No Causal Effect of Telomere Length on Ischemic Stroke and Its Subtypes: A Mendelian Randomization Study

 ,
,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Data Source and Single Nucleotide Polymorphism(SNP)Selection
2.2. Statistical Analysis
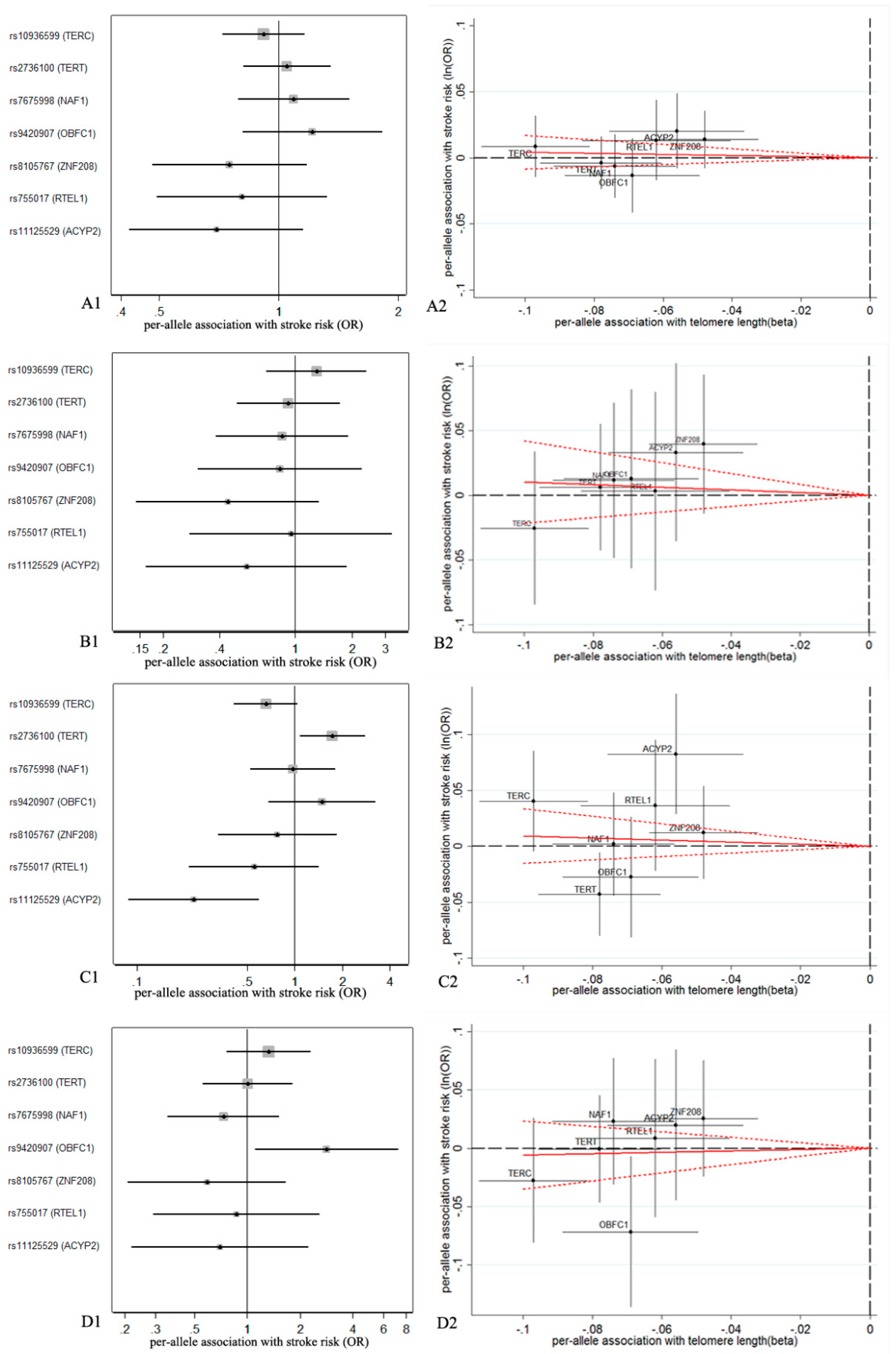
3. Results
3.1. Association of Genetic Variables with TL
3.2. Association of TL with IS
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pandian, J.D.; Gall, S.L.; Kate, M.P.; Silva, G.S.; Akinyemi, R.O.; Ovbiagele, B.I.; Lavados, P.M.; Gandhi, D.B.C.; Thrift, A.G. Prevention of stroke: A global perspective. Lancet 2018, 392, 1269–1278. [Google Scholar] [CrossRef]
- Hankey, G.J. Stroke. Lancet 2017, 389, 641–654. [Google Scholar] [CrossRef]
- Adams, H.P., Jr.; Bendixen, B.H.; Kappelle, L.J.; Biller, J.; Love, B.B.; Gordon, D.L.; Marsh, E.E., 3rd. Classification of subtype of acute ischemic stroke. Definitions for use in a multicenter clinical trial. TOAST. Trial of Org 10172 in Acute Stroke Treatment. Stroke 1993, 24, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Zhan, Y.; Liu, X.R.; Reynolds, C.A.; Pedersen, N.L.; Hagg, S.; Clements, M.S. Leukocyte telomere length and all-cause mortality: A between-within twin study with time-dependent effects using generalized survival models. Am. J. Epidemiol. 2018, 187, 2186–2191. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Zhan, Y.; Pedersen, N.L.; Fang, F.; Hagg, S. Telomere length and all-cause mortality: A meta-analysis. Ageing Res. Rev. 2018, 48, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Blackburn, E.H.; Epel, E.S.; Lin, J. Human telomere biology: A contributory and interactive factor in aging, disease risks, and protection. Science 2015, 350, 1193–1198. [Google Scholar] [CrossRef] [PubMed]
- Blackburn, E.H. Telomere states and cell fates. Nature 2000, 408, 53–56. [Google Scholar] [CrossRef]
- Laina, A.; Stellos, K.; Stamatelopoulos, K. Vascular ageing: Underlying mechanisms and clinical implications. Exp. Gerontol. 2018, 109, 16–30. [Google Scholar] [CrossRef]
- Yeh, J.K.; Wang, C.Y. Telomeres and telomerase in cardiovascular diseases. Genes (Basel) 2016, 7, 58. [Google Scholar] [CrossRef]
- Laurent, S.; Cockcroft, J.; Van Bortel, L.; Boutouyrie, P.; Giannattasio, C.; Hayoz, D.; Pannier, B.; Vlachopoulos, C.; Wilkinson, I.; Struijker-Boudier, H. Expert consensus document on arterial stiffness: Methodological issues and clinical applications. Eur. Heart J. 2006, 27, 2588–2605. [Google Scholar] [CrossRef]
- McDonnell, B.J.; Yasmin; Butcher, L.; Cockcroft, J.R.; Wilkinson, I.B.; Erusalimsky, J.D.; McEniery, C.M. The age-dependent association between aortic pulse wave velocity and telomere length. J. Physiol. 2017, 595, 1627–1635. [Google Scholar] [CrossRef] [PubMed]
- Amarenco, P.; Bogousslavsky, J.; Caplan, L.R.; Donnan, G.A.; Wolf, M.E.; Hennerici, M.G. The ASCOD phenotyping of ischemic stroke (Updated ASCO Phenotyping). Cerebrovasc. Dis. 2013, 36, 1–5. [Google Scholar] [CrossRef]
- Regenhardt, R.W.; Das, A.S.; Lo, E.H.; Caplan, L.R. Advances in understanding the pathophysiology of lacunar stroke: A review. JAMA Neurol. 2018, 75, 1273–1281. [Google Scholar] [CrossRef] [PubMed]
- Kamel, H.; Healey, J.S. Cardioembolic stroke. Circ. Res. 2017, 120, 514–526. [Google Scholar] [CrossRef]
- Zhang, W.; Chen, Y.; Wang, Y.; Liu, P.; Zhang, M.; Zhang, C.; Hu, F.B.; Hui, R. Short telomere length in blood leucocytes contributes to the presence of atherothrombotic stroke and haemorrhagic stroke and risk of post-stroke death. Clin. Sci. (Lond.) 2013, 125, 27–36. [Google Scholar] [CrossRef]
- Ding, H.; Chen, C.; Shaffer, J.R.; Liu, L.; Xu, Y.; Wang, X.; Hui, R.; Wang, D.W. Telomere length and risk of stroke in Chinese. Stroke 2012, 43, 658–663. [Google Scholar] [CrossRef] [PubMed]
- Schurks, M.; Prescott, J.; Dushkes, R.; De Vivo, I.; Rexrode, K.M. Telomere length and ischaemic stroke in women: A nested case-control study. Eur. J. Neurol. 2013, 20, 1068–1074. [Google Scholar] [CrossRef]
- Zee, R.Y.; Castonguay, A.J.; Barton, N.S.; Ridker, P.M. Relative leukocyte telomere length and risk of incident ischemic stroke in men: A prospective, nested case-control approach. Rejuvenation Res. 2010, 13, 411–414. [Google Scholar] [CrossRef]
- Sekula, P.; Del Greco, M.F.; Pattaro, C.; Kottgen, A. Mendelian randomization as an approach to assess causality using observational data. J. Am. Soc. Nephrol. 2016, 27, 3253–3265. [Google Scholar] [CrossRef]
- Hamad, R.; Walter, S.; Rehkopf, D.H. Telomere length and health outcomes: A two-sample genetic instrumental variables analysis. Exp. Gerontol. 2016, 82, 88–94. [Google Scholar] [CrossRef]
- Zhan, Y.; Karlsson, I.K.; Karlsson, R.; Tillander, A.; Reynolds, C.A.; Pedersen, N.L.; Hagg, S. Exploring the causal pathway from telomere length to coronary heart disease: A network mendelian randomization study. Circ. Res. 2017, 121, 214–219. [Google Scholar] [CrossRef] [PubMed]
- Larsson, S.C.; Scott, R.A.; Traylor, M.; Langenberg, C.C.; Hindy, G.; Melander, O.; Orho-Melander, M.; Seshadri, S.; Wareham, N.J.; Markus, H.S. Type 2 diabetes, glucose, insulin, BMI, and ischemic stroke subtypes: Mendelian randomization study. Neurology 2017, 89, 454–460. [Google Scholar] [CrossRef] [PubMed]
- Codd, V.; Nelson, C.P.; Albrecht, E.; Mangino, M.; Deelen, J.; Buxton, J.L.; Hottenga, J.J.; Fischer, K.; Esko, T.; Surakka, I.; et al. Identification of seven loci affecting mean telomere length and their association with disease. Nat. Genet. 2013, 45, 422–427. [Google Scholar] [CrossRef] [PubMed]
- Malik, R.; Chauhan, G.; Traylor, M.; Sargurupremraj, M.; Okada, Y.; Mishra, A.; Rutten-Jacobs, L.; Giese, A.K.; van der Laan, S.W.; Gretarsdottir, S.; et al. Multiancestry genome-wide association study of 520,000 subjects identifies 32 loci associated with stroke and stroke subtypes. Nat. Genet. 2018, 50, 524–537. [Google Scholar] [CrossRef] [PubMed]
- Burgess, S.; Scott, R.A.; Timpson, N.J.; Davey Smith, G.; Thompson, S.G. Using published data in Mendelian randomization: A blueprint for efficient identification of causal risk factors. Eur. J. Epidemiol. 2015, 30, 543–552. [Google Scholar] [CrossRef] [PubMed]
- Wensley, F.; Gao, P.; Burgess, S.; Kaptoge, S.; Di Angelantonio, E.; Shah, T.; Engert, J.C.; Clarke, R.; Davey-Smith, G.; Nordestgaard, B.G.; et al. Association between C reactive protein and coronary heart disease: Mendelian randomisation analysis based on individual participant data. BMJ 2011, 342, d548. [Google Scholar] [CrossRef] [PubMed]
- Yavorska, O.O.; Burgess, S. MendelianRandomization: An R package for performing Mendelian randomization analyses using summarized data. Int. J. Epidemiol. 2017, 46, 1734–1739. [Google Scholar] [CrossRef]
- Burgess, S.; Thompson, S.G. Mendelian Randomization: Methods for Using Genetic Variants in Causal Estimation, 1st ed.; CRC Press: New York, NY, USA, 2015; pp. 1–224. [Google Scholar]
- Burgess, S.; Butterworth, A.; Thompson, S.G. Mendelian randomization analysis with multiple genetic variants using summarized data. Genet. Epidemiol. 2013, 37, 658–665. [Google Scholar] [CrossRef]
- Didelez, V.; Sheehan, N. Mendelian randomization as an instrumental variable approach to causal inference. Stat. Methods Med. Res. 2007, 16, 309–330. [Google Scholar] [CrossRef]
- Brion, M.J.; Shakhbazov, K.; Visscher, P.M. Calculating statistical power in Mendelian randomization studies. Int. J. Epidemiol. 2013, 42, 1497–1501. [Google Scholar] [CrossRef]
- Burgess, S.; Thompson, S.G. Interpreting findings from Mendelian randomization using the MR-Egger method. Eur. J. Epidemiol. 2017, 32, 377–389. [Google Scholar] [CrossRef] [PubMed]
- Burgess, S.; Bowden, J.; Fall, T.; Ingelsson, E.; Thompson, S.G. Sensitivity analyses for robust causal inference from mendelian randomization analyses with multiple genetic variants. Epidemiology 2017, 28, 30–42. [Google Scholar] [CrossRef] [PubMed]
- Haycock, P.C.; Heydon, E.E.; Kaptoge, S.; Butterworth, A.S.; Thompson, A.; Willeit, P. Leucocyte telomere length and risk of cardiovascular disease: Systematic review and meta-analysis. BMJ 2014, 349, g4227. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Feng, C.; Li, L.; Yang, S.; Chen, Y.; Hui, R.; Zhang, M.; Zhang, W. The association of telomere attrition with first-onset stroke in Southern Chinese: A case-control study and meta-analysis. Sci. Rep. 2018, 8, 2290. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.; Pan, B.; Dang, X.; Wu, H.; Xu, D. Relationship between short telomere length and stroke: A meta-analysis. Medicine (Baltimore) 2018, 97, e12489. [Google Scholar] [CrossRef] [PubMed]
- Haycock, P.C.; Burgess, S.; Nounu, A.; Zheng, J.; Okoli, G.N.; Bowden, J.; Wade, K.H.; Timpson, N.J.; Evans, D.M.; Willeit, P.; et al. Association between telomere length and risk of cancer and non-neoplastic diseases: A Mendelian randomization study. JAMA Oncol. 2017, 3, 636–651. [Google Scholar] [CrossRef] [PubMed]
- Allende, M.; Molina, E.; Gonzalez-Porras, J.R.; Toledo, E.; Lecumberri, R.; Hermida, J. Short leukocyte telomere length is associated with cardioembolic stroke risk in patients with atrial fibrillation. Stroke 2016, 47, 863–865. [Google Scholar] [CrossRef]
- Zhan, Y.; Clements, M.S.; Roberts, R.O.; Vassilaki, M.; Druliner, B.R.; Boardman, L.A.; Petersen, R.C.; Reynolds, C.A.; Pedersen, N.L.; Hagg, S. Association of telomere length with general cognitive trajectories: A meta-analysis of four prospective cohort studies. Neurobiol. Aging 2018, 69, 111–116. [Google Scholar] [CrossRef]
- Zhan, Y.; Song, C.; Karlsson, R.; Tillander, A.; Reynolds, C.A.; Pedersen, N.L.; Hagg, S. Telomere length shortening and alzheimer disease--a mendelian randomization study. JAMA Neurol. 2015, 72, 1202–1203. [Google Scholar] [CrossRef]
- Cai, Z.; Wang, C.; He, W.; Tu, H.; Tang, Z.; Xiao, M.; Yan, L.J. Cerebral small vessel disease and Alzheimer’s disease. Clin. Interv. Aging 2015, 10, 1695–1704. [Google Scholar] [CrossRef]
- Silverwood, R.J.; Holmes, M.V.; Dale, C.E.; Lawlor, D.A.; Whittaker, J.C.; Smith, G.D.; Leon, D.A.; Palmer, T.; Keating, B.J.; Zuccolo, L.; et al. Testing for non-linear causal effects using a binary genotype in a Mendelian randomization study: Application to alcohol and cardiovascular traits. Int. J. Epidemiol. 2014, 43, 1781–1790. [Google Scholar] [CrossRef] [PubMed]
- Pierce, B.L.; Ahsan, H.; Vanderweele, T.J. Power and instrument strength requirements for Mendelian randomization studies using multiple genetic variants. Int. J. Epidemiol. 2011, 40, 740–752. [Google Scholar] [CrossRef] [PubMed]
- Herrmann, M.; Pusceddu, I.; Marz, W.; Herrmann, W. Telomere biology and age-related diseases. Clin. Chem. Lab. Med. 2018, 56, 1210–1222. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| SNP | Chr. | Locus | ‘Short’ Allele | ‘Other’ Allele | Beta Estimate | SE | Discovery p |
|---|---|---|---|---|---|---|---|
| rs10936599 | 3 | TERC | T | C | −0.097 | 0.008 | 2.5 × 10−31 |
| rs2736100 | 5 | TERT | A | C | −0.078 | 0.009 | 4.4 × 10−9 |
| rs7675998 | 4 | NAF1 | A | G | −0.074 | 0.009 | 4.3 × 10−16 |
| rs9420907 | 10 | OBFC1 | A | C | −0.069 | 0.010 | 6.9 × 10−11 |
| rs8105767 | 19 | ZNF208 | A | G | −0.048 | 0.008 | 1.1 × 10−9 |
| rs755017 | 20 | RTEL1 | A | G | −0.062 | 0.011 | 6.7 × 10−9 |
| rs11125529 | 2 | ACYP2 | C | A | −0.056 | 0.010 | 4.5 × 10−8 |
| Ischemic Stroke | IVW | RIVW | MR-Egger | ||||
|---|---|---|---|---|---|---|---|
| OR (95% CI) | p | OR (95% CI) | p | OR (95% CI) | p | Intercept (p) | |
| AIS | 0.96 (0.85, 1.09) | 0.535 | 0.96 (0.87, 1.07) | 0.452 | 1.32 (0.73, 2.37) | 0.354 | −0.02 (0.277) |
| LAS | |||||||
| ALL SNPs | 0.90 (0.66, 1.24) | 0.530 | 0.90 (0.62, 1.30) | 0.569 | 3.45 (0.79, 15.07) | 0.100 | −0.10 (0.054) |
| 6 SNPs 1 | 0.79 (0.54, 1.15) | 0.210 | 0.84 (0.29, 2.39) | 0.741 | 2.94 (0.34, 25.33) | 0.327 | −0.09 (0.224) |
| CES | |||||||
| ALL SNPs | 0.82 (0.52, 1.32) | 0.417 | 0.91 (0.56, 1.46) | 0.683 | 1.56 (0.17, 13.96) | 0.691 | −0.04 (0.080) |
| 5 SNPs 2 | 0.76 (0.39, 1.46) | 0.404 | 0.98 (0.53, 1.80) | 0.941 | 9.44 (0.55, 163.04) | 0.123 | −0.15 (0.112) |
| SVS | |||||||
| ALL SNPs | 1.06 (0.76, 1.48) | 0.722 | 1.03 (0.72, 1.48) | 0.867 | 2.59 (0.59, 11.38) | 0.208 | −0.07 (0.054) |
| 4 SNPs 3 | 0.72 (0.45, 1.16) | 0.176 | 0.72 (0.54, 0.97) | 0.028 | 1.13 (0.07, 18.38) | 0.932 | −0.03 (0.747) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cao, W.; Li, X.; Zhang, X.; Zhang, J.; Sun, Q.; Xu, X.; Sun, M.; Tian, Q.; Li, Q.; Wang, H.; et al. No Causal Effect of Telomere Length on Ischemic Stroke and Its Subtypes: A Mendelian Randomization Study. Cells 2019, 8, 159. https://doi.org/10.3390/cells8020159
Cao W, Li X, Zhang X, Zhang J, Sun Q, Xu X, Sun M, Tian Q, Li Q, Wang H, et al. No Causal Effect of Telomere Length on Ischemic Stroke and Its Subtypes: A Mendelian Randomization Study. Cells. 2019; 8(2):159. https://doi.org/10.3390/cells8020159
Chicago/Turabian StyleCao, Weijie, Xingang Li, Xiaoyu Zhang, Jie Zhang, Qi Sun, Xizhu Xu, Ming Sun, Qiuyue Tian, Qihuan Li, Hao Wang, and et al. 2019. "No Causal Effect of Telomere Length on Ischemic Stroke and Its Subtypes: A Mendelian Randomization Study" Cells 8, no. 2: 159. https://doi.org/10.3390/cells8020159
APA StyleCao, W., Li, X., Zhang, X., Zhang, J., Sun, Q., Xu, X., Sun, M., Tian, Q., Li, Q., Wang, H., Liu, J., Meng, X., Wu, L., Song, M., Hou, H., Wang, Y., & Wang, W. (2019). No Causal Effect of Telomere Length on Ischemic Stroke and Its Subtypes: A Mendelian Randomization Study. Cells, 8(2), 159. https://doi.org/10.3390/cells8020159