Transcriptomic Response to 1,25-Dihydroxyvitamin D in Human Fibroblasts with or without a Functional Vitamin D Receptor (VDR): Novel Target Genes and Insights into VDR Basal Transcriptional Activity


Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents and Cell Culture
2.2. Immunocytochemistry
2.3. Microarray Analysis of Global Gene Expression
2.4. Validation by Quantitative RT-PCR
2.5. Cell Proliferation Assays
3. Results
3.1. Transcriptomic Response to 1,25D in Human Fibroblasts
3.2. Comparative Analysis of Transcriptome Profiling in CO and MUT Fibroblasts
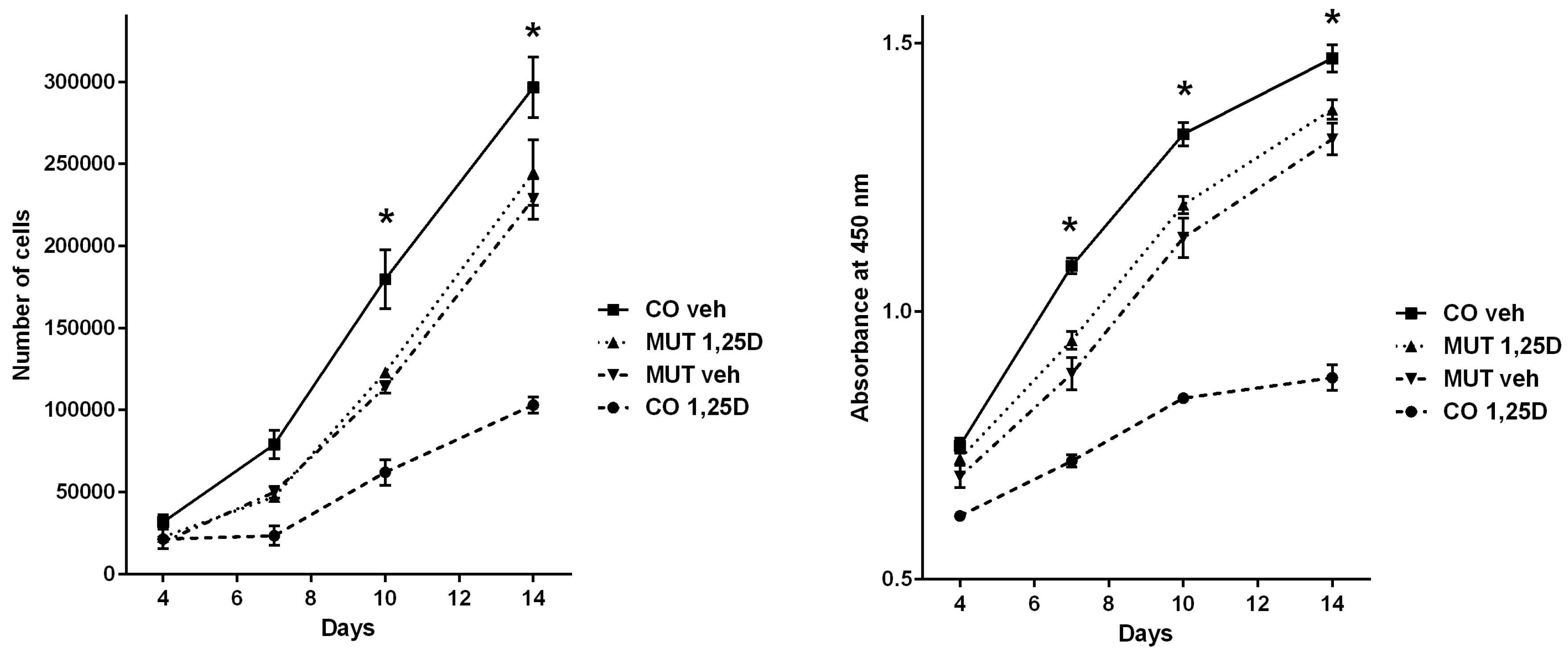
3.3. Analyses of Cell Proliferation Behaviour Corroborate Opposing Regulation by the VDR
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bouillon, R.; Carmeliet, G.; Verlinden, L.; van Etten, E.; Verstuyf, A.; Luderer, H.F.; Lieben, L.; Mathieu, C.; Demay, M. Vitamin D and human health: Lessons from vitamin D receptor null mice. Endocr. Rev. 2008, 29, 726–776. [Google Scholar] [CrossRef] [PubMed]
- Christakos, S.; Dhawan, P.; Verstuyf, A.; Verlinden, L.; Carmeliet, G. Vitamin D: Metabolism, Molecular Mechanism of Action, and Pleiotropic Effects. Physiol. Rev. 2016, 96, 365–408. [Google Scholar] [CrossRef] [PubMed]
- Mangelsdorf, D.J.; Thummel, C.; Beato, M.; Herrlich, P.; Schutz, G.; Umesono, K.; Blumberg, B.; Kastner, P.; Mark, M.; Chambon, P.; et al. The nuclear receptor superfamily: The second decade. Cell 1995, 83, 835–839. [Google Scholar] [CrossRef]
- Pike, J.W. Genome-wide principles of gene regulation by the vitamin D receptor and its activating ligand. Mol. Cell. Endocrinol. 2011, 347, 3–10. [Google Scholar] [CrossRef]
- Haussler, M.R.; Whitfield, G.K.; Kaneko, I.; Haussler, C.A.; Hsieh, D.; Hsieh, J.C.; Jurutka, P.W. Molecular mechanisms of vitamin D action. Calcif. Tissue Int. 2012, 92, 77–98. [Google Scholar] [CrossRef] [PubMed]
- Long, M.D.; Sucheston-Campbell, L.E.; Campbell, M.J. Vitamin D receptor and RXR in the post-genomic era. J. Cell. Physiol. 2015, 230, 758–766. [Google Scholar] [CrossRef] [PubMed]
- Meyer, M.B.; Watanuki, M.; Kim, S.; Shevde, N.K.; Pike, J.W. The human transient receptor potential vanilloid type 6 distal promoter contains multiple vitamin D receptor binding sites that mediate activation by 1,25-dihydroxyvitamin D3 in intestinal cells. Mol. Endocrinol. 2006, 20, 1447–1461. [Google Scholar] [CrossRef]
- Anderson, P.H.; Turner, A.G.; Morris, H.A. Vitamin D actions to regulate calcium and skeletal homeostasis. Clin. Biochem. 2012, 45, 880–886. [Google Scholar] [CrossRef] [PubMed]
- Adams, J.S.; Hewison, M. Extrarenal expression of the 25-hydroxyvitamin D-1-hydroxylase. Arch. Biochem. Biophys. 2012, 523, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Rosen, C.J.; Adams, J.S.; Bikle, D.D.; Black, D.M.; Demay, M.B.; Manson, J.E.; Murad, M.H.; Kovacs, C.S. The nonskeletal effects of vitamin D: An Endocrine Society scientific statement. Endocr. Rev. 2012, 33, 456–492. [Google Scholar] [CrossRef] [PubMed]
- Girgis, C.M.; Baldock, P.A.; Downes, M. Vitamin D, muscle and bone: Integrating effects in development, aging and injury. Mol. Cell. Endocrinol. 2015, 410, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Bikle, D.D.; Oda, Y.; Tu, C.L.; Jiang, Y. Novel mechanisms for the vitamin D receptor (VDR) in the skin and in skin cancer. J. Steroid Biochem. Mol. Biol. 2015, 148, 47–51. [Google Scholar] [CrossRef] [PubMed]
- Lazar, M.A. Thyroid hormone action: A binding contract. J. Clin. Investig. 2003, 112, 497–499. [Google Scholar] [CrossRef] [PubMed]
- Fuleihan Gel, H.; Bouillon, R.; Clarke, B.; Chakhtoura, M.; Cooper, C.; McClung, M.; Singh, R.J. Serum 25-Hydroxyvitamin D Levels: Variability, Knowledge Gaps, and the Concept of a Desirable Range. J. Bone Miner. Res. 2015, 30, 1119–1133. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, A.V.; Peehl, D.M.; Feldman, D. Inhibition of prostate cancer growth by vitamin D: Regulation of target gene expression. J. Cell. Biochem. 2003, 88, 363–371. [Google Scholar] [CrossRef] [PubMed]
- Eelen, G.; Verlinden, L.; van Camp, M.; van Hummelen, P.; Marchal, K.; de Moor, B.; Mathieu, C.; Carmeliet, G.; Bouillon, R.; Verstuyf, A. The effects of 1alpha,25-dihydroxyvitamin D3 on the expression of DNA replication genes. J. Bone Miner. Res. 2004, 19, 133–146. [Google Scholar] [CrossRef]
- Wang, T.T.; Tavera-Mendoza, L.E.; Laperriere, D.; Libby, E.; MacLeod, N.B.; Nagai, Y.; Bourdeau, V.; Konstorum, A.; Lallemant, B.; Zhang, R.; et al. Large-scale in silico and microarray-based identification of direct 1,25-dihydroxyvitamin D3 target genes. Mol. Endocrinol. 2005, 19, 2685–2695. [Google Scholar] [CrossRef] [PubMed]
- Lisse, T.S.; Chun, R.F.; Rieger, S.; Adams, J.S.; Hewison, M. Vitamin D activation of functionally distinct regulatory miRNAs in primary human osteoblasts. J. Bone Miner. Res. 2013, 28, 1478–1488. [Google Scholar] [CrossRef]
- Malloy, P.J.; Tasic, V.; Taha, D.; Tutunculer, F.; Ying, G.S.; Yin, L.K.; Wang, J.; Feldman, D. Vitamin D receptor mutations in patients with hereditary 1,25-dihydroxyvitamin D-resistant rickets. Mol. Genet. Metab. 2014, 111, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Mechica, J.B.; Leite, M.O.; Mendonca, B.B.; Frazzatto, E.S.; Borelli, A.; Latronico, A.C. A novel nonsense mutation in the first zinc finger of the vitamin D receptor causing hereditary 1,25-dihydroxyvitamin D3-resistant rickets. J. Clin. Endocrinol. Metab. 1997, 82, 3892–3894. [Google Scholar] [CrossRef]
- Damiani, F.M.; Martin, R.M.; Latronico, A.C.; Ferraz-de-Souza, B. Normal bone mass and normocalcemia in adulthood despite homozygous vitamin D receptor mutations. Osteoporos. Int. 2015, 26, 1819–1823. [Google Scholar] [CrossRef] [PubMed]
- Costa, P.L.; França, M.M.; Ferraz-de-Souza, B. Nonspecific binding of a frequently used vitamin D receptor (VDR) antibody: Important implications for vitamin D research in human health. Endocrine 2016, 54, 556–559. [Google Scholar] [CrossRef] [PubMed]
- Huang da, W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef] [PubMed]
- Pike, J.W.; Meyer, M.B. Fundamentals of vitamin D hormone-regulated gene expression. J. Steroid Biochem. Mol. Biol. 2014, 144 Pt A, 5–11. [Google Scholar] [CrossRef]
- LaPorta, E.; Welsh, J. Modeling vitamin D actions in triple negative/basal-like breast cancer. J. Steroid Biochem. Mol. Biol. 2014, 144 Pt A, 65–73. [Google Scholar] [CrossRef]
- Pike, J.W.; Meyer, M.B. Regulation of mouse Cyp24a1 expression via promoter-proximal and downstream-distal enhancers highlights new concepts of 1,25-dihydroxyvitamin D(3) action. Arch. Biochem. Biophys. 2012, 523, 2–8. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Craig, T.A.; Lutz, W.H.; Kumar, R. Identification of 1 alpha,25-dihydroxyvitamin D3 response elements in the human transforming growth factor beta 2 gene. Biochemistry 1999, 38, 2654–2660. [Google Scholar] [CrossRef] [PubMed]
- Campos, L.T.; Brentani, H.; Roela, R.A.; Katayama, M.L.; Lima, L.; Rolim, C.F.; Milani, C.; Folgueira, M.A.; Brentani, M.M. Differences in transcriptional effects of 1alpha,25 dihydroxyvitamin D3 on fibroblasts associated to breast carcinomas and from paired normal breast tissues. J. Steroid Biochem. Mol. Biol. 2013, 133, 12–24. [Google Scholar] [CrossRef]
- Neme, A.; Nurminen, V.; Seuter, S.; Carlberg, C. The vitamin D-dependent transcriptome of human monocytes. J. Steroid Biochem. Mol. Biol. 2016, 164, 180–187. [Google Scholar] [CrossRef]
- Bikle, D.D. Vitamin D metabolism and function in the skin. Mol. Cell. Endocrinol 2011, 347, 80–89. [Google Scholar] [CrossRef]
- Lisse, T.S.; Saini, V.; Zhao, H.; Luderer, H.F.; Gori, F.; Demay, M.B. The vitamin D receptor is required for activation of cWnt and hedgehog signaling in keratinocytes. Mol. Endocrinol. 2014, 28, 1698–1706. [Google Scholar] [CrossRef] [PubMed]
- Malloy, P.J.; Feldman, B.J. Cell-autonomous regulation of brown fat identity gene UCP1 by unliganded vitamin D receptor. Mol. Endocrinol. 2013, 27, 1632–1642. [Google Scholar] [CrossRef] [PubMed]
- Mangelsdorf, D.J.; Evans, R.M. The RXR heterodimers and orphan receptors. Cell 1995, 83, 841–850. [Google Scholar] [CrossRef]
- Bernal, J.; Morte, B. Thyroid hormone receptor activity in the absence of ligand: Physiological and developmental implications. Biochim. Biophys. Acta 2013, 1830, 3893–3899. [Google Scholar] [CrossRef] [PubMed]
- Alimirah, F.; Vaishnav, A.; McCormick, M.; Echchgadda, I.; Chatterjee, B.; Mehta, R.G.; Peng, X. Functionality of unliganded VDR in breast cancer cells: Repressive action on CYP24 basal transcription. Mol. Cell. Biochem. 2010, 342, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.M.; Pike, J.W. The vitamin D receptor functions as a transcription regulator in the absence of 1,25-dihydroxyvitamin D. J. Steroid Biochem. Mol. Biol. 2015. [Google Scholar] [CrossRef] [PubMed]
- Milani, C.; Katayama, M.L.; de Lyra, E.C.; Welsh, J.; Campos, L.T.; Brentani, M.M.; Maciel Mdo, S.; Roela, R.A.; del Valle, P.R.; Goes, J.C.; et al. Transcriptional effects of 1,25 dihydroxyvitamin D(3) physiological and supra-physiological concentrations in breast cancer organotypic culture. BMC Cancer 2013, 13, 119. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Gene Symbol | Transcript Cluster ID 1 | Fold Change | Adjusted p-Value 2 |
|---|---|---|---|
| Up-regulated by 1,25D | |||
| CYP24A1 | 16920497 | 87.7 | 4.8 × 10−16 |
| TGFB2 | 16677556 | 5.6 | 4.4 × 10−9 |
| RASSF2 | 16916901 | 4.0 | 1.2 × 10−10 |
| CILP | 16810713 | 3.9 | 9.1 × 10−12 |
| OSR2 | 17071298 | 3.9 | 1.2 × 10−9 |
| STC1 | 17075553 | 3.8 | 1.4 × 10−9 |
| RGNEF | 16986138 | 3.5 | 5.3 × 10−12 |
| NID2 | 16792954 | 3.3 | 2.8 × 10−9 |
| EFTUD1 | 16812475 | 3.3 | 1.5 × 10−9 |
| MREG | 16908138 | 3.1 | 8.9 × 10−9 |
| Down-regulated by 1,25D | |||
| KRTAP1-1 | 16844572 | −1.9 | 5.3 × 10−6 |
| LCE1F | 16671065 | −1.9 | 1.1 × 10−4 |
| PLK2 | 16996433 | −1.9 | 5.3 × 10−6 |
| LGR5 | 16754134 | −1.9 | 1.2 × 10−4 |
| EFNB2 | 16780859 | −1.9 | 1.0 × 10−3 |
| TRNAI2 | 17005782 | −2.2 | 3.8 × 10−5 |
| B3GALT2 | 16697471 | −2.2 | 3.5 × 10−4 |
| C11orf87 | 16730967 | −2.3 | 1.0 × 10−8 |
| LOC730755 | 16844585 | −2.3 | 9.8 × 10−11 |
| GALNT12 | 17087413 | −2.7 | 1.3 × 10−8 |
| Target Gene | Differential Expression | Microarray | qRT-PCR TaqMan | ||
|---|---|---|---|---|---|
| FC | p-Value | RQ | RQ Min-Max | ||
| CYP24A1 | CO 1,25D versus Veh up-reg | 87.7 | 10−16 | 1032.0 | 852.7–1249.2 |
| TGFB2 | CO 1,25D versus Veh up-reg | 5.6 | 10−9 | 11.8 | 9.6–14.5 |
| RASSF2 | CO 1,25D versus Veh up-reg | 4.0 | 10−10 | 9.0 | 6.2–13.0 |
| CILP | CO 1,25D versus Veh up-reg | 3.9 | 10−12 | 247.0 | 201.8–302.3 |
| STC1 | CO 1,25D versus Veh up-reg | 3.8 | 10−9 | 8.2 | 6.6–10.1 |
| HBEGF | CO 1,25D versus Veh down-reg | −1.7 | 10−5 | 0.44 | 0.33–0.58 |
| EFNB2 | CO 1,25D versus Veh down-reg | −1.9 | 10−3 | 0.61 | 0.43–0.85 |
| GALNT12 | CO 1,25D versus Veh down-reg | −2.7 | 10−8 | 0.36 | 0.24–0.53 |
| EGR1 | MUT 1,25D versus Veh up-reg | 2.7 | 10−7 | 3.9 | 3.6–4.4 |
| Target Gene | 1,25D-Activated VDR Regulation (CO 1,25D vs. CO veh) | Putative Unliganded VDR Regulation (CO Veh vs. MUT Veh) | ||
|---|---|---|---|---|
| Fold Change | Adj p-Value | Fold Change | Adj p-Value | |
| SERPINB7 | +2.7 | 3.1 × 10−7 | −1.9 | 4.7 × 10−5 |
| CCR1 | +2.1 | 3.6 × 10−6 | −1.5 | 1.6 × 10−3 |
| SPP1 | +2.0 | 3.7 × 10−7 | −1.7 | 7.7 × 10−6 |
| MIR548V | +1.9 | 1.2 × 10−4 | −1.7 | 9.9 × 10−4 |
| SULF2 | +1.8 | 2.7 × 10−7 | −1.8 | 3.1 × 10−7 |
| BMP6 | +1.7 | 3.0 × 10−5 | −2.4 | 7.7 × 10−7 |
| DNER | +1.7 | 7.4 × 10−7 | −1.4 | 6.0 × 10−5 |
| APBB1IP | +1.7 | 2.7 × 10−10 | −1.7 | 6.7 × 10−10 |
| SGIP1 | +1.5 | 1.4 × 10−4 | −1.4 | 1.7 × 10−3 |
| ANGPTL4 | +1.5 | 6.6 × 10−7 | −1.7 | 1.2 × 10−7 |
| DKK2 | −1.5 | 6.5 × 10−4 | +1.5 | 2.5 × 10−3 |
| HOXB5 | −1.5 | 7.3 × 10−5 | +1.7 | 1.9 × 10−5 |
| PITPNM3 | −1.6 | 1.9 × 10−5 | +1.5 | 2.9 × 10−4 |
| PDGFC | −1.6 | 1.9 × 10−8 | +1.9 | 1.6 × 10−9 |
| SPTLC3 | −1.7 | 3.3 × 10−7 | +1.7 | 1.1 × 10−6 |
| SNORA76 | −1.7 | 2.4 × 10−4 | +2.1 | 2.7 × 10−5 |
| CH25H | −1.8 | 1.3 × 10−4 | +2.0 | 3.6 × 10−5 |
| MIR4659A | −1.8 | 4.1 × 10−4 | +1.7 | 8.4 × 10−4 |
| LGR5 | −1.9 | 1.2 × 10−4 | +2.9 | 3.4 × 10−6 |
| EFNB2 | −1.9 | 1.0 × 10−3 | +1.9 | 1.8 × 10−3 |
| Target Gene Population | Direction of Regulation | Gene Ontology Enrichment (n of Targets Involved in Process) | DAVID p-Value |
|---|---|---|---|
| 1,25D-activated VDR targets | Up-reg | skeletal system development (n = 21) | 10−5 |
| Up-reg | steroid metabolic process (n = 16) | 10−5 | |
| Down-reg | cell cycle (n = 65) | 10−8 | |
| Down-reg | ribonucleotide binding (n = 110) | 10−6 | |
| Putative basal unligandedVDR targets | Up-reg | DNA repair (n = 77) | 10−23 |
| Up-reg | DNA replication (n = 62) | 10−23 | |
| Up-reg | mRNA processing (n = 65) | 10−13 | |
| Down-reg | endocytosis & membrane invagination (n = 19) | 10−5 | |
| Down-reg | cell adhesion (n = 37) | 10−4 | |
| Combined basal and 1,25D-dependent VDR targets | Opposing regulation | cell cycle (n = 34) | 10−8 |
| DNA repair and replication (n = 17) | 10−7 | ||
| nucleotide binding (n = 52) | 10−5 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Costa, P.L.F.; França, M.M.; Katayama, M.L.; Carneiro, E.T.; Martin, R.M.; Folgueira, M.A.K.; Latronico, A.C.; Ferraz-de-Souza, B. Transcriptomic Response to 1,25-Dihydroxyvitamin D in Human Fibroblasts with or without a Functional Vitamin D Receptor (VDR): Novel Target Genes and Insights into VDR Basal Transcriptional Activity. Cells 2019, 8, 318. https://doi.org/10.3390/cells8040318
Costa PLF, França MM, Katayama ML, Carneiro ET, Martin RM, Folgueira MAK, Latronico AC, Ferraz-de-Souza B. Transcriptomic Response to 1,25-Dihydroxyvitamin D in Human Fibroblasts with or without a Functional Vitamin D Receptor (VDR): Novel Target Genes and Insights into VDR Basal Transcriptional Activity. Cells. 2019; 8(4):318. https://doi.org/10.3390/cells8040318
Chicago/Turabian StyleCosta, Pedro L. F., Monica M. França, Maria L. Katayama, Eduardo T. Carneiro, Regina M. Martin, Maria A. K. Folgueira, Ana C. Latronico, and Bruno Ferraz-de-Souza. 2019. "Transcriptomic Response to 1,25-Dihydroxyvitamin D in Human Fibroblasts with or without a Functional Vitamin D Receptor (VDR): Novel Target Genes and Insights into VDR Basal Transcriptional Activity" Cells 8, no. 4: 318. https://doi.org/10.3390/cells8040318
APA StyleCosta, P. L. F., França, M. M., Katayama, M. L., Carneiro, E. T., Martin, R. M., Folgueira, M. A. K., Latronico, A. C., & Ferraz-de-Souza, B. (2019). Transcriptomic Response to 1,25-Dihydroxyvitamin D in Human Fibroblasts with or without a Functional Vitamin D Receptor (VDR): Novel Target Genes and Insights into VDR Basal Transcriptional Activity. Cells, 8(4), 318. https://doi.org/10.3390/cells8040318