M.; Inkson, C.A.; Embree, M.C.; Sonoyama, W.; Li, L.; Leet, A.I.; Seo, B.M.; Zhang, L.; et al. Identification of tendon stem/progenitor cells and the role of the extracellular matrix in their niche. Nat. Med. 2007, 13, 1219–1227. [Google Scholar] [CrossRef]
- Lui, P.P.; Kong, S.K.; Lau, P.M.; Wong, Y.M.; Lee, Y.W.; Tan, C.; Wong, O.T. Immunogenicity and escape mechanisms of allogeneic tendon-derived stem cells. Tissue Eng. Part A. 2014, 20, 3010–3020. [Google Scholar] [CrossRef]
- Hofer, H.R.; Tuan, R.S. Secreted trophic factors of mesenchymal stem cells support neurovascular and musculoskeletal therapies. Stem Cell Res. Ther. 2016, 7, 131. [Google Scholar] [CrossRef]
- Rosenthal, A.K.; Gohr, C.M.; Ninomiya, J.; Wakim, B.T. Proteomic analysis of articular cartilage vesicles from normal and osteoarthritic cartilage. Arthritis Rheum. 2011, 63, 401–411. [Google Scholar] [CrossRef]
- Withrow, J.; Murphy, C.; Liu, Y.; Hunter, M.; Fulzele, S.; Hamrick, M.W. Extracellular vesicles in the pathogenesis of rheumatoid arthritis and osteoarthritis. Arthritis Res. Ther. 2016, 18, 286. [Google Scholar] [CrossRef]
- Lee, K.J.; Comerford, E.J.; Simpson, D.M.; Clegg, P.D.; Canty-Laird, E.G. Identification and Characterization of Canine Ligament Progenitor Cells and Their Extracellular Matrix Niche. J. Proteome. Res. 2019, 18, 1328–1339. [Google Scholar] [CrossRef]
- Fu, W.; Li, Q.; Tang, X.; Chen, G.; Zhang, C.; Li, J. Mesenchymal stem cells reside in anterior cruciate ligament remnants in situ. Int. Orthop. 2016, 40, 1523–1530. [Google Scholar] [CrossRef]
- Huang, T.F.; Chen, Y.T.; Yang, T.H.; Chen, L.L.; Chiou, S.H.; Tsai, T.H.; Tsai, C.C.; Chen, M.H.; Ma, H.L.; Hung, S.C. Isolation and characterization of mesenchymal stromal cells from human anterior cruciate ligament. Cytotherapy 2008, 10, 806–814. [Google Scholar] [CrossRef]
- Cheng, M.T.; Liu, C.L.; Chen, T.H.; Lee, O.K. Optimization of culture conditions for stem cells derived from human anterior cruciate ligament and bone marrow. Cell Transplant. 2014, 23, 791–803. [Google Scholar] [CrossRef]
- Zhang, J.; Pan, T.; Im, H.J.; Fu, F.H.; Wang, J.H. Differential properties of human ACL and MCL stem cells may be responsible for their differential healing capacity. BMC Med. 2011, 9, 68. [Google Scholar] [CrossRef]
- Ogata, Y.; Mabuchi, Y.; Shinoda, K.; Horiike, Y.; Mizuno, M.; Otabe, K.; Suto, E.G.; Suzuki, N.; Sekiya, I.; Akazawa, C. Anterior cruciate ligament-derived mesenchymal stromal cells have a propensity to differentiate into the ligament lineage. Regen. Ther. 2018, 8, 20–28. [Google Scholar] [CrossRef]
- Prager, P.; Kunz, M.; Ebert, R.; Klein-Hitpass, L.; Sieker, J.; Barthel, T.; Jakob, F.; Konrads, C.; Steinert, A. Mesenchymal Stem Cells Isolated from the Anterior Cruciate Ligament: Characterization and Comparison of Cells from Young and Old Donors. Knee Surg. Relat. Res. 2018, 30, 193–205. [Google Scholar] [CrossRef]
- Dragoo, J.L.; Chang, W. Arthroscopic Harvest of Adipose-Derived Mesenchymal Stem Cells From the Infrapatellar Fat Pad. Am. J. Sports Med. 2017, 45, 3119–3127. [Google Scholar] [CrossRef]
- Toyoda, E.; Sato, M.; Takahashi, T.; Maehara, M.; Nakamura, Y.; Mitani, G.; Takagaki, T.; Hamahashi, K.; Watanabe, M. Multilineage-differentiating stress-enduring (Muse)-like cells exist in synovial tissue. Regen. Ther. 2019, 10, 17–26. [Google Scholar] [CrossRef]
- Bravo, B.; Guisasola, M.C.; Vaquero, J.; Tirado, I.; Gortazar, A.R.; Forriol, F. Gene expression, protein profiling, and chemotactic activity of infrapatellar fat pad mesenchymal stem cells in pathologies of the knee joint. J. Cell. Physiol. 2019. [Google Scholar] [CrossRef]
- Heldens, G.T.; Blaney Davidson, E.N.; Vitters, E.L.; Schreurs, B.W.; Piek, E.; van den Berg, W.B.; van der Kraan, P.M. Catabolic factors and osteoarthritis-conditioned medium inhibit chondrogenesis of human mesenchymal stem cells. Tissue Eng. Part A 2012, 18, 45–54. [Google Scholar] [CrossRef]
- Brandt, L.; Schubert, S.; Scheibe, P.; Brehm, W.; Franzen, J.; Gross, C.; Burk, J. Tenogenic Properties of Mesenchymal Progenitor Cells Are Compromised in an Inflammatory Environment. Int. J. Mol. Sci. 2018, 19, 2549. [Google Scholar] [CrossRef]
- De Sousa, E.B.; Dos Santos Junior, G.C.; Aguiar, R.P.; da Costa Sartore, R.; de Oliveira, A.C.L.; Almeida, F.C.L.; Neto, V.M.; Aguiar, D.P. Osteoarthritic Synovial Fluid Modulates Cell Phenotype and Metabolic Behavior In Vitro. Stem Cells Int. 2019, 2019, 8169172. [Google Scholar] [CrossRef]
- Wijekoon, H.M.S.; Toyota, K.; Kim, S.; Fang, J.; Bwalya, E.C.; Hosoya, K.; Okumura, M. Differentiation potential of synoviocytes derived from joints with cranial cruciate ligament rupture and medial patella luxation in dogs. Res. Vet. Sci. 2017, 114, 370–377. [Google Scholar] [CrossRef]
- Monibi, F.; Roller, B.L.; Stoker, A.; Garner, B.; Bal, S.; Cook, J.L. Identification of Synovial Fluid Biomarkers for Knee Osteoarthritis and Correlation with Radiographic Assessment. J. Knee Surg. 2016, 29, 242–247. [Google Scholar] [CrossRef]
- Edwards, J.C. Fibroblast biology. Development and differentiation of synovial fibroblasts in arthritis. Arthritis Res. 2000, 2, 344–347. [Google Scholar] [CrossRef]
- Bennike, T.; Ayturk, U.; Haslauer, C.M.; Froehlich, J.W.; Proffen, B.L.; Barnaby, O.; Birkelund, S.; Murray, M.M.; Warman, M.L.; Stensballe, A.; et al. A normative study of the synovial fluid proteome from healthy porcine knee joints. J. Proteome. Res. 2014, 13, 4377–4387. [Google Scholar] [CrossRef]
- Nio, J.; Yokoyama, A.; Okumura, M.; Iwanaga, T. Three-dimensional ultrastructure of synoviocytes in the knee joint of rabbits and morphological changes in osteoarthritis model. Arch. Histol. Cytol. 2002, 65, 189–200. [Google Scholar] [CrossRef]
- Sakuma, E.; Wada, I.; Mabuchi, Y.; Sugimura, I.; Yanagisono, T.; Yamada, E.; Amano, K.; Matsui, N.; Soji, T. Postnatal development of synovial capillaries of rats with special reference to permeability. Kaibogaku Zasshi 2000, 75, 407–411. [Google Scholar]
- Hillen, J.; Geyer, C.; Heitzmann, M.; Beckmann, D.; Krause, A.; Winkler, I.; Pavenstadt, H.; Bremer, C.; Pap, T.; Korb-Pap, A. Structural cartilage damage attracts circulating rheumatoid arthritis synovial fibroblasts into affected joints. Arthritis Res. Ther. 2017, 19, 40. [Google Scholar] [CrossRef]
- Berger, I.; Morawietz, L.; Jakobs, M.; Krenn, V. Value of histological work-up for synovial diseases. Orthopade 2009, 38, 484–490. [Google Scholar] [CrossRef]
- Krenn, V.; Morawietz, L.; Burmester, G.R.; Kinne, R.W.; Mueller-Ladner, U.; Muller, B.; Haupl, T. Synovitis score: Discrimination between chronic low-grade and high-grade synovitis. Histopathology 2006, 49, 358–364. [Google Scholar] [CrossRef]
- Jakobs, M.; Morawietz, L.; Rothschenk, H.; Hopf, T.; Weiner, S.; Schausten, H.; Krukemeyer, M.G.; Krenn, V. Synovitis score: Value of histopathological diagnostics in unclear arthritis. Case reports from rheumatological pathological practice. Z. Rheumatol. 2007, 66, 706–712. [Google Scholar] [CrossRef]
- Doring, A.K.; Junginger, J.; Hewicker-Trautwein, M. Cruciate ligament degeneration and stifle joint synovitis in 56 dogs with intact cranial cruciate ligaments: Correlation of histological findings and numbers and phenotypes of inflammatory cells with age, body weight and breed. Vet. Immunol. Immunopathol. 2018, 196, 5–13. [Google Scholar] [CrossRef]
- Kobayashi, S.; Baba, H.; Uchida, K.; Negoro, K.; Sato, M.; Miyazaki, T.; Nomura, E.; Murakami, K.; Shimizubata, M.; Meir, A. Microvascular system of anterior cruciate ligament in dogs. J. Orthop. Res. 2006, 24, 1509–1520. [Google Scholar] [CrossRef]
- Wang, C.; Mei, H.; Xie, J.; Jiang, J.; Chen, R.; Yin, L.; Fu, C.; Chen, C.; Sung, K.L. Gene expressions of LOXs and MMPs of the ACL fibroblasts cells co-cultured with synovial cells. Sheng Wu Yi Xue Gong Cheng Xue Za Zhi 2013, 30, 777–781. [Google Scholar]
- Nickerson, D.A.; Joshi, R.; Williams, S.; Ross, S.M.; Frank, C. Synovial fluid stimulates the proliferation of rabbit ligament. Fibroblasts in vitro. Clin. Orthop. Relat. Res. 1992, 274, 294–299. [Google Scholar]
- Mullaji, A.B.; Marawar, S.V.; Simha, M.; Jindal, G. Cruciate ligaments in arthritic knees: A histologic study with radiologic correlation. J. Arthroplast. 2008, 23, 567–572. [Google Scholar] [CrossRef]
- Biercevicz, A.M.; Akelman, M.R.; Rubin, L.E.; Walsh, E.G.; Merck, D.; Fleming, B.C. The uncertainty of predicting intact anterior cruciate ligament degeneration in terms of structural properties using T(2)(*) relaxometry in a human cadaveric model. J. Biomech. 2015, 48, 1188–1192. [Google Scholar] [CrossRef][Green Version]
- Cushner, F.D.; La Rosa, D.F.; Vigorita, V.J.; Scuderi, G.R.; Scott, W.N.; Insall, J.N. A quantitative histologic comparison: ACL degeneration in the osteoarthritic knee. J. Arthroplast. 2003, 18, 687–692. [Google Scholar] [CrossRef]
- Kumagai, K.; Sakai, K.; Kusayama, Y.; Akamatsu, Y.; Sakamaki, K.; Morita, S.; Sasaki, T.; Saito, T.; Sakai, T. The extent of degeneration of cruciate ligament is associated with chondrogenic differentiation in patients with osteoarthritis of the knee. Osteoarthr. Cartil. 2012, 20, 1258–1267. [Google Scholar] [CrossRef]
- Smith, K.D.; Clegg, P.D.; Innes, J.F.; Comerford, E.J. Elastin content is high in the canine cruciate ligament and is associated with degeneration. Vet. J. 2014, 199, 169–174. [Google Scholar] [CrossRef]
- Levy, Y.D.; Hasegawa, A.; Patil, S.; Koziol, J.A.; Lotz, M.K.; D’Lima, D.D. Histopathological changes in the human posterior cruciate ligament during aging and osteoarthritis: Correlations with anterior cruciate ligament and cartilage changes. Ann. Rheum. Dis. 2013, 72, 271–277. [Google Scholar] [CrossRef]
- Immonen, J.; Siefring, C.; Sanders, L. Osteoarthritis of the Anterior Cruciate Ligament and Medial Tibial Plateau: A Cadaveric Study. Cartilage 2019, 10, 11–18. [Google Scholar] [CrossRef]
- Vasseur, P.B.; Pool, R.R.; Arnoczky, S.P.; Lau, R.E. Correlative biomechanical and histologic study of the cranial cruciate ligament in dogs. Am. J. Vet. Res. 1985, 46, 1842–1854. [Google Scholar]
- Schwarz, S.; Gogele, C.; Ondruschka, B.; Hammer, N.; Kohl, B.; Schulze-Tanzil, G. Migrating Myofibroblastic Iliotibial Band-Derived Fibroblasts Represent a Promising Cell Source for Ligament Reconstruction. Int. J. Mol. Sci. 2019, 20, 1972. [Google Scholar] [CrossRef]
- Kleinbart, F.A.; Bryk, E.; Evangelista, J.; Scott, W.N.; Vigorita, V.J. Histologic comparison of posterior cruciate ligaments from arthritic and age-matched knee specimens. J. Arthroplast. 1996, 11, 726–731. [Google Scholar] [CrossRef]
- Kwee, R.M.; Ahlawat, S.; Kompel, A.J.; Morelli, J.N.; Fayad, L.M.; Zikria, B.A.; Demehri, S. Association of mucoid degeneration of anterior cruciate ligament with knee meniscal and cartilage damage. Osteoarthr. Cartil. 2015, 23, 1543–1550. [Google Scholar] [CrossRef]
- Kwee, R.M.; Hafezi-Nejad, N.; Roemer, F.W.; Zikria, B.A.; Hunter, D.J.; Guermazi, A.; Demehri, S. Association of Mucoid Degeneration of the Anterior Cruciate Ligament at MR Imaging with Medial Tibiofemoral Osteoarthritis Progression at Radiography: Data from the Osteoarthritis Initiative. Radiology 2018, 287, 912–921. [Google Scholar] [CrossRef]
- Cha, J.R.; Lee, C.C.; Cho, S.D.; Youm, Y.S.; Jung, K.H. Symptomatic mucoid degeneration of the anterior cruciate ligament. Knee Surg. Sports Traumatol. Arthrosc. 2013, 21, 658–663. [Google Scholar] [CrossRef]
- Fernandes, J.L.; Viana, S.L.; Mendonca, J.L.; Freitas, F.M.; Bezerra, A.S.; Lima, G.A.; Matos, V.L.; Cunha, N.F.; Martins, R.R.; Freitas, R.M. Mucoid degeneration of the anterior cruciate ligament: Magnetic resonance imaging findings of an underdiagnosed entity. Acta Radiol. 2008, 49, 75–79. [Google Scholar] [CrossRef] [PubMed]
- Lintz, F.; Pujol, N.; Boisrenoult, P.; Bargoin, K.; Beaufils, P.; Dejour, D. Anterior cruciate ligament mucoid degeneration: A review of the literature and management guidelines. Knee Surg. Sports Traumatol. Arthrosc. 2011, 19, 1326–1333. [Google Scholar] [CrossRef] [PubMed]
- McMonagle, J.S.; Helms, C.A.; Garrett, W.E., Jr.; Vinson, E.N. Tram-track appearance of the posterior cruciate ligament (PCL): Correlations with mucoid degeneration, ligamentous stability, and differentiation from PCL tears. AJR Am. J. Roentgenol. 2013, 201, 394–399. [Google Scholar] [CrossRef] [PubMed]
- Jung, K.H.; Cho, S.D.; Park, K.B.; Youm, Y.S. Relation between mucoid degeneration of the anterior cruciate ligament and posterior tibial slope. Arthroscopy 2012, 28, 502–506. [Google Scholar] [CrossRef]
- Kim, G.B.; Kim, K.I.; Song, S.J.; Lee, S.H. Increased Posterior Tibial Slope After Medial Open-Wedge High Tibial Osteotomy May Result in Degenerative Changes in Anterior Cruciate Ligament. J. Arthroplast. 2019. [Google Scholar] [CrossRef] [PubMed]
- Kyllar, M.; Cizek, P. Cranial cruciate ligament structure in relation to the tibial plateau slope and intercondylar notch width in dogs. J. Vet. Sci. 2018, 19, 699–707. [Google Scholar] [CrossRef]
- Yamagami, R.; Taketomi, S.; Inui, H.; Tahara, K.; Tanaka, S. The role of medial meniscus posterior root tear and proximal tibial morphology in the development of spontaneous osteonecrosis and osteoarthritis of the knee. Knee 2017, 24, 390–395. [Google Scholar] [CrossRef]
- Hovis, K.K.; Alizai, H.; Tham, S.C.; Souza, R.B.; Nevitt, M.C.; McCulloch, C.E.; Link, T.M. Non-traumatic anterior cruciate ligament abnormalities and their relationship to osteoarthritis using morphological grading and cartilage T2 relaxation times: Data from the Osteoarthritis Initiative (OAI). Skelet. Radiol. 2012, 41, 1435–1443. [Google Scholar] [CrossRef]
- Aggarwal, A.K.; Goel, A.; Radotra, B.D. Predictors of posterior cruciate ligament degeneration in osteoarthritic knees. J. Orthop. Surg. (Hong Kong) 2013, 21, 15–18. [Google Scholar] [CrossRef]
- Allain, J.; Goutallier, D.; Voisin, M.C. Macroscopic and histological assessments of the cruciate ligaments in arthrosis of the knee. Acta Orthop. Scand. 2001, 72, 266–269. [Google Scholar] [CrossRef]
- Trompeter, A.J.; Gill, K.; Appleton, M.A.; Palmer, S.H. Predicting anterior cruciate ligament integrity in patients with osteoarthritis. Knee Surg Sports Traumatol. Arthrosc. 2009, 17, 595–599. [Google Scholar] [CrossRef]
- Barrett, J.G.; Hao, Z.; Graf, B.K.; Kaplan, L.D.; Heiner, J.P.; Muir, P. Inflammatory changes in ruptured canine cranial and human anterior cruciate ligaments. Am. J. Vet. Res. 2005, 66, 2073–2080. [Google Scholar] [CrossRef]
- Tan, A.L.; Toumi, H.; Benjamin, M.; Grainger, A.J.; Tanner, S.F.; Emery, P.; McGonagle, D. Combined high-resolution magnetic resonance imaging and histological examination to explore the role of ligaments and tendons in the phenotypic expression of early hand osteoarthritis. Ann. Rheum. Dis. 2006, 65, 1267–1272. [Google Scholar] [CrossRef]
- Slobodin, G.; Rozenbaum, M.; Boulman, N.; Rosner, I. Varied presentations of enthesopathy. Semin. Arthritis Rheum. 2007, 37, 119–126. [Google Scholar] [CrossRef]
- Slobodin, G.; Rimar, D.; Boulman, N.; Kaly, L.; Rozenbaum, M.; Rosner, I.; Odeh, M. Entheseal involvement in systemic disorders. Clin. Rheumatol. 2015, 34, 2001–2010. [Google Scholar] [CrossRef]
- Benjamin, M.; McGonagle, D. Histopathologic changes at “synovio-entheseal complexes” suggesting a novel mechanism for synovitis in osteoarthritis and spondylarthritis. Arthritis Rheum. 2007, 56, 3601–3609. [Google Scholar] [CrossRef]
- Yamamoto, M.; Takada, H.; Takayama, T.; Katsumura, S.; Sato, M.; Abe, H.; Murakami, G.; Abe, S.I. Cartilage attachment morphology of the fetal cruciate ligaments of the knee: An immunohistochemical study using human fetal specimens. Okajimas Folia Anat. Jpn. 2016, 93, 67–72. [Google Scholar] [CrossRef][Green Version]
- Binks, D.A.; Bergin, D.; Freemont, A.J.; Hodgson, R.J.; Yonenaga, T.; McGonagle, D.; Radjenovic, A. Potential role of the posterior cruciate ligament synovio-entheseal complex in joint effusion in early osteoarthritis: A magnetic resonance imaging and histological evaluation of cadaveric tissue and data from the Osteoarthritis Initiative. Osteoarthr. Cartil. 2014, 22, 1310–1317. [Google Scholar] [CrossRef]
- Abraham, A.C.; Pauly, H.M.; Donahue, T.L. Deleterious effects of osteoarthritis on the structure and function of the meniscal enthesis. Osteoarthr. Cartil. 2014, 22, 275–283. [Google Scholar] [CrossRef]
- Nakahara, H.; Hasegawa, A.; Otabe, K.; Ayabe, F.; Matsukawa, T.; Onizuka, N.; Ito, Y.; Ozaki, T.; Lotz, M.K.; Asahara, H. Transcription factor Mohawk and the pathogenesis of human anterior cruciate ligament degradation. Arthritis Rheum. 2013, 65, 2081–2089. [Google Scholar] [CrossRef]
- Zhen, G.; Wen, C.; Jia, X.; Li, Y.; Crane, J.L.; Mears, S.C.; Askin, F.B.; Frassica, F.J.; Chang, W.; Yao, J.; et al. Inhibition of TGF-beta signaling in mesenchymal stem cells of subchondral bone attenuates osteoarthritis. Nat. Med. 2013, 19, 704–712. [Google Scholar] [CrossRef]
- Xie, L.; Tintani, F.; Wang, X.; Li, F.; Zhen, G.; Qiu, T.; Wan, M.; Crane, J.; Chen, Q.; Cao, X. Systemic neutralization of TGF-beta attenuates osteoarthritis. Ann. N.Y. Acad. Sci. 2016, 1376, 53–64. [Google Scholar] [CrossRef]
- Murata, K.; Kokubun, T.; Onitsuka, K.; Oka, Y.; Kano, T.; Morishita, Y.; Ozone, K.; Kuwabara, N.; Nishimoto, J.; Isho, T.; et al. Controlling joint instability after anterior cruciate ligament transection inhibits transforming growth factor-beta-mediated osteophyte formation. Osteoarthr. Cartil. 2019, 27, 1185–1196. [Google Scholar] [CrossRef]
- Lorda-Diez, C.I.; Montero, J.A.; Choe, S.; Garcia-Porrero, J.A.; Hurle, J.M. Ligand- and stage-dependent divergent functions of BMP signaling in the differentiation of embryonic skeletogenic progenitors in vitro. J. Bone Miner. Res. 2014, 29, 735–748. [Google Scholar] [CrossRef]
- Hoffmann, A.; Pelled, G.; Turgeman, G.; Eberle, P.; Zilberman, Y.; Shinar, H.; Keinan-Adamsky, K.; Winkel, A.; Shahab, S.; Navon, G.; et al. Neotendon formation induced by manipulation of the Smad8 signalling pathway in mesenchymal stem cells. J. Clin. Invest. 2006, 116, 940–952. [Google Scholar] [CrossRef]
- Schwarting, T.; Schenk, D.; Frink, M.; Benolken, M.; Steindor, F.; Oswald, M.; Ruchholtz, S.; Lechler, P. Stimulation with bone morphogenetic protein-2 (BMP-2) enhances bone-tendon integration in vitro. Connect. Tissue Res. 2016, 57, 99–112. [Google Scholar] [CrossRef]
- Li, B.; Bai, L.; Shen, P.; Sun, Y.; Chen, Z.; Wen, Y. Identification of differentially expressed microRNAs in knee anterior cruciate ligament tissues surgically removed from patients with osteoarthritis. Int. J. Mol. Med. 2017, 40, 1105–1113. [Google Scholar] [CrossRef]
- Struglics, A.; Okroj, M.; Sward, P.; Frobell, R.; Saxne, T.; Lohmander, L.S.; Blom, A.M. The complement system is activated in synovial fluid from subjects with knee injury and from patients with osteoarthritis. Arthritis Res. Ther. 2016, 18, 223. [Google Scholar] [CrossRef]
- Busch, C.; Girke, G.; Kohl, B.; Stoll, C.; Lemke, M.; Krasnici, S.; Ertel, W.; Silawal, S.; John, T.; Schulze-Tanzil, G. Complement gene expression is regulated by pro-inflammatory cytokines and the anaphylatoxin C3a in human tenocytes. Mol. Immunol. 2013, 53, 363–373. [Google Scholar] [CrossRef]
- Girke, G.; Kohl, B.; Busch, C.; John, T.; Godkin, O.; Ertel, W.; Schulze-Tanzil, G. Tenocyte activation and regulation of complement factors in response to in vitro cell injury. Mol. Immunol. 2014, 60, 14–22. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Score Details | Processing | References |
|---|---|---|
| (A) Synovitis scores | ||
| three features of chronic synovitis (enlargement of lining cell layer, cellular density of synovial stroma, leukocyte infiltrates) (each from 0= absent to 3 = strong) Sum: 9 (0–1, no synovitis; 2–4, low-grade synovitis; 5–9, high-grade synovitis) | Histological score, human synovial membrane HE staining | [83] |
| (B) Examples for cruciate ligament degeneration scores | ||
| 1: normal, 2: abnormal (thinner than normal and sclerotic), 3: ruptured (complete disappearance of the ligament or persistence of only a few fibers) | Macroscopical score, human ACL | [110] |
| [94] | ||
| [11] | ||
| [111] | ||
| 1: normal ACL with no visible signs of disease, 2: moderately damaged or obvious disease such as the visible presence of fissuring, yet still overall an intact ACL, 3: complete rupture of the ACL | Macroscopical score, human ACL | [112] |
| 0: normal, 1: degenerative lesions of collagen fibers in less than one third of the ligament thickness, 2: degenerative lesions of collagen fibers between one third and two thirds, 3: degenerative lesions of collagen fibers in more than two thirds | Histological score, human ACL longitudinal sections HE staining | [111] |
| abnormal PCL: -abrupt zones of loose fibrous connective tissue -cystic, myxoid, and/or mucoid alterations. Cystic degeneration: acellular cysts, myxoid pattern: replacement of the normal collagen pattern by spindle and stellate-type cells. Mucinous degeneration: pools of mucinous-like tissue. Categories: normal, slight, mild, moderate, and marked | Histological score, human PCL cross section of the tibial and femoral sites, two (lateral and medial) paramedian longitudinal sections HE staining | [98] |
| 1: inflammation in the ACL, 2: mucoid degeneration, 3: chondroid metaplasia, 4: cystic changes, 5: orientation of collagen fibers changes were scored as follows: 0: no changes, 0.5: minimal changes, 1: mild changes, 2: moderate changes, 3: severe changes, sum: 0–15 | Histological score, human ACL transversely and longitudinally (median) from the proximal 1/3 of the midsubstance and femur attachment site, HE staining (+Alcian blue staining) | [89] Was slightly modified and used by others: [11,13,94] |
| 1: ligamentous structure with parallel bundles of closely packed collagen fibers, low cellularity with well spaced parallel fibroblasts showing extremely elongated nuclei: no secondary degenerative features 2: parallel arrangement of collagen and fibroblasts still detectable but some loss of compaction of collagen fibers and/or increased fibroblast nuclei. Nuclei are plumper, although still elongated. A few secondary degenerative features may be present 3: disorganization and disruption of the parallel arrangement of collagen fibers is more pronounced. Increased cellularity is often apparent and the nuclei may show loss of their bipolar nature. Secondary degenerative changes, such as calcification, ossification, cyst formation and myxoid changes may be prominent | Histological score, human ACL longitudinal sections HE staining | [112] |
| chondroid metaplasia (e.g., loss of spindle-shaped fibroblasts with transformation into ovoid or round nuclei with normal or clonal appearance; formation of perinuclear “halo” areas) and matrix degradation (e.g., loss of typical collagen fiber organization; increase in proteoglycans; woven collagen fiber, and fiber disorganization, Chondroid metaplasia and matrix degradation were graded: 1: no changes, 2: mild changes in a small area, 3: moderate, 4: severe, 5: severe diffuse changes | Histological score, canine CrCL longitudinal sections HE staining Alcian blue staining | [107,113] |
| (modified) Vasseur score grades 0–3: with several subcategories Grad 1: mild degenerative changes affecting the ligamentocytes and collagen bundles, small solitary and multiple areas of degeneration in the central region. Grade 2: large acellular areas, chondroid metaplasia, focal cell proliferation, collagen fibrils not within dense primary bundles, loss of typical bundling pattern Grade 3: a variety of severe degenerative changes (more than half of the ligament diameter involved) with tearing of deteriorated axial fibers. | Histological score, canine CrCL longitudinal sections HE staining | [93,96] |
| microfibril staining, increased staining of interfascicular and interbundle regions, ligament substance (intrabundle), extent and degree pericellular staining, each category up to 2 points, sum: 0–10 | Microfibril staining, canine CrCL and CaCL, longitudinal sections Miller’s stain: elastin, oxytalan fibers | “Miller’s score” [93] |
| calcium distribution (none, single, multiple foci, throughout), quality of deposits (none, diffuse, compact), intensity of staining/color (none, low = brown, pronounced: brown + black, intense = black), sum: 0–8 | Calcium deposition Van Kossa score, longitudinal sections Van Kossa staining | supplemental Table 1 in: [13] |
© 2019 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schulze-Tanzil, G. Intraarticular Ligament Degeneration Is Interrelated with Cartilage and Bone Destruction in Osteoarthritis. Cells 2019, 8, 990. https://doi.org/10.3390/cells8090990
Schulze-Tanzil G. Intraarticular Ligament Degeneration Is Interrelated with Cartilage and Bone Destruction in Osteoarthritis. Cells. 2019; 8(9):990. https://doi.org/10.3390/cells8090990
Chicago/Turabian StyleSchulze-Tanzil, Gundula. 2019. "Intraarticular Ligament Degeneration Is Interrelated with Cartilage and Bone Destruction in Osteoarthritis" Cells 8, no. 9: 990. https://doi.org/10.3390/cells8090990
APA StyleSchulze-Tanzil, G. (2019). Intraarticular Ligament Degeneration Is Interrelated with Cartilage and Bone Destruction in Osteoarthritis. Cells, 8(9), 990. https://doi.org/10.3390/cells8090990