A Disintegrin and Metalloprotease 15 is Expressed on Rheumatoid Arthritis Synovial Tissue Endothelial Cells and may Mediate Angiogenesis


Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients
2.2. Cell Culture
2.3. Enzyme-Linked Immunosorbent Assay (ELISA) of ADAM-15, CXCL16, Fractalkine/CX3CL1, ENA78/CXCL5, Intercellular Adhesion Molecule (ICAM)-1, Vascular Cell Adhesion Molecule (VCAM)-1, MCP-1/CCL2, IL-8, and TGF-β
2.4. RNA Extraction and Quantitative Polymerase Chain Reaction (qPCR) of HUVECs
2.5. Immunohistochemistry
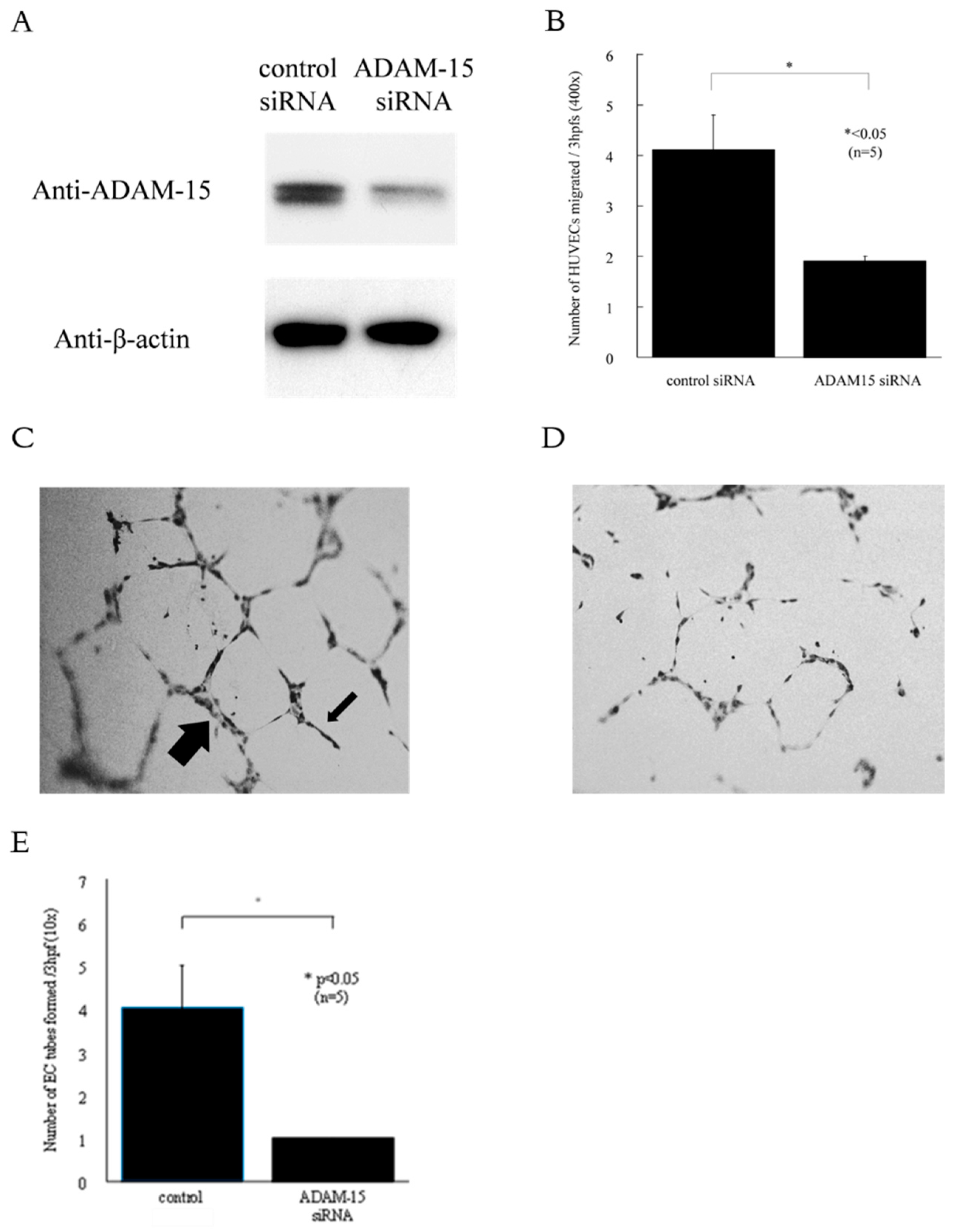
2.6. RNA Silencing
2.7. In Vitro HUVEC Chemotaxis Assay
2.8. In vitro Matrigel Tube Formation Assay
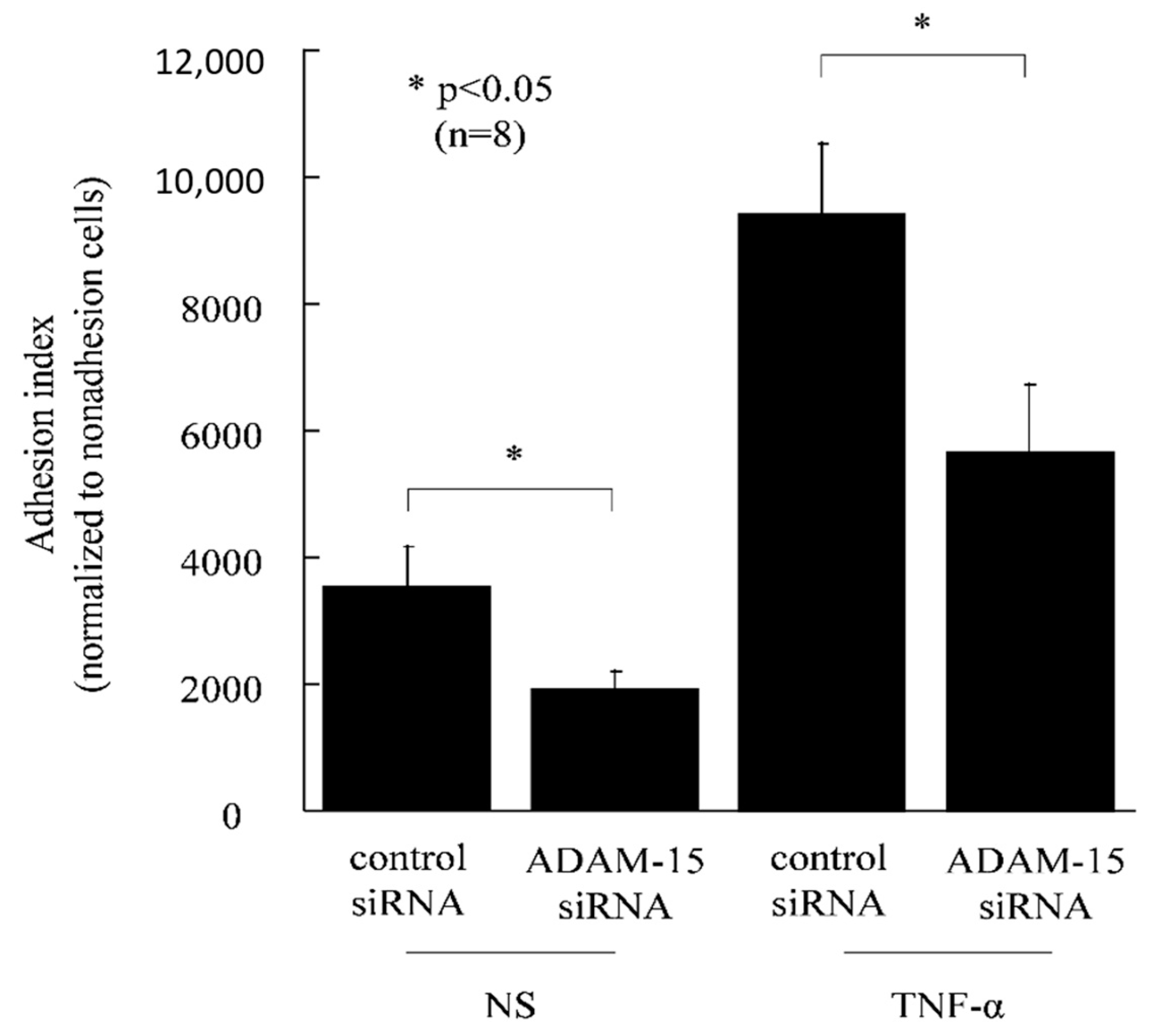
2.9. Monocyte Adhesion Assay
2.10. Statistical Analysis
3. Results
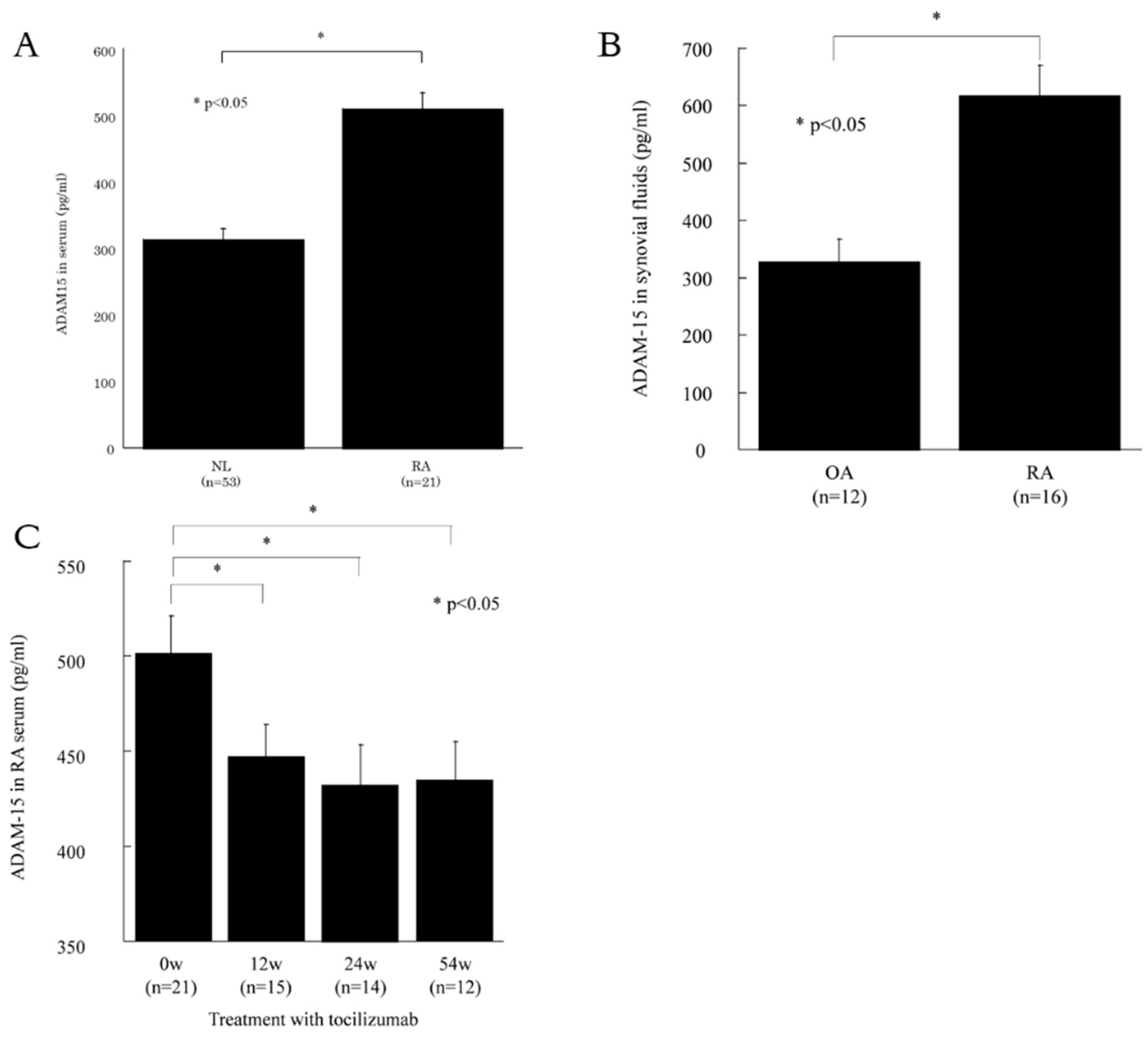
3.1. ADAM15 is Expressed in RA Sera and Synovial Fluids
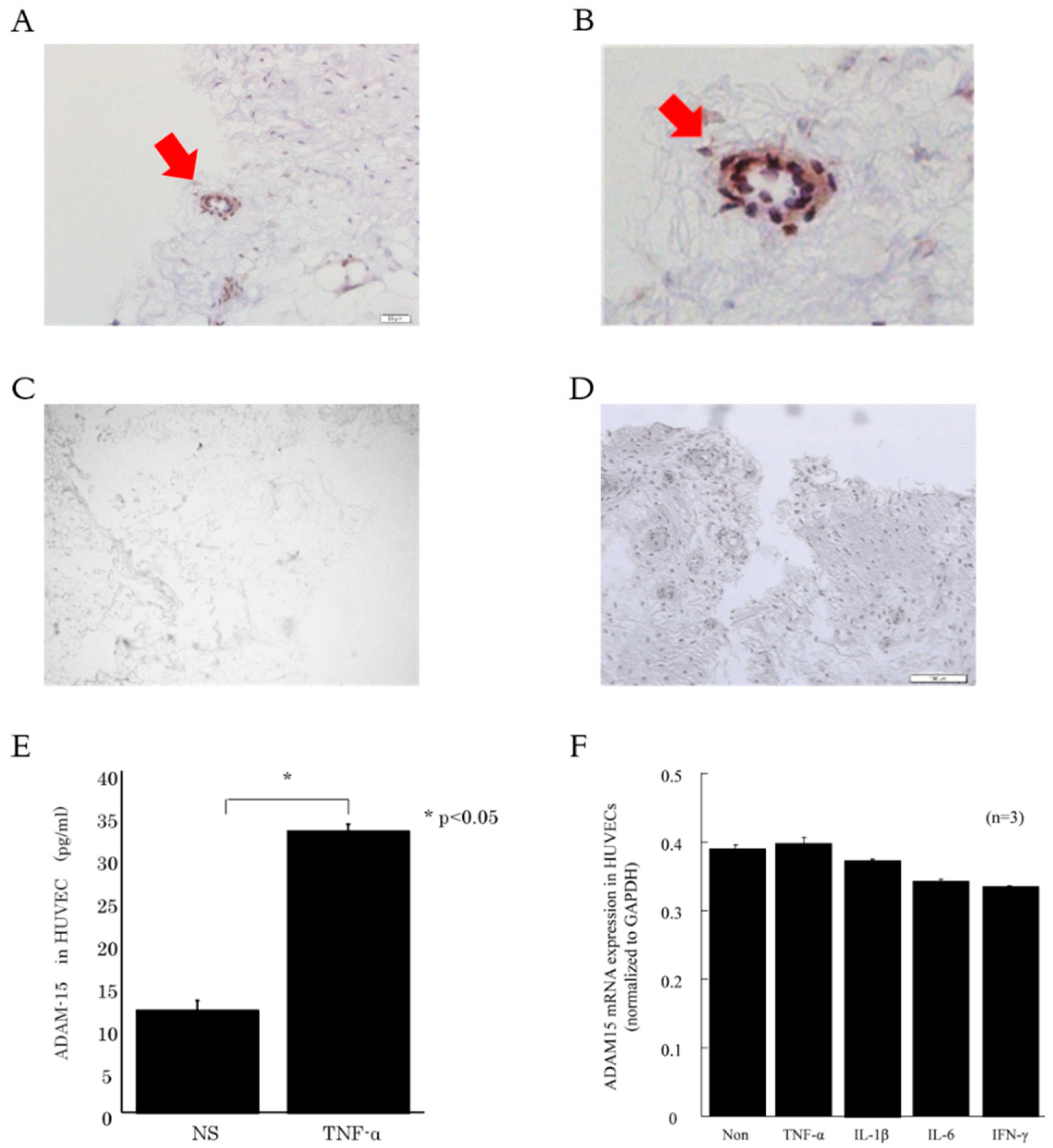
3.2. ADAM15 is Expressed in RA ST Endothelial Cells (ECs)
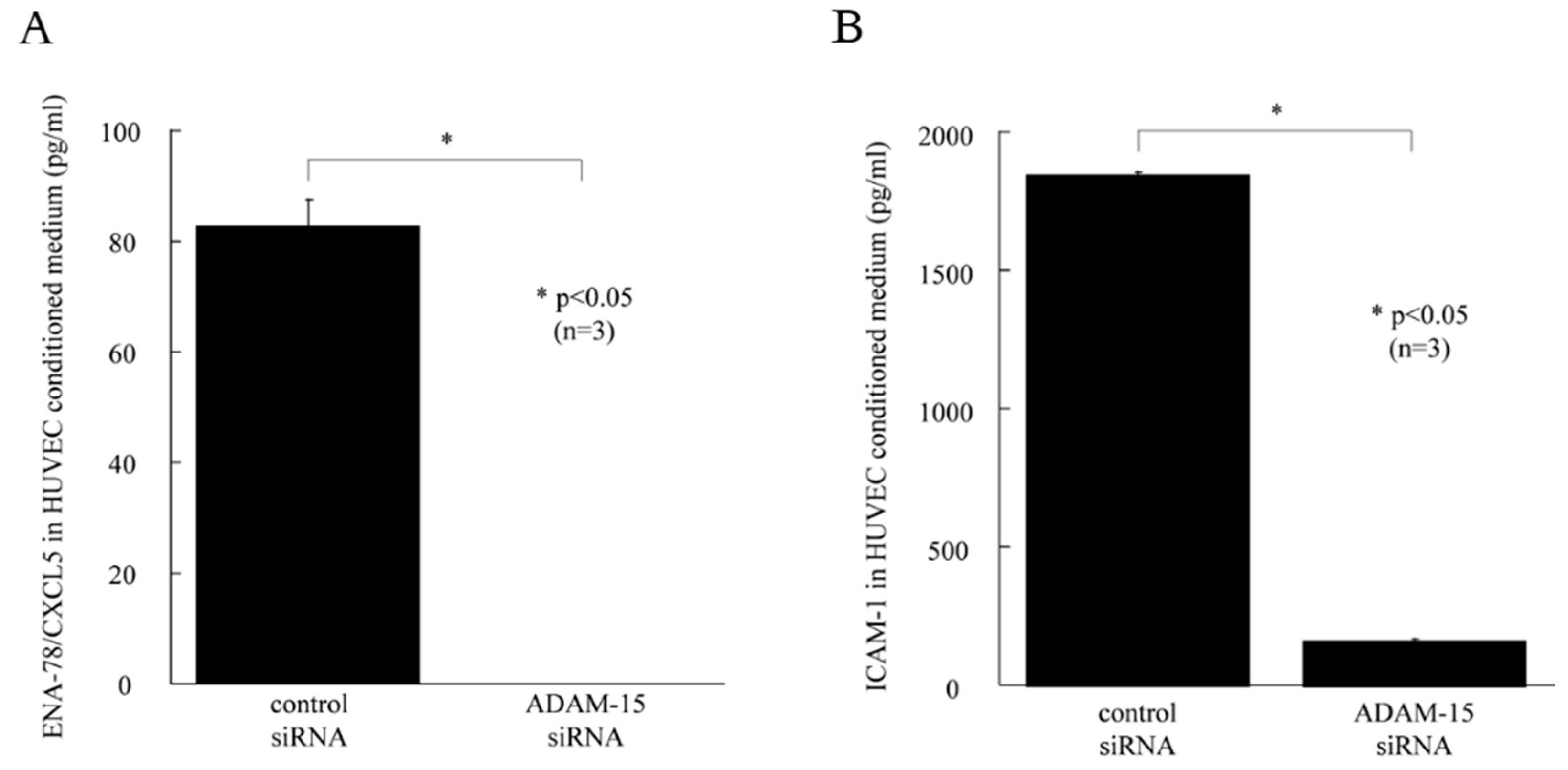
3.3. ADAM15 Mediates the Release of Potent Angiogenic Factors
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Feldmann, M.; Brennan, F.M.; Maini, R.N. Rheumatoid arthritis. Cell 1996, 85, 307–310. [Google Scholar] [CrossRef]
- Schaible, H.G. Nociceptive neurons detect cytokines in arthritis. Arthritis Res. Ther. 2014, 16, 470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cutolo, M.; Sulli, A.; Barone, A.; Seriolo, B.; Accardo, S. Macrophages, synovial tissue and rheumatoid arthritis. Clin. Exp. Rheumatol. 1993, 11, 331–339. [Google Scholar] [PubMed]
- Schett, G.; Gravallese, E. Bone erosion in rheumatoid arthritis: Mechanisms, diagnosis and treatment. Nat. Rev. Rheumatol. 2012, 8, 656–664. [Google Scholar]
- Kasama, T.; Isozaki, T.; Takahashi, R.; Miwa, Y. Clinical effects of tocilizumab on cytokines and immunological factors in patients with rheumatoid arthritis. Int. Immunopharmacol. 2016, 35, 301–306. [Google Scholar] [CrossRef] [PubMed]
- Neovius, M.; Arkema, E.V.; Olsson, H. Drug survival on TNF inhibitors in patients with rheumatoid arthritis comparison of adalimumab, etanercept and infliximab. Ann. Rheum. Dis. 2015, 74, 354–360. [Google Scholar] [CrossRef] [PubMed]
- Bohm, B.B.; Aigner, T.; Gehrsitz, A.; Blobel, C.P.; Kalden, J.R.; Burkhardt, H. Up-regulation of MDC15 (metargidin) messenger RNA in human osteoarthritic cartilage. Arthritis Rheum. 1999, 42, 1946–1950. [Google Scholar] [CrossRef] [Green Version]
- Scheller, J.; Chalaris, A.; Garbers, C.; Rose-John, S. ADAM17: A molecular switch to control inflammation and tissue regeneration. Trends immunol. 2011, 32, 380–387. [Google Scholar] [CrossRef] [PubMed]
- Wolf, C.; Qian, Y.; Brooke, M.A.; Kelsell, D.P.; Franzke, C.W. ADAM17/EGFR axis promotes transglutaminase-dependent skin barrier formation through phosholipase C gamma1 and protein kinase C pathways. Sci. Rep. 2016, 6, 39780. [Google Scholar] [CrossRef]
- Kuefer, R.; Day, K.C.; Kleer, C.G. ADAM15 disintegrin is associated with aggressive prostate and breast cancer disease. Neoplasia 2006, 8, 319–329. [Google Scholar] [CrossRef]
- Komiya, K.; Enomoto, H.; Inoki, I. Expression of ADAM15 in rheumatoid synovium: Up-regulation by vascular endothelial growth factor and possible implications for angiogenesis. Arthritis Res. Ther. 2005, 7, R1158–R1173. [Google Scholar] [CrossRef] [PubMed]
- Matsunawa, M.; Isozaki, T.; Odai, T. Increased serum levels of soluble fractalkine (CX3CL1) correlate with disease activity in rheumatoid vasculitis. Arthritis Rheum. 2006, 54, 3408–3416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isozaki, T.; Rabquer, B.J.; Ruth, J.H.; Haines, G.K.; Koch, A.E. ADAM-10 is overexpressed in rheumatoid arthritis synovial tissue and mediates angiogenesis. Arthritis Rheum. 2013, 65, 98–108. [Google Scholar] [CrossRef] [PubMed]
- Rose-John, S. ADAM17, shedding, TACE as therapeutic targets. Pharmacol. Res. 2013, 71, 19–22. [Google Scholar] [CrossRef] [PubMed]
- Ortiz, R.M.; Karkkainen, I.; Huovila, A.P. Aberrant alternative exon use and increased copy number of human metalloprotease-disintegrin ADAM15 gene in breast cancer cells. Genes Chromosom. Cancer 2004, 41, 366–378. [Google Scholar]
- Lucas, N.; Day, M.L. The role of the disintegrin metalloproteinase ADAM15 in prostate cancer progression. J. Cell. Biochem. 2009, 106, 967–974. [Google Scholar] [CrossRef] [PubMed]
- Mosnier, J.F.; Jarry, A.; Bou-Hanna, C.; Denis, M.G.; Merlin, D.; Laboisse, C.L. ADAM15 upregulation and interaction with multiple binding partners in inflammatory bowel disease. Lab Investig. 2006, 86, 1064–1073. [Google Scholar] [CrossRef]
- Gao, J.; Zheng, W.; Wang, L.; Song, B. A disintegrin and metallproteinase 15 knockout decreases migration of fibroblast-like synoviocytes and inflammation in rheumatoid arthritis. Mol. Med. Rep. 2015, 11, 4389–4396. [Google Scholar] [CrossRef]
- Bohm, B.B.; Schirner, A.; Burkhardt, H. ADAM15 modulates outside-in signalling in chondrocyte-matrix interactions. J. Cell. Mol. Med. 2009, 13, 2634–2644. [Google Scholar] [CrossRef]
- Sun, C.; Wu, M.H.; Guo, M.; Day, M.L.; Lee, E.S.; Yuan, S.Y. ADAM15 regulates endothelial permeability and neutrophil migration via Src/ERK1/2 signalling. Cardiovasc. Res. 2010, 87, 348–355. [Google Scholar] [CrossRef] [Green Version]
- Elshabrawy, H.A.; Chen, Z.; Volin, M.V.; Ravella, S.; Virupannavar, S.; Shahrara, S. The pathogenic role of angiogenesis in rheumatoid arthritis. Angiogenesis 2015, 18, 433–448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aporntewan, C.; Ballard, D.H.; Lee, J.Y.; Lee, J.S.; Wu, Z.; Zhao, H. Gene hunting of the Genetic Analysis Workshop 16 rheumatoid arthritis data using rough set theory. BMC Proc. 2009, 3, S126. [Google Scholar] [CrossRef] [Green Version]
- Koch, A.E.; Kunkel, S.L.; Harlow, L.A. Epithelial neutrophil activating peptide-78: A novel chemotactic cytokine for neutrophils in arthritis. J. Clin. Investig. 1994, 94, 1012–1018. [Google Scholar] [CrossRef] [PubMed]
- Amin, M.A.; Campbell, P.L.; Ruth, J.H. A key role for Fut1-regulated angiogenesis and ICAM-1 expression in K/BxN arthritis. Ann. Rheum. Dis. 2015, 74, 1459–1466. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Total (n) | 21 |
|---|---|
| Gender (female:male) | 20:1 |
| Mean age (years) | 51.7 ± 3.43 |
| Duration of RA (years) | 7.3 ± 1.9 |
| Baseline DAS28 (ESR) | 4.83 ± 0.29 |
| Dosage of MTX (mg) | 6.48 ± 1.16 |
| Dosage of prednisolone (mg) | 3.64 ± 0.56 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nishimi, S.; Isozaki, T.; Wakabayashi, K.; Takeuchi, H.; Kasama, T. A Disintegrin and Metalloprotease 15 is Expressed on Rheumatoid Arthritis Synovial Tissue Endothelial Cells and may Mediate Angiogenesis. Cells 2019, 8, 32. https://doi.org/10.3390/cells8010032
Nishimi S, Isozaki T, Wakabayashi K, Takeuchi H, Kasama T. A Disintegrin and Metalloprotease 15 is Expressed on Rheumatoid Arthritis Synovial Tissue Endothelial Cells and may Mediate Angiogenesis. Cells. 2019; 8(1):32. https://doi.org/10.3390/cells8010032
Chicago/Turabian StyleNishimi, Shinichiro, Takeo Isozaki, Kuninobu Wakabayashi, Hiroko Takeuchi, and Tsuyoshi Kasama. 2019. "A Disintegrin and Metalloprotease 15 is Expressed on Rheumatoid Arthritis Synovial Tissue Endothelial Cells and may Mediate Angiogenesis" Cells 8, no. 1: 32. https://doi.org/10.3390/cells8010032