Plasma Fibrinogen-Like 1 as a Potential Biomarker for Radiation-Induced Liver Injury

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Experimental Section
2.1. Animal Experiments
2.2. Liver Imaging
2.3. Irradiation
2.4. Sample Preparation
2.5. Histopathological Analysis
2.6. Western Blotting
2.7. Cell Culture and Reagents
2.8. Quantitative Real-Time Reverse Transcription-PCR (RT-qPCR)
2.9. ELISA
2.10. Statistical Analysis
3. Results
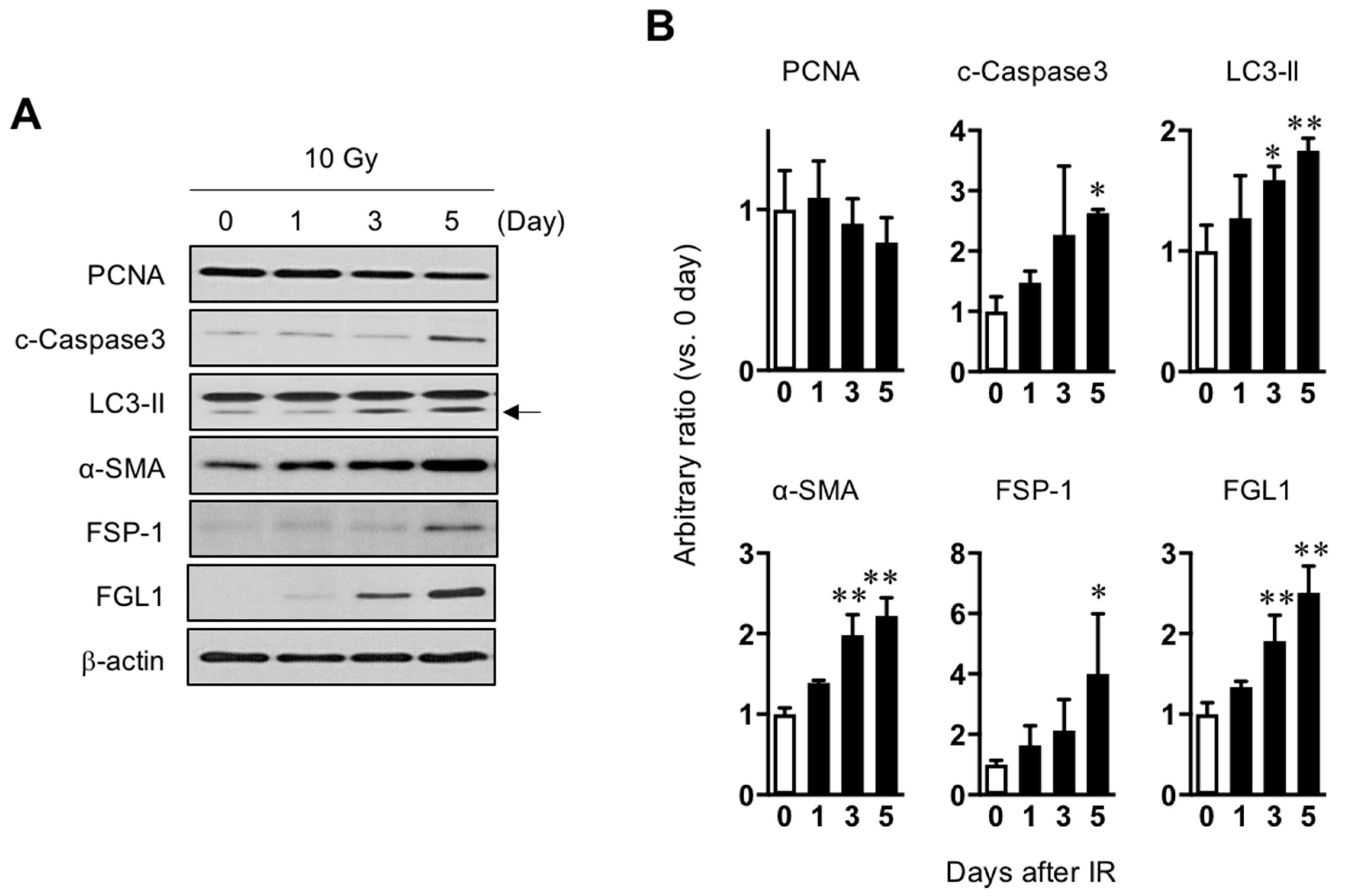
3.1. Up-Regulation of FGL1 in Mouse Liver by Irradiation
3.1.1. Association between FGL1 and Radiation-Induced Liver Injury
3.1.2. Up-Regulation of FGL1 and 8-OHdG in the Liver at the Subacute Phase after IR
3.2. Up-Regulation of FGL1 in Human Hepatocytes Responding to Radiation
3.2.1. FGL1 Expression in HHs Following IR
3.2.2. FGL1 Upregulation by Radiation or Oxidative Stress from Hydrogen Peroxidase in HHs
3.3. Plasma FGL1 as a Biomarker for Radiation Liver Injury
3.3.1. FGL1 Expression in the Liver and Plasma Following Total Body Exposure
3.3.2. FGL1 Expression in the Liver and Plasma Following Liver IR
3.3.3. Association between Plasma FGL1 and the Radiation Dose or Irradiation Field Size Relative to the Liver
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Emami, B.; Lyman, J.; Brown, A.; Coia, L.; Goitein, M.; Munzenrider, J.E.; Shank, B.; Solin, L.J.; Wesson, M. Tolerance of normal tissue to therapeutic radiation. Int. J. Radiat. Oncol. Biol. Phys. 1991, 21, 109–122. [Google Scholar] [CrossRef]
- Fleckenstein, K.; Gauter-Fleckenstein, B.; Jackson, I.L.; Rabbani, Z.; Anscher, M.; Vujaskovic, Z. Using biological markers to predict risk of radiation injury. Semin. Radiat. Oncol. 2007, 17, 89–98. [Google Scholar] [CrossRef]
- Okunieff, P.; Chen, Y.; Maguire, D.J.; Huser, A.K. Molecular markers of radiation-related normal tissue toxicity. Cancer Metastasis Rev. 2008, 27, 363–374. [Google Scholar] [CrossRef]
- Tanaka, H.; Hayashi, S.; Ohtakara, K.; Hoshi, H. Hepatic dysfunction after radiotherapy for primary gastric lymphoma. J. Radiat. Res. 2012, 54, 92–97. [Google Scholar] [CrossRef]
- Lawrence, T.S.; Robertson, J.M.; Anscher, M.S.; Jirtle, R.L.; Ensminger, W.D.; Fajardo, L.F. Hepatic toxicity resulting from cancer treatment. Int. J. Radiat. Oncol. Biol. Phys. 1995, 31, 1237–1248. [Google Scholar] [CrossRef]
- Liang, S.X.; Zhu, X.D.; Xu, Z.Y.; Zhu, J.; Zhao, J.D.; Lu, H.J.; Yang, Y.L.; Chen, L.; Wang, A.Y.; Fu, X.L.; et al. Radiation-induced liver disease in three-dimensional conformal radiation therapy for primary liver carcinoma: The risk factors and hepatic radiation tolerance. Int. J. Radiat. Oncol. Biol. Phys. 2006, 65, 426–434. [Google Scholar] [CrossRef]
- Kim, J.; Jung, Y. Radiation-induced liver disease: Current understanding and future perspectives. Exp. Mol. Med. 2017, 49, e359. [Google Scholar] [CrossRef]
- Luedde, T.; Kaplowitz, N.; Schwabe, R.F. Cell death and cell death responses in liver disease: Mechanisms and clinical relevance. Gastroenterology 2014, 147, 765–783 e4. [Google Scholar] [CrossRef]
- Hara, H.; Uchida, S.; Yoshimura, H.; Aoki, M.; Toyoda, Y.; Sakai, Y.; Morimoto, S.; Fukamachi, H.; Shiokawa, K.; Hanada, K. Isolation and characterization of a novel liver-specific gene, hepassocin, upregulated during liver regeneration. Biochim. Biophys. Acta 2000, 1492, 31–44. [Google Scholar] [CrossRef]
- Hara, H.; Yoshimura, H.; Uchida, S.; Toyoda, Y.; Aoki, M.; Sakai, Y.; Morimoto, S.; Shiokawa, K. Molecular cloning and functional expression analysis of a cDNA for human hepassocin, a liver-specific protein with hepatocyte mitogenic activity. Biochim. Biophys. Acta 2001, 1520, 45–53. [Google Scholar] [CrossRef]
- Li, C.Y.; Cao, C.Z.; Xu, W.X.; Cao, M.M.; Yang, F.; Dong, L.; Yu, M.; Zhan, Y.Q.; Gao, Y.B.; Li, W.; et al. Recombinant human hepassocin stimulates proliferation of hepatocytes in vivo and improves survival in rats with fulminant hepatic failure. Gut 2010, 59, 817–826. [Google Scholar] [CrossRef]
- Gao, M.; Zhan, Y.Q.; Yu, M.; Ge, C.H.; Li, C.Y.; Zhang, J.H.; Wang, X.H.; Ge, Z.Q.; Yang, X.M. Hepassocin activates the EGFR/ERK cascade and induces proliferation of L02 cells through the Src-dependent pathway. Cell Signal. 2014, 26, 2161–2166. [Google Scholar] [CrossRef]
- Yan, J.; Yu, Y.; Wang, N.; Chang, Y.; Ying, H.; Liu, W.; He, J.; Li, S.; Jiang, W.; Li, Y.; et al. LFIRE-1/HFREP-1, a liver-specific gene, is frequently downregulated and has growth suppressor activity in hepatocellular carcinoma. Oncogene 2004, 23, 1939–1949. [Google Scholar] [CrossRef] [Green Version]
- Cao, M.M.; Xu, W.X.; Li, C.Y.; Cao, C.Z.; Wang, Z.D.; Yao, J.W.; Yu, M.; Zhan, Y.Q.; Wang, X.H.; Tang, L.J.; et al. Hepassocin regulates cell proliferation of the human hepatic cells L02 and hepatocarcinoma cells through different mechanisms. J. Cell Biochem. 2011, 112, 2882–2890. [Google Scholar] [CrossRef]
- Demchev, V.; Malana, G.; Vangala, D.; Stoll, J.; Desai, A.; Kang, H.W.; Li, Y.; Nayeb-Hashemi, H.; Niepel, M.; Cohen, D.E.; et al. Targeted deletion of fibrinogen like protein 1 reveals a novel role in energy substrate utilization. PLoS ONE 2013, 8, e58084. [Google Scholar] [CrossRef]
- Wu, H.T.; Lu, F.H.; Ou, H.Y.; Su, Y.C.; Hung, H.C.; Wu, J.S.; Yang, Y.C.; Wu, C.L.; Chang, C.J. The role of hepassocin in the development of non-alcoholic fatty liver disease. J. Hepatol. 2013, 59, 1065–1072. [Google Scholar] [CrossRef]
- Kim, J.H.; Jenrow, K.A.; Brown, S.L. Mechanisms of radiation-induced normal tissue toxicity and implications for future clinical trials. Radiat. Oncol. J. 2014, 32, 103–115. [Google Scholar] [CrossRef] [Green Version]
- Friedman, M.A.; Volberding, P.A.; Cassidy, M.J.; Resser, K.J.; Wasserman, T.H.; Phillips, T.L. Therapy for hepatocellular cancer with intrahepatic arterial adriamycin and 5-fluorouracil combined with whole-liver irradiation: A Northern California Oncology Group Study. Cancer Treat. Rep. 1979, 63, 1885–1888. [Google Scholar]
- Stillwagon, G.B.; Order, S.E.; Guse, C.; Klein, J.L.; Leichner, P.K.; Leibel, S.A.; Fishman, E.K. 194 hepatocellular cancers treated by radiation and chemotherapy combinations: Toxicity and response: A Radiation Therapy Oncology Group Study. Int. J. Radiat. Oncol. Biol. Phys. 1989, 17, 1223–1229. [Google Scholar] [CrossRef]
- Benson, R.; Madan, R.; Kilambi, R.; Chander, S. Radiation induced liver disease: A clinical update. J. Egypt Natl Canc Inst 2016, 28, 7–11. [Google Scholar] [CrossRef] [Green Version]
- Codogno, P.; Meijer, A.J. Autophagy and signaling: Their role in cell survival and cell death. Cell Death Differ. 2005, 12, 1509–1518. [Google Scholar] [CrossRef]
- Hofer, S.; Brenner, T.; Bopp, C.; Steppan, J.; Lichtenstern, C.; Weitz, J.; Bruckner, T.; Martin, E.; Hoffmann, U.; Weigand, M.A. Cell death serum biomarkers are early predictors for survival in severe septic patients with hepatic dysfunction. Crit. Care 2009, 13, R93. [Google Scholar] [CrossRef]
- Xagorari, A.; Siotou, E.; Yiangou, M.; Tsolaki, E.; Bougiouklis, D.; Sakkas, L.; Fassas, A.; Anagnostopoulos, A. Protective effect of mesenchymal stem cell-conditioned medium on hepatic cell apoptosis after acute liver injury. Int. J. Clin. Exp. Pathol. 2013, 6, 831–840. [Google Scholar]
- Lorente, L.; Martin, M.M.; González-Rivero, A.F.; Ferreres, J.; Solé-Violan, J.; Labarta, L.; Díaz, C.; Jiménez, A.; Borreguero-León, J.M. Serum levels of caspase-cleaved cytokeratin-18 and mortality are associated in severe septic patients: Pilot study. PLoS ONE 2014, 9, e109618. [Google Scholar] [CrossRef]
- Ausch, C.; Buxhofer-Ausch, V.; Olszewski, U.; Hinterberger, W.; Ogris, E.; Schiessel, R.; Hamilton, G. Caspase-cleaved cytokeratin 18 fragment (M30) as marker of postoperative residual tumor load in colon cancer patients. Eur. J. Surg. Oncol. 2009, 35, 1164–1168. [Google Scholar] [CrossRef] [Green Version]
- Sultan, S.; Cameron, S.; Ahmad, S.; Malik, I.A.; Schultze, F.C.; Hielscher, R.; Rave-Fränk, M.; Hess, C.F.; Ramadori, G.; Christiansen, H. Serum Lipocalin2 is a potential biomarker of liver irradiation damage. Liver Int. 2013, 33, 459–468. [Google Scholar] [CrossRef]
- Perrotti, A.; Miltgen, G.; Chevet-Noel, A.; Durst, C.; Vernerey, D.; Bardonnet, K.; Davani, S.; Chocron, S. Neutrophil gelatinase-associated lipocalin as early predictor of acute kidney injury after cardiac surgery in adults with chronic kidney failure. Ann. Thorac. Surg. 2015, 99, 864–869. [Google Scholar] [CrossRef]
- Jin, H.; Kang, G.Y.; Jeon, S.; Kim, J.M.; Park, Y.N.; Cho, J.; Lee, Y.S. Identification of molecular signatures involved in radiation-induced lung fibrosis. J. Mol. Med.(Berl) 2019, 97, 37–47. [Google Scholar] [CrossRef]
- Potters, L.; Steinberg, M.; Rose, C.; Timmerman, R.; Ryu, S.; Hevezi, J.M.; Welsh, J.; Mehta, M.; Larson, D.A.; Janjan, N.A. American Society for Therapeutic Radiology and Oncology and American College of Radiology practice guideline for the performance of stereotactic body radiation therapy. Int. J. Radiat. Oncol. Biol. Phys. 2004, 60, 1026–1032. [Google Scholar] [CrossRef]
- Khozouz, R.F.; Huq, S.Z.; Perry, M.C. Radiation-induced liver disease. J. Clin. Oncol. 2008, 26, 4844–4845. [Google Scholar] [CrossRef]
- Pan, C.C.; Kavanagh, B.D.; Dawson, L.A.; Li, X.A.; Das, S.K.; Miften, M.; Ten Haken, R.K. Radiation-associated liver injury. Int. J. Radiat. Oncol. Biol. Phys. 2010, 76, S94–S100. [Google Scholar] [CrossRef]
- Guha, C.; Kavanagh, B.D. Hepatic radiation toxicity: Avoidance and amelioration. Semin. Radiat. Oncol. 2011, 21, 256–263. [Google Scholar] [CrossRef]
- Bae, S.H.; Kim, M.S.; Jang, W.I.; Cho, C.K.; Yoo, H.J.; Kim, K.B.; Han, C.J.; Park, S.C.; Lee, D.H. Low Hepatic Toxicity in Primary and Metastatic Liver Cancers after Stereotactic Ablative Radiotherapy Using 3 Fractions. J. Korean Med. Sci. 2015, 30, 1055–1061. [Google Scholar] [CrossRef] [Green Version]







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, N.-K.; Jung, M.G.; Jeong, Y.J.; Son, Y.; Han, S.C.; Park, S.; Lim, Y.-B.; Lee, Y.-J.; Kim, S.-H.; Park, S.C.; et al. Plasma Fibrinogen-Like 1 as a Potential Biomarker for Radiation-Induced Liver Injury. Cells 2019, 8, 1042. https://doi.org/10.3390/cells8091042
Han N-K, Jung MG, Jeong YJ, Son Y, Han SC, Park S, Lim Y-B, Lee Y-J, Kim S-H, Park SC, et al. Plasma Fibrinogen-Like 1 as a Potential Biomarker for Radiation-Induced Liver Injury. Cells. 2019; 8(9):1042. https://doi.org/10.3390/cells8091042
Chicago/Turabian StyleHan, Na-Kyung, Myung Gu Jung, Ye Ji Jeong, Yeonghoon Son, Su Chul Han, Seungwoo Park, Young-Bin Lim, Yoon-Jin Lee, Sung-Ho Kim, Su Cheol Park, and et al. 2019. "Plasma Fibrinogen-Like 1 as a Potential Biomarker for Radiation-Induced Liver Injury" Cells 8, no. 9: 1042. https://doi.org/10.3390/cells8091042
APA StyleHan, N.-K., Jung, M. G., Jeong, Y. J., Son, Y., Han, S. C., Park, S., Lim, Y.-B., Lee, Y.-J., Kim, S.-H., Park, S. C., & Lee, H.-J. (2019). Plasma Fibrinogen-Like 1 as a Potential Biomarker for Radiation-Induced Liver Injury. Cells, 8(9), 1042. https://doi.org/10.3390/cells8091042