Ameliorating the Fibrotic Remodeling of the Heart through Direct Cardiac Reprogramming


Department of Medicine, Heart and Vascular Research Center, The MetroHealth System, Case Western Reserve University, Cleveland, OH 44106, USA
*
Author to whom correspondence should be addressed.
Cells 2019, 8(7), 679; https://doi.org/10.3390/cells8070679
Submission received: 5 May 2019
/
Revised: 21 June 2019
/
Accepted: 23 June 2019
/
Published: 4 July 2019
(This article belongs to the Special Issue Direct Cell Reprogramming: From Basic Science to Translational Medicine)
Abstract
:Coronary artery disease is the most common form of cardiovascular diseases, resulting in the loss of cardiomyocytes (CM) at the site of ischemic injury. To compensate for the loss of CMs, cardiac fibroblasts quickly respond to injury and initiate cardiac remodeling in an injured heart. In the remodeling process, cardiac fibroblasts proliferate and differentiate into myofibroblasts, which secrete extracellular matrix to support the intact structure of the heart, and eventually differentiate into matrifibrocytes to form chronic scar tissue. Discovery of direct cardiac reprogramming offers a promising therapeutic strategy to prevent/attenuate this pathologic remodeling and replace the cardiac fibrotic scar with myocardium in situ. Since the first discovery in 2010, many progresses have been made to improve the efficiency and efficacy of reprogramming by understanding the mechanisms and signaling pathways that are activated during direct cardiac reprogramming. Here, we overview the development and recent progresses of direct cardiac reprogramming and discuss future directions in order to translate this promising technology into an effective therapeutic paradigm to reverse cardiac pathological remodeling in an injured heart.
1. Introduction
Mortality by cardiovascular diseases accounts for 31.5% of all deaths worldwide [1]. The most common form of heart disease is coronary artery disease where functional cardiomyocytes (CMs) die out in the ischemic area of the heart and are replaced by a fibrotic scar, which leads to the dysfunction of the heart and eventually heart failure. Since adult CMs possess very limited to no self-regenerative capability, heart transplantation is still the final solution for patients with end-stage heart failure, however, this remains a limited option for most patients due to the shortage of donor organs. Therefore, developing new strategies of cellular therapies offers more accessible options for a broader group of coronary heart patients and prevents a diseased heart from end-stage failure. Currently, there are five major cellular therapy strategies that have been actively investigated in the past few decades for cardiac regenerative medicine: (1) Transplantation of autologous adult stem cells [2,3,4,5], (2) transplantation of embryonic stem cell (ESC)- or induced pluripotent stem cell (iPSC)-derived CMs [6,7,8], (3) activation of endogenous progenitors [9,10,11,12], (4) cell-cycle reentry of adult CMs [13,14,15,16], and (5) direct cell fate reprogramming [17,18,19,20,21]. Each strategy has shown promising benefits but are also facing different technical challenges [22,23]. Many research progresses of direct cardiac reprogramming have been discussed in recently review articles [24,25,26]; in this article, we reviewed the newest discoveries of direct cardiac reprogramming, including cell cycle regulation, chemokine signaling, inflammatory immune signaling, and single-cell RNA-seq findings during reprogramming of induced cardiomyocyte-like cells (iCM), and incorporated a new understanding of matrifibrocytes during cardiac fibrotic remodeling to discuss future directions of translating this promising technology into clinical applications.
2. Pathological Remodeling of the Heart
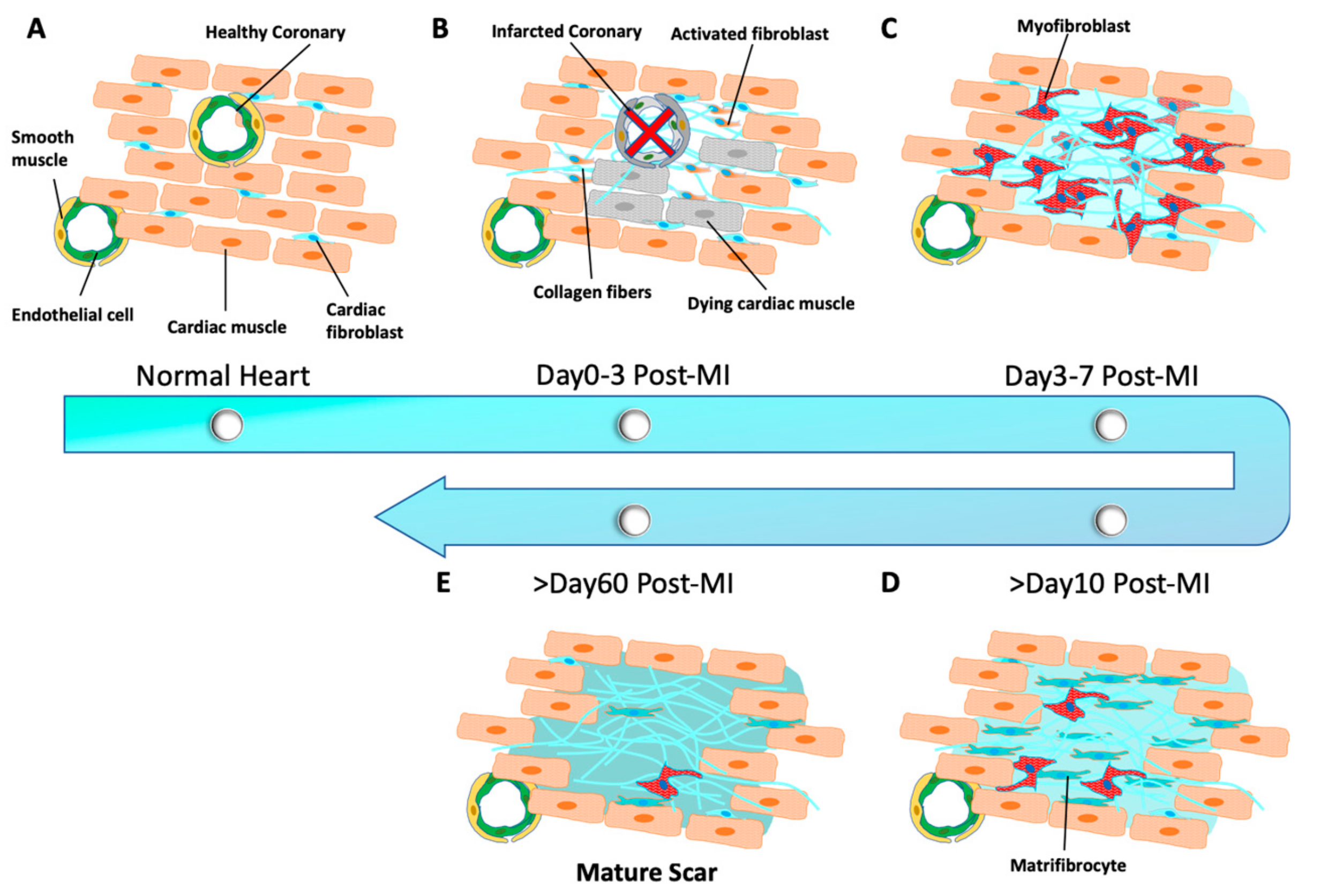
During the development of coronary artery diseases, such as myocardial infarction (MI), the heart tissue undergoes a pathological remodeling where necrotic myocardium is replaced by non-myocyte cells to preserve functional and structural integrity of the heart. Remodeling is characterized by excessive accumulation of collagen-based extracellular matrix (ECM) majorly from resident cardiac fibroblasts [27]. Previous studies [28,29] have suggested that fibroblasts constitute the majority of non-myocytes in the heart, which was considered recently to be overestimated in the adult murine and healthy human heart [30]. In the normal healthy heart (Figure 1A), cardiac fibroblasts remain quiescent and play an essential role in the maintenance of mechanical, structural, and electrophysiological functions of the heart [31,32]. However, in an infarcted myocardium, quiescent fibroblasts are activated to secrete various pro-inflammatory factors [33] and quickly re-enter the cell cycle (Figure 1B). Following a massive proliferation, cardiac fibroblasts differentiate into contractile myofibroblasts that are characterized by the expression of α-smooth muscle actin (αSMA) within a week of a MI [33,34,35] (Figure 1C). Toward the end of remodeling, myofibroblasts exit the cell-cycle and provide a stronger contractile support to the damaged heart by secreting much more ECM than fibroblasts [36,37,38]. In most tissues, myofibroblasts undergo apoptosis when the scar is fully matured and the healing process is completed [39,40]. However, in the heart, a significant number of quiescent myofibroblasts persist in the formed mature scar for long time (>60 days post-MI) to mechanically support and protect the heart against any further damage [41]. Very recently, Fu et al. identified a more specialized type of cell, termed matrifibrocytes, with expression of new marker genes, such as bone-cartilage markers Chad and Comp. They found that matrifibrocytes were further differentiated from myofibroblasts and persisted in the chronic myocardial scar [42] (Figure 1D). Their lineage-tracing studies has updated our understanding of the pathological remodeling in an infarcted heart. In brief (Figure 1), quiescent cardiac fibroblasts are activated upon injury and reach a maximum proliferation rate within Day 2–4 post-MI, followed by αSMA+ myofibroblast differentiation and proliferation by Day 3–7 post-MI; from Day 10 post-MI, myofibroblasts lose proliferative ability and αSMA expression and then further differentiate into matrifibrocytes and finally form a mature scar within 60 days [39]. However, the role of matrifibrocytes in heart remodeling and regeneration has yet to be understood.
3. Development of Direct Cardiac Reprogramming
3.1. Direct Reprogramming of Mouse Fibroblasts into iCMs In Vitro
Since lost CMs in an injured heart are replaced by cardiac fibroblasts, it will be a promising therapy for cardiac regenerative medicine if proliferated cardiac fibroblasts can be transdifferentiated into functional CMs. Transdifferentiation was initially reported in the early 1990s whereby a single transcription factor, MyoD, was sufficient to convert fibroblasts and epithelial cells into skeletal muscle cells [43]. After decades of efforts, the discovery of iPSCs [44,45] demonstrated that, rather than a single transcription factor, a combination of several transcription factors might be required to directly convert a terminally-differentiated cell type into another. Indeed, in 2010, Ieda et al. [17] cultured cardiac fibroblasts, isolated from αMHC-GFP transgenic mice, and successfully identified a minimal combination of three transcription factors (GMT: Gata4, Mef2c, and Tbx5) that could directly convert cardiac fibroblasts into induced cardiomyocyte-like cells (iCMs) without undergoing an intermediate pluripotency or progenitor state. Reprogrammed αMHC-GFP+ iCMs expressed a group of cardiac genes, e.g., Myh6, Actc1, Actn2, etc., and showed assembled sarcomere structures. Spontaneous calcium oscillations/transients were observed in many iCMs two weeks after reprogramming induction, but action potential and cell contraction were developed only in a very small population (0.01–0.1%) of reprogrammed cells.
Soon after the first study by Ieda et al. [17], Song et al. [21] included Hand2 in the GMT cocktail (GHMT), which yielded more αMHC-GFP/cardiac Troponin-T (cTnT) double-positive iCMs in adult murine tail-tip fibroblasts and cardiac fibroblasts than GMT only. Addis et al. [46] used a different reporter system in which a genetically encoded calcium indicator GCaMP was driven by cTnT promoter. They found that Nkx2.5 could significantly enhance the efficiency of GHMT to reprogram more functional iCMs with spontaneous calcium oscillations and beating. Protze et al. [47] screened triple combinations of a group of candidate transcription factors and found that an optimal combination of Mef2c, Tbx5, and Myocd could activate expression of a broader spectrum of cardiac genes than GMT did. Hirai et al. [48] fused MyoD domain to Mef2c and found that MyoD-Mef2c fused together with three other wild-type genes (Gata4, Hand2 and Tbx5) could yield 15-fold more beating iCMs than wild-type GHMT. In contrast to the transcription-factor-based approach, Jayawardena et al. [19] investigated various combinations of cardiac enriched microRNAs (miRs) for iCM induction and found that a combination of muscle-specific miRs (miR combo: miR1, miR133, miR208, and miR409) could induce iCMs from mouse cardiac fibroblasts. Moreover, JAK inhibitor improved the reprogramming capability of the miR combo. Consistently, Muraoka et al. [49] found that miR133 suppressed Snai1, a master regulator of epithelial-to-mesenchymal transition improved GMT-mediated iCM reprogramming, including a higher yield and shortened induction time of beating iCMs. Importantly, the reprogramming trajectory is acquired within 48 h of GMT virus infection into mouse cardiac fibroblasts [50], and a sufficient expression of all reprogramming factors in individual fibroblasts is critical to overcome the initial barrier for a successful iCM reprogramming [50,51].
3.2. In Situ Reprogramming of iCMs in The Heart
After the success of in vitro iCM-reprogramming, converting resident cardiac fibroblasts into functional CMs in situ is critical to translating this promising approach into a practical therapeutic paradigm for cardiac regenerative medicine. The first success of in vivo reprogramming was reported by Qian et al. [20]. They found that a local injection of GMT retroviruses into the ischemic region of the heart, right after coronary artery ligation, could reprogram new iCMs with improved ejection-fraction of the heart and decreased scar size. They utilized multiple lineage-tracing animals, including periostin-Cre and fibroblasts-specific protein 1 (Fsp1)-Cre mice, and demonstrated that in vivo reprogrammed iCMs predominantly originated from resident cardiac fibroblasts. Importantly, in vivo reprogrammed iCMs were highly similar to native CMs with a well-formed sarcomere and functionally coupled well with endogenous CMs. Meanwhile, Song et al. [21] also showed the success of in vivo iCM-reprogramming with GHMT factors. Most importantly, both studies have shown that in vivo generated iCMs improve the ejection-fraction and decrease scar size four weeks after MI. Additional Thymosin-β4 could improve the outcome of GMT-reprogramming and further enhance the functional recovery of the infarcted heart through improved neovascularization [52]. The success of in vivo reprogramming has also been reproduced with a miR combo (miR-1, miR-133, miR-208, miR-499), which decreased scar size and improved heart function in the mouse MI model [19,53]. Inagawa et al. [18] found that GMT viruses majorly infected fibroblasts and other non-myocyte cells within the scar, but only 3% of infected cells expressed αMHC-GFP and formed striated sarcomeres. A single polycistronic vector of three factors (GMT) with self-cleaving peptides could increase the in vivo-reprogramming yield of αMHC-GFP+/α-Actinin+ cells, of which 30% showed well-formed sarcomere structures. Consistently, Wang et al. [54] studied different orders of three factors in single polycistronic vector and found that a polycistronic vector of MGT, in which Mef2c is expressed at relatively higher levels than Gata4 and Tbx5, could improve the efficiency and the quality of in vitro-reprogrammed iCMs. However, MGT construct only increased the number but not the quality of in vivo-reprogrammed iCMs [55], suggesting that the in vivo environment of the heart has a more significant role in iCM quality than MGT itself. Nonetheless, single-cell transcriptome analysis revalidated that Mef2c is expressed relatively higher than Gata4 and Tbx5 in successfully reprogramming iCMs [56].
3.3. Direct Cardiac Reprogramming of Human Fibroblasts
Another critical step in translating direct cardiac reprogramming into a therapeutic approach is to achieve reprogramming of human cells with optimized reprogramming cocktails. Several research teams, including ours, has shown that neither GMT nor GHMT were sufficient to reprogram human fibroblasts. Wada et al. [57] found that the addition of Myocd and Mesp1 to GMT (GMTMM) could convert human cardiac and dermal fibroblasts into iCMs, in which the expression of cardiac genes were increased and fibroblast genes were decreased. In our laboratory, we re-performed a screening of 21 cardiac enriched transcription factors and found that Esrrg and Mesp1 together with GMT (5F) was sufficient to induce αMHC and cTnT double-positive iCMs from human fibroblasts [58]. Moreover, addition of Myocd and ZFPM2 to 5F combination (7F) further increased the yield and improved the quality of reprogrammed iCMs. Global gene expression in iCMs shifted from fibroblasts to cardiomyocyte-like state. Calcium transients and a depolarized resting membrane potential were observed in both 5F- and 7F-reprogrammed cells; however, spontaneous contraction was not observed. Our single cell qPCR studies demonstrated that both Hand2 and miR1 could further improve the quality of 7F-reprogrammed iCMs, indicated by more activated cardiac genes and silenced fibroblast genes [59], although the yield of iCMs was not improved. Meanwhile, TGF-β and Wnt signaling inhibitors could improve the 7F-iCM reprogramming and induced more spontaneous calcium oscillations in 7F-iCMs [60]. Nam et al. [61] started from GHMT and screened additional transcription factors for human iCM reprogramming. They found that Myocd together with GHMT could activate more cTnT expression in human fibroblasts. They further included miR1 and miR133 and identified a reprogramming cocktail of six factors (Gata4, Hand2, Tbx5, Myocd, miR1, and miR133) that could directly convert human fibroblasts into iCMs. Spontaneous calcium oscillation was observed in 8-week reprogrammed iCMs and spontaneous contraction was observed in a very small portion of iCMs 11 weeks after reprogramming.
3.4. Direct Reprogramming to Multipotent Cardiac Progenitors
In addition to iCM reprogramming, reprogramming fibroblasts into expandable cardiac progenitor cells has also been studied. A combination of five factors (Mesp1, Tbx5, Gata4, Nkx2.5, and Baf60c) in combination with activation of canonical Wnt and JAK/STAT pathways could reprogram adult mouse cardiac, lung, and tail tip fibroblasts into expandable progenitor cells, termed induced cardiac progenitor cells (iCPCs) [62]. iCPCs display cardiac-mesoderm-restricted multipotency with the ability to be differentiated into cardiac myocytes, smooth muscle, and endothelial cells. Considering the chemically defined condition that can capture and expand CPCs in vitro [63], reprogramming of iCPCs provides an alternative cell source for patient-specific autologous cell transplantation therapy.
4. Mechanistical Understanding of Direct Cardiac Reprogramming
With the goal of translating reprogramming into a therapeutic paradigm, scientists around the world have been enthusiastically investigating the reprogramming mechanism and developing new approaches/tools to improving the efficiency of direct cardiac reprogramming.
4.1. Activation of Signaling Pathways During Reprogramming
It has been found that different signaling pathways are involved in and directly regulate iCM reprogramming (Figure 2A and Table 1). Inhibition of transforming growth factor beta (TGF-β) by SB431542 compound [64,65] could enhance GHMT-reprogramming and yield more iCMs from mouse embryonic fibroblasts (MEFs) and adult cardiac fibroblasts. Inhibition of Rho-associated kinase (ROCK) pathway increased the conversion rate of iCM-reprogramming in MEFs, tail tip fibroblasts, and cardiac fibroblasts [65], probably by the prevention of cell apoptosis that happens in some newly-reprogrammed iCMs [66]. Inhibition of Notch signaling increased the binding of Mef2c to the promoter regions of cardiac structural genes and subsequently enhanced cardiac reprogramming [67]. Furthermore, a combination of Wnt-inhibitor and TGF-β inhibitor could significantly augment GMT reprogramming and accelerate the conversion progression of beating iCMs [60]. In addition, activation of IGF1/PI3K/Akt1 signal pathway, in which mTORC1 and Foxo3a act as downstream mediators, enhanced GHMT-mediated reprogramming [68]. The reprogramming benefit of activation of p38 MAP kinase and PI3K/Akt pathways was consistently observed with a serum-free culture condition of FGF2, FGF10, and VEGF (FFV) [69]. On the other hand, suppression of inflammatory signaling has been shown to enhance direct cardiac reprogramming. For example; the zinc finger transcription factor 281 (ZNF281) enhanced cardiac reprogramming in part by repressing the inflammatory markers in adult mouse tail-tip fibroblasts [70]. Recently, inhibition of C-C chemokine signaling in MEFs or neonatal cardiac fibroblasts enhanced cardiac reprogramming efficiency, emphasizing a requirement of immune-response suppression for iCM-reprogramming progression [71]. Consistently, inhibition of inflammatory immune signaling through suppression of cyclooxygenase-2 highly improved the yield and quality of reprogrammed iCMs in neonatal and adult mouse fibroblasts [72], which was not observed in MEFs.
4.2. Epigenetic Barriers of Reprogramming
Direct reprogramming of somatic cells into other type of cells, including iPSCs [73,74], neurons [75,76] and CMs, requires overcoming epigenetic barriers and the opening of chromatin structures on critical genes related to specific cell-fate identity. Indeed, the enrichment of trimethylated histone H3 of lysine 4 (H3K4me3), a mark of actively-transcribed genes, was increased and trimethylated histone H3 of lysine 27 (H3K27me3), a mark of inactive genes, was decreased in cardiac gene promoter regions as early as Day 3 of cardiac reprogramming [77]. Interestingly, the most effective timing of H3K27me3 diminishment to improve iCM-reprogramming was observed from Day 1 to Day 4 after virus infection, while diminishment of H3K9me2, another mark of inactivated genes, was most effective in enhancing the reprogramming from Day 3 to Day 7 [78], suggesting that the timing of epigenetic regulation is critical for a successful direct cardiac reprogramming. Inhibition of H3K27 methyltransferases or of polycomb repressive complex 2 (PRC2) could induce cardiac gene expression in fibroblasts; while inhibition of H3K27 demethylases blocked the induction of cardiac gene expression in miR-combo-mediated reprogramming [79]. Consistently, suppression of Bmi1, one component of PRC1, enhanced open chromatin states of cardiac genes, especially of Gata4, which improved cardiac reprogramming and eliminated the need for exogenous Gata4 [80]. Nonetheless, ZNF281, a transcriptional regulator, co-occupied cardiac gene enhancers with Gata4 and promoted GHMT(+Akt1)-mediated cardiac reprogramming in adult mouse fibroblasts through synergistic activation of diverse set of cardiac genes [70]. Very recently, it was found that Mef2c orchestrates chromatin accessibility of Gata4 and Tbx5 factors by recruiting them to Mef2 binding sites [81]. On the other hand, ectopic Gata4 was shown to be enriched at low levels on additional target gene loci and co-expression with another pioneer factor, Foxa2, increased enrichment on sampling genomic sites [82].
4.3. Cell-Cycle Regulation During Direct Cardiac Reprogramming
Although cardiac fibroblasts are quiescent in the healthy heart [31]; fibroblasts in an injured heart or in culture are quickly activated to re-enter the cell cycle. It is known that each cell-cycle phase constitutes a chain of interconnected events with a dynamic fluctuation of epigenetic chromatin modifications, including genomic DNA methylation and histone modifications [83,84]; therefore, it is important to understand how cell cycle is regulated in reprogrammed cells during direct cardiac reprogramming. Time-lapse imaging from Day 2 to Day 4 post-GMT-infection showed directly that nearly 40% of αMHC-GFP+ iCMs reprogrammed from MEFs were still active in the cell cycle at early stages of reprogramming [66], but αMHC-GFP+ iCMs gradually exited the cell cycle along with the progression of reprogramming. Interestingly, time-lapse imaging of early-stage reprogrammed cells suggested that iCM-reprogramming was mostly initiated at late-G1- or S-phase [66], indicating that a particular phase of the cell cycle might offer a more optimal epigenetic status for reprogramming induction. Indeed, depletion of Foxa2 in human foreskin fibroblasts resulted in a decreased demethylation on Foxa2 target genes in S-phase, but not in G1-arrested cells [82], suggesting that a loosened structure of S-phase chromosomes assists transcription factors access to their targeted genes. In our study, cell-cycle synchronization at S-phase facilitated cell-cycle exit and accelerated iCM reprogramming with enhanced expression of cardiac genes [66]. Other studies have also demonstrated that cell-cycle exit is critical for the maturation of reprogrammed iCMs from MEFs and mouse cardiac fibroblasts [66,85], while overexpression of large T antigen, which endows cardiac fibroblasts a constitutive proliferation ability, inhibited reprogramming induction [56]. Recently, by single-cell analysis techniques, critical function of cell-cycle exit is validated in iCM reprogramming of human cardiac fibroblasts [86]. In summary, cell-cycle exit, which is a developmentally required process of CM maturation in mammalian hearts [87], is an important and necessary event for direct cardiac reprogramming.
4.4. Modification of Extracellular Matrix
Extracellular matrix (ECM) has a significant impact on the function of cardiomyocytes. Matrices that mimic mechanical stiffness of the developing heart is optimal for promoting actomyosin striation and for transmitting contraction of cardiomyocytes to the matrix; neither harder matrices, which mimic the ECM of infarcted heart, nor softer matrices are good for developing well organized sarcomeres in cardiomyocytes [88,89]. Manipulating the stiffness of culture matrix could improve the functional maturation of human iPSC-derived cardiomyocytes [90]. Indeed, ECM also plays an important role in cardiac reprogramming (Table 1). Although adjusting the substrate stiffness alone didn’t improve reprogramming yield, a micro-grooved substrate could enhance reprogramming efficiency and yielded more mature iCMs with improved quality, including the higher expression of cardiac genes and more beating cells, through an increased nuclear localization of Mkl1, a mechanosensitive transcription factor [91]. Li et al. [92] encapsulated reprogrammed fibroblasts in three-dimensional (3D) hydrogels and found that the 3D tissue bundle environment increased the expression of matrix metalloproteinases (MMPs) and enhanced direct cardiac reprogramming with increased yield and improved quality of iCMs. These findings demonstrate the importance of a proper extracellular-matrix for reprogramming, which is one of the mysterious in vivo environmental factors in the native heart. Although these studies partly mimic the in vivo environment of a scar, an actual scar exhibits different levels of stiffness [93] with different amounts of collagen deposition [94] at various time points after infarction, which may affect reprogramming quality differently. Therefore, a specific period of post-MI time window may have an optimal stiffness environment to maximize the reprogramming efficiency in vivo. Indeed, infarcted myocardium progressively increased stiffness within a month and the highest level of regeneration was achieved at Day 7–14 post-MI, which provided an optimal stiffness for the endothelial progenitor lineage commitment of transplanted bone-marrow mononuclear cells [93]. Therefore, future reprogramming studies should pay greater attention to the mechanical microenvironments in cell cultures and in vivo studies.
4.5. Non-Genomic Integration Methods for Direct Cardiac Reprogramming
It is a clinical concern that retroviruses/lentivirus integrate into the genome of host cells and might insert random mutations [21,95,96,97,98,99]. Therefore, non-genomic integration methods have also been investigated actively for a safer approach for iCM reprogramming. In addition to adeno and adeno-associated virus that were studied in reprogramming [98,100], Miyamoto et al. [101] utilized Sendai virus (SeV) and developed a non-integrating, polycistronic GMT vector system (SeV-GMT) that successfully reprogrammed cardiac fibroblasts into iCMs in vitro with higher reprogramming efficiency. In vivo delivery of SeV-GMT yielded a better functional recovery of the heart with ~3.5 times smaller scar size than retroviral-GMT did. Chang et al. [102] utilized cationic gold nanoparticles (AuNPs) to deliver GMT transcription factors into MEFs. AuNPs allowed integration-free transfection of reprogramming factors with no measurable cytotoxicity on cells. Fibroblasts were much more efficiently reprogrammed into iCMs and started beating with AuNP-GMT compared to regular GMT transfection. Moreover, in vivo delivery of AuNP-GMT in a mouse model improved heart function by measurably decreasing scar size and fibrotic area two weeks post-MI. Alternative to gene therapy, a cocktail of small-molecule chemical compounds, consisted of CHIR99021, RepSox, Forskolin, VPA, Parnate, and TTNPB (CRFVPT), had been identified to successfully reprogram MEFs and tail-tip fibroblasts into chemical-induced cardiomyocyte-like cells (CiCMs) [103]. The induction of CiCMs by CRFVPT passes through a cardiac progenitor stage with upregulated expression of Sca-1, Wt1, Flk1, Abcg2, and Mesp1 in the early stages of reprogramming, but not a pluripotent stage. Importantly, in vivo delivery of CRFVPT plus Rolipram into Fsp1-Cre:R26RtdTomato mice successfully generated tdTomato+ CiCMs in the heart and reduced the formation of fibrotic scar tissues in MI hearts with functional improvement of the heart [104]. Similarly, CiCMs could be also reprogrammed from human foreskin fibroblasts by a combination of nine chemical compounds (CHIR99021, A83-01, BIX01294, AS8351, SC1, Y27632, OAC2, SU16F, and JNJ10198409) [105]. Human CiCMs also initially passed through a progenitor stage before the final conversion into functional iCMs, suggesting a different mechanism of CiCM-reprogramming from gene-mediated cardiac reprogramming. All these studies with non-integrated approaches will facilitate the translation of direct cardiac reprogramming into an efficient and safer therapy for future clinical applications.
5. Perspective to Translate Direct Cardiac Reprogramming
Despite the enormous progress, direct cardiac reprogramming remains an immature technology and a better mechanistic understanding of iCM reprogramming, especially in vivo reprogramming, is required for future clinical applications.
5.1. Understanding the Mystery of In Vivo Reprogramming
Compared to in vitro reprogramming in petri dishes, in vivo reprogramming generated a higher yield and better quality of iCMs in mouse heart with acute MI [18,20,21], suggesting that the in vivo environment of the heart after acute MI appears to overcome more cellular and epigenetic barriers and improves direct cardiac reprogramming. Upon myocardial injury, fibroblasts as well as other non-myocyte cells, e.g., macrophages, are being activated to remodel infarcted heart tissue. Cardiac resident macrophage proliferation occurs within the first week following pressure overload hypertrophy and is a requisite for the heart’s adaptive response [106]. It is still unknown how much of those activated inflammation signals contribute to the enhancement of in vivo iCM-reprogramming. The close interactions between various cell types in the infarct and peri-infarct zone could play a critical role in improving cardiac reprogramming. For example, cardiac resident mesenchymal stem cells (MSCs) secrete molecules that protect the heart against damage [107]; therefore, secreted factors might improve the efficiency of direct cardiac reprogramming, while reprogramming factors may also have positive effects on cardiac MSCs in terms of secreting more cardioprotective and regenerative factors. In addition, the extracellular matrix in the heart provides proper stiffness and a topographical 3-D microenvironment, which is important for the induction and maturation of reprogrammed iCMs [91,92]. We speculate that direct mechanical and/or electrical interactions between iCMs and neighbor myocytes/non-myocytes/ECM factors, combined with endogenously secreted factors, may cooperatively work to improve direct cardiac reprogramming, which should be carefully studied and well understood before its clinical application.
5.2. Converting Chronic Fibrotic Scar into Myocardium
Noticeably, all published studies of in vivo reprogramming were performed with an acute MI model, with gene delivery right after coronary artery ligation (Figure 3A). Therefore, those freshly activated cardiac fibroblasts are the major cells targeted by different combinations of reprogramming factors. However, the majority of activated fibroblasts have been differentiated into αSMA+ myofibroblasts and further become αSMA− matrifibrocytes in a chronic scar [42]. So far, it is unknown whether these differentiated myofibroblasts/matrifibrocytes are reprogrammable in vitro (Figure 2B) and in vivo (Figure 3B). Several studies have demonstrated that achieving a successful cardiac reprogramming in vitro requires healthy and non-senescent fibroblasts [108,109,110]. In addition, treatment of virus-infected fibroblasts with TGFβ, which induces myofibroblasts differentiation, inhibits cardiac reprogramming [65], indicating the difficulty of reprogramming iCMs from myofibroblasts/ matrifibrocytes. Since there is still no effective treatment for a chronic cardiac scar, it will be priceless to develop a novel approach that can reprogram myofibroblasts/matrifibrocytes into iCMs, which is the first step to eventually converting chronic cardiac scar into functional muscle tissue, and will accelerate the translation of direct cardiac reprogramming. As we discussed earlier, variations of scar stiffness and size across the infarct might be another important aspect of reprogramming the chronic scar into iCMs, which needs to be addressed in future studies.
5.3. Direct Cardiac Reprogramming of Human Cells
Conversion of human fibroblasts into beating functional iCMs is still very challenging and not much progress has been made since the first three publications [57,58,61], although many mechanistic understandings of cardiac reprogramming have been achieved with mouse cells (Table 1). One realistic hurdle is that there is a very limited source to obtain high-quality human cardiac fibroblasts that, as found in mouse cells, is critical for the success of direct cardiac reprogramming. It is a consistent finding that human iCM reprogramming requires more reprogramming factors and takes a longer time than mouse reprogramming [57,58,61], especially because the conversion rate of human iCMs is still very low. The most recent study by Zhou et al. [86] developed a cell fate index algorithm from single-cell RNA-Seq data to assess reprogramming progression and found a slower progression of iCM reprogramming in human cells than that in mouse cells. Therefore, it is very important to develop an optimized protocol for the efficient reprogramming of human cells through a better understanding of its mechanism and eliminating more barriers. Indeed, an analytic technique, called trajectory alignment, identified barriers of MyoD-mediated human myogenic reprogramming, which could also be a helpful technique for identifying the barriers of human iCM reprogramming [111]. Furthermore, studying the safety and functional benefits of human iCM reprogramming with a large animal model (i.e., pig) is required for clinical translation of this technology; however, there is no successful report yet, although we have speculated that the in vivo environment of the heart should improve human cardiac reprogramming and yield fully functional iCMs in pig hearts [112]. Therefore, more effort should be invested in human cardiac reprogramming. A full understanding of direct cardiac reprogramming in human cells is a necessary requirement in order to consider its clinical applications.
6. Conclusions
Direct in vivo cardiac reprogramming allows the direct injection of reprogramming cocktails into injured heart and in situ regeneration without cell transplantation and is a promising technology for the replacement of damaged myocardium in ischemic heart diseases. Although enormous progresses have been made recently, the technology of direct cardiac reprogramming is still under development for future clinical applications. Better mechanistic understanding is required, particularly for in vivo reprogramming, the possibility of reprogramming a chronic cardiac scar, and human iCM reprogramming, in order to translate it to a promising alternative to cell-based therapies. Nevertheless, we are optimistic that the rapid development of iCM reprogramming technology will offer a new therapeutic treatment for millions of patients with ischemic heart diseases in the future.
Author Contributions
E.B.; writing—original draft preparation, J.-D.F.; writing—review and editing.
Funding
This study was supported by the Start-up Fund from The MetroHealth System (to J.-D.F) and grants from American Heart Association-13SDG14580035 (to J.-D.F.) and NIH-1R01HL139006 (to J.-D.F.).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Benjamin, E.J.; Blaha, M.J.; Chiuve, S.E.; Cushman, M.; Das, S.R.; Deo, R.; de Ferranti, S.D.; Floyd, J.; Fornage, M.; Gillespie, C.; et al. Heart Disease and Stroke Statistics-2017 Update: A Report from the American Heart Association. Circulation 2017, 135, e146–e603. [Google Scholar] [CrossRef] [PubMed]
- Savi, M.; Bocchi, L.; Fiumana, E.; Karam, J.P.; Frati, C.; Bonafe, F.; Cavalli, S.; Morselli, P.G.; Guarnieri, C.; Caldarera, C.M.; et al. Enhanced engraftment and repairing ability of human adipose-derived stem cells, conveyed by pharmacologically active microcarriers continuously releasing HGF and IGF-1, in healing myocardial infarction in rats. J. Biomed. Mater. Res. Part A 2015, 103, 3012–3025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, G.; Iyer, V.; Lee, T.C.; Canty, J.M., Jr. Autologous mesenchymal stem cells mobilize cKit+ and CD133+ bone marrow progenitor cells and improve regional function in hibernating myocardium. Circ. Res. 2011, 109, 1044–1054. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Chiu, S.; Liang, X.; Gao, F.; Zhang, Z.; Liao, S.; Liang, Y.; Chai, Y.H.; Low, D.J.; Tse, H.F.; et al. Rap1-mediated nuclear factor-kappaB (NF-kappaB) activity regulates the paracrine capacity of mesenchymal stem cells in heart repair following infarction. Cell Death Discov. 2015, 1, 15007. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Liu, X.; Zhang, Y.; Liang, X.; Ding, Y.; Xu, Y.; Fang, Z.; Zhang, F. Enhanced cell survival and paracrine effects of mesenchymal stem cells overexpressing hepatocyte growth factor promote cardioprotection in myocardial infarction. Exp. Cell Res. 2016, 344, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Giacomelli, E.; Mummery, C.L.; Bellin, M. Human heart disease: Lessons from human pluripotent stem cell-derived cardiomyocytes. Cell. Mol. Life Sci. 2017, 74, 3711–3739. [Google Scholar] [CrossRef] [PubMed]
- Mandel, Y.; Weissman, A.; Schick, R.; Barad, L.; Novak, A.; Meiry, G.; Goldberg, S.; Lorber, A.; Rosen, M.R.; Itskovitz-Eldor, J.; et al. Human embryonic and induced pluripotent stem cell-derived cardiomyocytes exhibit beat rate variability and power-law behavior. Circulation 2012, 125, 883–893. [Google Scholar] [CrossRef]
- Moreau, A.; Boutjdir, M.; Chahine, M. Induced pluripotent stem-cell-derived cardiomyocytes: Cardiac applications, opportunities, and challenges. Can. J. Physiol. Pharmacol. 2017, 95, 1108–1116. [Google Scholar] [CrossRef]
- Smart, N.; Bollini, S.; Dube, K.N.; Vieira, J.M.; Zhou, B.; Davidson, S.; Yellon, D.; Riegler, J.; Price, A.N.; Lythgoe, M.F.; et al. De novo cardiomyocytes from within the activated adult heart after injury. Nature 2011, 474, 640. [Google Scholar] [CrossRef]
- Ellison, G.M.; Torella, D.; Dellegrottaglie, S.; Perez-Martinez, C.; Perez de Prado, A.; Vicinanza, C.; Purushothaman, S.; Galuppo, V.; Iaconetti, C.; Waring, C.D.; et al. Endogenous cardiac stem cell activation by insulin-like growth factor-1/hepatocyte growth factor intracoronary injection fosters survival and regeneration of the infarcted pig heart. J. Am. Coll. Cardiol. 2011, 58, 977–986. [Google Scholar] [CrossRef]
- Urbanek, K.; Rota, M.; Cascapera, S.; Bearzi, C.; Nascimbene, A.; De Angelis, A.; Hosoda, T.; Chimenti, S.; Baker, M.; Limana, F.; et al. Cardiac stem cells possess growth factor-receptor systems that after activation regenerate the infarcted myocardium, improving ventricular function and long-term survival. Circ. Res. 2005, 97, 663–673. [Google Scholar] [CrossRef] [PubMed]
- Linke, A.; Muller, P.; Nurzynska, D.; Casarsa, C.; Torella, D.; Nascimbene, A.; Castaldo, C.; Cascapera, S.; Bohm, M.; Quaini, F.; et al. Stem cells in the dog heart are self-renewing, clonogenic, and multipotent and regenerate infarcted myocardium, improving cardiac function. Proc. Natl. Acad. Sci. USA 2005, 102, 8966–8971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campa, V.M.; Gutierrez-Lanza, R.; Cerignoli, F.; Diaz-Trelles, R.; Nelson, B.; Tsuji, T.; Barcova, M.; Jiang, W.; Mercola, M. Notch activates cell cycle reentry and progression in quiescent cardiomyocytes. J. Cell Biol. 2008, 183, 129–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engel, F.B.; Hauck, L.; Cardoso, M.C.; Leonhardt, H.; Dietz, R.; von Harsdorf, R. A mammalian myocardial cell-free system to study cell cycle reentry in terminally differentiated cardiomyocytes. Circ. Res. 1999, 85, 294–301. [Google Scholar] [CrossRef] [PubMed]
- Miyawaki, A.; Obana, M.; Mitsuhara, Y.; Orimoto, A.; Nakayasu, Y.; Yamashita, T.; Fukada, S.I.; Maeda, M.; Nakayama, H.; Fujio, Y. Adult murine cardiomyocytes exhibit regenerative activity with cell cycle reentry through STAT3 in the healing process of myocarditis. Sci. Rep. 2017, 7, 1407. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, T.M.A.; Ang, Y.S.; Radzinsky, E.; Zhou, P.; Huang, Y.; Elfenbein, A.; Foley, A.; Magnitsky, S.; Srivastava, D. Regulation of Cell Cycle to Stimulate Adult Cardiomyocyte Proliferation and Cardiac Regeneration. Cell 2018, 173, 104–116. [Google Scholar] [CrossRef]
- Ieda, M.; Fu, J.D.; Delgado-Olguin, P.; Vedantham, V.; Hayashi, Y.; Bruneau, B.G.; Srivastava, D. Direct Reprogramming of Fibroblasts into Functional Cardiomyocytes by Defined Factors. Cell 2010, 142, 375–386. [Google Scholar] [CrossRef] [Green Version]
- Inagawa, K.; Miyamoto, K.; Yamakawa, H.; Muraoka, N.; Sadahiro, T.; Umei, T.; Wada, R.; Katsumata, Y.; Kaneda, R.; Nakade, K.; et al. Induction of cardiomyocyte-like cells in infarct hearts by gene transfer of Gata4, Mef2c, and Tbx5. Circ. Res. 2012, 111, 1147–1156. [Google Scholar] [CrossRef]
- Jayawardena, T.M.; Egemnazarov, B.; Finch, E.A.; Zhang, L.; Payne, J.A.; Pandya, K.; Zhang, Z.; Rosenberg, P.; Mirotsou, M.; Dzau, V.J. MicroRNA-mediated in vitro and in vivo direct reprogramming of cardiac fibroblasts to cardiomyocytes. Circ. Res. 2012, 110, 1465–1473. [Google Scholar] [CrossRef]
- Qian, L.; Huang, Y.; Spencer, C.I.; Foley, A.; Vedantham, V.; Liu, L.; Conway, S.J.; Fu, J.-d.D.; Srivastava, D. In vivo reprogramming of murine cardiac fibroblasts into induced cardiomyocytes. Nature 2012, 485, 593–598. [Google Scholar] [CrossRef]
- Song, K.; Nam, Y.J.; Luo, X.; Qi, X.; Tan, W.; Huang, G.N.; Acharya, A.; Smith, C.L.; Tallquist, M.D.; Neilson, E.G.; et al. Heart repair by reprogramming non-myocytes with cardiac transcription factors. Nature 2012, 485, 599–604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tzahor, E.; Poss, K.D. Cardiac regeneration strategies: Staying young at heart. Science 2017, 356, 1035–1039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Z.; Pu, W.T. Strategies for cardiac regeneration and repair. Sci. Transl. Med. 2014, 6, 239rv1. [Google Scholar] [CrossRef] [PubMed]
- Tani, H.; Sadahiro, T.; Ieda, M. Direct Cardiac Reprogramming: A Novel Approach for Heart Regeneration. Int. J. Mol. Sci. 2018, 19, 2629. [Google Scholar] [CrossRef] [PubMed]
- Vaseghi, H.; Liu, J.; Qian, L. Molecular barriers to direct cardiac reprogramming. Protein Cell 2017. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Yang, Z.; Zhao, Z.-A.A.; Shen, Z. Direct reprogramming of fibroblasts into cardiomyocytes. Stem Cell Res. Ther. 2017, 8, 118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frangogiannis, N.G. Pathophysiology of Myocardial Infarction. Compr. Physiol. 2015, 5, 1841–1875. [Google Scholar] [CrossRef]
- Bergmann, O.; Zdunek, S.; Felker, A.; Salehpour, M.; Alkass, K.; Bernard, S.; Sjostrom, S.L.; Szewczykowska, M.; Jackowska, T.; Dos Remedios, C.; et al. Dynamics of Cell Generation and Turnover in the Human Heart. Cell 2015, 161, 1566–1575. [Google Scholar] [CrossRef]
- Banerjee, I.; Fuseler, J.W.; Price, R.L.; Borg, T.K.; Baudino, T.A. Determination of cell types and numbers during cardiac development in the neonatal and adult rat and mouse. Am. J. Physiol. Heart Circ. Physiol. 2007, 293, H1883–H1891. [Google Scholar] [CrossRef] [Green Version]
- Pinto, A.R.; Ilinykh, A.; Ivey, M.J.; Kuwabara, J.T.; D’Antoni, M.L.; Debuque, R.; Chandran, A.; Wang, L.; Arora, K.; Rosenthal, N.A.; et al. Revisiting Cardiac Cellular Composition. Circ. Res. 2016, 118, 400–409. [Google Scholar] [CrossRef] [Green Version]
- Souders, C.A.; Bowers, S.L.; Baudino, T.A. Cardiac fibroblast: The renaissance cell. Circ. Res. 2009, 105, 1164–1176. [Google Scholar] [CrossRef] [PubMed]
- Zeisberg, E.M.; Kalluri, R. Origins of cardiac fibroblasts. Circ. Res. 2010, 107, 1304–1312. [Google Scholar] [CrossRef] [PubMed]
- Dewald, O.; Zymek, P.; Winkelmann, K.; Koerting, A.; Ren, G.; Abou-Khamis, T.; Michael, L.H.; Rollins, B.J.; Entman, M.L.; Frangogiannis, N.G. CCL2/Monocyte Chemoattractant Protein-1 regulates inflammatory responses critical to healing myocardial infarcts. Circ. Res. 2005, 96, 881–889. [Google Scholar] [CrossRef] [PubMed]
- Frangogiannis, N.G.; Michael, L.H.; Entman, M.L. Myofibroblasts in reperfused myocardial infarcts express the embryonic form of smooth muscle myosin heavy chain (SMemb). Cardiovasc. Res. 2000, 48, 89–100. [Google Scholar] [CrossRef] [Green Version]
- Ivey, M.J.; Kuwabara, J.T.; Pai, J.T.; Moore, R.E.; Sun, Z.; Tallquist, M.D. Resident fibroblast expansion during cardiac growth and remodeling. J. Mol. Cell. Cardiol. 2018, 114, 161–174. [Google Scholar] [CrossRef] [PubMed]
- Petrov, V.V.; Fagard, R.H.; Lijnen, P.J. Stimulation of collagen production by transforming growth factor-beta1 during differentiation of cardiac fibroblasts to myofibroblasts. Hypertension 2002, 39, 258–263. [Google Scholar] [CrossRef] [PubMed]
- Saxena, A.; Bujak, M.; Frunza, O.; Dobaczewski, M.; Gonzalez-Quesada, C.; Lu, B.; Gerard, C.; Frangogiannis, N.G. CXCR3-independent actions of the CXC chemokine CXCL10 in the infarcted myocardium and in isolated cardiac fibroblasts are mediated through proteoglycans. Cardiovasc. Res. 2014, 103, 217–227. [Google Scholar] [CrossRef] [Green Version]
- Bujak, M.; Dobaczewski, M.; Gonzalez-Quesada, C.; Xia, Y.; Leucker, T.; Zymek, P.; Veeranna, V.; Tager, A.M.; Luster, A.D.; Frangogiannis, N.G. Induction of the CXC chemokine interferon-gamma-inducible protein 10 regulates the reparative response following myocardial infarction. Circ. Res. 2009, 105, 973–983. [Google Scholar] [CrossRef]
- Czubryt, M.P. Common threads in cardiac fibrosis, infarct scar formation, and wound healing. Fibrogenes. Tissue Repair 2012, 5, 19. [Google Scholar] [CrossRef]
- Desmouliere, A.; Redard, M.; Darby, I.; Gabbiani, G. Apoptosis mediates the decrease in cellularity during the transition between granulation tissue and scar. Am. J. Pathol. 1995, 146, 56–66. [Google Scholar]
- Willems, I.E.; Havenith, M.G.; De Mey, J.G.; Daemen, M.J. The alpha-smooth muscle actin-positive cells in healing human myocardial scars. Am. J. Pathol. 1994, 145, 868–875. [Google Scholar] [PubMed]
- Fu, X.; Khalil, H.; Kanisicak, O.; Boyer, J.G.; Vagnozzi, R.J.; Maliken, B.D.; Sargent, M.A.; Prasad, V.; Valiente-Alandi, I.; Blaxall, B.C.; et al. Specialized fibroblast differentiated states underlie scar formation in the infarcted mouse heart. J. Clin. Investig. 2018, 128, 2127–2143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, J.; Costa, M.L.; Mermelstein, C.S.; Chagas, C.; Holtzer, S.; Holtzer, H. MyoD converts primary dermal fibroblasts, chondroblasts, smooth muscle, and retinal pigmented epithelial cells into striated mononucleated myoblasts and multinucleated myotubes. Proc. Natl. Acad. Sci. USA 1990, 87, 7988–7992. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Yamanaka, S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 2006, 126, 663–676. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Tanabe, K.; Ohnuki, M.; Narita, M.; Ichisaka, T.; Tomoda, K.; Yamanaka, S. Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell 2007, 131, 861–872. [Google Scholar] [CrossRef]
- Addis, R.C.; Ifkovits, J.L.; Pinto, F.; Kellam, L.D.; Esteso, P.; Rentschler, S.; Christoforou, N.; Epstein, J.A.; Gearhart, J.D. Optimization of direct fibroblast reprogramming to cardiomyocytes using calcium activity as a functional measure of success. J. Mol. Cell. Cardiol. 2013, 60, 97–106. [Google Scholar] [CrossRef] [Green Version]
- Protze, S.; Khattak, S.; Poulet, C.; Lindemann, D.; Tanaka, E.M.; Ravens, U. A new approach to transcription factor screening for reprogramming of fibroblasts to cardiomyocyte-like cells. J. Mol. Cell. Cardiol. 2012, 53, 323–332. [Google Scholar] [CrossRef]
- Hirai, H.; Katoku-Kikyo, N.; Keirstead, S.A.; Kikyo, N. Accelerated direct reprogramming of fibroblasts into cardiomyocyte-like cells with the MyoD transactivation domain. Cardiovasc. Res. 2013, 100, 105–113. [Google Scholar] [CrossRef] [Green Version]
- Muraoka, N.; Yamakawa, H.; Miyamoto, K.; Sadahiro, T.; Umei, T.; Isomi, M.; Nakashima, H.; Akiyama, M.; Wada, R.; Inagawa, K.; et al. MiR-133 promotes cardiac reprogramming by directly repressing Snai1 and silencing fibroblast signatures. EMBO J. 2014, 33, 1565–1581. [Google Scholar] [CrossRef]
- Stone, N.R.; Gifford, C.A.; Thomas, R.; Pratt, K.J.B.; Samse-Knapp, K.; Mohamed, T.M.A.; Radzinsky, E.M.; Schricker, A.; Yu, P.; Ivey, K.N.; et al. Unique Transcription Factor Functions Regulate Epigenetic and Transcriptional Dynamics During Cardiac Reprogramming. bioRxiv 2019. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhang, A.D.; Kim, L.J.; Nam, Y.J. Ensuring expression of four core cardiogenic transcription factors enhances cardiac reprogramming. Sci. Rep. 2019, 9, 6362. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, D.; Ieda, M.; Fu, J.; Qian, L. Cardiac repair with thymosin β4 and cardiac reprogramming factors. Ann. N. Y. Acad. Sci. 2012, 1270, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Jayawardena, T.M.; Finch, E.A.; Zhang, L.; Zhang, H.; Hodgkinson, C.P.; Pratt, R.E.; Rosenberg, P.B.; Mirotsou, M.; Dzau, V.J. MicroRNA induced cardiac reprogramming in vivo: Evidence for mature cardiac myocytes and improved cardiac function. Circ. Res. 2015, 116, 418–424. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Liu, Z.; Yin, C.; Asfour, H.; Chen, O.; Li, Y.; Bursac, N.; Liu, J.; Qian, L. Stoichiometry of Gata4, Mef2c, and Tbx5 Influences the Efficiency and Quality of Induced Cardiac Myocyte Reprogramming. Circ. Res. 2015, 116, 237–244. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.; Wang, L.; Yin, C.; Liu, J.; Qian, L. In vivo cardiac reprogramming using an optimal single polycistronic construct. Cardiovasc. Res. 2015, 108, 217–219. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Wang, L.; Welch, J.D.; Ma, H.; Zhou, Y.; Vaseghi, H.R.; Yu, S.; Wall, J.B.; Alimohamadi, S.; Zheng, M.; et al. Single-cell transcriptomics reconstructs fate conversion from fibroblast to cardiomyocyte. Nature 2017. [Google Scholar] [CrossRef]
- Wada, R.; Muraoka, N.; Inagawa, K.; Yamakawa, H.; Miyamoto, K.; Sadahiro, T.; Umei, T.; Kaneda, R.; Suzuki, T.; Kamiya, K.; et al. Induction of human cardiomyocyte-like cells from fibroblasts by defined factors. Proc. Natl. Acad. Sci. USA 2013, 110, 12667–12672. [Google Scholar] [CrossRef] [Green Version]
- Fu, J.-D.D.; Stone, N.R.; Liu, L.; Spencer, C.I.; Qian, L.; Hayashi, Y.; Delgado-Olguin, P.; Ding, S.; Bruneau, B.G.; Srivastava, D. Direct reprogramming of human fibroblasts toward a cardiomyocyte-like state. Stem Cell Rep. 2013, 1, 235–247. [Google Scholar] [CrossRef]
- Bektik, E.; Dennis, A.; Prasanna, P.; Madabhushi, A.; Fu, J.-D.D. Single cell qPCR reveals that additional HAND2 and microRNA-1 facilitate the early reprogramming progress of seven-factor-induced human myocytes. PLoS ONE 2017, 12, e0183000. [Google Scholar] [CrossRef]
- Mohamed, T.M.A.; Stone, N.R.; Berry, E.C.; Radzinsky, E.; Huang, Y.; Pratt, K.; Ang, Y.-S.; Yu, P.; Wang, H.; Tang, S.; et al. Chemical Enhancement of In Vitro and In Vivo Direct Cardiac Reprogramming Clinical Perspective. Circulation 2016, 135, 978–995. [Google Scholar] [CrossRef]
- Nam, Y.J.; Song, K.; Luo, X.; Daniel, E.; Lambeth, K.; West, K.; Hill, J.A.; DiMaio, J.M.; Baker, L.A.; Bassel-Duby, R.; et al. Reprogramming of human fibroblasts toward a cardiac fate. Proc. Natl. Acad. Sci. USA 2013, 110, 5588–5593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lalit, P.A.; Salick, M.R.; Nelson, D.O.; Squirrell, J.M.; Shafer, C.M.; Patel, N.G.; Saeed, I.; Schmuck, E.G.; Markandeya, Y.S.; Wong, R.; et al. Lineage Reprogramming of Fibroblasts into Proliferative Induced Cardiac Progenitor Cells by Defined Factors. Cell Stem Cell 2016, 18, 354–367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Cao, N.; Huang, Y.; Spencer, I.C.; Fu, J.-d.; Yu, C.; Liu, K.; Nie, B.; Xu, T.; Li, K.; et al. Expandable Cardiovascular Progenitor Cells Reprogrammed from Fibroblasts. Cell Stem Cell 2016, 18, 368–381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ifkovits, J.L.; Addis, R.C.; Epstein, J.A.; Gearhart, J.D. Inhibition of TGFbeta signaling increases direct conversion of fibroblasts to induced cardiomyocytes. PLoS ONE 2014, 9, e89678. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Londono, P.; Cao, Y.; Sharpe, E.J.; Proenza, C.; O’Rourke, R.; Jones, K.L.; Jeong, M.Y.; Walker, L.A.; Buttrick, P.M.; et al. High-efficiency reprogramming of fibroblasts into cardiomyocytes requires suppression of pro-fibrotic signalling. Nat. Commun. 2015, 6, 8243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bektik, E.; Dennis, A.; Pawlowski, G.; Zhou, C.; Maleski, D.; Takahashi, S.; Laurita, K.R.; Deschênes, I.; Fu, J.-D.D. S-phase Synchronization Facilitates the Early Progression of Induced-Cardiomyocyte Reprogramming through Enhanced Cell-Cycle Exit. Int. J. Mol. Sci. 2018, 19, 1364. [Google Scholar] [CrossRef] [PubMed]
- Abad, M.; Hashimoto, H.; Zhou, H.; Morales, M.G.; Chen, B.; Bassel-Duby, R.; Olson, E.N. Notch Inhibition Enhances Cardiac Reprogramming by Increasing MEF2C Transcriptional Activity. Stem Cell Rep. 2017, 8, 548–560. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Dickson, M.E.; Kim, M.S.; Bassel-Duby, R.; Olson, E.N. Akt1/protein kinase B enhances transcriptional reprogramming of fibroblasts to functional cardiomyocytes. Proc. Natl. Acad. Sci. USA 2015, 112, 11864–11869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamakawa, H.; Muraoka, N.; Miyamoto, K.; Sadahiro, T.; Isomi, M.; Haginiwa, S.; Kojima, H.; Umei, T.; Akiyama, M.; Kuishi, Y.; et al. Fibroblast Growth Factors and Vascular Endothelial Growth Factor Promote Cardiac Reprogramming under Defined Conditions. Stem Cell Rep. 2015, 5, 1128–1142. [Google Scholar] [CrossRef] [Green Version]
- Zhou, H.; Morales, M.G.; Hashimoto, H.; Dickson, M.E.; Song, K.; Ye, W.; Kim, M.S.; Niederstrasser, H.; Wang, Z.; Chen, B.; et al. ZNF281 enhances cardiac reprogramming by modulating cardiac and inflammatory gene expression. Genes Dev. 2017, 31, 1770–1783. [Google Scholar] [CrossRef] [Green Version]
- Yijing, G.; Ienglam, L.; Shuo, T.; Wenbin, G.; Karatas, H.; Yangbing, L.; Shaomeng, W.; Liu, L.; Zhong, W. Enhancing Cardiac Reprogramming by Suppressing Specific C-C Chemokine Signaling Pathways. bioRxiv 2019. [Google Scholar] [CrossRef]
- Muraoka, N.; Nara, K.; Tamura, F.; Kojima, H.; Yamakawa, H.; Sadahiro, T.; Miyamoto, K.; Isomi, M.; Haginiwa, S.; Tani, H.; et al. Role of cyclooxygenase-2-mediated prostaglandin E2-prostaglandin E receptor 4 signaling in cardiac reprogramming. Nat. Commun. 2019, 10, 674. [Google Scholar] [CrossRef] [PubMed]
- Soufi, A.; Donahue, G.; Zaret, K.S. Facilitators and impediments of the pluripotency reprogramming factors’ initial engagement with the genome. Cell 2012, 151, 994–1004. [Google Scholar] [CrossRef] [PubMed]
- Chronis, C.; Fiziev, P.; Papp, B.; Butz, S.; Bonora, G.; Sabri, S.; Ernst, J.; Plath, K. Cooperative Binding of Transcription Factors Orchestrates Reprogramming. Cell 2017, 168, 442–459. [Google Scholar] [CrossRef] [PubMed]
- Wapinski, O.L.; Vierbuchen, T.; Qu, K.; Lee, Q.Y.; Chanda, S.; Fuentes, D.R.; Giresi, P.G.; Ng, Y.H.; Marro, S.; Neff, N.F.; et al. Hierarchical mechanisms for direct reprogramming of fibroblasts to neurons. Cell 2013, 155, 621–635. [Google Scholar] [CrossRef] [PubMed]
- Chanda, S.; Ang, C.E.; Davila, J.; Pak, C.; Mall, M.; Lee, Q.Y.; Ahlenius, H.; Jung, S.W.; Sudhof, T.C.; Wernig, M. Generation of induced neuronal cells by the single reprogramming factor ASCL1. Stem Cell Rep. 2014, 3, 282–296. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Chen, O.; Zheng, M.; Wang, L.; Zhou, Y.; Yin, C.; Liu, J.; Qian, L. Re-patterning of H3K27me3, H3K4me3 and DNA methylation during fibroblast conversion into induced cardiomyocytes. Stem Cell Res. 2016, 16, 507–518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirai, H.; Kikyo, N. Inhibitors of suppressive histone modification promote direct reprogramming of fibroblasts to cardiomyocyte-like cells. Cardiovasc. Res. 2014, 102, 188–190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dal-Pra, S.; Hodgkinson, C.P.; Mirotsou, M.; Kirste, I.; Dzau, V.J. Demethylation of H3K27 Is Essential for the Induction of Direct Cardiac Reprogramming by miR Combo. Circ. Res. 2017, 120, 1403–1413. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Wang, L.; Vaseghi, H.R.; Liu, Z.; Lu, R.; Alimohamadi, S.; Yin, C.; Fu, J.-D.D.; Wang, G.G.; Liu, J.; et al. Bmi1 Is a Key Epigenetic Barrier to Direct Cardiac Reprogramming. Cell Stem Cell 2016, 18, 382–395. [Google Scholar] [CrossRef] [Green Version]
- Hashimoto, H.; Wang, Z.; Garry, G.A.; Malladi, V.S.; Botten, G.A.; Ye, W.; Zhou, H.; Osterwalder, M.; Dickel, D.E.; Visel, A.; et al. Cardiac Reprogramming Factors Synergistically Activate Genome-wide Cardiogenic Stage-Specific Enhancers. Cell Stem Cell 2019. [Google Scholar] [CrossRef] [PubMed]
- Donaghey, J.; Thakurela, S.; Charlton, J.; Chen, J.S.; Smith, Z.D.; Gu, H.; Pop, R.; Clement, K.; Stamenova, E.K.; Karnik, R.; et al. Genetic determinants and epigenetic effects of pioneer-factor occupancy. Nat. Genet. 2018, 50, 250–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, S.E.; Fraga, M.F.; Weaver, I.C.; Berdasco, M.; Szyf, M. Variations in DNA methylation patterns during the cell cycle of HeLa cells. Epigenetics 2007, 2, 54–65. [Google Scholar] [CrossRef] [PubMed]
- Bou Kheir, T.; Lund, A.H. Epigenetic dynamics across the cell cycle. Essays Biochem. 2010, 48, 107–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Wang, L.; Liu, Z.; Alimohamadi, S.; Yin, C.; Liu, J.; Qian, L. Comparative Gene Expression Analyses Reveal Distinct Molecular Signatures between Differentially Reprogrammed Cardiomyocytes. Cell Rep. 2017, 20, 3014–3024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Liu, Z.; Welch, J.D.; Gao, X.; Wang, L.; Garbutt, T.; Keepers, B.; Ma, H.; Prins, J.F.; Shen, W.; et al. Single-Cell Transcriptomic Analyses of Cell Fate Transitions during Human Cardiac Reprogramming. Cell Stem Cell 2019. [Google Scholar] [CrossRef]
- Takeuchi, T. Regulation of cardiomyocyte proliferation during development and regeneration. Dev. Growth Differ. 2014, 56, 402–409. [Google Scholar] [CrossRef] [Green Version]
- Chopra, A.; Lin, V.; McCollough, A.; Atzet, S.; Prestwich, G.D.; Wechsler, A.S.; Murray, M.E.; Oake, S.A.; Kresh, J.Y.; Janmey, P.A. Reprogramming cardiomyocyte mechanosensing by crosstalk between integrins and hyaluronic acid receptors. J. Biomech. 2012, 45, 824–831. [Google Scholar] [CrossRef] [Green Version]
- Engler, A.J.; Carag-Krieger, C.; Johnson, C.P.; Raab, M.; Tang, H.Y.; Speicher, D.W.; Sanger, J.W.; Sanger, J.M.; Discher, D.E. Embryonic cardiomyocytes beat best on a matrix with heart-like elasticity: Scar-like rigidity inhibits beating. J. Cell Sci. 2008, 121, 3794–3802. [Google Scholar] [CrossRef]
- Ribeiro, A.J.; Ang, Y.S.; Fu, J.D.; Rivas, R.N.; Mohamed, T.M.; Higgs, G.C.; Srivastava, D.; Pruitt, B.L. Contractility of single cardiomyocytes differentiated from pluripotent stem cells depends on physiological shape and substrate stiffness. Proc. Natl. Acad. Sci. USA 2015, 112, 12705–12710. [Google Scholar] [CrossRef] [Green Version]
- Sia, J.; Yu, P.; Srivastava, D.; Li, S. Effect of biophysical cues on reprogramming to cardiomyocytes. Biomaterials 2016, 103, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.Z.; Dal-Pra, S.; Mirotsou, M.; Jayawardena, T.M.; Hodgkinson, C.P.; Bursac, N.; Dzau, V.J. Tissue-engineered 3-dimensional (3D) microenvironment enhances the direct reprogramming of fibroblasts into cardiomyocytes by microRNAs. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Sun, A.; Ma, H.; Yao, K.; Zhou, N.; Shen, L.; Zhang, C.; Zou, Y.; Ge, J. Infarcted myocardium-like stiffness contributes to endothelial progenitor lineage commitment of bone marrow mononuclear cells. J. Cell. Mol. Med. 2011, 15, 2245–2261. [Google Scholar] [CrossRef] [PubMed]
- Voorhees, A.P.; DeLeon-Pennell, K.Y.; Ma, Y.; Halade, G.V.; Yabluchanskiy, A.; Iyer, R.P.; Flynn, E.; Cates, C.A.; Lindsey, M.L.; Han, H.C. Building a better infarct: Modulation of collagen cross-linking to increase infarct stiffness and reduce left ventricular dilation post-myocardial infarction. J. Mol. Cell. Cardiol. 2015, 85, 229–239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doi, K.; Takeuchi, Y. Gene therapy using retrovirus vectors: Vector development and biosafety at clinical trials. Uirusu 2015, 65, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Vrljicak, P.; Tao, S.; Varshney, G.K.; Quach, H.N.; Joshi, A.; LaFave, M.C.; Burgess, S.M.; Sampath, K. Genome-Wide Analysis of Transposon and Retroviral Insertions Reveals Preferential Integrations in Regions of DNA Flexibility. G3 2016, 6, 805–817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mathison, M.; Gersch, R.P.; Nasser, A.; Lilo, S.; Korman, M.; Fourman, M.; Hackett, N.; Shroyer, K.; Yang, J.; Ma, Y.; et al. In vivo cardiac cellular reprogramming efficacy is enhanced by angiogenic preconditioning of the infarcted myocardium with vascular endothelial growth factor. J. Am. Heart Assoc. 2012, 1. [Google Scholar] [CrossRef] [PubMed]
- Mathison, M.; Singh, V.P.; Chiuchiolo, M.J.; Sanagasetti, D.; Mao, Y.; Patel, V.B.; Yang, J.; Kaminsky, S.M.; Crystal, R.G.; Rosengart, T.K. In situ reprogramming to transdifferentiate fibroblasts into cardiomyocytes using adenoviral vectors: Implications for clinical myocardial regeneration. J. Thorac. Cardiovasc. Surg. 2017, 153, 329–339. [Google Scholar] [CrossRef]
- Nam, Y.-J.J.; Lubczyk, C.; Bhakta, M.; Zang, T.; Fernandez-Perez, A.; McAnally, J.; Bassel-Duby, R.; Olson, E.N.; Munshi, N.V. Induction of diverse cardiac cell types by reprogramming fibroblasts with cardiac transcription factors. Development 2014, 141, 4267–4278. [Google Scholar] [CrossRef] [Green Version]
- Yoo, S.Y.; Jeong, S.-N.N.; Kang, J.-I.I.; Lee, S.-W.W. Chimeric Adeno-Associated Virus-Mediated Cardiovascular Reprogramming for Ischemic Heart Disease. ACS Omega 2018, 3, 5918–5925. [Google Scholar] [CrossRef]
- Miyamoto, K.; Akiyama, M.; Tamura, F.; Isomi, M.; Yamakawa, H.; Sadahiro, T.; Muraoka, N.; Kojima, H.; Haginiwa, S.; Kurotsu, S.; et al. Direct In Vivo Reprogramming with Sendai Virus Vectors Improves Cardiac Function after Myocardial Infarction. Cell Stem Cell 2018, 22, 91–103. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.; Lee, E.; Kim, J.; Kwon, Y.-W.; Kwon, Y.; Kim, J. Efficient in vivo direct conversion of fibroblasts into cardiomyocytes using a nanoparticle-based gene carrier. Biomaterials 2018. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Huang, C.; Xu, X.; Gu, H.; Ye, Y.; Jiang, C.; Qiu, Z.; Xie, X. Direct reprogramming of mouse fibroblasts into cardiomyocytes with chemical cocktails. Cell Res. 2015, 25, 1013–1024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, C.; Tu, W.; Fu, Y.; Wang, J.; Xie, X. Chemical-induced cardiac reprogramming in vivo. Cell Res. 2018, 28, 686–689. [Google Scholar] [CrossRef] [PubMed]
- Cao, N.; Huang, Y.; Zheng, J.; Spencer, C.I.; Zhang, Y.; Fu, J.-D.D.; Nie, B.; Xie, M.; Zhang, M.; Wang, H.; et al. Conversion of human fibroblasts into functional cardiomyocytes by small molecules. Science 2016, 352, 1216–1220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, X.; Shen, Y.; Zhang, R.; Sugi, K.; Vasudevan, N.T.; Alaiti, M.A.; Sweet, D.R.; Zhou, L.; Qing, Y.; Gerson, S.L.; et al. Distinct roles of resident and nonresident macrophages in nonischemic cardiomyopathy. Proc. Natl. Acad. Sci. USA 2018, 115, E4661–E4669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Golpanian, S.; Wolf, A.; Hatzistergos, K.E.; Hare, J.M. Rebuilding the Damaged Heart: Mesenchymal Stem Cells, Cell-Based Therapy, and Engineered Heart Tissue. Physiol. Rev. 2016, 96, 1127–1168. [Google Scholar] [CrossRef] [PubMed]
- Qian, L.; Berry, E.C.; Fu, J.D.; Ieda, M.; Srivastava, D. Reprogramming of mouse fibroblasts into cardiomyocyte-like cells in vitro. Nat. Protoc. 2013, 8, 1204–1215. [Google Scholar] [CrossRef]
- Srivastava, D.; Ieda, M. Critical factors for cardiac reprogramming. Circ. Res. 2012, 111, 5–8. [Google Scholar] [CrossRef]
- Chen, J.X.; Krane, M.; Deutsch, M.A.; Wang, L.; Rav-Acha, M.; Gregoire, S.; Engels, M.C.; Rajarajan, K.; Karra, R.; Abel, E.D.; et al. Inefficient reprogramming of fibroblasts into cardiomyocytes using Gata4, Mef2c, and Tbx5. Circ. Res. 2012, 111, 50–55. [Google Scholar] [CrossRef]
- Cacchiarelli, D.; Qiu, X.; Srivatsan, S.; Manfredi, A.; Ziller, M.; Overbey, E.; Grimaldi, A.; Grimsby, J.; Pokharel, P.; Livak, K.J.; et al. Aligning Single-Cell Developmental and Reprogramming Trajectories Identifies Molecular Determinants of Myogenic Reprogramming Outcome. Cell Syst. 2018, 7, 258–268. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.D.; Srivastava, D. Direct reprogramming of fibroblasts into cardiomyocytes for cardiac regenerative medicine. Circ. J. 2015, 79, 245–254. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Schematic representation of cardiac remodeling progression in post-infarction mouse hearts. (A) Cardiac fibroblasts are normally quiescent in the heart. (B) Cardiac fibroblasts are activated and quickly reenter the cell cycle after myocardium infarction (MI). (C) Cardiac fibroblasts differentiated into myofibroblasts. (D) Matrifibrocytes are further differentiated from myofibroblasts. (E) A mature scar is formed to mechanically support and protect the heart.
Figure 1.
Schematic representation of cardiac remodeling progression in post-infarction mouse hearts. (A) Cardiac fibroblasts are normally quiescent in the heart. (B) Cardiac fibroblasts are activated and quickly reenter the cell cycle after myocardium infarction (MI). (C) Cardiac fibroblasts differentiated into myofibroblasts. (D) Matrifibrocytes are further differentiated from myofibroblasts. (E) A mature scar is formed to mechanically support and protect the heart.

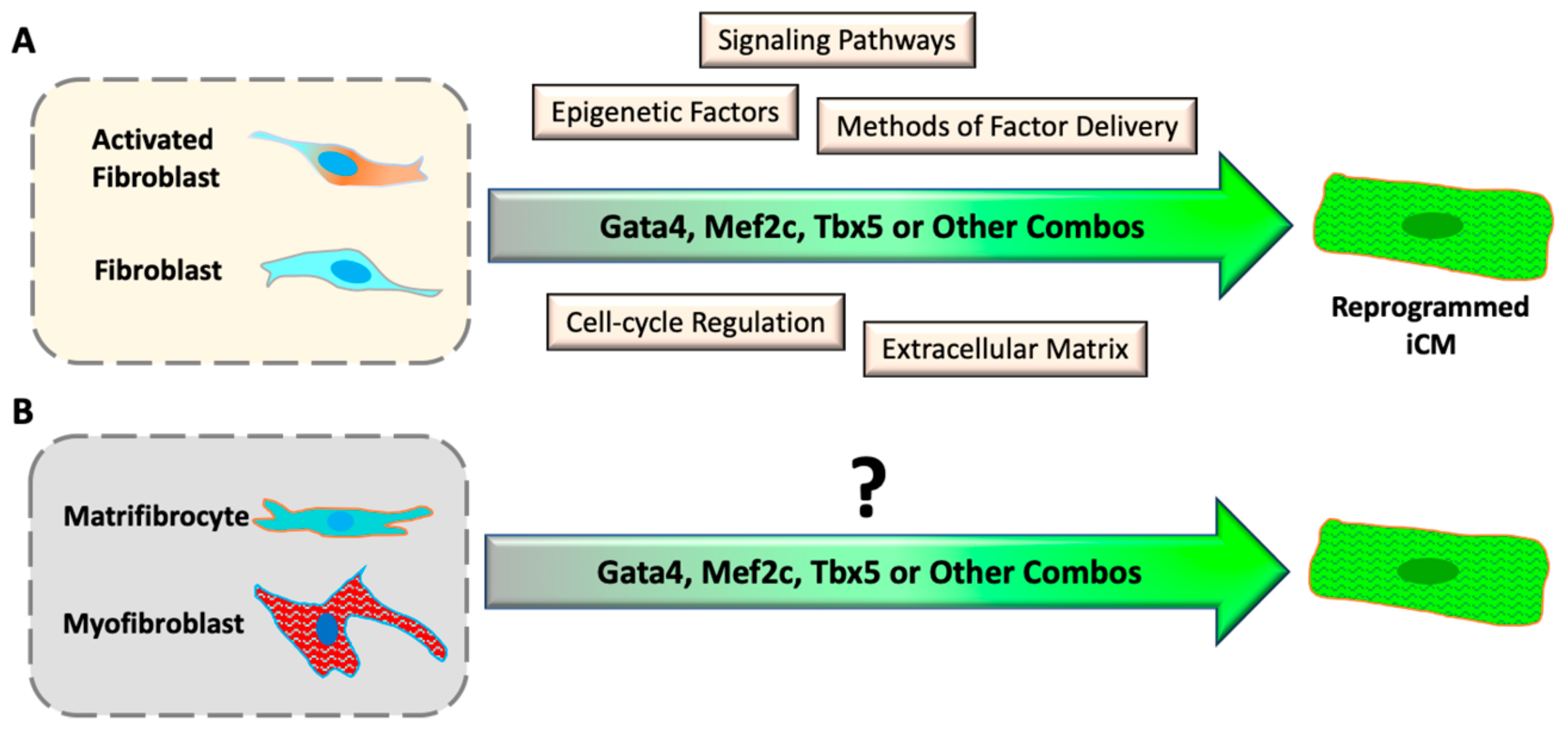
Figure 2.
Progresses and challenges of direct cardiac reprogramming in vitro. (A) Understanding the mechanism of iCM reprogramming with cultured inactive and activated fibroblasts. (B) It is unknown if differentiated myofibroblasts and matrifibrocytes can be reprogrammed into iCMs.
Figure 2.
Progresses and challenges of direct cardiac reprogramming in vitro. (A) Understanding the mechanism of iCM reprogramming with cultured inactive and activated fibroblasts. (B) It is unknown if differentiated myofibroblasts and matrifibrocytes can be reprogrammed into iCMs.
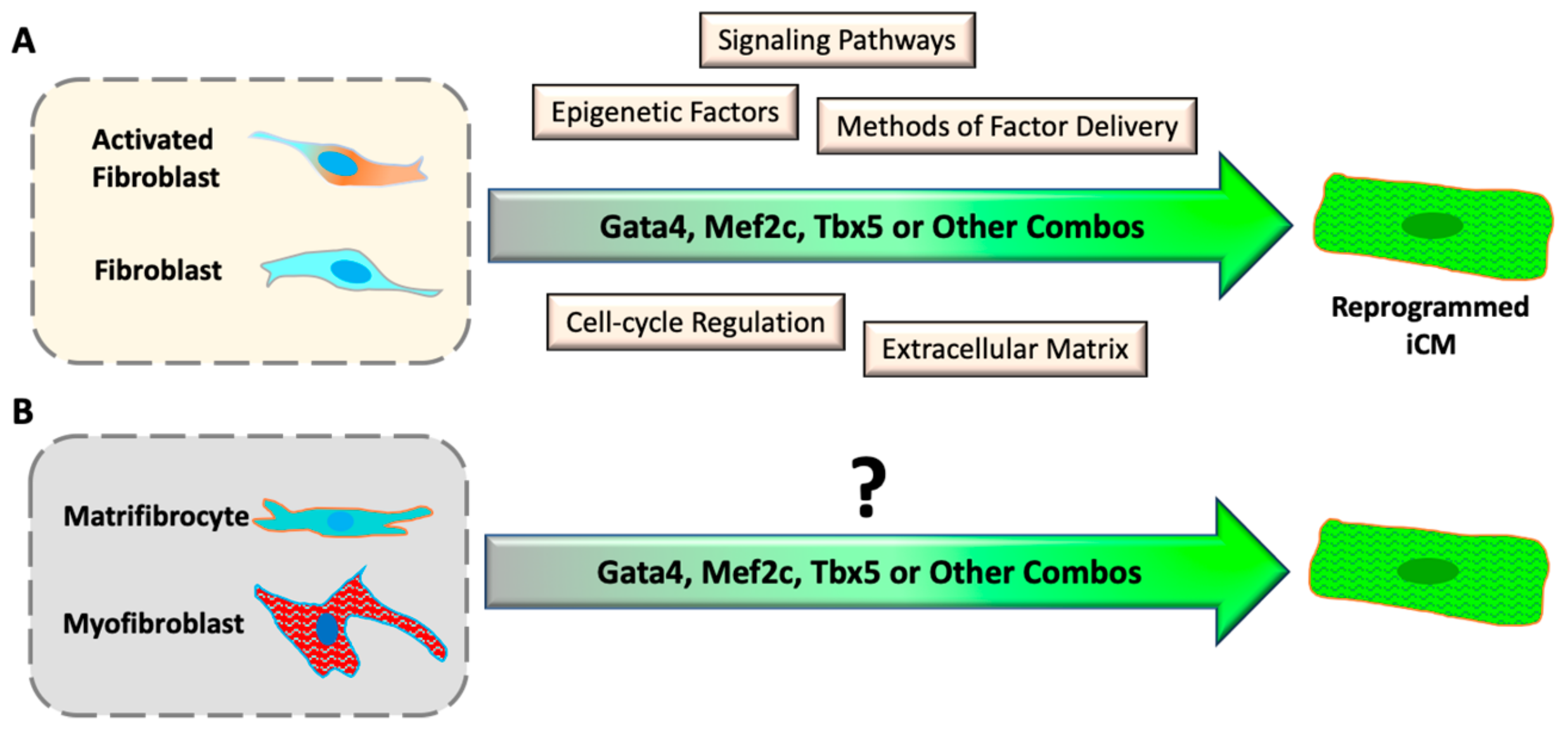
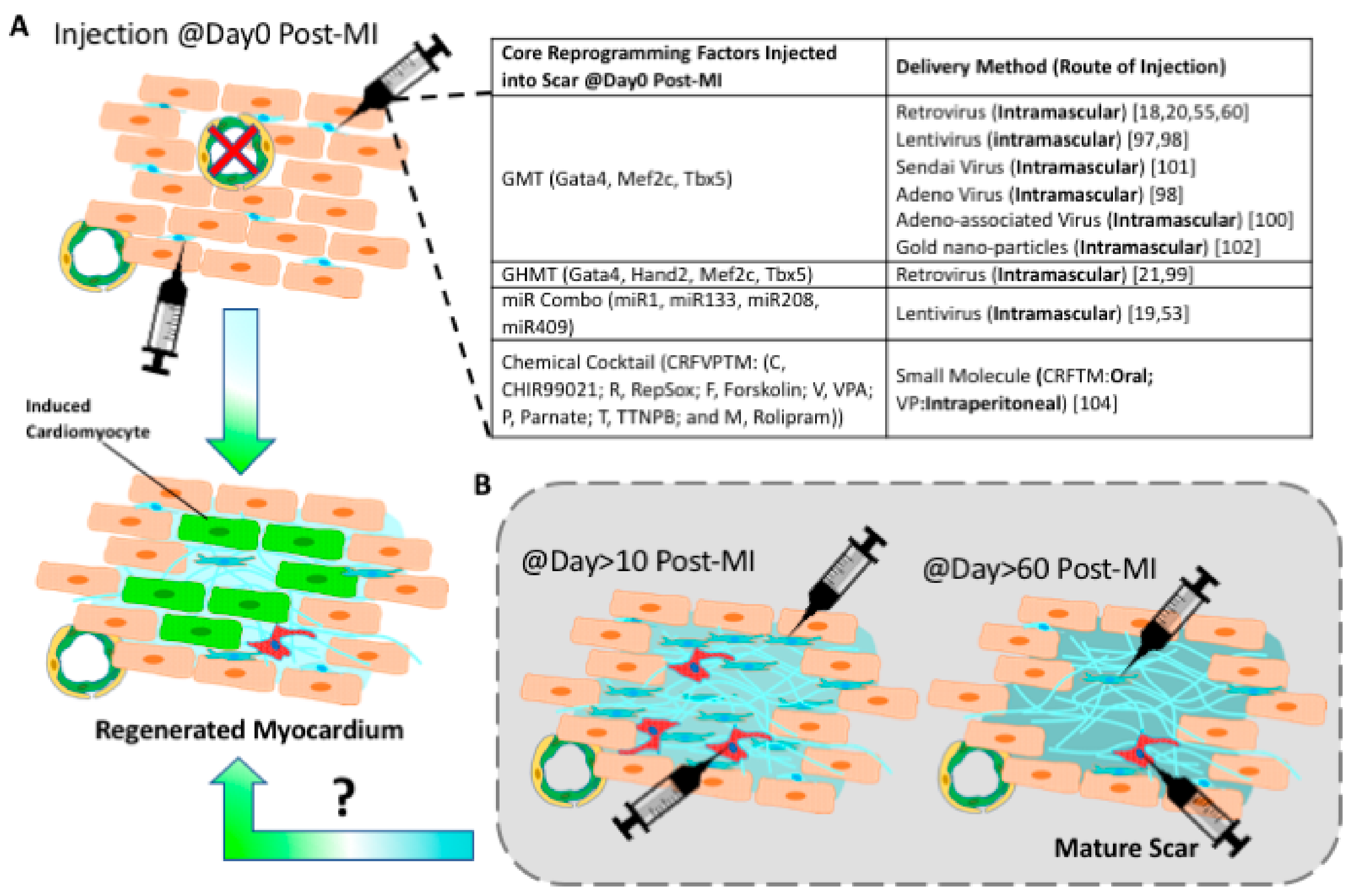
Figure 3.
Progresses and challenges of in vivo reprogramming. (A) Cardiac reprogramming factors were all delivered right after acute myocardial infarction. The inserted table summarizes in vivo reprogramming studies that use various viral or chemical cocktails [18,19,20,21,53,55,60,97,98,99,100,101,102,104]. (B) It is unknown if a cardiac chronic scar can be reprogrammed into induced cardiac muscle tissue.
Figure 3.
Progresses and challenges of in vivo reprogramming. (A) Cardiac reprogramming factors were all delivered right after acute myocardial infarction. The inserted table summarizes in vivo reprogramming studies that use various viral or chemical cocktails [18,19,20,21,53,55,60,97,98,99,100,101,102,104]. (B) It is unknown if a cardiac chronic scar can be reprogrammed into induced cardiac muscle tissue.


{kind=link}
{kind=link}
{kind=link}
Table 1.
A summary of the mechanistic understanding of recent publications on mouse and human iCM reprogramming.
Table 1.
A summary of the mechanistic understanding of recent publications on mouse and human iCM reprogramming.
| Signaling Pathways | Mouse iCM Reprogramming | Human iCM Reprogramming |
|---|---|---|
| TGFβ Inhibition | Enhanced by suppression of Smad2 and 3 phosphorylation [64,65] | Enhanced [60] |
| Akt1 Activation | Enhanced through activation of mTORC1 and Foxo3a in embryonic but not in adult fibroblasts [68] | Unknown |
| JAK inhibition | Enhanced iCM quality [19] | Unknown |
| RhoA-ROCK Inhibition | Enhanced through suppression of SRF-signaling [65] | Unknown |
| Notch Inhibition | Enhanced through increased Mef2c activity [67] | Unknown |
| Wnt Inhibition | Enhanced through suppression of canonical Wnt [60] | Enhanced [60] |
| FFV (Fgf2, FGF10, VEGF) | Enhanced through activation of p38 MAPK and PI3K/Akt pathways [69] | Unknown |
| Inflammatory Signaling Pathways | ||
| Cyclooxygenase-2 Inhibition (by diclofenac) | Improved iCM quality through suppression of E2/PGE R4, cAMP/PKA, and IL1β [72] | Unknown |
| C-C chemokine inhibition | Enhanced by suppression of chemokine receptors [71] | Unknown |
| Transcriptional Regulators | ||
| ZNF281 Activation | Enhanced by cooperation with Gata4 and suppression of inflammatory response [70] | Unknown |
| ZFPM2 Activation | Unknown | Enhanced [58] |
| Epigenetic Factors | ||
| Bmi1 Inhibition | Improved induction of beating iCMs [80] | Unknown |
| Ezh2 Inhibition | Enhanced by suppression of H3K27me2 & H3K27me3 [78,79] | Unknown |
| G9a and GLP Inhibition | Enhanced by suppression of H3K9me & H3K9me2 [78] | Unknown |
| Topological Factors | ||
| Microgroove ECM | Improved through nuclear localization of Mkl1 [91] | Unknown |
| 3D Hydrogels | Improved by increased expression of MMPs [92] | Unknown |
| Cell-cycle Manipulations | ||
| S-phase Synchronization | Accelerated by enhancing cell-cycle exit [66] | Unknown |
| G2/M-phase Synchronization | Improved iCM quality and yield [56] | Unknown |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Bektik, E.; Fu, J.-d. Ameliorating the Fibrotic Remodeling of the Heart through Direct Cardiac Reprogramming. Cells 2019, 8, 679. https://doi.org/10.3390/cells8070679
AMA Style
Bektik E, Fu J-d. Ameliorating the Fibrotic Remodeling of the Heart through Direct Cardiac Reprogramming. Cells. 2019; 8(7):679. https://doi.org/10.3390/cells8070679
Chicago/Turabian StyleBektik, Emre, and Ji-dong Fu. 2019. "Ameliorating the Fibrotic Remodeling of the Heart through Direct Cardiac Reprogramming" Cells 8, no. 7: 679. https://doi.org/10.3390/cells8070679
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.