Altered Levels of Toll-Like Receptors in Circulating Extracellular Vesicles in Multiple Sclerosis

 , , and
, , and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Participants, Standard Protocol Approvals, and Patient Consents
2.2. Isolation of Extracellular Vesicles
2.3. EV Array
2.4. Measurement of Markers in EVs
2.5. Statistical Analysis
3. Results
3.1. Study Cohort
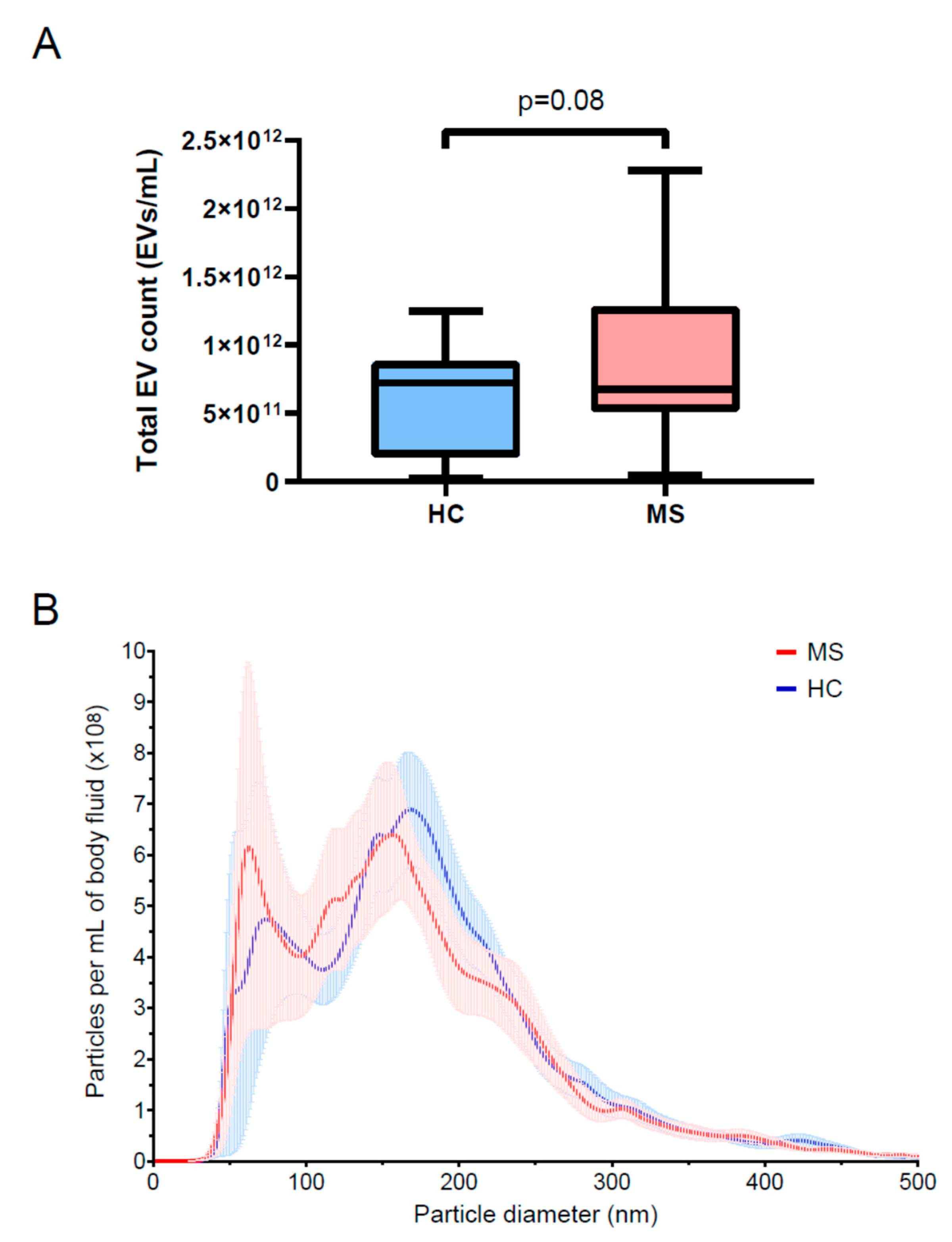
3.2. EV Array Identified Lower Levels of Multiple Surface Markers in MS Compared to Controls
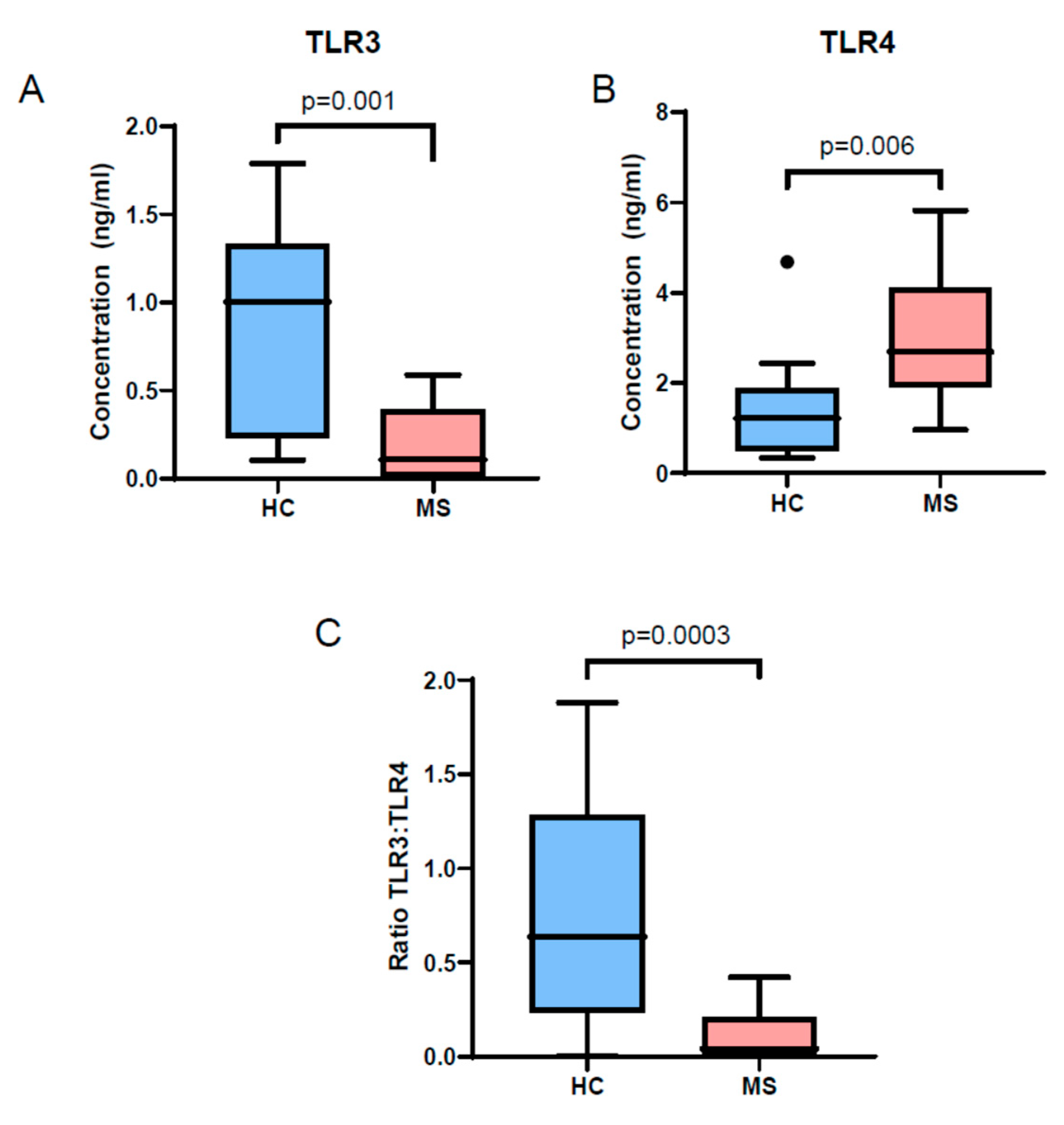
3.3. TLR3 and TLR4 Levels Are Altered in MS Patient EVs
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Reich, D.S.; Lucchinetti, C.F.; Calabresi, P.A. Multiple Sclerosis. N. Engl. J. Med. 2018, 378, 169–180. [Google Scholar] [CrossRef] [PubMed]
- Greening, D.W.; Gopal, S.K.; Xu, R.; Simpson, R.J.; Chen, W. Exosomes and their roles in immune regulation and cancer. Semin. Cell Dev. Biol. 2015, 40, 72–81. [Google Scholar] [CrossRef] [PubMed]
- Barreca, M.M.; Aliotta, E.; Geraci, F. Extracellular Vesicles in Multiple Sclerosis as Possible Biomarkers: Dream or Reality? Adv. Exp. Med. Biol. 2017, 958, 1–9. [Google Scholar] [PubMed]
- Athauda, D.; Gulyani, S.; Karnati, H.; Li, Y.; Tweedie, D.; Mustapic, M.; Chawla, S.; Chowdhury, K.; Skene, S.S.; Greig, N.H.; et al. Utility of Neuronal-Derived Exosomes to Examine Molecular Mechanisms That Affect Motor Function in Patients With Parkinson Disease: A Secondary Analysis of the Exenatide-PD Trial. JAMA Neurol. 2019, 76, 420. [Google Scholar] [CrossRef] [PubMed]
- Pulliam, L.; Sun, B.; Mustapic, M.; Chawla, S.; Kapogiannis, D. Plasma neuronal exosomes serve as biomarkers of cognitive impairment in HIV infection and Alzheimer’s disease. J. Neurovirol. 2019. [Google Scholar] [CrossRef]
- Mustapic, M.; Eitan, E.; Werner, J.K.; Berkowitz, S.T.; Lazaropoulos, M.P.; Tran, J.; Goetzl, E.J.; Kapogiannis, D. Plasma Extracellular Vesicles Enriched for Neuronal Origin: A Potential Window into Brain Pathologic Processes. Front. Neurosci. 2017, 11, 278. [Google Scholar] [CrossRef]
- Minagar, A.; Jy, W.; Jimenez, J.J.; Sheremata, W.A.; Mauro, L.M.; Mao, W.W.; Horstman, L.L.; Ahn, Y.S. Elevated plasma endothelial microparticles in multiple sclerosis. Neurology 2001, 56, 1319–1324. [Google Scholar] [CrossRef]
- Ebrahimkhani, S.; Vafaee, F.; Young, P.E.; Hur, S.S.J.; Hawke, S.; Devenney, E.; Beadnall, H.; Barnett, M.H.; Suter, C.M.; Buckland, M.E. Exosomal microRNA signatures in multiple sclerosis reflect disease status. Sci. Rep. 2017, 7, 14293. [Google Scholar] [CrossRef]
- Manna, I.; Iaccino, E.; Dattilo, V.; Barone, S.; Vecchio, E.; Mimmi, S.; Filippelli, E.; Demonte, G.; Polidoro, S.; Granata, A.; et al. Exosome-associated miRNA profile as a prognostic tool for therapy response monitoring in multiple sclerosis patients. FASEB J. 2018, 32, 4241–4246. [Google Scholar] [CrossRef]
- Geraci, F.; Ragonese, P.; Barreca, M.M.; Aliotta, E.; Mazzola, M.A.; Realmuto, S.; Vazzoler, G.; Savettieri, G.; Sconzo, G.; Salemi, G. Differences in Intercellular Communication During Clinical Relapse and Gadolinium-Enhanced MRI in Patients with Relapsing Remitting Multiple Sclerosis: A Study of the Composition of Extracellular Vesicles in Cerebrospinal Fluid. Front. Cell Neurosci. 2018, 12, 418. [Google Scholar] [CrossRef]
- Kimura, K.; Hohjoh, H.; Fukuoka, M.; Sato, W.; Oki, S.; Tomi, C.; Yamaguchi, H.; Kondo, T.; Takahashi, R.; Yamamura, T. Circulating exosomes suppress the induction of regulatory T cells via let-7i in multiple sclerosis. Nat. Commun. 2018, 9, 17. [Google Scholar] [CrossRef] [PubMed]
- Moyano, A.L.; Li, G.; Boullerne, A.I.; Feinstein, D.L.; Hartman, E.; Skias, D.; Balavanov, R.; van Breemen, R.B.; Bongarzone, E.R.; Månsson, J.-E.; et al. Sulfatides in extracellular vesicles isolated from plasma of multiple sclerosis patients. J. Neurosci. Res. 2016, 94, 1579–1587. [Google Scholar] [CrossRef] [PubMed]
- Pieragostino, D.; Cicalini, I.; Lanuti, P.; Ercolino, E.; di Ioia, M.; Zucchelli, M.; Zappacosta, R.; Miscia, S.; Marchisio, M.; Sacchetta, P.; et al. Enhanced release of acid sphingomyelinase-enriched exosomes generates a lipidomics signature in CSF of Multiple Sclerosis patients. Sci. Rep. 2018, 8, 3071. [Google Scholar] [CrossRef] [PubMed]
- Galazka, G.; Mycko, M.P.; Selmaj, I.; Raine, C.S.; Selmaj, K.W. Multiple sclerosis: Serum-derived exosomes express myelin proteins. Mult. Scler. J. 2018, 24, 449–458. [Google Scholar] [CrossRef] [PubMed]
- Jørgensen, M.M.; Bæk, R.; Varming, K. Potentials and capabilities of the Extracellular Vesicle (EV) Array. J. Extracell. Vesicles 2015, 4, 26048. [Google Scholar] [CrossRef] [PubMed]
- Bæk, R.; Jørgensen, M.M. Multiplexed Phenotyping of Small Extracellular Vesicles Using Protein Microarray (EV Array). Methods Mol. Biol. 2017, 1545, 117–127. [Google Scholar]
- Fernández-Paredes, L.; de Diego, R.P.; de Andrés, C.; Sánchez-Ramón, S. Close Encounters of the First Kind: Innate Sensors and Multiple Sclerosis. Mol. Neurobiol. 2017, 54, 101–114. [Google Scholar] [CrossRef]
- Deckx, N.; Willekens, B.; Wens, I.; Eijnde, B.O.; Goossens, H.; Van Damme, P.; Berneman, Z.N.; Cools, N. Altered molecular expression of TLR-signaling pathways affects the steady-state release of IL-12p70 and IFN-α in patients with relapsing-remitting multiple sclerosis. Innate Immun. 2016, 22, 266–273. [Google Scholar] [CrossRef]
- Okada, Y.; Ochi, H.; Fujii, C.; Hashi, Y.; Hamatani, M.; Ashida, S.; Kawamura, K.; Kusaka, H.; Matsumoto, S.; Nakagawa, M.; et al. Signaling via toll-like receptor 4 and CD40 in B cells plays a regulatory role in the pathogenesis of multiple sclerosis through interleukin-10 production. J. Autoimmun. 2018, 88, 103–113. [Google Scholar] [CrossRef]
- Crowley, T.; Fitzpatrick, J.-M.; Kuijper, T.; Cryan, J.F.; O’Toole, O.; O’Leary, O.F.; Downer, E.J. Modulation of TLR3/TLR4 inflammatory signaling by the GABAB receptor agonist baclofen in glia and immune cells: Relevance to therapeutic effects in multiple sclerosis. Front. Cell. Neurosci. 2015, 9, 284. [Google Scholar] [CrossRef]
- Kawasaki, T.; Kawai, T. Toll-like receptor signaling pathways. Front. Immunol. 2014, 5, 461. [Google Scholar] [CrossRef] [PubMed]
- Touil, T.; Fitzgerald, D.; Zhang, G.-X.; Rostami, A.; Gran, B. Cutting Edge: TLR3 stimulation suppresses experimental autoimmune encephalomyelitis by inducing endogenous IFN-beta. J. Immunol. 2006, 177, 7505–7509. [Google Scholar] [CrossRef] [PubMed]
- Fitzgerald, D.C.; Ciric, B.; Touil, T.; Harle, H.; Grammatikopolou, J.; Das Sarma, J.; Gran, B.; Zhang, G.-X.; Rostami, A. Suppressive effect of IL-27 on encephalitogenic Th17 cells and the effector phase of experimental autoimmune encephalomyelitis. J. Immunol. 2007, 179, 3268–3275. [Google Scholar] [CrossRef] [PubMed]
- Saresella, M.; Gatti, A.; Tortorella, P.; Marventano, I.; Piancone, F.; La Rosa, F.; Caputo, D.; Rovaris, M.; Biasin, M.; Clerici, M. Toll-like receptor 3 differently modulates inflammation in progressive or benign multiple sclerosis. Clin. Immunol. 2014, 150, 109–120. [Google Scholar] [CrossRef] [PubMed]
- Andersson, Å.; Covacu, R.; Sunnemark, D.; Danilov, A.I.; Dal Bianco, A.; Khademi, M.; Wallström, E.; Lobell, A.; Brundin, L.; Lassmann, H.; et al. Pivotal Advance: HMGB1 expression in active lesions of human and experimental multiple sclerosis. J. Leukoc. Biol. 2008, 84, 1248–1255. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Characteristic | RRMS | HC | |
|---|---|---|---|
| Number | 18 | 16 | |
| Age, years (mean ± SD) | 43.9 ± 10.8 | 39.4 ± 8.9 | |
| Female sex, n (%) | 13 (72.2) | 11 (68.5) | |
| Caucasian Race, n (%) | 16 (88.9) | 14 (87.5) | |
| Expanded Disability Status Scale (EDSS), median (IQR) | 2.0 (1.5) | - | |
| Disease duration, years (mean ± SD) | 9.8 ± 6.2 | - | |
| Current disease-modifying therapy | None | 9 | - |
| Glatiramer acetate | 4 | - | |
| Interferon beta | 3 | - | |
| Natalizumab | 2 | - | |
| MS Rel. Intensity (log2 Mean ± SEM) n = 3 | Control Rel. Intensity (log2 Mean ± SEM) n = 3 | Total Serum EVs (p Values) | Total Serum EVs Normalized by EV Concentration by (Nanoparticle Tracking Analysis) NTA (p Values) | |
|---|---|---|---|---|
| CD171 | 0.35 ± 0.07 | 1.97 ± 0.24 | 0.003 | 0.004 |
| TLR3 | 3.49 ± 0.87 | 7.55 ± 0.35 | 0.011 | 0.011 |
| Annexin V | 0.93 ± 0.63 | 5.52 ± 0.83 | 0.012 | 0.012 |
| CD83 | 1.46 ± 0.49 | 4.83 ± 0.64 | 0.014 | 0.016 |
| CD3 | 3.72 ± 0.51 | 6.75 ± 0.54 | 0.015 | 0.018 |
| CD45RA | 0.20 ± 0.20 | 2.94 ± 0.64 | 0.015 | 0.018 |
| LAMP-1 | 0.07 ± 0.07 | 3.03 ± 0.74 | 0.016 | 0.018 |
| a-SYN | 0.72 ± 0.43 | 3.52 ± 0.57 | 0.017 | 0.021 |
| CD45RO | 1.11 ± 0.51 | 4.31 ± 0.76 | 0.025 | 0.028 |
| CD25 | 3.12 ± 0.88 | 6.60 ± 0.59 | 0.031 | 0.032 |
| CHRM4 | 1.45 ± 0.39 | 3.40 ± 0.49 | 0.036 | 0.039 |
| CD197 | 0.67 ± 0.47 | 3.85 ± 0.91 | 0.037 | 0.04 |
| ADBB2 | 2.82 ± 0.23 | 4.38 ± 0.46 | 0.037 | 0.04 |
| TSG101 | 0.07 ± 0.07 | 2.26 ± 0.73 | 0.041 | 0.044 |
| TGFb1 | 1.93 ± 0.43 | 4.80 ± 0.85 | 0.04 | 0.046 |
| CD206 | 1.68 ± 1.07 | 5.66 ± 0.89 | 0.046 | 0.046 |
| Alix | 1.26 ± 0.64 | 4.74 ± 0.96 | 0.039 | 0.041 |
| HLA ABC | 0.16 ± 0.16 | 3.05 ± 1.38 | 0.106 | 0.11 |
| CD8a | 0.00 ± 0.00 | 1.66 ± 0.83 | 0.118 | 0.117 |
| CD56 | 0.00 ± 0.00 | 1.38 ± 0.69 | 0.118 | 0.118 |
| Fas L | 0.00 ± 0.00 | 1.89 ± 0.96 | 0.118 | 0.123 |
| TNF RII | 0.00 ± 0.00 | 2.15 ± 1.20 | 0.147 | 0.152 |
| TRAIL | 0.00 ± 0.00 | 1.90 ± 1.07 | 0.151 | 0.155 |
| CD19 | 0.00 ± 0.00 | 1.88 ± 1.10 | 0.162 | 0.167 |
| CD45 | 0.06 ± 0.06 | 2.25 ± 1.29 | 0.164 | 0.17 |
| CD14 | 0.00 ± 0.00 | 2.13 ± 1.27 | 0.169 | 0.174 |
| LAMP2 | 0.00 ± 0.00 | 1.94 ± 1.21 | 0.183 | 0.188 |
| CD106 | 0.00 ± 0.00 | 1.77 ± 1.12 | 0.187 | 0.193 |
| TNF RI | 0.30 ± 0.30 | 2.12 ± 1.15 | 0.191 | 0.199 |
| Hsp90 | 0.00 ± 0.00 | 1.86 ± 1.23 | 0.203 | 0.208 |
| CD4 | 0.14 ± 0.14 | 1.62 ± 1.02 | 0.226 | 0.233 |
| ICAM-1 | 0.00 ± 0.00 | 1.53 ± 1.21 | 0.277 | 0.281 |
| CD28 | 0.67 ± 0.67 | 2.44 ± 1.36 | 0.309 | 0.324 |
| Apo E | 0.56 ± 0.56 | 2.08 ± 1.27 | 0.337 | 0.351 |
| Flotillin-1 | 0.96 ± 0.53 | 1.91 ± 0.96 | 0.431 | 0.435 |
| APP | 0.00 ± 0.00 | 0.00 ± 0.00 | nd | nd |
| Hsp70 | 0.00 ± 0.00 | 0.00 ± 0.00 | nd | nd |
| General EV surface markers | ||||
| CD81 | 2.27 ± 0.78 | 3.95 ± 0.65 | 0.171 | 0.167 |
| CD9 | 2.11 ± 0.17 | 3.32 ± 0.71 | 0.172 | 0.187 |
| CD63 | 0.32 ± 0.32 | 1.79 ± 1.25 | 0.318 | 0.322 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bhargava, P.; Nogueras-Ortiz, C.; Chawla, S.; Bæk, R.; Jørgensen, M.M.; Kapogiannis, D. Altered Levels of Toll-Like Receptors in Circulating Extracellular Vesicles in Multiple Sclerosis. Cells 2019, 8, 1058. https://doi.org/10.3390/cells8091058
Bhargava P, Nogueras-Ortiz C, Chawla S, Bæk R, Jørgensen MM, Kapogiannis D. Altered Levels of Toll-Like Receptors in Circulating Extracellular Vesicles in Multiple Sclerosis. Cells. 2019; 8(9):1058. https://doi.org/10.3390/cells8091058
Chicago/Turabian StyleBhargava, Pavan, Carlos Nogueras-Ortiz, Sahil Chawla, Rikke Bæk, Malene Møller Jørgensen, and Dimitrios Kapogiannis. 2019. "Altered Levels of Toll-Like Receptors in Circulating Extracellular Vesicles in Multiple Sclerosis" Cells 8, no. 9: 1058. https://doi.org/10.3390/cells8091058
APA StyleBhargava, P., Nogueras-Ortiz, C., Chawla, S., Bæk, R., Jørgensen, M. M., & Kapogiannis, D. (2019). Altered Levels of Toll-Like Receptors in Circulating Extracellular Vesicles in Multiple Sclerosis. Cells, 8(9), 1058. https://doi.org/10.3390/cells8091058