S. cerevisiae Strain Lacking Mitochondrial IF3 Shows Increased Levels of Tma19p during Adaptation to Respiratory Growth

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains, Media, and Growth Conditions
2.2. Mitochondria Isolation
2.3. Cytochrome c Oxidase Activity Assay
2.4. Blue Native Polyacrylamide Gel Electrophoresis (PAGE)
2.5. 2D Differential Proteomic Analysis (2D-DIGE)
2.6. Growth Curves
3. Results
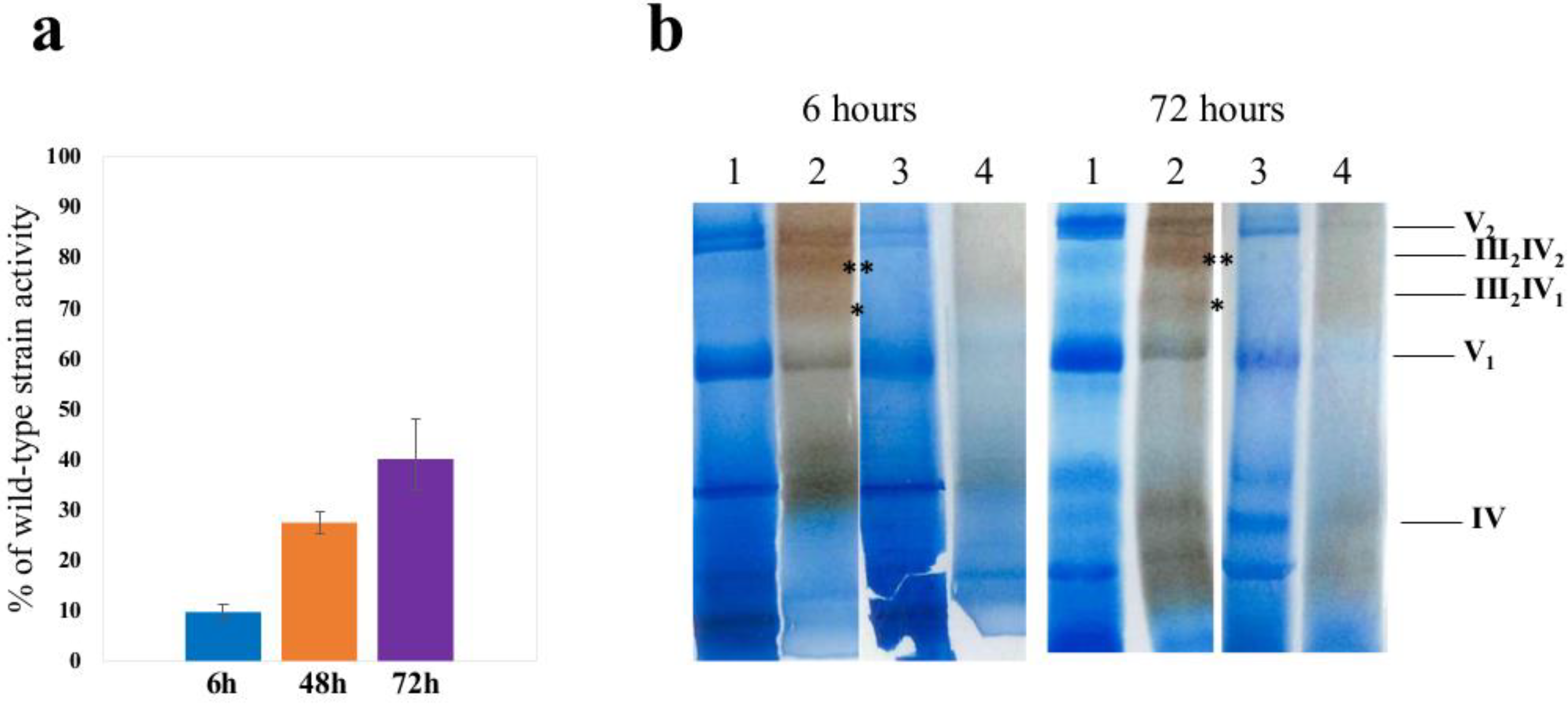
3.1. Adaptation of aim23Δ Yeast Strain to Respiratory Growth Does Not Completely Restore Cytochrome Oxidase Activity
3.2. Deletion of AIM23 Completely Prevents Supercomplexes Formation, Which Does Not Restore after Adaptation to Respiratory Growth
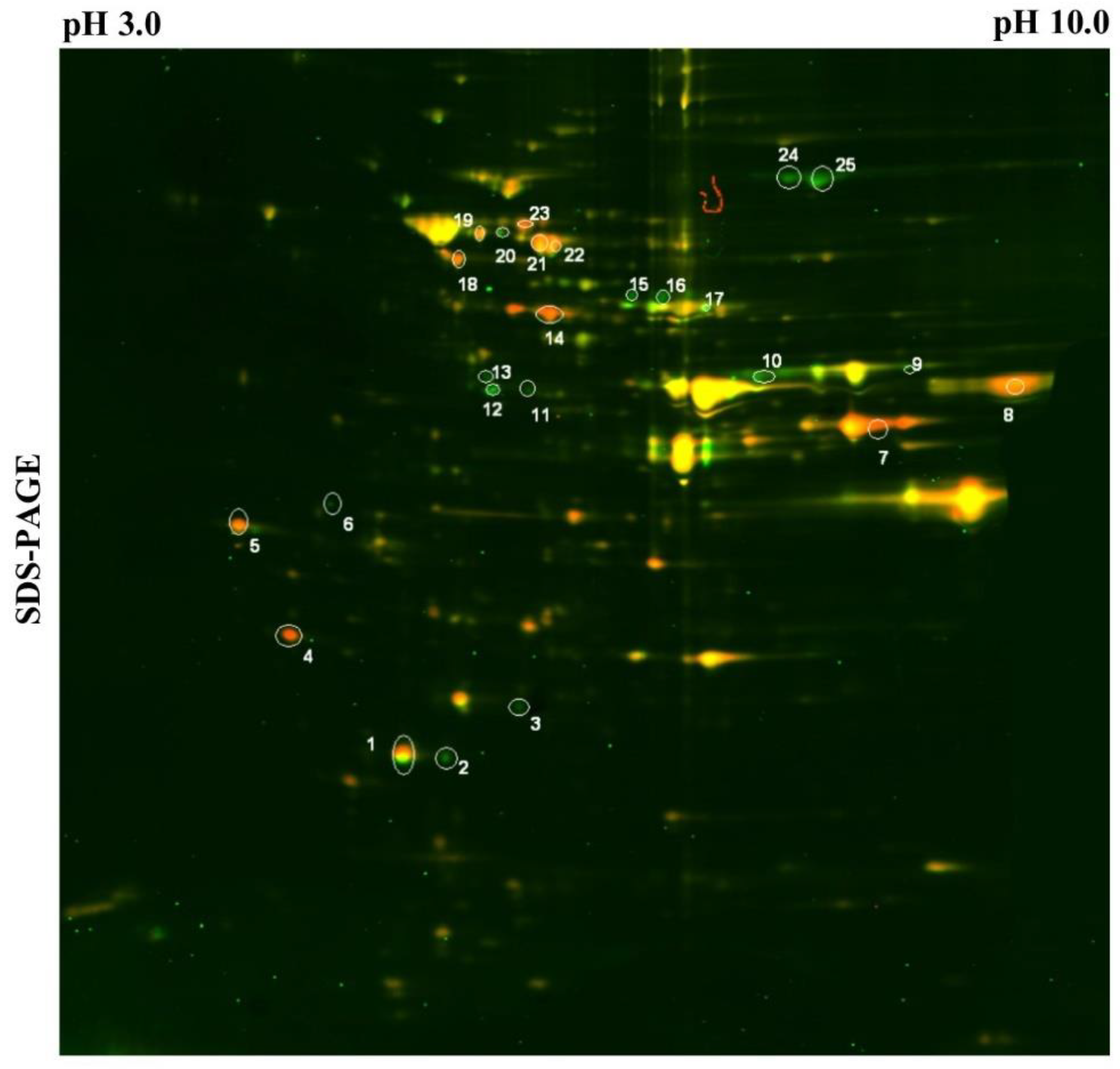
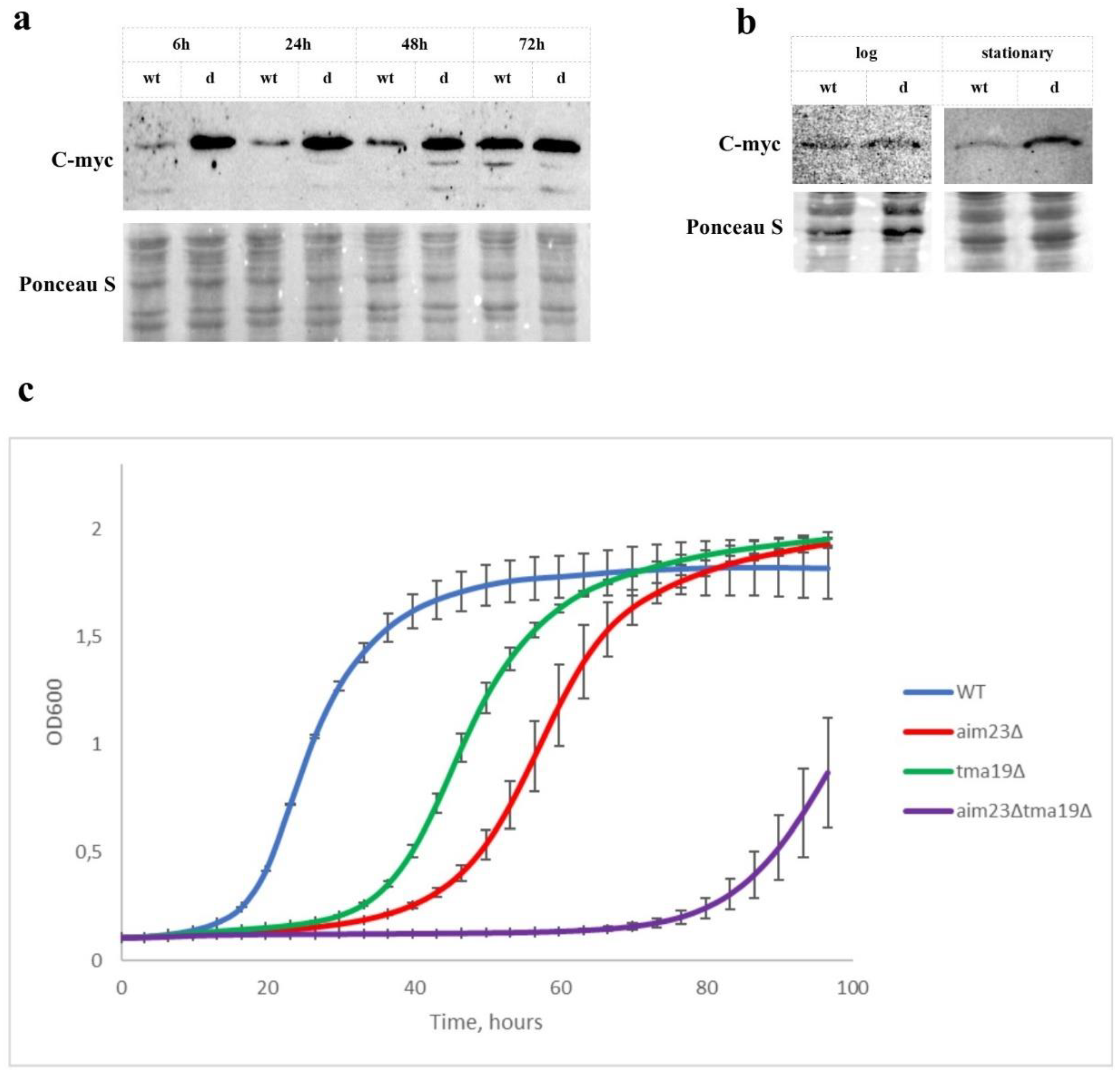
3.3. Tma19p is Overproduced During Adaptation of Aim23p-Deficient Yeasts and is Indispensable for Its Respiratory Growth
3.4. Deletion of TMA19 Gene is Suppressed by Addition of Antioxidants in Both tma19Δ and aim23Δtma19Δ Strains
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Martin, W.F.; Garg, S.; Zimorski, V. Endosymbiotic theories for eukaryote origin. Philos. Trans. R. Soc. B Biol. Sci. 2015, 370, 20140330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hansen, K.G.; Herrmann, J.M. Transport of Proteins into Mitochondria. Protein J. 2019, 38, 330–342. [Google Scholar] [CrossRef] [PubMed]
- Kolesnikov, A.A.; Gerasimov, E.S. Diversity of mitochondrial genome organization. Biochem. Mosc. 2012, 77, 1424–1435. [Google Scholar] [CrossRef] [PubMed]
- Chacinska, A.; Koehler, C.M.; Milenkovic, D.; Lithgow, T.; Pfanner, N. Importing Mitochondrial Proteins: Machineries and Mechanisms. Cell 2009, 138, 628–644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ott, M.; Amunts, A.; Brown, A. Organization and Regulation of Mitochondrial Protein Synthesis. Annu. Rev. Biochem. 2016, 85, 77–101. [Google Scholar] [CrossRef] [PubMed]
- Gaur, R.; Grasso, D.; Datta, P.P.; Krishna, P.D.V; Das, G.; Spencer, A.; Agrawal, R.K.; Spremulli, L.; Varshney, U. A single mammalian mitochondrial translation initiation factor functionally replaces two bacterial factors. Mol. Cell 2008, 29, 180–190. [Google Scholar] [CrossRef] [PubMed]
- Kuzmenko, A.V.; Levitskii, S.A.; Vinogradova, E.N.; Atkinson, G.C.; Hauryliuk, V.; Zenkin, N.; Kamenski, P.A. Protein biosynthesis in mitochondria. Biochemistry 2013, 78, 855–866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atkinson, G.C.; Kuzmenko, A.; Kamenski, P.; Vysokikh, M.Y.; Lakunina, V.; Tankov, S.; Smirnova, E.; Soosaar, A.; Tenson, T.; Hauryliuk, V. Evolutionary and genetic analyses of mitochondrial translation initiation factors identify the missing mitochondrial IF3 in S. cerevisiae. Nucleic Acids Res. 2012, 40, 6122–6134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuzmenko, A.; Derbikova, K.; Salvatori, R.; Tankov, S.; Atkinson, G.C.; Tenson, T.; Ott, M.; Kamenski, P.; Hauryliuk, V. Aim-less translation: Loss of Saccharomyces cerevisiae mitochondrial translation initiation factor mIF3/Aim23 leads to unbalanced protein synthesis. Sci. Rep. 2016, 6, 18749. [Google Scholar] [CrossRef]
- Hegemann, J.H.; Güldener, U.; Köhler, G.J. Gene disruption in the budding yeast Saccharomyces cerevisiae. Methods Mol. Biol. 2006, 313, 129–144. [Google Scholar]
- Wach, A.; Brachat, A.; Pöhlmann, R.; Philippsen, P. New heterologous modules for classical or PCR-based gene disruptions in Saccharomyces cerevisiae. Yeast 1994, 10, 1793–1808. [Google Scholar] [CrossRef] [PubMed]
- Gietz, R.D.; Schiestl, R.H. Quick and easy yeast transformation using the LiAc/SS carrier DNA/PEG method. Nat. Protoc. 2007, 2, 35–37. [Google Scholar] [CrossRef] [PubMed]
- Janke, C.; Magiera, M.M.; Rathfelder, N.; Taxis, C.; Reber, S.; Maekawa, H.; Moreno-Borchart, A.; Doenges, G.; Schwob, E.; Schiebel, E.; et al. A versatile toolbox for PCR-based tagging of yeast genes: New fluorescent proteins, more markers and promoter substitution cassettes. Yeast 2004, 21, 947–962. [Google Scholar] [CrossRef] [PubMed]
- Gregg, C.; Kyryakov, P.; Titorenko, V.I. Purification of Mitochondria from Yeast Cells. J. Vis. Exp. 2009, 30, e1417. [Google Scholar] [CrossRef] [PubMed]
- Barrientos, A.; Fontanesi, F.; Díaz, F. Evaluation of the mitochondrial respiratory Chain and oxidative phosphorylation system using polarography and spectrophotometric enzyme assays. Curr. Protoc. Hum. Genet. 2009, 63, 19.3.1–19.3.14. [Google Scholar]
- Wittig, I.; Braun, H.-P.; Schägger, H. Blue native PAGE. Nat. Protoc. 2006, 1, 418–428. [Google Scholar] [CrossRef] [PubMed]
- Jha, P.; Wang, X.; Auwerx, J. Analysis of Mitochondrial Respiratory Chain Supercomplexes Using Blue Native Polyacrylamide Gel Electrophoresis (BN-PAGE). In Current Protocols in Mouse Biology; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2016; Volume 6, pp. 1–14. [Google Scholar]
- Meleady, P. Two-Dimensional Gel Electrophoresis and 2D-DIGE. In Methods in Molecular Biology; Springer: Clifton, NJ, USA, 2018; Volume 1664, pp. 3–14. [Google Scholar]
- Schägger, H.; Pfeiffer, K. Supercomplexes in the respiratory chains of yeast and mammalian mitochondria. EMBO J. 2000, 19, 1777–1783. [Google Scholar] [CrossRef] [Green Version]
- Rinnerthaler, M.; Jarolim, S.; Heeren, G.; Palle, E.; Perju, S.; Klinger, H.; Bogengruber, E.; Madeo, F.; Braun, R.J.; Breitenbach-Koller, L.; et al. MMI1 (YKL056c, TMA19), the yeast orthologue of the translationally controlled tumor protein (TCTP) has apoptotic functions and interacts with both microtubules and mitochondria. Biochim. Biophys. Acta Bioenerg. 2006, 1757, 631–638. [Google Scholar] [CrossRef] [Green Version]
- Rossignol, R.; Faustin, B.; Rocher, C.; Malgat, M.; Mazat, J.-P.; Letellier, T. Mitochondrial threshold effects. Biochem. J. 2003, 370, 751–762. [Google Scholar] [CrossRef] [Green Version]
- Davey, G.P.; Clark, J.B. Threshold Effects and Control of Oxidative Phosphorylation in Nonsynaptic Rat Brain Mitochondria. J. Neurochem. 2002, 66, 1617–1624. [Google Scholar] [CrossRef]
- Kobayashi, D.; Tokuda, T.; Sato, K.; Okanishi, H.; Nagayama, M.; Hirayama-Kurogi, M.; Ohtsuki, S.; Araki, N. Identification of a Specific Translational Machinery via TCTP-EF1A2 Interaction Regulating NF1-associated Tumor Growth by Affinity Purification and Data-independent Mass Spectrometry Acquisition (AP-DIA). Mol. Cell. Proteom. 2019, 18, 245–262. [Google Scholar] [CrossRef] [PubMed]
- Trancíková, A.; Weisová, P.; Kissová, I.; Zeman, I.; Kolarov, J. Production of reactive oxygen species and loss of viability in yeast mitochondrial mutants: Protective effect of Bcl-xL. FEMS Yeast Res. 2004, 5, 149–156. [Google Scholar] [CrossRef] [PubMed]
- Boy-Marcotte, E.; Perrot, M.; Bussereau, F.; Boucherie, H.; Jacquet, M. Msn2p and Msn4p control a large number of genes induced at the diauxic transition which are repressed by cyclic AMP in Saccharomyces cerevisiae. J. Bacteriol. 1998, 180, 1044–1052. [Google Scholar] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Spot Number | aim23Δ Versus WT Ratio | Protein Name | Spot Number | aim23Δ versus WT Ratio | Protein Name |
|---|---|---|---|---|---|
| 1 | 2.06 | Ahp1p | 14 | 4.65 | Hxk1p |
| 2 | 0.39 | NI * | 15 | 0.47 | Ino1p |
| 3 | 0.27 | Pst2p | 16 | 0.87 | Ino1p |
| 4 | 11.06 | Tma19p | 17 | 0.78 | Ino1p |
| 5 | 28.92 | Efb1p | 18 | 2.47 | Hsp60p |
| 6 | 0.38 | Tpm1p | 19 | 3.35 | Vma1p |
| 7 | 3.58 | Adh1p | 20 | 0.65 | Vma1p |
| 8 | 1.44 | Pgk1p | 21 | 2.32 | Ssb1p |
| 9 | 0.38 | Erg13p | 22 | 3.1 | Ssb1p |
| 10 | 0.56 | Eno1p | 23 | 2.78 | NI * |
| 11 | 0.55 | Gpd1p | 24 | 0.39 | Met6p |
| 12 | 0.22 | Sam2p | 25 | 0.35 | Met6p |
| 13 | 0.26 | Lys9p |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Levitskii, S.; Baleva, M.V.; Chicherin, I.; Krasheninnikov, I.A.; Kamenski, P. S. cerevisiae Strain Lacking Mitochondrial IF3 Shows Increased Levels of Tma19p during Adaptation to Respiratory Growth. Cells 2019, 8, 645. https://doi.org/10.3390/cells8070645
Levitskii S, Baleva MV, Chicherin I, Krasheninnikov IA, Kamenski P. S. cerevisiae Strain Lacking Mitochondrial IF3 Shows Increased Levels of Tma19p during Adaptation to Respiratory Growth. Cells. 2019; 8(7):645. https://doi.org/10.3390/cells8070645
Chicago/Turabian StyleLevitskii, Sergey, Maria V. Baleva, Ivan Chicherin, Igor A. Krasheninnikov, and Piotr Kamenski. 2019. "S. cerevisiae Strain Lacking Mitochondrial IF3 Shows Increased Levels of Tma19p during Adaptation to Respiratory Growth" Cells 8, no. 7: 645. https://doi.org/10.3390/cells8070645
APA StyleLevitskii, S., Baleva, M. V., Chicherin, I., Krasheninnikov, I. A., & Kamenski, P. (2019). S. cerevisiae Strain Lacking Mitochondrial IF3 Shows Increased Levels of Tma19p during Adaptation to Respiratory Growth. Cells, 8(7), 645. https://doi.org/10.3390/cells8070645