Oligodendrocytes in Development, Myelin Generation and Beyond


Wellcome-Wolfson Institute for Experimental Medicine, Queen’s University Belfast, Belfast BT9 7BL, UK
*
Author to whom correspondence should be addressed.
†
These authors contributed equally.
Cells 2019, 8(11), 1424; https://doi.org/10.3390/cells8111424
Submission received: 15 October 2019
/
Revised: 7 November 2019
/
Accepted: 7 November 2019
/
Published: 12 November 2019
(This article belongs to the Collection Oligodendrocyte Physiology and Pathology Function)
Abstract
:Oligodendrocytes are the myelinating cells of the central nervous system (CNS) that are generated from oligodendrocyte progenitor cells (OPC). OPC are distributed throughout the CNS and represent a pool of migratory and proliferative adult progenitor cells that can differentiate into oligodendrocytes. The central function of oligodendrocytes is to generate myelin, which is an extended membrane from the cell that wraps tightly around axons. Due to this energy consuming process and the associated high metabolic turnover oligodendrocytes are vulnerable to cytotoxic and excitotoxic factors. Oligodendrocyte pathology is therefore evident in a range of disorders including multiple sclerosis, schizophrenia and Alzheimer’s disease. Deceased oligodendrocytes can be replenished from the adult OPC pool and lost myelin can be regenerated during remyelination, which can prevent axonal degeneration and can restore function. Cell population studies have recently identified novel immunomodulatory functions of oligodendrocytes, the implications of which, e.g., for diseases with primary oligodendrocyte pathology, are not yet clear. Here, we review the journey of oligodendrocytes from the embryonic stage to their role in homeostasis and their fate in disease. We will also discuss the most common models used to study oligodendrocytes and describe newly discovered functions of oligodendrocytes.
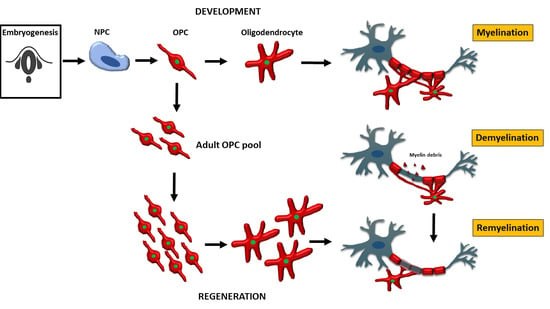
1. Introduction
Oligodendrocytes are the myelinating cells of the central nervous system (CNS). They are generated from oligodendrocyte progenitor cells following tightly orchestrated processes of migration, proliferation and differentiation [1]. Oligodendrocytes are fundamental to myelin formation in the developing CNS and critical for myelin regeneration following injury including in the most common demyelinating disease multiple sclerosis (MS) [2,3]. In this review, we discuss the journey of oligodendrocytes in development from the embryonic stage to oligodendrocyte function in health and their fate in disease. We will also review the most common models used to study oligodendrocyte behaviour and function and discuss recent advances in our knowledge of oligodendrocytes.
2. Oligodendrocyte Progenitor Cells (OPC) and Oligodendrocytes in Development
2.1. OPC and Oligodendrocytes during Embryonic Development
Oligodendrocytes are one of the major glial cell types in the CNS besides microglia and astroglia. Oligodendrocytes were first described by Virchow [4], Deiters [5] and Golgi [6] in the 19th century. While neurons had been well characterised at that time, an abundant unknown cell type called “neuroglia” became the focus of attention. Analysing the fine structure of the brain led Virchow to introduce the term “Nervenkitt” (German: nerve glue) as cells appeared in between nerves [4,7]. Distinct from neurons, these cells could not grow axons [5] and were able to proliferate even postnatally. Initially, they were not thought to have a role other than being connective tissue. Pio del Rio-Hortega eventually differentiated neuroglia into microglia and four types of oligodendroglia by using more advanced staining techniques, including silver carbonate [8,9].
Amongst the glial populations, bipolar cells were found to be highly proliferative and migratory, whereas filamentous myelin-producing glia were mainly found in white matter suggesting a bipolar precursor cell type, later termed oligodendrocyte progenitor cell (OPC), and a distinct, differentiated cell type, the oligodendrocyte [10].
The origin of neurons was well characterised within the CNS yet less was known about the origin of oligodendrocytes and microglia [11,12]. Microglia originate from the yolk sack [13], while oligodendrocytes develop from multiple origins in the brain and anterior spinal cord with radial glial cells, the ventral ventricular zone and dorsal spinal cord as sources [14,15,16,17,18,19]. Neural progenitor cells (NPC) arising in the neural tube during embryonic development are the common precursors for oligodendrocytes, astrocytes and neurons [20,21].
Murine OPC in brain and spinal cord are characterised by the expression of DM-20 mRNA [16,22]. Using this marker, three temporal waves of OPC development were identified starting in the ventral ventricular zone after neural tube closure from E9.5 [16]. Rat OPC first appeared at around E14 [17] and human OPC at E45, reflecting the gestational week 6.5 [23]. Across species, several waves of OPC generation were identified. The first wave of OPC generation in the forebrain is followed by a smaller second wave from the dorsal ventricular zone and by a third postnatal wave originating in the cortex [24]. In the spinal cord, ventrally-derived OPC are followed by a wave of dorsally-derived OPC [25,26], which make up to 20% of total OPC [19,25].
These waves subsequently lead to an overproduction of progenitor cells, which compete for space and survival factors provided by astrocytes and axons [24,27,28,29]. It was shown that most OPC from the first cortical wave die [24,27]. However, Orduz et al. recently demonstrated that a subpopulation of first-wave cortical OPC not only survive but have non-redundant functions [30].
In the adult normal CNS, the majority of proliferating cells are of oligodendroglial lineage origin, which by dividing self-renew and generate mature oligodendrocytes, whereas little evidence exists that neurons are able to proliferate in adulthood [31,32,33,34].
During vertebrate development, one of the major and best-established morphogens is the Sonic hedgehog (Shh) protein, required for most OPC to originate during early embryogenesis [26,35]. Shh is produced by the notochord, which is essential for the CNS dorsal–ventral axis development [36,37]. OPC as well as motor neurons originate from similar areas in the ventral neural tube during development and need equal amounts of Shh in order to arise, which suggests an interlinked generation of both cell types [38,39,40]. However, in vitro experiments with Shh deficient cells indicated that Shh is not an absolute requirement for OPC origination [41], while mice lacking a notochord did not develop any OPC [38]. Cai et al. corroborated these findings in vivo demonstrating Nkx6- and Shh-independent generation of an OPC subpopulation, which suggests the involvement of other molecular pathways [26]. Indeed, Shh is important for the timing of OPC generation as a recent study has shown [42].
Tightly regulated epigenetic mechanisms, such as DNA methylation and histone modification, have recently been discovered in the regulation of OPC differentiation that are distinct in the different developmental stages and in myelin regeneration (reviewed in detail in [43] ).
2.2. Distribution of OPC and Oligodendrocytes within the CNS
Only 5%–8% of total glial cells are OPC [48], which are evenly distributed in white (WM) and grey matter (GM), with OPC being slightly less abundant in GM [48]. The location gives rise to behavioural differences between WM and GM OPC; while WM NG2+ OPC in organotypic brain slices had a greater proliferative response to PDGF-A, GM OPC were less responsive to PDGF-A and morphologically and genetically less mature than WM OPC [49,50]. In vivo, more WM OPC differentiate into myelinating oligodendrocytes than GM OPC, many of which remain NG2+ progenitors as shown by Dimou et al. [51,52], suggesting a potential backup pool of OPC during adulthood. In the adult CNS, oligodendrocyte generation from OPC is slowed down and WM OPC generate about 20% of total differentiated and myelinating oligodendrocytes in the murine corpus callosum vs. 5% in the cortex [53]. However, 20% of cortical GM oligodendroglial lineage cells are differentiated CNP+ NG2- oligodendrocytes yet these cells do not myelinate [53]. Recently, Hughes et al. demonstrated that cortical NG2+ cells are highly dynamic, balancing their population by proliferation, differentiation and self-repulsion to maintain homeostasis [54].
In order for axonal myelination to occur, migration of OPC from their site of origin into the developing WM tracts of the CNS is required [55]. To overcome this spatial distance, OPC migrate in a jumping or crawling mode along blood vessels within the CNS, which is dependent on WNT signalling [56,57]. Their subsequent excessive proliferation, especially in the WM, leads to an abundant pool of progenitors throughout the brain and spinal cord [58].
2.3. Developmental Markers of OPC and Oligodendrocytes
New-born OPC are characterised by the expression of DM-20 mRNA, an isoform of protein proteolipid protein (PLP), the most abundant myelin protein [16]. There are numerous additional markers that determine the oligodendroglial cell lineage and reflect their developmental stage, the most prominent are summarised in Figure 1. Once committed to the oligodendroglial lineage, cell surface antigens can be recognized by specific antibodies such as A2B5 [59]. In vitro, A2B5 positive cells can differentiate into both oligodendrocytes and astrocytes, which were therefore termed oligodendrocyte-type-2 astrocyte (O-2A) progenitor cells [60]. O-2A progenitor cells constitutively differentiate into oligodendrocytes unless specific environmental cues redirect differentiation into astrocytes [61].
The best characterised marker for OPC is PDGFR-α, the receptor for PDGF-A, the most potent OPC mitogen and survival factor, which is produced by both astrocytes and neurons [15,62,63,64]. Consequently, overexpression of this growth factor, e.g., during development, leads to increase in OPC numbers [64].
Pre-oligodendrocytes engage with a target axon, thereby losing their bipolarity, and start to build filamentous myelin outgrowths. At this differentiation stage, pre-oligodendrocytes are characterised by the expression of three main myelin associated markers, 2′, 3′-cyclic-nucleotide 3′-phosphodiesterase (CNPase) and the cell surface markers O4 and O1 [65,66]. CNPase has two different isoforms which are differentially expressed: oligodendrocyte precursors only seem to express the larger isoform, whereas myelinating oligodendrocytes were shown to express both isoforms [67]. O4 is already expressed in late progenitors, whereas O1 is typical for pre-myelinating oligodendrocytes [68] (Figure 1).
Mature, differentiated oligodendrocytes are characterised by the production of myelin and myelin proteins, and in combination with a cell lineage specific marker such as Olig2 can be used to identify this maturation stage (Figure 1). The myelin proteins include myelin basic protein (MBP) [69,70], which is expressed on the cytoplasmic surface of the plasma membrane [71], the transmembrane protein PLP [16,72], myelin associated glycoprotein (MAG) [73] as well as the membrane marker galactocerebroside (GalC) [74] and surface marker myelin-oligodendrocyte glycoprotein (MOG) [69]. MBP and MAG first appear between postnatal day 5 and 7 in murine CNS-derived oligodendrocytes, whereas MOG emerges one to two days later [75]. Interestingly, PLP was recently described to be expressed in murine Olig2+PDGFR-α+ cells, which makes it an early marker for OPC with a role in process extension [76].
Two genetically related, but functionally different, transcription factors Olig1 and Olig2 are present throughout the oligodendroglial lineage [77,78]. Olig2 is essential for NPC to develop into OPC [79] and Olig2 gain of function in OPC can promote remyelination in mice [80]. The role of Olig1 is less well known. Olig1 has a non-redundant role in OPC differentiation and myelination in murine brain development; however, spinal cord OPC are less dependent on Olig1 for differentiation and myelination [81,82]. In repair, however, Olig1 deficient mice showed delayed oligodendrocyte differentiation and impaired remyelination of demyelinated CNS lesions [83]. Another transcription factor involved in oligodendrocyte development that characterises the entire oligodendroglial lineage is SOX10 [84]. SOX10 is essential for NPC derived oligodendroglial lineage specification during early development and for OPC differentiation [85,86,87], while transcription factor Nkx2.2 promotes and regulates timing of oligodendrocyte differentiation [88,89]. A marker for early oligodendroglial lineage cells is the proteoglycan NG2, however, NG2+ cells are able to differentiate into oligodendrocytes but also into astrocytes [90]. Antibodies against the markers described here are commercially available.
2.4. Other Myelin-Producing Cells
Schwann cells are the oligodendrocyte counterpart in the peripheral nervous system (PNS), derived from the neural crest [91]. Oligodendrocytes and Schwann cells share main functions in providing support and insulation for axons. However, Schwann cells are only able to myelinate single axons rather than multiple axons contrary to oligodendrocytes (reviewed in [92,93]). This feature is dependent on an E3 ligase component, deficiency of which leads to increased myelination potential in murine Schwann cells in vivo [94].
Schwann cells as well as oligodendrocytes are region specific for the CNS and the peripheral nervous system and a glial barrier at the motor exit points was found to prevent oligodendrocytes exiting from the CNS. The cells forming this barrier are motor exit point (MEP) glia, which were recently described as the third cell type capable of producing myelin [95,96]. MEP glia share communalities with both oligodendrocytes and Schwann cells; like OPC, they are derived from the ventral neural tube and they express Olig2 as do oligodendroglial lineage cells. MEP glia are also characterised by the expression of SOX10 and WIF1. This, combined with Olig2 and Foxd3 expression, identifies this population [96]. Similar to Schwann cells, MEP glia express Foxd3 and they are able to myelinate just one axon, although the molecular mechanism underlying the process is different. MEP glia lack krox20, a key initiator of myelination for Schwann cells, and are also not affected by gpr126 deficiency as opposed to Schwann cells. Moreover, MEP glia selectively myelinate spinal motor root axons [95,96,97] and were described to have a role in preventing the migration of OPC into the periphery by blocking off OPC in the CNS via direct contact [96].
3. Oligodendrocyte Function in Health
3.1. Myelination of Axons
The myelin sheath is an extension of the oligodendrocyte and Schwann cell plasma membrane that wraps around nerve axons in a concentric fashion [98] as shown in Figure 2. In 1717, Leeuwenhoek described a ‘nervule’ with ‘fatty parts’ around it, likely being the first ever description of myelin [99].
Since then, technical advances in histology and optical techniques have allowed for both the structure and function of the myelin sheath to be explored in detail [3,98,100]. For example, high-resolution time-lapse live imaging, electron microscopy and magnetic resonance imaging of the dynamic processes of myelination and remyelination in vivo have been hugely beneficial to the advancement of our understanding of oligodendrocyte biology in health and disease [3,98,101].
Myelination is a complex and tightly regulated process (reviewed in [102]). In vivo time-lapse imaging in transgenic zebrafish revealed that oligodendrocytes continually extend and retract their processes towards axons before settling into their final positions [103]. The oligodendrocyte processes also sense neighbouring cells to ensure uniform spacing of myelin segments with evenly spaced nodes [104]. Once a mature oligodendrocyte connects with an axon, the oligodendrocyte plasma membrane architecture changes rapidly. Numerous hypotheses have been suggested to explain how the oligodendrocyte extends its plasma membrane to wrap axons and form a compact myelin sheath [99]. Two of the most prominent hypotheses suggest that myelin either extends an inner tongue repeatedly around the axon [105] or alternatively that ‘croissant-like’ layers of myelin sheath are formed on top of pre-existing ones [106]. Sophisticated studies that combined high resolution in vivo imaging and 3D reconstructions of optic nerve fibres fixed with high pressure freezing facilitated improved visualisation of the dynamic process of myelination [107]. These studies revealed that myelination occurs via plasma membrane extension laterally down the axon to form the paranodal loops [107], a discovery consistent with the original mechanism proposed by Geren and Schmitt [105].
Myelination is a highly regulated process that is governed by several molecular cues. Only axons with a large diameter are myelinated [108,109,110], and myelination itself increases the axonal diameter not just simply by the extra myelin sheaths around the axon, but also due to localised neurofilament accumulation and phosphorylation [111,112].
Myelin sheath formation is regulated by several factors such as neuregulin 1 in Schwann cells [113] and Ca2+ activity in oligodendrocytes [101], as well as by neuronal activity, which can identify axons for myelination [46]. Intriguingly, neuronal activity regulates also OPC proliferation, differentiation and survival [44,45,46,47].
3.2. Functions of the Myelin Sheath
Functionally, the myelin sheath facilitates rapid transmission of axon potentials and provides metabolic support to the axons it ensheaths [3,100]. Sodium channels are located at the intermittent interruptions (nodes of Ranvier), where short portions of the axon are left unwrapped [100,114]. Myelin is an electrical insulator and when the membrane at the node is excited, the axon potential ‘jumps’ from one node of Ranvier to the next, due to the low capacitance of the sheath, in a process called saltatory conduction. Little energy is required to depolarise the remaining membrane between the nodes given the low capacitance of the sheath, which results in transmission of the action potential [100,115]. This form of transmission is much faster than in non-myelinated axons [3].
Oligodendrocytes and the myelin sheath metabolically support axons. Oligodendrocytes can generate lactate, which can then be transferred to axons to generate metabolic energy in the form of ATP [116]. In the brain, lactate is shuttled through the most abundant lactate transporter MCT1, and both MCT2 and MCT4 [117]. MCT1 expression has been detected in endothelial cells, however oligodendrocytes and the myelin sheath also harness MCT1 to transport lactate to axons [117,118]. Lee et al. depleted MCT1 gene expression in spinal cord cultures, which led to extensive neuronal death; an effect that could be rescued with exogenous lactate supplementation [117]. Indeed, a number of glycolytic and Krebs cycle enzymes such as succinate dehydrogenase and fumarase are expressed in the myelin sheath, which contribute to glucose catabolism and ATP production [119].
The central function of oligodendrocytes in the CNS is the generation of myelin during development, adaptive myelination in adulthood and remyelination after damage, while OPC predominantly serve as a backup pool to generate new oligodendrocytes. However, CNS remyelination can also be mediated by infiltrating Schwann cells, which was described for multiple conditions including MS and traumatic brain injury [2]. Post-developmental OPC can also differentiate into other neural lineage cells such as astrocytes and Schwann cells [120]. OPC-derived Schwann cells reside in CNS regions deficient in astrocytes, suggesting that severely demyelinated lesions that are partially necrotic may inhibit astrogliosis, and that a lack of astrocytic scarring may encourage Schwann cell infiltration [120,121]. The factors governing transition from OPC to Schwann cells remain elusive [2], but these data suggest that an astrocyte-derived factor is necessary for OPC differentiation into oligodendrocytes [120].
3.3. Non-Myelinating Functions of Oligodendrocytes and OPC
Emerging evidence suggests OPC have an immunomodulatory capacity. OPC express cytokine receptors [122,123,124] and assess their microenvironment through filopodia extension [103]. In response to inflammatory cues, OPC can migrate to sites of injury, in a manner similar to that of microglia [54,125]. Upon exposure to IFNγ, OPC cross-present antigen to cytotoxic CD8+ T cells in vitro and in vivo, leading to their cytotoxic death [126,127]. This newly discovered pro-inflammatory OPC phenotype promotes tissue damage and blocks remyelination. This suggests that suppression of OPC-mediated inflammation may ameliorate cell death in favour of promoting OPC differentiation into myelin-producing oligodendrocytes [127].
4. Models to Study Oligodendrocytes and OPC
4.1. Rodent and Human In Vitro Models
Several protocols have been developed to isolate and culture oligodendrocytes from rodent brains at different stages from embryonic into adulthood [128,129,130]. The isolation of OPC and oligodendrocytes from murine CNS tissue is based on the expression of markers typical of this maturation stage, for example PDGFR-α or A2B5 for OPC or O4 or GalC for oligodendrocytes [129,130]. These primary cultures are suitable for studying the proliferation, survival and differentiation of OPC as well as the effect of molecules of interest on OPC and oligodendrocyte biology. Moreover, different protocols have been developed to study myelination by co-culturing oligodendrocytes with neurons or synthetic nanofibers [110,131]. Primary OPC cultures derived from e.g., rodents, are particularly interesting for studying the proliferation and differentiation of OPC and are suitable for high throughput screening of pharmacological compounds that may interfere with these processes [132].
However, primary OPC cultures are restricted by the availability of animals and related ethical issues. To overcome these limitations, several laboratories developed spontaneously immortalised cell lines derived from O-2A rat precursors, such as CG-4 or OLN-93 immortalised OPC [133,134], or primary murine OPC immortalised by viral infection, such as Oli-neu [135]. These cell lines are karyotypically normal, express the marker typical of their developmental stages and they can be differentiated into mature oligodendrocytes by withdrawing mitogenic factors, such as in CG-4, or by adding factors, such as dibutyryl cAMP for Oli-neu and OLN-93 [133,134,135]. Immortalised cell lines have undeniable advantages, such as the consistency, the robustness and the low costs related to the model, but because of the unpredictable nature of mutations that lead to the immortalisation, any assumption and translation to any physiological function need to be corroborated by an additional model.
Although the biology of oligodendroglial cells is conserved between rodents and humans, there are some differences between species that should be considered for any translation from murine models to the human physiology (reviewed in [136,137]). The peculiar structure and position of the brain and spinal cord reduces the possibilities of obtaining cells directly from human CNS tissue. One strategy that can be adopted is to isolate OPC from surgery biopsies, e.g., from healthy resection margin of brain tumours (excluding the tumour itself), resected tissue from epileptic or traumatic brain tissue [138]. Another strategy involves the use of post mortem brains or spinal cords; as cells can be isolated up to 12 h after death, from any area of the CNS [139]. Adult human oligodendrocytes can be cultured for 2–3 weeks or directly used for further analysis (for example for mRNA or protein analysis). Although these approaches are precious to unravel the differences between rodent and human biology, the availability of these tissues and the possibility to isolate only oligodendrocytes are a limiting factor.
In recent years, the establishment of induced pluripotent stem cells (iPSC) overcame the restricted availability of human CNS biopsies [140]. iPSC are reprogrammed from somatic cells (e.g., fibroblasts, peripheral blood mononuclear cells) that can be obtained in a non-invasive way. iPSC can subsequently be differentiated to any cell type of interest by inducing their differentiation via specific growth factors (e.g., T3, NT3, IGF, PDGF-A for inducing OPC) or inducing the expression of transcription factors necessary for the transition (e.g., SOX10, Olig2 and NKX6.2 for OPC/oligodendrocytes) [141,142,143]. Recently, protocols with increasing efficiency and reduced time to obtain oligodendrocytes from human iPSC (iOL) were developed; from 170 days in vitro to 20 days to obtain a culture with mature oligodendrocytes (around 70% efficiency of which 20% expressing MBP) ([141,142,143] and reviewed in [137]). The possibility of having OPC and oligodendrocytes derived from healthy donors and patients with neurological diseases as a model of study made iPSC popular in the last decade as proven by the variety of protocols developed. Moreover, the possibility of following the differentiation of OPC into iOL is a valuable tool to study this process in a human system. iOL can also represent a precious model to screen drugs promoting differentiation and/or myelin production directly on human cells, possibly directly derived from patients, as exemplified by Ehrlich et al. [142]. Unfortunately, the elevated costs and the time necessary to obtain iOL reduce the accessibility to this technique on a routine basis in the majority of laboratories. More efficient and cheaper protocols to obtain iOL are desirable for the future to broaden our knowledge in human oligodendrocyte biology.
The methods described above (summarised in Table 1) entail the use of an isolated population. Other methods have been developed to mimic the complexity of the environment in which OPC and oligodendrocytes reside, such as mixed glia cultures [144], organotypic brain slices [145], and iPSC derived organoids [146]. These models are useful to study the impact of other cell types on OPC and oligodendrocytes, to decipher any indirect effect exerted by other CNS cells and to complement in vivo studies.
4.2. In Vivo Models
One of the most used strategies to study oligodendroglial lineage cells is to genetically label markers typical of the lineage or of the maturation stage of interest with fluorescent proteins (see Table 1). These are particular suitable models to study the biology of oligodendrocytes in the context of a complex biological system. The most used species used for this purpose are mice or zebrafish. Mice generated to detect OPC during development and maturation are for example Sox10-Venus [147] or CNP-EGFP mice [148]. Other mice were generated targeting specific proteins, such as PLP-EGFP mice [149]. In recent years, zebrafish became popular as a model to study remyelination, due to some unique features: large numbers of offspring that develop quickly and outside of the mother, the transparency and small size of the animals that make live imaging of genetically modified zebrafish possible, the reduced costs and reduced level of self-consciousness compared to mammalian models. In fact, zebrafish share 70% of the genome with humans and most of the genes related to myelin are conserved [152,153]. Different transgenic zebrafish have been developed to study different stages and function of OPC and oligodendrocytes such as Tg(sox10:mRFP), Tg(olig2:EGFP) and Tg(mbp:EGFP) [103,150,151]. Given the consistency of the model, transgenic zebrafish are also suitable for high-throughput screening: recently, Early et al. developed an automatic analysis tool where zebrafish larvae are automatically delivered to a spinning disk confocal microscope and the images handled by an image analysis pipeline, facilitating the screening of pro-myelinating compounds [154].
4.3. Animal Models to study Oligodendrocytes and Remyelination in Multiple Sclerosis
There are no animal models that entirely recapitulate all of the features of MS, but different murine models are established to investigate different aspects of the pathology [155]. Experimental autoimmune encephalomyelitis (EAE) is an acute demyelinating episode triggered by the immune system stimulated by myelin protein with adjuvants [156]. EAE is a reproducible, well established model with a defined clinical pattern making it suitable to investigate the relevance of different molecular pathways for MS progression or the efficacy of new treatments.
Two models of non-immune-mediated demyelination use either dietary given cuprizone (a copper chelator) or injected lysolecithin (a lipidolitic detergent) [157,158]. Although the initiation of these models lacks immune engagement, immune cells are recruited to the demyelinated areas, indicating the importance of these cells also in the repair phase [159,160]. These models do not recapitulate any of the typical symptoms that patients with MS experience, but are very useful to investigate molecular and cellular mechanisms involved in the damage and repair phase. Cuprizone is a toxin that induces acute demyelination and is characterised by a peak of OPC infiltration 4 weeks after the start of the cuprizone diet in the corpus callosum, one of the most demyelinated areas in this model [161,162]. The infiltrated OPC are able to differentiate into oligodendrocytes in the following 2 weeks, representing the most abundant population after 6 weeks of cuprizone diet [162]. However, remyelination starts only after cuprizone is withdrawn: in one week, indeed, it is possible to observe an increased expression of myelin associated markers in the corpus callosum that stabilises upon complete remyelination 5 weeks after the re-establishment of a normal diet [162]. Interestingly, Sachs et al. recently developed a modified protocol able to slow down remyelination by the administration of rapamycin, an mTOR inhibitor, to the cuprizone treated mice, giving an extended temporal window to study the processes involved in myelin regeneration [162].
Lysolecithin induced demyelination is also a valuable model to study the remyelination process given the well described and distinct phases of OPC recruitment and proliferation in the lesion site between day five and 10 after the lesion induction, OPC differentiation between day 10 and 14 followed by myelin regeneration [83,158,163,164]. This well-established model is suitable for distinguishing the effect of molecules and cells of interest, including oligodendrocytes, in the different phases of myelin regeneration in an accurate manner.
The aforementioned models are able to recapitulate some features of MS; a comprehensive in vivo model reflecting the complexity of the disease is still lacking. Combination models have been developed to align animal models closer with human disease. For example, Rüther et al. combined cuprizone induced demyelination with EAE to induce a chronic model with persistent clinical signs and chronic lesions even in the forebrain, a region that is not usually affected in EAE, and with a robust immune component [165].
5. Oligodendrocytes in Disease
5.1. Oligodendrocytes in Diseases with Myelin Pathology
Oligodendrocyte death is not necessarily a sign of disease. Oligodendrocytes can die throughout development and adulthood without underlying pathology to enable neuronal plasticity and lifelong learning [166,167]. However, OPC and particularly oligodendrocytes are highly vulnerable to oxidative stress due to low antioxidant levels, for instance glutathione, and high iron content, which is required for enzyme activity. Iron in combination with hydrogen peroxide, from dismutation of superoxide, can produce highly reactive hydroxyl radicals via the Fenton reaction [168].
Especially during myelination oligodendrocytes are vulnerable to cytotoxic by-products from a high metabolic turnover (e.g., reactive oxygen species (ROS), hydrogen peroxide) [169,170]. Likewise, oligodendrocytes are susceptible to excitotoxicity from high glutamate and ATP concentrations [171,172]. Due to this vulnerability oligodendrocytes are affected in a range of disorders.
The most common pathological causes of oligodendrocyte death in the CNS are trauma, ischaemia or autoimmune attacks. Oligodendrocyte death can lead to subsequent demyelination or it can follow as a result of primary myelin damage [173]. In traumatic injuries and ischaemia, oligodendrocyte death and demyelination can follow the original injury [120,174], whereas in autoimmune diseases such as MS, oligodendrocytes are a primary target of an immune attack against myelin and oligodendrocyte specific proteins.
Much rarer are genetic defects that lead to oligodendrocyte damage as seen in some leukodystrophies [175]. The main cause of oligodendrocyte death in these diseases seem to be the accumulation of mutated PLP1, a prevalent protein in the myelin sheath, that fails to be transported to the plasma membrane leading to apoptosis of the cell [176].
White matter pathology is a characteristic of Alzheimer’s disease (AD), however oligodendrocyte death and demyelination are believed to occur secondary to neurodegeneration [177]. In post mortem AD tissue Olig2+ oligodendrocyte lineage cells were decreased [178] as were myelinating oligodendrocytes in a mouse model of AD [179]. The underlying cause is not established; however toxicity of beta-amyloid is likely a contributing factor [179,180]. Yet, oligodendrocyte pathology can be evident even before any neurodegenerative events materialise as a study by Fischer showed [181]. Intriguingly, WM pathology in AD is predominately affecting those CNS regions that were myelinated last during development (neuropathologic retrogenesis) [182,183], suggesting a connection between late myelin development and AD.
While oligodendrocyte pathology is not regarded as the primary cause, oligodendrocytes are downstream targets in some neuropsychiatric disorders including schizophrenia, bipolar disorders, autism, ADHD, mood disorders and depression (reviewed in [184] and [185]). For instance, oligodendrocyte density was region-specifically reduced in patients with bipolar disorders and schizophrenia [186,187,188], and animal models to study demyelination, such as the cuprizone model, have also been used to model aspects of schizophrenia and anxiety disorders (reviewed in [189]). In a chronic stress model oligodendrocyte specific genes were downregulated in the amygdala and prefrontal cortex [190], while dysregulation of oligodendrocytes and nodes of Ranvier is associated with depression [191].
5.2. Oligodendrocytes in Demyelination
Demyelinating diseases are the most common pathologies affecting oligodendrocytes. The most common demyelinating disease is MS [192]. MS is characterised by an immune-mediated attack against myelin sheaths and oligodendrocytes, primarily by myelin-specific CD8+ T cells [193]. This results in demyelination, which is characterised by destruction of the myelin sheath and death of oligodendrocytes [194] leaving axons denuded and vulnerable to neurodegeneration [1].
Genetic fate-mapping studies allowed adult OPC differentiation fates to be tracked after demyelination. This confirmed that adult OPC that represent approximately 6% of the total CNS cell numbers [48] are the main source of new oligodendrocytes after myelin damage [195]. In response to demyelination, adult OPC undergo a switch to an activated state that is characterised by an increased expression of numerous transcription factors [196,197]. These include SOX2 and TCF7L2, the latter of which is central to WNT signalling and maintains OPC in the cell cycle to enable proliferation and colonisation of areas of demyelination [2,197]. OPC migrate to the lesion, guided by microglia and astrocyte-derived factors including fibroblast growth factor and brain-derived neurotrophic factor [2,198,199], before differentiating into mature oligodendrocytes by exiting the cell cycle [200], a conversion led by the transcription factor E2F1 and proto-oncogene MYC [201,202]. The new oligodendrocyte is able to replace the destroyed myelin with a shorter and thinner myelin sheath [157] as shown in Figure 2.
5.3. Oligodendrocytes in Ageing
Ageing is associated with increased white matter atrophy, a decline in motor learning and diminished remyelination capacity [203,204]. The latter is due to changes in the ageing CNS environment [204] and to the decreased ability to recruit OPC that subsequently fail to differentiate into oligodendrocytes [205,206]. Rejuvenation can restore the reparative capacity in the CNS as the Franklin lab has shown: in a parabiosis experiment with young and old mice that were subjected to demyelination macrophages were key to restore the remyelination capacity in old animals by clearing myelin debris [204].
Due to their high metabolic rate during myelination, for the maintenance of myelin and for the trophic support of axons [117] oligodendrocytes are exposed to prolonged periods of cytotoxic by-products such as ROS; for instance, cholesterol synthesis is highly energy intensive [207] and also declines with age [208]. Additionally, compared to astrocytes oligodendrocytes have reduced antioxidants levels [169,209], which further decline with age [210]. Oligodendrocytes are highly vulnerable to oxidative stress, which can result in DNA damage [211]. With age, DNA repair mechanisms are either not efficient or are disturbed thus leading to oligodendrocyte apoptosis. As OPC recruitment and differentiation into oligodendrocytes is diminished with age [205,212], dead oligodendrocytes are not as efficiently replaced in old age as they were in younger age leading to white matter atrophy over time.
6. Oligodendrocytes in Myelin Regeneration
Remyelination is a natural regenerative process, which is believed to prevent neurodegeneration and restore function [2,213]. In MS and animal models of MS, the ability to remyelinate efficiently declines with age and disease progression (reviewed in [214]). Our knowledge about the mechanisms involved is mainly based on animal studies, which although powerful, deliver a simplified picture of what occurs in humans. This becomes increasingly clear with results from recent cell population studies in human healthy and MS brain tissue demonstrating the existence of different oligodendrocyte lineage cells, dependent on location, origin, disease phenotype and age [215,216,217]. New functions of oligodendrocyte lineage cells beyond their central roles in myelination and trophic support were recently identified including immunomodulatory properties as shown by Falcao et al. [126]. Given that MS is an immune-mediated disease, it would be striking if oligodendrocytes are not just the targets of an immune attack in MS but are actively involved in disease pathology.
OPC are central to remyelination by generating new oligodendrocytes that replace those lost after injury [218,219]. In contrast, surviving adult oligodendrocytes were long thought to not be involved in remyelination. This dogma has recently been revised for both rodents and humans as studies by Duncan et al. and Yeung et al. demonstrated [215,220]. Using rodent and non-human primate animal models, Duncan et al. showed that oligodendrocytes are connected to both myelinated and remyelinated myelin sheaths indicating that existing oligodendrocytes contribute to remyelination [220]. Comparing C14 in genomic DNA with atmospheric C14, Yeung et al. calculated the age of oligodendrocytes in MS lesions. The authors concluded that rather than newly formed oligodendrocytes from OPC old ‘spared’ oligodendrocytes regenerated myelin in remyelinated MS lesions [215].
Remyelination seems to replicate aspects of developmental myelination, at least in parts. For instance, similar transcription factors are involved in both processes, such as Myrf and Zfp488 [221,222]. Conversely, while Klf9 and Stat3 seem non-essential in developmental myelination they are critical for efficient remyelination [223,224] and Arnett et al. showed that the transcription factor Olig1 is more important than Olig2 in remyelination suggesting their role is reversed compared to developmental myelination [83].
In human MS lesions, Chang et al. and Kuhlmann et al. identified a block in OPC differentiation into oligodendrocytes as the bottleneck for efficient remyelination in MS [225,226]. In early MS and in successfully remyelinated lesions, differentiated oligodendrocytes are found within WM lesions, while in chronic stages of the disease, which is characterised by predominantly non-remyelinating lesions, differentiated oligodendrocyte are rare [225]. Albeit in lower numbers, OPC are still found in these chronic MS lesions indicating that OPC deficiency is not the primary cause for remyelination failure; it can, however, contribute to the impaired differentiation of OPC into remyelinating oligodendrocytes [225,227].
A study by Boyd et al. showed that deficiency of OPC within lesions either due to impaired recruitment and/or proliferation is another cause of remyelination failure for 37% of MS lesions, predominantly chronic active lesions. OPC deficiency in these lesions is linked to Semaphorin 3A that prevents OPC migration into the lesion area [205,228].
To overcome remyelination failure in MS new targets to enhance remyelination therapeutically are currently examined. Originally thought to only contribute to the pathology of MS, the immune system is now recognised in facilitating remyelination. Depletion of macrophages or lymphocytes leads to impaired remyelination following toxin-induced demyelination in animal models [159,229]. For instance, M2 macrophages and microglia promote oligodendrocyte differentiation via activin-A [230] and we showed that regulatory T cells promote oligodendrocyte differentiation and remyelination via CCN3 secretion [231].
7. Concluding Remarks
Oligodendrocytes have come a long way from the original description as ‘Nervenkitt’ to their central importance as the myelinating cells of the CNS. Adding to this key function, oligodendrocytes provide trophic support to axons and more recently, likely have additional immunomodulatory capacity. While we have a good understanding of the role of oligodendrocytes in myelination and remyelination, there is still a gap of knowledge of the molecular mechanisms involved in these processes. Given the similarities between myelination and remyelination, a better understanding of myelination can provide clues that can be harnessed to improve myelin regeneration, an unmet need for the treatment of demyelinating diseases, such as MS.
Only recently, new roles of oligodendrocytes have been uncovered such as potential immunomodulatory functions, which warrant further investigation, particularly in disease context. For instance, the question of whether oligodendrocytes are actively participating in disease progression e.g., in MS, by perpetuating inflammation would be interesting to address.
Advances in the models to study oligodendrocyte biology and function, such as cell population-based approaches, has broadened our knowledge beyond their classical roles.
Despite the mechanistic similarities between animals and humans in development and maturation of OPC and oligodendrocytes, there are differences that need to be accounted for when translating results to the human system. Different in vitro approaches based, e.g., on human iPSC will help to overcome the intrinsic interspecies differences that can complement and validate the discoveries from animal models.
In conclusion, oligodendrocytes have long left behind the ‘Nervenkitt’ attribute and are recognised as critical regulators of neuronal functions in CNS development, homeostasis and regeneration.
Author Contributions
Conceptualization, Y.D., L.G., S.K. and D.C.; Writing—Original Draft Preparation, S.K., L.G., D.C. and Y.D.; Writing—Review & Editing, S.K., L.G., D.C. and Y.D.; Visualization, S.K.; Supervision, Y.D.; Project Administration, L.G. and Y.D.; Funding Acquisition, Y.D.
Funding
Part of this work was supported by a grant to Y.D. from the Medical Research Foundation of the Freemasons of Ireland (PhD studentship D.C.).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Bradl, M.; Lassmann, H. Oligodendrocytes: Biology and pathology. Acta Neuropathol. 2010, 119, 37–53. [Google Scholar] [CrossRef] [PubMed]
- Franklin, R.J.M.; Ffrench-Constant, C. Regenerating CNS myelin-from mechanisms to experimental medicines. Nat. Rev. Neurosci 2017, 18, 753–769. [Google Scholar] [CrossRef] [PubMed]
- Nave, K.A. Myelination and support of axonal integrity by glia. Nature 2010, 468, 244–252. [Google Scholar] [CrossRef] [PubMed]
- Virchow, R. Gesammelte Abhandlungen zur Wissenschaftlichen Medicin; Hamm, Frankfurt a.M. 1856. Available online: https://archive.org/details/b21462161 (accessed on 9 November 2019).
- Deiters, O.F.C. Untersuchungen über Gehirn und Rückenmark des Menschen und der Säugetiere; Braunschweig. 1865. Available online: https://archive.org/details/untersuchungen00deit/page/n8 (accessed on 9 November 2019).
- Golgi, C. Sulla Fina Anatomia Degli Organi Centrali del Sistema Nervoso. Milano, 1885. Available online: https://archive.org/details/b21978724/page/n8 (accessed on 9 November 2019).
- Virchow, R. Die Cellularpathologie in ihrer Begründung auf physiologische und pathologische Gewebelehre. Berlin, 1858. Available online: https://archive.org/details/diecellularpatho00virc/page/n8 (accessed on 9 November 2019).
- Del Rio-Hortega, P. La microglia y su transformacíon en células en bastoncito y cuerpos gránulo-adiposos. Trab. Lab. Invest. Biol. Madrid. 1920, 18, 37–82. [Google Scholar]
- Del Rio-Hortega, P. Arte y artificio de la ciencia histológica. Residencia Revista de la Residencia de Estudiantes IV. 1933, 4, 191–206. [Google Scholar]
- Hardy, R.; Reynolds, R. Proliferation and differentiation potential of rat forebrain oligodendroglial progenitors both in vitro and in vivo. Development 1991, 111, 1061–1080. [Google Scholar] [PubMed]
- Rexed, B. The cytoarchitectonic organization of the spinal cord in the cat. J. Comp. Neurol. 1952, 96, 415–495. [Google Scholar] [CrossRef] [PubMed]
- Nornes, H.O.; Das, G.D. Temporal pattern of neurogenesis in spinal cord of rat. I. An autoradiographic study — time and sites of origin and migration and settling patterns of neuroblasts. Brain Res. 1974, 73, 121–138. [Google Scholar] [CrossRef]
- Ginhoux, F.; Prinz, M. Origin of microglia: Current concepts and past controversies. Cold Spring Harb. Perspect. Biol. 2015, 7, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Raff, M.C.; Miller, R.H.; Noble, M. A glial progenitor cell that develops in vitro into an astrocyte or an oligodendrocyte depending on culture medium. Nature 1983, 303, 390–396. [Google Scholar] [CrossRef] [PubMed]
- Pringle, N.P.; Richardson, W.D. A singularity of PDGF alpha-receptor expression in the dorsoventral axis of the neural tube may define the origin of the oligodendrocyte lineage. Development 1993, 117. [Google Scholar]
- Timsit, S.; Martinez, S.; Allinquant, B.; Peyron, F.; Puelles, L.; Zalc, B. Oligodendrocytes originate in a restricted zone of the embryonic ventral neural tube defined by DM-20 mRNA expression. J. Neurosci. 1995, 15, 1012–1024. [Google Scholar] [CrossRef] [PubMed]
- Warf, B.C.; Fok-Seang, J.; Miller, R.H. Evidence for the ventral origin of oligodendrocyte precursors in the rat spinal cord. J. Neurosci. 1991, 11, 2477–2488. [Google Scholar] [CrossRef] [PubMed]
- Vallstedt, A.; Klos, J.M.; Ericson, J. Multiple dorsoventral origins of oligodendrocyte generation in the spinal cord and hindbrain. Neuron 2005, 45, 55–67. [Google Scholar] [CrossRef] [PubMed]
- Fogarty, M.; Richardson, W.D.; Kessaris, N. A subset of oligodendrocytes generated from radial glia in the dorsal spinal cord. Development 2005, 132, 1951–1959. [Google Scholar] [CrossRef] [PubMed]
- Davis, A.A.; Temple, S. A self-renewing multipotential stem cell in embryonic rat cerebral cortex. Nature 1994, 372, 263–266. [Google Scholar] [CrossRef] [PubMed]
- Rogister, B.; Ben-Hur, T.; Dubois-Dalcq, M. From neural stem cells to myelinating oligodendrocytes. Mol. Cell. Neurosci. 1999, 14, 287–300. [Google Scholar] [CrossRef] [PubMed]
- Spassky, N.; Goujet-Zalc, C.; Parmantier, E.; Olivier, C.; Martinez, S.; Ivanova, A.; Ikenaka, K.; Macklin, W.; Cerruti, I.; Zalc, B.; et al. Multiple restricted origin of oligodendrocytes. J. Neurosci. 1998, 18, 8331–8343. [Google Scholar] [CrossRef] [PubMed]
- Hajihosseini, M.; Tham, T.N.; Dubois-Dalcq, M. Origin of oligodendrocytes within the human spinal cord. J. Neurosci. 1996, 16, 7981–7994. [Google Scholar] [CrossRef] [PubMed]
- Kessaris, N.; Fogarty, M.; Iannarelli, P.; Grist, M.; Wegner, M.; Richardson, W.D. Competing waves of oligodendrocytes in the forebrain and postnatal elimination of an embryonic lineage. Nat. Neurosci. 2006, 9, 173–179. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, R.B.; Clarke, L.E.; Burzomato, V.; Kessaris, N.; Anderson, P.N.; Attwell, D.; Richardson, W.D. Dorsally and Ventrally Derived Oligodendrocytes Have Similar Electrical Properties but Myelinate Preferred Tracts. J. Neurosci. 2011, 31, 6809–6819. [Google Scholar] [CrossRef] [PubMed]
- Cai, J.; Qi, Y.; Hu, X.; Tan, M.; Liu, Z.; Zhang, J.; Li, Q.; Sander, M.; Qiu, M. Generation of oligodendrocyte precursor cells from mouse dorsal spinal cord independent of Nkx6 regulation and Shh signaling. Neuron 2005, 45, 41–53. [Google Scholar] [CrossRef] [PubMed]
- Barres, B.A.; Hart, I.K.; Coles, H.S.; Burne, J.F.; Voyvodic, J.T.; Richardson, W.D.; Raff, M.C. Cell death and control of cell survival in the oligodendrocyte lineage. Cell 1992, 70, 31–46. [Google Scholar] [CrossRef]
- Trapp, B.D.; Nishiyama, A.; Cheng, D.; Macklin, W. Differentiation and death of premyelinating oligodendrocytes in developing rodent brain. J. Cell Biol. 1997, 137, 459–468. [Google Scholar] [CrossRef] [PubMed]
- Barres, B.A.; Raff, M.C. Axonal control of oligodendrocyte development. J. Cell Biol. 1999, 147, 1123–1128. [Google Scholar] [CrossRef] [PubMed]
- Orduz, D.; Benamer, N.; Ortolani, D.; Coppola, E.; Vigier, L.; Pierani, A.; Angulo, M.C. Developmental cell death regulates lineage-related interneuron-oligodendroglia functional clusters and oligodendrocyte homeostasis. Nat. Commun. 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Bhardwaj, R.D.; Curtis, M.A.; Spalding, K.L.; Buchholz, B.A.; Fink, D.; Björk-Eriksson, T.; Nordborg, C.; Gage, F.H.; Druid, H.; Eriksson, P.S.; et al. Neocortical neurogenesis in humans is restricted to development. Proc. Natl. Acad. Sci. USA 2006, 103, 12564–12568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bandeira, F.; Lent, R.; Herculano-Houzel, S. Changing numbers of neuronal and non-neuronal cells underlie postnatal brain growth in the rat. Proc. Natl. Acad. Sci. USA 2009, 106, 14108–14113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barnabé-Heider, F.; Göritz, C.; Sabelström, H.; Takebayashi, H.; Pfrieger, F.W.; Meletis, K.; Frisén, J. Origin of New Glial Cells in Intact and Injured Adult Spinal Cord. Cell Stem Cell 2010, 7, 470–482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inta, D.; Alfonso, J.; von Engelhardt, J.; Kreuzberg, M.M.; Meyer, A.H.; van Hooft, J.A.; Monyer, H. Neurogenesis and widespread forebrain migration of distinct GABAergic neurons from the postnatal subventricular zone. Proc. Natl. Acad. Sci. USA 2008, 105, 20994–20999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orentas, D.M.; Hayes, J.E.; Dyer, K.L.; Miller, R.H. Sonic hedgehog signaling is required during the appearance of spinal cord oligodendrocyte precursors. Development 1999, 126, 2419–2429. [Google Scholar] [PubMed]
- Van Straaten, H.W.M.; Hekking, J.W.M.; Beursgens, J.P.W.M.; Terwindt-Rouwenhorst, E.; Drukker, J. Effect of the notochord on proliferation and differentiation in the neural tube of the chick embryo. Development 1989, 107, 793–803. [Google Scholar] [CrossRef]
- Echelard, Y.; Epstein, D.J.; St-Jacques, B.; Shen, L.; Mohler, J.; McMahon, J.A.; McMahon, A.P. Sonic hedgehog, a member of a family of putative signaling molecules, is implicated in the regulation of CNS polarity. Cell 1993, 75, 1417–1430. [Google Scholar] [CrossRef]
- Pringle, N.P.; Yu, W.-P.; Guthrie, S.; Roelink, H.; Lumsden, A.; Peterson, A.C.; Richardson, W.D. Determination of Neuroepithelial Cell Fate: Induction of the Oligodendrocyte Lineage by Ventral Midline Cells and Sonic Hedgehog. Dev. Biol. 1996, 177, 30–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richardson, M.K.; Hanken, J.; Gooneratne, M.L.; Pieau, C.; Raynaud, A.; Selwood, L.; Wright, G.M. There is no highly conserved embryonic stage in the vertebrates: Implications for current theories of evolution and development. Anat. Embryol. 1997, 196, 91–106. [Google Scholar] [CrossRef] [PubMed]
- Roelink, H.; Augsburger, A.; Heemskerk, J.; Korzh, V.; Norlin, S.; Ruiz i Altaba, A.; Tanabe, Y.; Placzek, M.; Edlund, T.; Jessell, T.M.; et al. Floor plate and motor neuron induction by vhh-1, a vertebrate homolog of hedgehog expressed by the notochord. Cell 1994, 76, 761–775. [Google Scholar] [CrossRef]
- Nery, S.; Wichterle, H.; Fishell, G. Sonic hedgehog contributes to oligodendrocyte specification in the mammalian forebrain. Development 2001, 128, 527–540. [Google Scholar] [PubMed]
- Hashimoto, H.; Jiang, W.; Yoshimura, T.; Moon, K.H.; Bok, J.; Ikenaka, K. Strong sonic hedgehog signaling in the mouse ventral spinal cord is not required for oligodendrocyte precursor cell (OPC) generation but is necessary for correct timing of its generation. Neurochem. Int. 2018, 119, 178–183. [Google Scholar] [CrossRef] [PubMed]
- Tiane, A.; Schepers, M.; Rombaut, B.; Hupperts, R.; Prickaerts, J.; Hellings, N.; van den Hove, D.; Vanmierlo, T. From OPC to Oligodendrocyte: An Epigenetic Journey. Cells 2019, 8, 1236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barres, B.A.; Raff, M.C. Proliferation of oligodendrocyte precursor cells depends on electrical activity in axons. Nature 1993, 361, 258–260. [Google Scholar] [CrossRef] [PubMed]
- Hughes, E.G.; Orthmann-Murphy, J.L.; Langseth, A.J.; Bergles, D.E. Myelin remodeling through experience-dependent oligodendrogenesis in the adult somatosensory cortex. Nat. Neurosci. 2018, 21, 696–706. [Google Scholar] [CrossRef] [PubMed]
- Gibson, E.M.; Purger, D.; Mount, C.W.; Goldstein, A.K.; Lin, G.L.; Wood, L.S.; Inema, I.; Miller, S.E.; Bieri, G.; Zuchero, J.B.; et al. Neuronal Activity Promotes Oligodendrogenesis and Adaptive Myelination in the Mammalian Brain. Science 2014, 344, 1252304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitew, S.; Gobius, I.; Fenlon, L.R.; McDougall, S.J.; Hawkes, D.; Xing, Y.L.; Bujalka, H.; Gundlach, A.L.; Richards, L.J.; Kilpatrick, T.J.; et al. Pharmacogenetic stimulation of neuronal activity increases myelination in an axon-specific manner. Nat. Commun. 2018, 9, 306. [Google Scholar] [CrossRef] [PubMed]
- Dawson, M.R.L.; Polito, A.; Levine, J.M.; Reynolds, R. NG2-expressing glial progenitor cells: An abundant and widespread population of cycling cells in the adult rat CNS. Mol. Cell. Neurosci. 2003, 24, 476–488. [Google Scholar] [CrossRef]
- Hill, R.A.; Patel, K.D.; Medved, J.; Reiss, A.M.; Nishiyama, A. NG2 cells in white matter but not gray matter proliferate in response to PDGF. J. Neurosci. 2013, 33, 14558–14566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lentferink, D.H.; Jongsma, J.M.; Werkman, I.; Baron, W. Grey matter OPCs are less mature and less sensitive to IFNγ than white matter OPCs: consequences for remyelination. Sci. Rep. 2018, 8, 2113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dimou, L.; Simon, C.; Kirchhoff, F.; Takebayashi, H.; Götz, M. Progeny of Olig2-expressing progenitors in the gray and white matter of the adult mouse cerebral cortex. J. Neurosci. 2008, 28, 10434–10442. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.H.; Fukaya, M.; Yang, J.K.; Rothstein, J.D.; Bergles, D.E. NG2+ CNS glial progenitors remain committed to the oligodendrocyte lineage in postnatal life and following neurodegeneration. Neuron 2010, 68, 668–681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rivers, L.E.; Young, K.M.; Rizzi, M.; Jamen, F. PDGFRA/NG2 glia generate myelinating oligodendrocytes and piriform projection neurons in adult mice. Nat. Neurosci. 2008, 11, 1–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hughes, E.G.; Kang, S.H.; Fukaya, M.; Bergles, D.E. Oligodendrocyte progenitors balance growth with self-repulsion to achieve homeostasis in the adult brain. Nat. Neurosci. 2013, 16, 668–676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lachapelle, F.; Gumpel, M.; Baulac, M.; Jacque, C.; Duc, P.; Baumann, N. Transplantation of CNS Fragments into the Brain of Shiverer Mutant Mice: Extensive Myelination by Implanted Oligodendrocytes. Dev. Neurosci. 1983, 6, 325–334. [Google Scholar] [CrossRef] [PubMed]
- Tsai, H.H.; Niu, J.; Munji, R.; Davalos, D.; Chang, J.; Zhang, H.; Tien, A.C.; Kuo, C.J.; Chan, J.R.; Daneman, R.; et al. Oligodendrocyte precursors migrate along vasculature in the developing nervous system. Science 2016, 351, 379–384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niu, J.; Tsai, H.H.; Hoi, K.K.; Huang, N.; Yu, G.; Kim, K.; Baranzini, S.E.; Xiao, L.; Chan, J.R.; Fancy, S.P.J. Aberrant oligodendroglial–vascular interactions disrupt the blood–brain barrier, triggering CNS inflammation. Nat. Neurosci. 2019, 22, 709–718. [Google Scholar] [CrossRef] [PubMed]
- Miller, R.H.; Payne, J.; Milner, L.; Zhang, H.; Orentas, D.M. Spinal cord oligodendrocytes develop from a limited number of migratory, highly proliferative precursors. J. Neurosci. Res. 1997, 50, 157–168. [Google Scholar] [CrossRef]
- Fok-Seang, J.; Miller, R.H. Distribution and differentiation of A2B5+ glial precursors in the developing rat spinal cord. J Neurosci Res 1994, 37, 219–235. [Google Scholar] [CrossRef] [PubMed]
- Raff, M.C.; Abney, E.R.; Miller, R.H. Two glial cell lineages diverge prenatally in rat optic nerve. Dev. Biol. 1984, 106, 53–60. [Google Scholar] [CrossRef]
- Hughes, S.M.; Lillien, L.E.; Raff, M.C.; Rohrer, H.; Sendtner, M. Ciliary neurotrophic factor induces type-2 astrocyte differentiation in culture. Nature 1988, 335, 70–73. [Google Scholar] [CrossRef] [PubMed]
- Noble, M.; Murray, K.; Stroobant, P.; Waterfield, M.D.; Riddle, P. Platelet-derived growth factor promotes division and motility and inhibits premature differentiation of the oligodendrocyte/type-2 astrocyte progenitor cell. Nature 1988, 333, 560–562. [Google Scholar] [CrossRef] [PubMed]
- Yeh, H.J.; Ruit, K.G.; Wang, Y.X.; Parks, W.C.; Snider, W.D.; Deuel, T.F. PDGF A-chain gene is expressed by mammalian neurons during development and in maturity. Cell 1991, 64, 209–216. [Google Scholar] [CrossRef]
- Calver, A.R.; Hall, A.C.; Yu, W.P.; Walsh, F.S.; Heath, J.K.; Betsholtz, C.; Richardson, W.D. Oligodendrocyte population dynamics and the role of PDGF in vivo. Neuron 1998, 20, 869–882. [Google Scholar] [CrossRef] [Green Version]
- Sommer, I.; Schachner, M. Monoclonal antibodies (O1 to O4) to oligodendrocyte cell surfaces: An immunocytological study in the central nervous system. Dev. Biol. 1981, 83, 311–327. [Google Scholar] [CrossRef]
- Braun, P.E.; Sandillon, F.; Edwards, A.; Matthieu, J.-M.; Privat, A. lmmunocytochemical Localization by Electron Microscopy of 2’,3’-Cyclic Nucleotide 3’-Phosphodiesterase in Developing Oligodendrocytes of Normal and Mutant Brain. J. Neurosci. 1988, 8, 3057–3066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scherer, S.S.; Braun, P.E.; Grinspan, J.; Collarini, E.; Wang, D.Y.; Kamholz, J. Differential regulation of the 2′,3′-cyclic nucleotide 3′-phosphodiesterase gene during oligodendrocyte development. Neuron 1994, 12, 1363–1375. [Google Scholar] [CrossRef]
- Jakovcevski, I.; Filipovic, R.; Mo, Z.; Rakic, S.; Zecevic, N. Oligodendrocyte development and the onset of myelination in the human fetal brain. Front. Neuroanat. 2009, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brunner, C.; Lassmann, H.; Waehneldt, T.V.; Matthieu, J.-M.; Linington, C. Differential Ultrastructural Localization of Myelin Basic Protein, Myelin/Oligodendroglial Glycoprotein, and 2′,3′-Cyclic Nucleotide 3′-Phosphodiesterase in the CNS of Adult Rats. J. Neurochem. 1989, 52, 296–304. [Google Scholar] [CrossRef] [PubMed]
- Linnington, C.; Webb, M.; Woodhams, P.L. A novel myelin-associated glycoprotein defined by a mouse monoclonal antibody. J. Neuroimmunol. 1984, 6, 387–396. [Google Scholar] [CrossRef]
- Barbarese, E.; Barry, C.; Chou, C.J.; Goldstein, D.J.; Nakos, G.A.; Hyde-DeRuyscher, R.; Scheld, K.; Carson, J.H. Expression and Localization of Myelin Basic Protein in Oligodendrocytes and Transfected Fibroblasts. J. Neurochem. 1988, 51, 1737–1745. [Google Scholar] [CrossRef] [PubMed]
- Michalski, J.P.; Anderson, C.; Beauvais, A.; de Repentigny, Y.; Kothary, R. The proteolipid protein promoter drives expression outside of the oligodendrocyte lineage during embryonic and early postnatal development. PLoS ONE 2011, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trapp, B.D. Myelin-Associated Glycoprotein Location and Potential Functions. Ann. N. Y. Acad. Sci. 1990, 605, 29–43. [Google Scholar] [CrossRef] [PubMed]
- Raff, M.C.; Mirsky, R.; Fields, K.L.; Lisak, R.P.; Dorfman, S.H.; Silberberg, D.H.; Gregson, N.A.; Leibowitz, S.; Kennedy, M.C. Galactocerebroside is a specific cell-surface antigenic marker for oligodendrocytes in culture. Nature 1978, 274, 813–816. [Google Scholar] [CrossRef] [PubMed]
- Dubois-Dalcq, M.; Behar, T.; Hudson, L.; Lazzarini, R.A. Emergence of three myelin proteins in oligodendrocytes cultured without neurons. J. Cell Biol. 1986, 102, 384–392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harlow, D.E.; Saul, K.E.; Culp, C.M.; Vesely, E.M.; Macklin, W.B. Expression of proteolipid protein gene in spinal cord stem cells and early oligodendrocyte progenitor cells is dispensable for normal cell migration and myelination. J. Neurosci. 2014, 34, 1333–1343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Q.; Wang, S.; Anderson, D.J. Identification of a novel family of oligodendrocyte lineage-specific basic helix-loop-helix transcription factors. Neuron 2000, 25, 331–343. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Q.; Anderson, D.J. The bHLH transcription factors OLIG2 and OLIG1 couple neuronal and glial subtype specification. Cell 2002, 109, 61–73. [Google Scholar] [CrossRef] [Green Version]
- Rowitch, D.H. Glial specification in the vertebrate neural tube. Nat. Rev. Neurosci. 2004, 5, 409–419. [Google Scholar] [CrossRef] [PubMed]
- Wegener, A.; Deboux, C.; Bachelin, C.; Frah, M.; Kerninon, C.; Seilhean, D.; Weider, M.; Wegner, M.; Nait-Oumesmar, B. Gain of Olig2 function in oligodendrocyte progenitors promotes remyelination. Brain 2015, 138, 120–135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xin, M.; Yue, T.; Ma, Z.; Wu, F.; Gow, A.; Lu, Q.R. Myelinogenesis and axonal recognition by oligodendrocytes in brain are uncoupled in Olig1-null mice. J. Neurosci. 2005, 25, 1354–1365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, J.; Bercury, K.K.; Ahrendsen, J.T.; Macklin, W.B. Olig1 function is required for oligodendrocyte differentiation in the mouse brain. J. Neurosci. 2015, 35, 4386–4402. [Google Scholar] [CrossRef] [PubMed]
- Arnett, H.A.; Fancy, S.P.J.; Alberta, J.A.; Zhao, C.; Plant, S.R.; Kaing, S.; Raine, C.S.; Rowitch, D.H.; Franklin, R.J.M.; Stiles, C.D. bHLH Transcription Factor Olig1 Is Required to Repair Demyelinated Lesions in the CNS. Science 2004, 306, 2111–2115. [Google Scholar] [CrossRef] [PubMed]
- Kuhlbrodt, K.; Herbarth, B.; Sock, E.; Hermans-Borgmeyer, I.; Wegner, M. Sox10, a novel transcriptional modulator in glial cells. J. Neurosci. 1998, 18, 237–250. [Google Scholar] [CrossRef] [PubMed]
- Pozniak, C.D.; Langseth, A.J.; Dijkgraaf, G.J.P.; Choe, Y.; Werb, Z.; Pleasure, S.J. Sox10 directs neural stem cells toward the oligodendrocyte lineage by decreasing Suppressor of Fused expression. Proc. Natl. Acad. Sci. USA 2010, 107, 21795–21800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stolt, C.C.; Rehberg, S.; Ader, M.; Lommes, P.; Riethmacher, D.; Schachner, M.; Bartsch, U.; Wegner, M. Terminal differentiation of myelin-forming oligodendrocytes depends on the transcription factor Sox10. Genes Dev. 2002, 16, 165–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stolt, C.C.; Lommes, P.; Friedrich, R.P.; Wegner, M. Transcription factors Sox8 and Sox10 perform non-equivalent roles during oligodendrocyte development despite functional redundancy. Development 2004, 131, 2349–2358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, Y.; Cai, J.; Wu, Y.; Wu, R.; Lee, J.; Fu, H.; Rao, M.; Sussel, L.; Rubenstein, J.; Qiu, M. Control of oligodendrocyte differentiation by the Nkx2.2 homeodomain transcription factor. Development 2001, 128, 2723–2733. [Google Scholar] [PubMed]
- Zhu, Q.; Zhao, X.; Zheng, K.; Li, H.; Huang, H.; Zhang, Z.; Mastracci, T.; Wegner, M.; Chen, Y.; Sussel, L.; et al. Genetic evidence that Nkx2.2 and Pdgfra are major determinants of the timing of oligodendrocyte differentiation in the developing CNS. Development 2014, 141, 548–555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, X.; Bergles, D.E.; Nishiyama, A. NG2 cells generate both oligodendrocytes and gray matter astrocytes. Development 2008, 135, 145–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woodhoo, A.; Sommer, L. Development of the Schwann cell lineage: From the neural crest to the myelinated nerve. Glia 2008, 56, 1481–1490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jessen, K.R.; Mirsky, R. The origin and development of glial cells in peripheral nerves. Nat. Rev. Neurosci. 2005, 6, 671–682. [Google Scholar] [CrossRef] [PubMed]
- Salzer, J.L. Schwann cell myelination. Cold Spring Harb. Perspect. Biol. 2015, 7, a020529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harty, B.L.; Coelho, F.; Pease-Raissi, S.E.; Mogha, A.; Ackerman, S.D.; Herbert, A.L.; Gereau, R.W.; Golden, J.P.; Lyons, D.A.; Chan, J.R.; et al. Myelinating Schwann cells ensheath multiple axons in the absence of E3 ligase component Fbxw7. Nat. Commun. 2019, 10, 2976. [Google Scholar] [CrossRef] [PubMed]
- Kucenas, S.; Wang, W.-D.; Knapik, E.W.; Appel, B. A selective glial barrier at motor axon exit points prevents oligodendrocyte migration from the spinal cord. J. Neurosci. 2009, 29, 15187–15194. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.J.; Morris, A.D.; Welsh, T.G.; Kucenas, S. Contact-Mediated Inhibition Between Oligodendrocyte Progenitor Cells and Motor Exit Point Glia Establishes the Spinal Cord Transition Zone. PLoS Biol. 2014, 12, e1001961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fontenas, L.; Welsh, T.G.; Piller, M.; Coughenour, P.; Gandhi, A.V.; Prober, D.A.; Kucenas, S. The Neuromodulator Adenosine Regulates Oligodendrocyte Migration at Motor Exit Point Transition Zones. Cell Rep. 2019, 27, 115–128.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raine, C.S. Morphology of Myelin and Myelination. In Myelin; Springer US: Boston, MA, USA, 1984; pp. 1–50. [Google Scholar]
- Rosenbluth, J. A brief history of myelinated nerve fibers: one hundred and fifty years of controversy. J. Neurocytol. 1999, 28, 251–262. [Google Scholar] [CrossRef] [PubMed]
- Morell, P.; Quarles, R.H. The Myelin Sheath. 1999. Available online: https://www.ncbi.nlm.nih.gov/books/NBK27954/ (accessed on 9 November 2019).
- Baraban, M.; Koudelka, S.; Lyons, D.A. Ca2+ activity signatures of myelin sheath formation and growth in vivo. Nat. Neurosci. 2018, 21, 19–23. [Google Scholar] [CrossRef] [PubMed]
- Nave, K.-A.; Werner, H.B. Myelination of the Nervous System: Mechanisms and Functions. Annu. Rev. Cell Dev. Biol. 2014, 30, 503–533. [Google Scholar] [CrossRef] [PubMed]
- Kirby, B.B.; Takada, N.; Latimer, A.J.; Shin, J.; Carney, T.J.; Kelsh, R.N.; Appel, B. In vivo time-lapse imaging shows dynamic oligodendrocyte progenitor behavior during zebrafish development. Nat. Neurosci. 2006, 9, 1506–1511. [Google Scholar] [CrossRef] [PubMed]
- Simons, M.; Trotter, J. Wrapping it up: The cell biology of myelination. Curr. Opin. Neurobiol. 2007, 17, 533–540. [Google Scholar] [CrossRef] [PubMed]
- Geren, B.B.; Schmitt, F.O. The structure of the Schwann cell and its relation to the axon in certain invertebrate nerve fibers. Proc. Natl. Acad. Sci. USA 1954, 40, 863–870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sobottka, B.; Ziegler, U.; Kaech, A.; Becher, B.; Goebels, N. CNS live imaging reveals a new mechanism of myelination: The liquid croissant model. Glia 2011, 59, 1841–1849. [Google Scholar] [CrossRef] [PubMed]
- Snaidero, N.; Möbius, W.; Czopka, T.; Hekking, L.H.P.; Mathisen, C.; Verkleij, D.; Goebbels, S.; Edgar, J.; Merkler, D.; Lyons, D.A.; et al. Myelin Membrane Wrapping of CNS Axons by PI(3,4,5)P3-Dependent Polarized Growth at the Inner Tongue. Cell 2014, 156, 277–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friede, R.L. Control of myelin formation by axon caliber. (With a model of the control mechanism). J. Comp. Neurol. 1972, 144, 233–252. [Google Scholar] [CrossRef] [PubMed]
- Windebank, A.J.; Wood, P.; Bunge, R.P.; Dyck, P.J. Myelination determines the caliber of dorsal root ganglion neurons in culture. J. Neurosci. 1985, 5, 1563–1569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.; Leach, M.K.; Redmond, S.A.; Chong, S.Y.C.; Mellon, S.H.; Tuck, S.J.; Feng, Z.-Q.; Corey, J.M.; Chan, J.R. A culture system to study oligodendrocyte myelination processes using engineered nanofibers. Nat. Methods 2012, 9, 917–922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sánchez, I.; Hassinger, L.; Paskevich, P.A.; Shine, H.D.; Nixon, R.A. Oligodendroglia regulate the regional expansion of axon caliber and local accumulation of neurofilaments during development independently of myelin formation. J. Neurosci. 1996, 16, 5095–5105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sánchez, I.; Hassinger, L.; Sihag, R.K.; Cleveland, D.W.; Mohan, P.; Nixon, R.A. Local Control of Neurofilament Accumulation during Radial Growth of Myelinating Axons in Vivo. J. Cell Biol. 2000, 151, 1013–1024. [Google Scholar] [CrossRef] [PubMed]
- Nave, K.-A.; Salzer, J.L. Axonal regulation of myelination by neuregulin 1. Curr. Opin. Neurobiol. 2006, 16, 492–500. [Google Scholar] [CrossRef] [PubMed]
- Waxman, S.G.; Ritchie, J.M. Molecular dissection of the myelinated axon. Ann. Neurol. 1993, 33, 121–136. [Google Scholar] [CrossRef] [PubMed]
- Seidl, A.H. Regulation of conduction time along axons. Neuroscience 2014, 276, 126–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bercury, K.K.; Macklin, W.B. Dynamics and Mechanisms of CNS Myelination. Dev. Cell 2015, 32, 447–458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.; Morrison, B.M.; Li, Y.; Lengacher, S.; Farah, M.H.; Hoffman, P.N.; Liu, Y.; Tsingalia, A.; Jin, L.; Zhang, P.-W.; et al. Oligodendroglia metabolically support axons and contribute to neurodegeneration. Nature 2012, 487, 443–448. [Google Scholar] [CrossRef] [PubMed]
- Fünfschilling, U.; Supplie, L.M.; Mahad, D.; Boretius, S.; Saab, A.S.; Edgar, J.; Brinkmann, B.G.; Kassmann, C.M.; Tzvetanova, I.D.; Möbius, W.; et al. Glycolytic oligodendrocytes maintain myelin and long-term axonal integrity. Nature 2012, 485, 517–521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ravera, S.; Bartolucci, M.; Calzia, D.; Aluigi, M.G.; Ramoino, P.; Morelli, A.; Panfoli, I. Tricarboxylic acid cycle-sustained oxidative phosphorylation in isolated myelin vesicles. Biochimie 2013, 95, 1991–1998. [Google Scholar] [CrossRef] [PubMed]
- Assinck, P.; Duncan, G.J.; Plemel, J.R.; Lee, M.J.; Stratton, J.A.; Manesh, S.B.; Liu, J.; Ramer, L.M.; Kang, S.H.; Bergles, D.E.; et al. Myelinogenic Plasticity of Oligodendrocyte Precursor Cells following Spinal Cord Contusion Injury. J. Neurosci. 2017, 37, 8635–8654. [Google Scholar] [CrossRef] [PubMed]
- Itoyama, Y.; Ohnishi, A.; Tateishi, J.; Kuroiwa, Y.; Webster, H.D. Spinal cord multiple sclerosis lesions in Japanese patients: Schwann cell remyelination occurs in areas that lack glial fibrillary acidic protein (GFAP). Acta Neuropathol. 1985, 65, 217–223. [Google Scholar] [CrossRef] [PubMed]
- Baerwald, K.D.; Popko, B. Developing and mature oligodendrocytes respond differently to the immune cytokine interferon-gamma. J. Neurosci. Res. 1998, 52, 230–239. [Google Scholar] [CrossRef]
- Arnett, H.A.; Mason, J.; Marino, M.; Suzuki, K.; Matsushima, G.K.; Ting, J.P.-Y. TNFα promotes proliferation of oligodendrocyte progenitors and remyelination. Nat. Neurosci. 2001, 4, 1116–1122. [Google Scholar] [CrossRef] [PubMed]
- Bonora, M.; De Marchi, E.; Patergnani, S.; Suski, J.M.; Celsi, F.; Bononi, A.; Giorgi, C.; Marchi, S.; Rimessi, A.; Duszyński, J.; et al. Tumor necrosis factor-α impairs oligodendroglial differentiation through a mitochondria-dependent process. Cell Death Differ. 2014, 21, 1198–1208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, Z.; Wang, C.; Zepp, J.; Wu, L.; Sun, K.; Zhao, J.; Chandrasekharan, U.; DiCorleto, P.E.; Trapp, B.D.; Ransohoff, R.M.; et al. Act1 mediates IL-17–induced EAE pathogenesis selectively in NG2+ glial cells. Nat. Neurosci. 2013, 16, 1401–1408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Falcão, A.M.; van Bruggen, D.; Marques, S.; Meijer, M.; Jäkel, S.; Agirre, E.; Samudyata; Floriddia, E.M.; Vanichkina, D.P.; ffrench-Constant, C.; et al. Disease-specific oligodendrocyte lineage cells arise in multiple sclerosis. Nat. Med. 2018, 24, 1837–1844. [Google Scholar]
- Kirby, L.; Jin, J.; Cardona, J.G.; Smith, M.D.; Martin, K.A.; Wang, J.; Strasburger, H.; Herbst, L.; Alexis, M.; Karnell, J.; et al. Oligodendrocyte precursor cells present antigen and are cytotoxic targets in inflammatory demyelination. Nat. Commun. 2019, 10, 3887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dincman, T.A.; Beare, J.E.; Ohri, S.S.; Whittemore, S.R. Isolation of cortical mouse oligodendrocyte precursor cells. J. Neurosci. Methods 2012, 209, 219–226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emery, B.; Dugas, J.C. Purification of oligodendrocyte lineage cells from mouse cortices by immunopanning. Cold Spring Harb. Protoc. 2013, 2013, 854–868. [Google Scholar] [CrossRef] [PubMed]
- Flores-Obando, R.E.; Freidin, M.M.; Abrams, C.K. Rapid and specific immunomagnetic isolation of mouse primary oligodendrocytes. J. Vis. Exp. 2018, 2018. [Google Scholar] [CrossRef] [PubMed]
- Pang, Y.; Zheng, B.; Kimberly, S.L.; Cai, Z.; Rhodes, P.G.; Lin, R.C.S. Neuron-oligodendrocyte myelination co-culture derived from embryonic rat spinal cord and cerebral cortex. Brain Behav. 2012, 2, 53–67. [Google Scholar] [CrossRef] [PubMed]
- Mei, F.; Fancy, S.P.J.; Shen, Y.A.A.; Niu, J.; Zhao, C.; Presley, B.; Miao, E.; Lee, S.; Mayoral, S.R.; Redmond, S.A.; et al. Micropillar arrays as a high-throughput screening platform for therapeutics in multiple sclerosis. Nat. Med. 2014, 20, 954–960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Louis, J.C.; Magal, E.; Muir, D.; Manthorpe, M.; Varon, S. CG-4, A new bipotential glial cell line from rat brain, is capable of differentiating in vitro into either mature oligodendrocytes or type-2 astrocytes. J. Neurosci. Res. 1992, 31, 193–204. [Google Scholar] [CrossRef] [PubMed]
- Richter-Landsberg, C.; Heinrich, M. OLN-93: A new permanent oligodendroglia cell line derived from primary rat brain glial cultures. J. Neurosci. Res. 1996, 45, 161–173. [Google Scholar] [CrossRef]
- Jungl, M.; Kramer’, E.; Grzenkowskil, M.; Tang2, K.; Biakemore2, W.; Aguzzi3, A.; Khazaie4, K.; Chiichlia4, K.; Von Blankenfeld5, G.; Kettenmann5, H.; et al. Lines of Murine Oligodendroglial Precursor Cells Immortalized by an Activated neu Tvrosine Kinase Show Distinct Degrees of Interaction with Axons In Wtro and In Wvo. EJN 1995, 7, 1245–1265. [Google Scholar]
- Van Tilborg, E.; de Theije, C.G.M.; van Hal, M.; Wagenaar, N.; de Vries, L.S.; Benders, M.J.; Rowitch, D.H.; Nijboer, C.H. Origin and dynamics of oligodendrocytes in the developing brain: Implications for perinatal white matter injury. Glia 2018, 66, 221–238. [Google Scholar] [CrossRef] [PubMed]
- Chanoumidou, K.; Mozafari, S.; Baron-Van Evercooren, A.; Kuhlmann, T. Stem cell derived oligodendrocytes to study myelin diseases. Glia 2019, glia.23733. [Google Scholar] [CrossRef] [PubMed]
- Medina-Rodríguez, E.M.; Arenzana, F.J.; Bribián, A.; De Castro, F. Protocol to isolate a large amount of functional oligodendrocyte precursor cells from the cerebral cortex of adult mice and humans. PLoS ONE 2013, 8, e81620. [Google Scholar]
- De Groot, J.A.; Montagne, L.; Janssen, I.; Ravid, R.; Van Der Valk, P.; Veerhuis, R. Isolation and characterization of adult microglial cells and oligodendrocytes derived from postmortem human brain tissue. Brain Res. Protoc. 2000, 5, 85–94. [Google Scholar] [CrossRef]
- Takahashi, K.; Yamanaka, S. Induction of Pluripotent Stem Cells from Mouse Embryonic and Adult Fibroblast Cultures by Defined Factors. Cell 2006, 126, 663–676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Bates, J.; Li, X.; Schanz, S.; Chandler-Militello, D.; Levine, C.; Maherali, N.; Studer, L.; Hochedlinger, K.; Windrem, M.; et al. Human iPSC-derived oligodendrocyte progenitor cells can myelinate and rescue a mouse model of congenital hypomyelination. Cell Stem Cell 2013, 12, 252–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ehrlich, M.; Mozafari, S.; Glatza, M.; Starost, L.; Velychko, S.; Hallmann, A.L.; Cui, Q.L.; Schambach, A.; Kim, K.P.; Bachelin, C.; et al. Rapid and efficient generation of oligodendrocytes from human induced pluripotent stem cells using transcription factors. Proc. Natl. Acad. Sci. USA 2017, 114, E2243–E2252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-León, J.A.; Kumar, M.; Boon, R.; Chau, D.; One, J.; Wolfs, E.; Eggermont, K.; Berckmans, P.; Gunhanlar, N.; de Vrij, F.; et al. SOX10 Single Transcription Factor-Based Fast and Efficient Generation of Oligodendrocytes from Human Pluripotent Stem Cells. Stem Cell Rep. 2018, 10, 655–672. [Google Scholar]
- Dittmer, M.; Young, A.; O’Hagan, T.; Eleftheriadis, G.; Bankhead, P.; Dombrowski, Y.; Medina, R.J.; Fitzgerald, D.C. Characterization of a murine mixed neuron-glia model and cellular responses to regulatory T cell-derived factors. Mol Brain 2018, 11, 25. [Google Scholar] [CrossRef] [PubMed]
- Harrer, M.D.; von Büdingen, H.C.; Stoppini, L.; Alliod, C.; Pouly, S.; Fischer, K.; Goebels, N. Live imaging of remyelination after antibody-mediated demyelination in an ex-vivo model for immune mediated CNS damage. Exp. Neurol. 2009, 216, 431–438. [Google Scholar] [CrossRef] [PubMed]
- Marton, R.M.; Miura, Y.; Sloan, S.A.; Li, Q.; Revah, O.; Levy, R.J.; Huguenard, J.R.; Pașca, S.P. Differentiation and maturation of oligodendrocytes in human three-dimensional neural cultures. Nat. Neurosci. 2019, 22, 484–491. [Google Scholar] [CrossRef] [PubMed]
- Shibata, S.; Yasuda, A.; Renault-Mihara, F.; Suyama, S.; Katoh, H.; Inoue, T.; Inoue, Y.U.; Nagoshi, N.; Sato, M.; Nakamura, M.; et al. Sox10-Venus mice: A new tool for real-time labeling of neural crest lineage cells and oligodendrocytes. Mol. Brain 2010, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, X.; Chittajallu, R.; Belachew, S.; Anderson, S.; McBain, C.J.; Gallo, V. Expression of the green fluorescent protein in the oligodendrocyte lineage: A transgenic mouse for developmental and physiological studies. J. Neurosci. Res. 2002, 70, 529–545. [Google Scholar] [CrossRef] [PubMed]
- Mallon, B.S.; Shick, H.E.; Kidd, G.J.; Macklin, W.B. Proteolipid Promoter Activity Distinguishes Two Populations of NG2-Positive Cells throughout Neonatal Cortical Development. J. Neurosci. 2002, 22, 876–885. [Google Scholar] [CrossRef] [PubMed]
- Almeida, R.G.; Czopka, T.; Ffrench-Constant, C.; Lyons, D.A. Individual axons regulate the myelinating potential of single oligodendrocytes in vivo. Development 2011, 138, 4443–4450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, J.; Park, H.-C.; Topczewska, J.M.; Mawdsley, D.J.; Appel, B. Neural cell fate analysis in zebrafish using olig2 BAC transgenics. Methods Cell Sci. 2003, 25, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Howe, K.; Clark, M.D.; Torroja, C.F.; Torrance, J.; Berthelot, C.; Muffato, M.; Collins, J.E.; Humphray, S.; McLaren, K.; Matthews, L.; et al. The zebrafish reference genome sequence and its relationship to the human genome. Nature 2013, 496, 498–503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nawaz, S.; Schweitzer, J.; Jahn, O.; Werner, H.B. Molecular evolution of myelin basic protein, an abundant structural myelin component. Glia 2013, 61, 1364–1377. [Google Scholar] [CrossRef] [PubMed]
- Early, J.J.; Cole, K.L.; Williamson, J.M.; Swire, M.; Kamadurai, H.; Muskavitch, M.; Lyons, D.A. An automated high-resolution in vivo screen in zebrafish to identify chemical regulators of myelination. Elife 2018, 7. [Google Scholar] [CrossRef] [PubMed]
- Ransohoff, R.M. Animal models of multiple sclerosis: The good, the bad and the bottom line. Nat. Neurosci. 2012, 15, 1074–1077. [Google Scholar] [CrossRef] [PubMed]
- Croxford, A.L.; Kurschus, F.C.; Waisman, A. Mouse models for multiple sclerosis: Historical facts and future implications. Biochim. Biophys. Acta Mol. Basis Dis. 2011, 1812, 177–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blakemore, W.F. Pattern of remyelination in the CNS. Nature 1974, 249, 577–578. [Google Scholar] [CrossRef] [PubMed]
- Hall, S.M. The effect of injections of lysophosphatidyl choline into white matter of the adult mouse spinal cord. J. Cell Sci. 1972, 10, 535–546. [Google Scholar] [PubMed]
- Kotter, M.R.; Setzu, A.; Sim, F.J.; Van Rooijen, N.; Franklin, R.J.M. Macrophage depletion impairs oligodendrocyte remyelination following lysolecithin-induced demyelination. Glia 2001, 35, 204–212. [Google Scholar] [CrossRef] [PubMed]
- Kipp, M.; Clarner, T.; Dang, J.; Copray, S.; Beyer, C. The cuprizone animal model: New insights into an old story. Acta Neuropathol. 2009, 118, 723–736. [Google Scholar] [CrossRef] [PubMed]
- Matsushima, G.K.; Morell, P. The Neurotoxicant, Cuprizone, as a Model to Study Demyelination and Remyelination in the Central Nervous System. Brain Pathol. 2006, 11, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Sachs, H.H.; Bercury, K.K.; Popescu, D.C.; Narayanan, S.P.; Macklin, W.B. A new model of Cuprizone-Mediated demyelination/remyelination. ASN Neuro. 2014, 6. [Google Scholar] [CrossRef] [PubMed]
- Woodruff, R.H.; Fruttiger, M.; Richardson, W.D.; Franklin, R.J.M. Platelet-derived growth factor regulates oligodendrocyte progenitor numbers in adult CNS and their response following CNS demyelination. Mol. Cell. Neurosci. 2004, 25, 252–262. [Google Scholar] [CrossRef] [PubMed]
- Fancy, S.P.J.; Baranzini, S.E.; Zhao, C.; Yuk, D.I.; Irvine, K.A.; Kaing, S.; Sanai, N.; Franklin, R.J.M.; Rowitch, D.H. Dysregulation of the Wnt pathway inhibits timely myelination and remyelination in the mammalian CNS. Genes Dev. 2009, 23, 1571–1585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rüther, B.J.; Scheld, M.; Dreymueller, D.; Clarner, T.; Kress, E.; Brandenburg, L.O.; Swartenbroekx, T.; Hoornaert, C.; Ponsaerts, P.; Fallier-Becker, P.; et al. Combination of cuprizone and experimental autoimmune encephalomyelitis to study inflammatory brain lesion formation and progression. Glia 2017, 65, 1900–1913. [Google Scholar] [CrossRef] [PubMed]
- Sampaio-Baptista, C.; Johansen-Berg, H. White Matter Plasticity in the Adult Brain. Neuron 2017, 96, 1239–1251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McKenzie, I.A.; Ohayon, D.; Li, H.; Paes de Faria, J.; Emery, B.; Tohyama, K.; Richardson, W.D. Motor skill learning requires active central myelination. Science 2014, 346, 318–322. [Google Scholar] [CrossRef] [PubMed]
- Butts, B.D.; Houde, C.; Mehmet, H. Maturation-dependent sensitivity of oligodendrocyte lineage cells to apoptosis: Implications for normal development and disease. Cell Death Differ. 2008, 15, 1178–1186. [Google Scholar] [CrossRef] [PubMed]
- Juurlink, B.H.J.; Thorburne, S.K.; Hertz, L. Peroxide-scavenging deficit underlies oligodendrocyte susceptibility to oxidative stress. Glia 1998, 22, 371–378. [Google Scholar] [CrossRef]
- McTigue, D.M.; Tripathi, R.B. The life, death, and replacement of oligodendrocytes in the adult CNS. J. Neurochem. 2008, 107, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Matute, C.; Torre, I.; Pérez-Cerdá, F.; Pérez-Samartín, A.; Alberdi, E.; Etxebarria, E.; Arranz, A.M.; Ravid, R.; Rodríguez-Antigüedad, A.; Sánchez-Gómez, M.V.; et al. P2X 7 receptor blockade prevents ATP excitotoxicity in oligodendrocytes and ameliorates experimental autoimmune encephalomyelitis. J. Neurosci. 2007, 27, 9525–9533. [Google Scholar] [CrossRef] [PubMed]
- Matute, C.; Sánchez-Gómez, M.V.; Martínez-Millán, L.; Miledi, R. Glutamate receptor-mediated toxicity in optic nerve oligodendrocytes. Proc. Natl. Acad. Sci. USA 1997, 94, 8830–8835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Traka, M.; Podojil, J.R.; McCarthy, D.P.; Miller, S.D.; Popko, B. Oligodendrocyte death results in immune-mediated CNS demyelination. Nat. Neurosci. 2016, 19, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Fancy, S.P.J.; Harrington, E.P.; Yuen, T.J.; Silbereis, J.C.; Zhao, C.; Baranzini, S.E.; Bruce, C.C.; Otero, J.J.; Huang, E.J.; Nusse, R.; et al. Axin2 as regulatory and therapeutic target in newborn brain injury and remyelination. Nat. Neurosci. 2011, 14, 1009–1016. [Google Scholar] [CrossRef] [PubMed]
- Torii, T.; Miyamoto, Y.; Yamauchi, J.; Tanoue, A. Pelizaeus-Merzbacher disease: Cellular pathogenesis and pharmacologic therapy. Pediatr. Int. 2014, 56, 659–666. [Google Scholar] [CrossRef] [PubMed]
- Simons, M.; Krämer, E.M.; Macchi, P.; Rathke-Hartlieb, S.; Trotter, J.; Nave, K.A.; Schulz, J.B. Overexpression of the myelin proteolipid protein leads to accumulation of cholesterol and proteolipid protein in endosomes/lysosomes: Implications for Pelizaeus-Merzbacher disease. J. Cell Biol. 2002, 157, 327–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McAleese, K.E.; Walker, L.; Graham, S.; Moya, E.L.J.; Johnson, M.; Erskine, D.; Colloby, S.J.; Dey, M.; Martin-Ruiz, C.; Taylor, J.P.; et al. Parietal white matter lesions in Alzheimer’s disease are associated with cortical neurodegenerative pathology, but not with small vessel disease. Acta Neuropathol. 2017, 134, 459–473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Behrendt, G.; Baer, K.; Buffo, A.; Curtis, M.A.; Faull, R.L.; Rees, M.I.; Götz, M.; Dimou, L. Dynamic changes in myelin aberrations and oligodendrocyte generation in chronic amyloidosis in mice and men. Glia 2013, 61, 273–286. [Google Scholar] [CrossRef] [PubMed]
- Desai, M.K.; Mastrangelo, M.A.; Ryan, D.A.; Sudol, K.L.; Narrow, W.C.; Bowers, W.J. Early oligodendrocyte/myelin pathology in Alzheimer’s disease mice constitutes a novel therapeutic target. Am. J. Pathol. 2010, 177, 1422–1435. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Chen, S.; Ahmed, S.H.; Chen, H.; Ku, G.; Goldberg, M.P.; Hsu, C.Y. Amyloid-beta peptides are cytotoxic to oligodendrocytes. J. Neurosci. 2001, 21, 1–5. [Google Scholar] [CrossRef]
- Fischer, F.U.; Wolf, D.; Scheurich, A.; Fellgiebel, A. Altered whole-brain white matter networks in preclinical Alzheimer’s disease. NeuroImage Clin. 2015, 8, 660–666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braak, H.; Braak, E. Development of Alzheimer-related neurofibrillary changes in the neocortex inversely recapitulates cortical myelogenesis. Acta Neuropathol. 1996, 92, 197–201. [Google Scholar] [CrossRef] [PubMed]
- Reisberg, B.; Franssen, E.H.; Souren, L.E.M.; Auer, S.R.; Akram, I.; Kenowsky, S. Evidence and mechanisms of retrogenesis in Alzheimer’s and other dementias: Management and treatment import. Am. J. Alzheimers. Dis. Other Demen. 2002, 17, 202–212. [Google Scholar] [CrossRef] [PubMed]
- Fields, R.D. ScienceDirect-Trends in Neurosciences: White matter in learning, cognition and psychiatric disorders. Trends Neurosci. 2009, 31, 361–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, N.; Sibille, E. A putative functional role for oligodendrocytes in mood regulation. Transl. Psychiatry 2012, 2, e109. [Google Scholar] [CrossRef] [PubMed]
- Rajkowska, G. Cell pathology in bipolar disorder. Bipolar Disord. 2002, 4, 105–116. [Google Scholar] [CrossRef] [PubMed]
- Vostrikov, V.M.; Uranova, N.A.; Orlovskaya, D.D. Deficit of perineuronal oligodendrocytes in the prefrontal cortex in schizophrenia and mood disorders. Schizophr. Res. 2007, 94, 273–280. [Google Scholar] [CrossRef] [PubMed]
- Tkachev, D.; Mimmack, M.L.; Ryan, M.M.; Wayland, M.; Freeman, T.; Jones, P.B.; Starkey, M.; Webster, M.J.; Yolken, R.H.; Bahn, S. Oligodendrocyte dysfunction in schizophrenia and bipolar disorder. Lancet 2003, 362, 798–805. [Google Scholar] [CrossRef]
- Herring, N.R.; Konradi, C. Myelin, copper, and the cuprizone model of schizophrenia. Front. Biosci. Sch. 2011, 3 S, 23–40. [Google Scholar]
- Cathomas, F.; Azzinnari, D.; Bergamini, G.; Sigrist, H.; Buerge, M.; Hoop, V.; Wicki, B.; Goetze, L.; Soares, S.; Kukelova, D.; et al. Oligodendrocyte gene expression is reduced by and influences effects of chronic social stress in mice. Genes, Brain Behav. 2019, 18, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Miyata, S.; Taniguchi, M.; Koyama, Y.; Shimizu, S.; Tanaka, T.; Yasuno, F.; Yamamoto, A.; Iida, H.; Kudo, T.; Katayama, T.; et al. Association between chronic stress-induced structural abnormalities in Ranvier nodes and reduced oligodendrocyte activity in major depression. Sci. Rep. 2016, 6, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Love, S. Demyelinating diseases. J. Clin. Pathol. 2006, 59, 1151–1159. [Google Scholar] [CrossRef] [PubMed]
- Saxena, A.; Bauer, J.; Scheikl, T.; Zappulla, J.; Audebert, M.; Desbois, S.; Waisman, A.; Lassmann, H.; Liblau, R.S.; Mars, L.T. Cutting Edge: Multiple Sclerosis-Like Lesions Induced by Effector CD8 T Cells Recognizing a Sequestered Antigen on Oligodendrocytes. J. Immunol. 2008, 181, 1617–1621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duncan, I.D.; Radcliff, A.B. Inherited and acquired disorders of myelin: The underlying myelin pathology. Exp. Neurol. 2016, 283, 452–475. [Google Scholar] [CrossRef] [PubMed]
- Hesp, Z.C.; Goldstein, E.A.; Miranda, C.J.; Kaspar, B.K.; McTigue, D.M.; Kaspar, B.K.; McTigue, D.M. Chronic Oligodendrogenesis and Remyelination after Spinal Cord Injury in Mice and Rats. J. Neurosci. 2015, 35, 1274–1290. [Google Scholar] [CrossRef] [PubMed]
- Levine, J.M.; Reynolds, R. Activation and Proliferation of Endogenous Oligodendrocyte Precursor Cells during Ethidium Bromide-Induced Demyelination. Exp. Neurol. 1999, 160, 333–347. [Google Scholar] [CrossRef] [PubMed]
- Fancy, S.P.J.; Zhao, C.; Franklin, R.J.M. Increased expression of Nkx2.2 and Olig2 identifies reactive oligodendrocyte progenitor cells responding to demyelination in the adult CNS. Mol. Cell. Neurosci. 2004, 27, 247–254. [Google Scholar] [CrossRef] [PubMed]
- Arai, K.; Lo, E.H. An oligovascular niche: Cerebral endothelial cells promote the survival and proliferation of oligodendrocyte precursor cells. J. Neurosci. 2009, 29, 4351–4355. [Google Scholar] [CrossRef] [PubMed]
- Hammond, T.R.; Gadea, A.; Dupree, J.; Kerninon, C.; Nait-Oumesmar, B.; Aguirre, A.; Gallo, V. Astrocyte-Derived Endothelin-1 Inhibits Remyelination through Notch Activation. Neuron 2014, 81, 588–602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casaccia-Bonnefil, P.; Tikoo, R.; Kiyokawa, H.; Friedrich, V.; Chao, M.V.; Koff, A. Oligodendrocyte precursor differentiation is perturbed in the absence of the cyclin-dependent kinase inhibitor p27Kip1. Genes Dev. 1997, 11, 2335–2346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magri, L.; Gacias, M.; Wu, M.; Swiss, V.A.; Janssen, W.G.; Casaccia, P. c-Myc-dependent transcriptional regulation of cell cycle and nucleosomal histones during oligodendrocyte differentiation. Neuroscience 2014, 276, 72–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magri, L.; Swiss, V.A.; Jablonska, B.; Lei, L.; Pedre, X.; Walsh, M.; Zhang, W.; Gallo, V.; Canoll, P.; Casaccia, P. E2F1 coregulates cell cycle genes and chromatin components during the transition of oligodendrocyte progenitors from proliferation to differentiation. J. Neurosci. 2014, 34, 1481–1493. [Google Scholar] [CrossRef] [PubMed]
- Habes, M.; Erus, G.; Toledo, J.B.; Zhang, T.; Bryan, N.; Launer, L.J.; Rosseel, Y.; Janowitz, D.; Doshi, J.; Van Der Auwera, S.; et al. White matter hyperintensities and imaging patterns of brain ageing in the general population. Brain 2016, 139, 1164–1179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruckh, J.M.; Zhao, J.W.; Shadrach, J.L.; Van Wijngaarden, P.; Rao, T.N.; Wagers, A.J.; Franklin, R.J.M. Rejuvenation of regeneration in the aging central nervous system. Cell Stem Cell 2012, 10, 96–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sim, F.J.; Zhao, C.; Penderis, J.; Franklin, R.J.M. The age-related decrease in CNS remyelination efficiency is attributable to an impairment of both oligodendrocyte progenitor recruitment and differentiation. J. Neurosci. 2002, 22, 2451–2459. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Hill, R.A.; Dietrich, D.; Komitova, M.; Suzuki, R.; Nishiyama, A. Age-dependent fate and lineage restriction of single NG2 cells. Development 2011, 138, 745–753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saher, G.; Brügger, B.; Lappe-Siefke, C.; Möbius, W.; Tozawa, R.I.; Wehr, M.C.; Wieland, F.; Ishibashi, S.; Nave, K.A. High cholesterol level is essential for myelin membrane growth. Nat. Neurosci. 2005, 8, 468–475. [Google Scholar] [CrossRef] [PubMed]
- Thelen, K.M.; Falkai, P.; Bayer, T.A.; Lütjohann, D. Cholesterol synthesis rate in human hippocampus declines with aging. Neurosci. Lett. 2006, 403, 15–19. [Google Scholar] [CrossRef] [PubMed]
- Thorburne, S.K.; Juurlink, B.H.J. Low Glutathione and High Iron Govern the Susceptibility of Oligodendroglial Precursors to Oxidative Stress. J. Neurochem. 1996, 67, 1014–1022. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.M.; Choi, J. Age-associated decline in γ-glutamylcysteine synthetase gene expression in rats. Free Radic. Biol. Med. 2000, 28, 566–574. [Google Scholar] [CrossRef]
- Back, S.A.; Luo, N.L.; Borenstein, N.S.; Levine, J.M.; Volpe, J.J.; Kinney, H.C. Late oligodendrocyte progenitors coincide with the developmental window of vulnerability for human perinatal white matter injury. J. Neurosci. 2001, 21, 1302–1312. [Google Scholar] [CrossRef] [PubMed]
- Shields, S.A.; Gilson, J.M.; Blakemore, W.F.; Franklin, R.J.M. Remyelination occurs as extensively but more slowly in old rats compared to young rats following gliotoxin-induced CNS demyelination. Glia 1999, 28, 77–83. [Google Scholar] [CrossRef]
- Duncan, I.D.; Brower, A.; Kondo, Y.; Curlee, J.F.; Schultz, R.D. Extensive remyelination of the CNS leads to functional recovery. Proc. Natl. Acad. Sci. USA 2009, 106, 6832–6836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gruchot, J.; Weyers, V.; Göttle, P.; Förster, M.; Hartung, H.-P.; Küry, P.; Kremer, D. The Molecular Basis for Remyelination Failure in Multiple Sclerosis. Cells 2019, 8, 825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeung, M.S.; Djelloul, M.; Steiner, E.; Bernard, S.; Salehpour, M.; Possnert, G.; Brundin, L.; Frisén, J. Oligodendrocyte generation dynamics in multiple sclerosis. Nature 2019, 24, 1837–1844. [Google Scholar] [CrossRef] [PubMed]
- Marques, S.; Zeisel, A.; Codeluppi, S.; van Bruggen, D.; Falcão, A.M.; Xiao, L.; Li, H.; Häring, M.; Hochgerner, H.; Romanov, R.A.; et al. Oligodendrocyte heterogeneity in the mouse juvenile and adult central nervous system. Science 2016, 352, 1326–1329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viganò, F.; Möbius, W.; Götz, M.; Dimou, L. Transplantation reveals regional differences in oligodendrocyte differentiation in the adult brain. Nat. Neurosci. 2013, 16, 1370–1372. [Google Scholar] [CrossRef] [PubMed]
- Crawford, A.H.; Tripathi, R.B.; Foerster, S.; McKenzie, I.; Kougioumtzidou, E.; Grist, M.; Richardson, W.D.; Franklin, R.J.M. Pre-existing mature oligodendrocytes do not contribute to remyelination following toxin-induced spinal cord demyelination. Am. J. Pathol. 2016, 186, 511–516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Targett, M.P.; Sussman, J.; Scolding, N.; O’Leary, M.T.; Compston, D.A.; Blakemore, W.F. Failure to achieve remyelination of demyelinated rat axons following transplantation of glial cells obtained from the adult human brain. Neuropathol. Appl. Neurobiol. 1996, 22, 199–206. [Google Scholar] [CrossRef] [PubMed]
- Duncan, I.D.; Radcliff, A.B.; Heidari, M.; Kidd, G.; August, B.K.; Wierenga, L.A. The adult oligodendrocyte can participate in remyelination. Proc. Natl. Acad. Sci. USA 2018, 115, E11807–E11816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soundarapandian, M.M.; Selvaraj, V.; Lo, U.G.; Golub, M.S.; Feldman, D.H.; Pleasure, D.E.; Deng, W. Zfp488 promotes oligodendrocyte differentiation of neural progenitor cells in adult mice after demyelination. Sci. Rep. 2011, 1, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duncan, G.J.; Plemel, J.R.; Assinck, P.; Manesh, S.B.; Muir, F.G.W.; Hirata, R.; Berson, M.; Liu, J.; Wegner, M.; Emery, B.; et al. Myelin regulatory factor drives remyelination in multiple sclerosis. Acta Neuropathol. 2017, 134, 403–422. [Google Scholar] [CrossRef] [PubMed]
- Dugas, J.C.; Ibrahim, A.; Barres, B.A. The T3-induced gene KLF9 regulates oligodendrocyte differentiation and myelin regeneration. Mol. Cell. Neurosci. 2012, 50, 45–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steelman, A.J.; Zhou, Y.; Koito, H.; Kim, S.J.; Payne, H.R.; Lu, Q.R.; Li, J. Activation of oligodendroglial Stat3 is required for efficient remyelination. Neurobiol. Dis. 2016, 91, 336–346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuhlmann, T.; Miron, V.; Cuo, Q.; Wegner, C.; Antel, J.; Bru« Ck, W.; Kuhlmann, T. Differentiation block of oligodendroglial progenitor cells as a cause for remyelination failure in chronic multiple sclerosis. Brain 2008, 131, 1749–1758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, A.; Tourtellotte, W.W.; Rudick, R.; Trapp, B.D. Premyelinating oligodendrocytes in chronic lesions of multiple sclerosis. N. Engl. J. Med. 2002, 346, 165–173. [Google Scholar] [CrossRef] [PubMed]
- Mason, J.L.; Toews, A.; Hostettler, J.D.; Morell, P.; Suzuki, K.; Goldman, J.E.; Matsushima, G.K. Oligodendrocytes and Progenitors Become Progressively Depleted within Chronically Demyelinated Lesions. Am. J. Pathol. 2004, 164, 1673–1682. [Google Scholar] [CrossRef] [Green Version]
- Boyd, A.; Zhang, H.; Williams, A. Insufficient OPC migration into demyelinated lesions is a cause of poor remyelination in MS and mouse models. Acta Neuropathol. 2013, 125, 841–859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bieber, A.J.; Kerr, S.; Rodriguez, M. Efficient central nervous system remyelination requires T cells. Ann. Neurol. 2003, 53, 680–684. [Google Scholar] [CrossRef] [PubMed]
- Miron, V.E.; Boyd, A.; Zhao, J.-W.; Yuen, T.J.; Ruckh, J.M.; Shadrach, J.L.; van Wijngaarden, P.; Wagers, A.J.; Williams, A.; Franklin, R.J.M.; et al. M2 microglia and macrophages drive oligodendrocyte differentiation during CNS remyelination. Nat. Neurosci. 2013, 16, 1211–1218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dombrowski, Y.; O’Hagan, T.; Dittmer, M.; Penalva, R.; Mayoral, S.R.; Bankhead, P.; Fleville, S.; Eleftheriadis, G.; Zhao, C.; Naughton, M.; et al. Regulatory T cells promote myelin regeneration in the central nervous system. Nat. Neurosci. 2017, 20, 674–680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Schematic depiction of oligodendroglial lineage markers specific for different developmental stages from neuronal progenitor cells (NPC) to myelinating oligodendrocyte (OL). A2B5 recognises progenitor cells, NPC and oligodendrocyte progenitor cells (OPC), while oligodendroglial cell lineage markers Olig1 and 2 as well as Sox10 and Nkx2.2 are expressed in all cells of the lineage, OPC and pre-oligodendrocytes (pre-OL) are characterised by PDGFR-α and NG2 expression. PLP, O4, O1 and CNPase are expressed during transition from progenitor to differentiated oligodendrocytes, while differentiated, axon-myelinating oligodendrocytes are characterised by myelin protein expression (MBP, MAG, MOG, GalC). NPC: neuronal progenitor cell; OPC: oligodendrocyte progenitor cell; OL: oligodendrocyte; PDGFR-α: platelet-derived growth factor receptor A; NG2: neuron-glial antigen 2; PLP: proteolipid protein; CNPase: 2’,3’-Cyclic-nucleotide 3’-phosphodiesterase; MBP: myelin basic protein; MAG: myelin associated glycoprotein; MOG: myelin-oligodendrocyte glycoprotein; GalC: galactocerebroside.
Figure 1.
Schematic depiction of oligodendroglial lineage markers specific for different developmental stages from neuronal progenitor cells (NPC) to myelinating oligodendrocyte (OL). A2B5 recognises progenitor cells, NPC and oligodendrocyte progenitor cells (OPC), while oligodendroglial cell lineage markers Olig1 and 2 as well as Sox10 and Nkx2.2 are expressed in all cells of the lineage, OPC and pre-oligodendrocytes (pre-OL) are characterised by PDGFR-α and NG2 expression. PLP, O4, O1 and CNPase are expressed during transition from progenitor to differentiated oligodendrocytes, while differentiated, axon-myelinating oligodendrocytes are characterised by myelin protein expression (MBP, MAG, MOG, GalC). NPC: neuronal progenitor cell; OPC: oligodendrocyte progenitor cell; OL: oligodendrocyte; PDGFR-α: platelet-derived growth factor receptor A; NG2: neuron-glial antigen 2; PLP: proteolipid protein; CNPase: 2’,3’-Cyclic-nucleotide 3’-phosphodiesterase; MBP: myelin basic protein; MAG: myelin associated glycoprotein; MOG: myelin-oligodendrocyte glycoprotein; GalC: galactocerebroside.

Figure 2.
Oligodendrocytes in myelination, demyelination and remyelination. Oligodendrocytes myelinate large diameter axons in the CNS and provide trophic support for the underlying axon. Oligodendrocytes are highly vulnerable and insults such as trauma, immune-mediated attacks or ischaemia can lead to oligodendrocyte death and demyelination. Newly differentiated oligodendrocytes derived from an adult OPC pool can replace deceased oligodendrocytes, which can reinstate the myelin sheath around demyelinated axons (remyelination). Regenerated myelin is thinner than the original myelin sheath.
Figure 2.
Oligodendrocytes in myelination, demyelination and remyelination. Oligodendrocytes myelinate large diameter axons in the CNS and provide trophic support for the underlying axon. Oligodendrocytes are highly vulnerable and insults such as trauma, immune-mediated attacks or ischaemia can lead to oligodendrocyte death and demyelination. Newly differentiated oligodendrocytes derived from an adult OPC pool can replace deceased oligodendrocytes, which can reinstate the myelin sheath around demyelinated axons (remyelination). Regenerated myelin is thinner than the original myelin sheath.


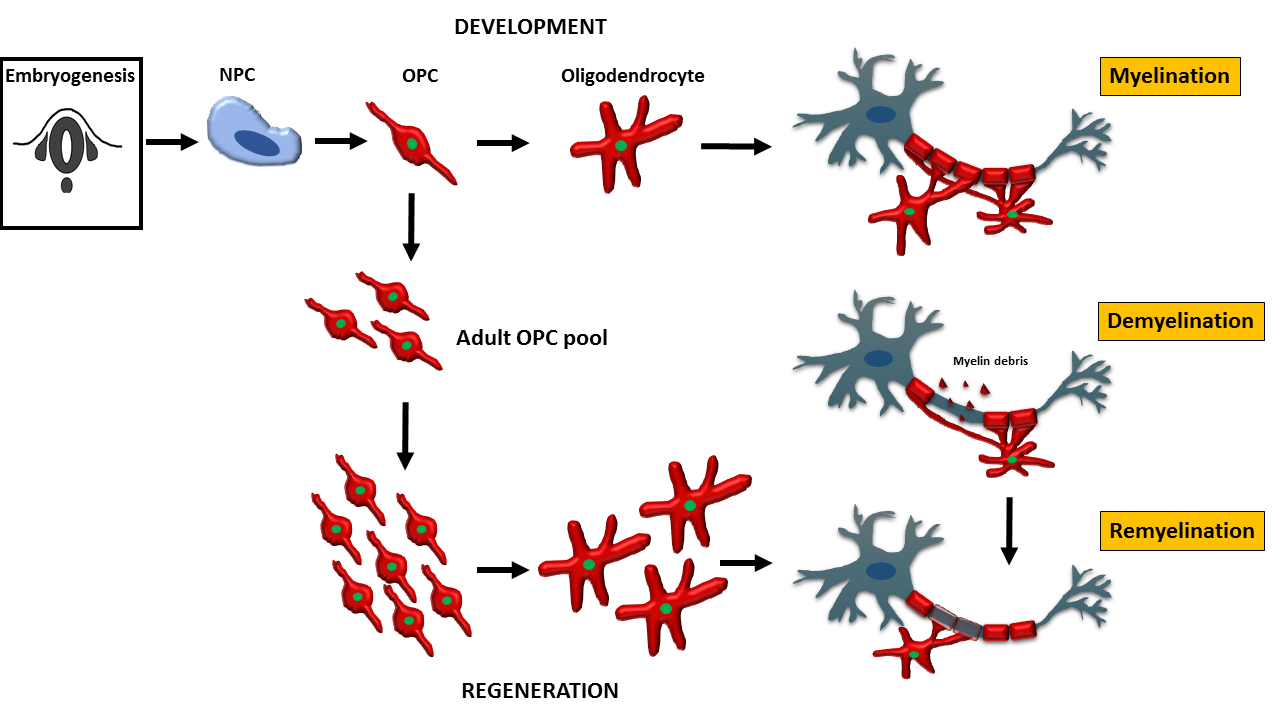{kind=link}
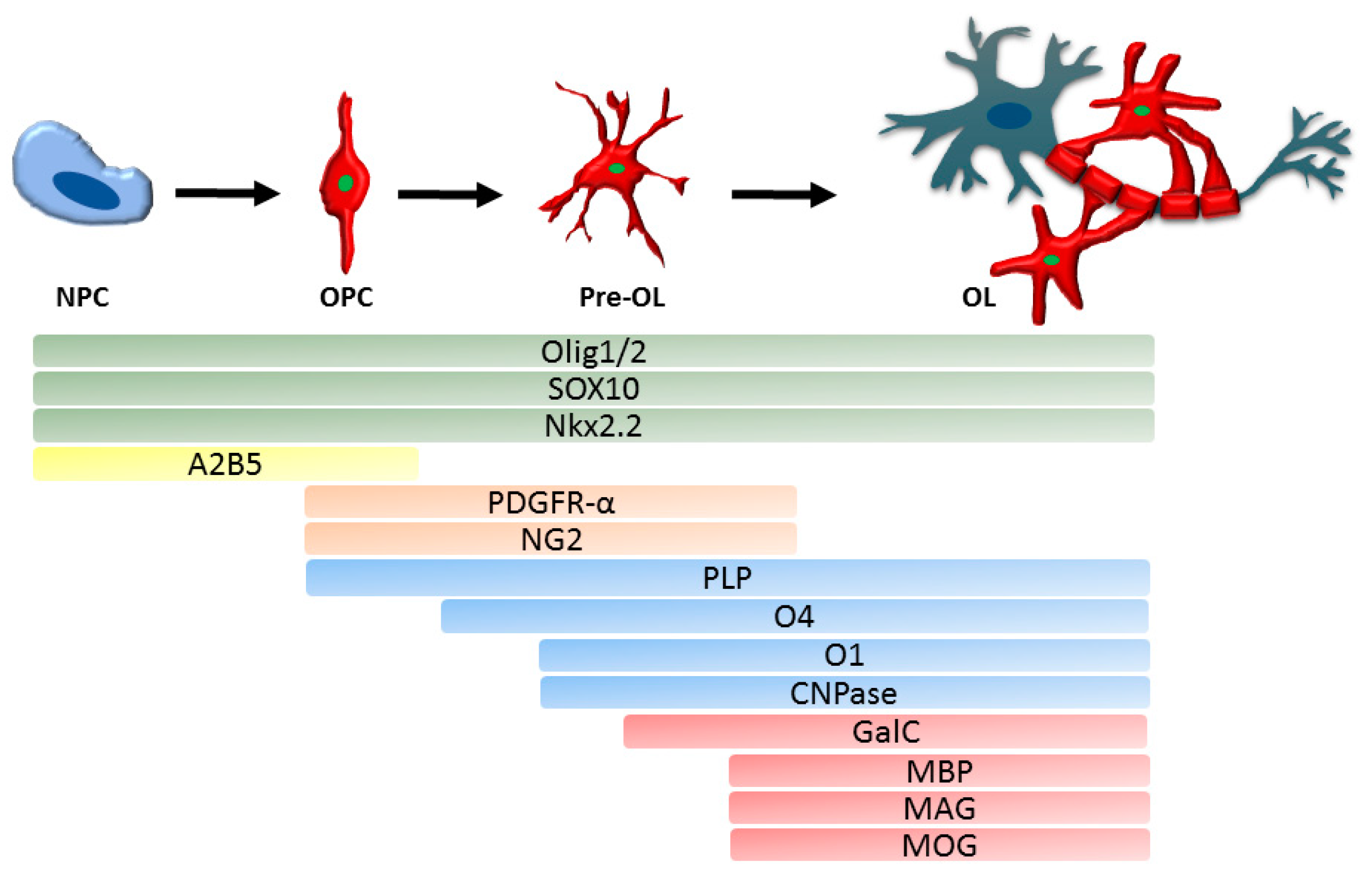{kind=link}
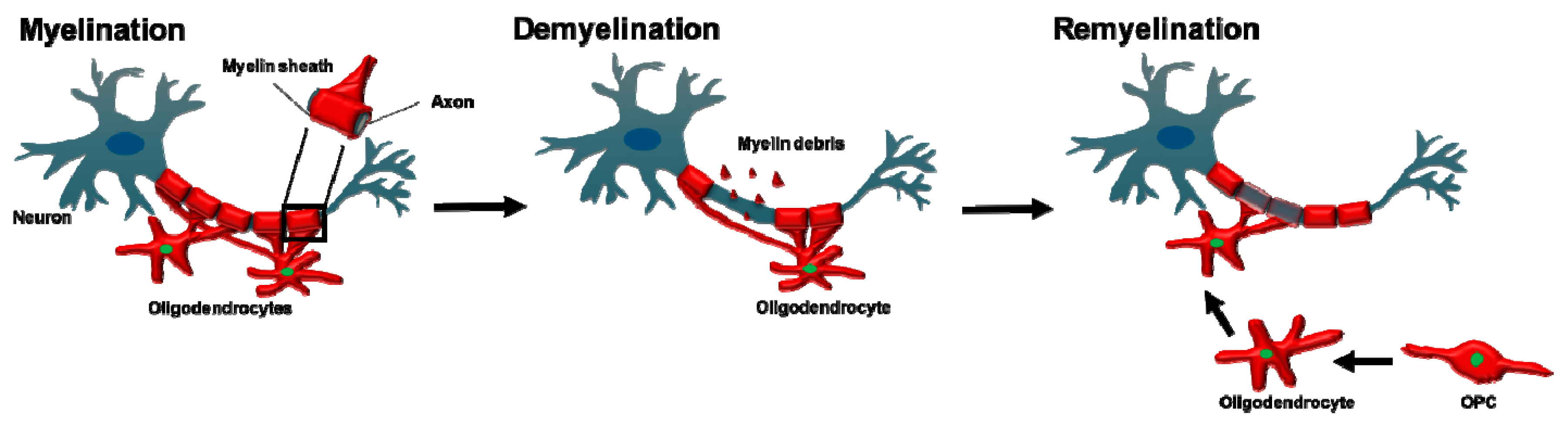{kind=link}
Table 1.
Summary of animal and human models listed in Section 4.1 and Section 4.2. This list provides examples of a variety of models available to study OPC and oligodendrocyte (OL) biology.
Table 1.
Summary of animal and human models listed in Section 4.1 and Section 4.2. This list provides examples of a variety of models available to study OPC and oligodendrocyte (OL) biology.
| In vitro Animal Models | In vitro Human Models | In Vivo Animal Models |
|---|---|---|
| OPC/OL cultures [128,129,130] | OPC derived from biopsies [138] or post mortem CNS [139] | Reporter mice (e.g., Sox10-Venus mice, CNP-EGFP mice, PLP-EGFP mice) [147,148,149] |
| Immortalized oligodendroglial cells lines (e.g., CG-4, OLN-93, Oli-neu) [133,134,135] | iOL derived from human iPSC [137,141,142,143] | Reporter zebrafish (e.g., Tg(sox10:mRFP), Tg(olig2:EGFP), Tg(mbp:EGFP)) [103,150,151] |
OPC: Oligodendrocyte Progenitor Cell, OL: oligodendrocyte, CNS: Central Nervous System, iOL: induced Oligodendrocyte, iPSC: induced Pluripotent Stem Cell.
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kuhn, S.; Gritti, L.; Crooks, D.; Dombrowski, Y. Oligodendrocytes in Development, Myelin Generation and Beyond. Cells 2019, 8, 1424. https://doi.org/10.3390/cells8111424
AMA Style
Kuhn S, Gritti L, Crooks D, Dombrowski Y. Oligodendrocytes in Development, Myelin Generation and Beyond. Cells. 2019; 8(11):1424. https://doi.org/10.3390/cells8111424
Chicago/Turabian StyleKuhn, Sarah, Laura Gritti, Daniel Crooks, and Yvonne Dombrowski. 2019. "Oligodendrocytes in Development, Myelin Generation and Beyond" Cells 8, no. 11: 1424. https://doi.org/10.3390/cells8111424
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.