
Transgene-Free Cynomolgus Monkey iPSCs Generated under Chemically Defined Conditions
by
 and
and
Yuliia Tereshchenko
1,2,†, 
Nesil Esiyok
1,†,
Enrique Garea-Rodríguez
3,
Daniele Repetto
3,
Rüdiger Behr
1,2,* and
Ignacio Rodríguez-Polo
1,2,4,*,†,‡ 1
Research Platform Degenerative Diseases, German Primate Center-Leibniz Institute for Primate Research, Kellnerweg 4, 37077 Göttingen, Germany
2
German Center for Cardiovascular Research (DZHK), Partner Site Göttingen, 37075 Göttingen, Germany
3
Charles River Laboratories Germany GmbH, Hans-Adolf-Krebs-Weg 9, 37077 Göttingen, Germany
4
Department of Developmental Biology, Göttingen Center for Molecular Biosciences, University of Göttingen, Justus-von-Liebig Weg 11, 37077 Göttingen, Germany
*
Authors to whom correspondence should be addressed.
†
These authors contributed equally to this work.
‡
Current address: Stem Cell & Human Development Laboratory, The Francis Crick Institute, 1 Midland Road, London NW1 1AT, UK.
Cells 2024, 13(6), 558; https://doi.org/10.3390/cells13060558
Submission received: 20 December 2023
/
Revised: 11 March 2024
/
Accepted: 15 March 2024
/
Published: 21 March 2024
(This article belongs to the Collection Stem Cells in Tissue Engineering and Regeneration)
Abstract
:Non-human primates (NHPs) are pivotal animal models for translating novel cell replacement therapies into clinical applications, including validating the safety and efficacy of induced pluripotent stem cell (iPSC)-derived products. Preclinical development and the testing of cell-based therapies ideally comprise xenogeneic (human stem cells into NHPs) and allogenic (NHP stem cells into NHPs) transplantation studies. For the allogeneic approach, it is necessary to generate NHP-iPSCs with generally equivalent quality to the human counterparts that will be used later on in patients. Here, we report the generation and characterization of transgene- and feeder-free cynomolgus monkey (Macaca fascicularis) iPSCs (Cyno-iPSCs). These novel cell lines have been generated according to a previously developed protocol for the generation of rhesus macaque, baboon, and human iPSC lines. Beyond their generation, we demonstrate the potential of the novel Cyno-iPSCs to differentiate into two clinically relevant cell types, i.e., cardiomyocytes and neurons. Overall, we provide a resource of novel iPSCs from the most frequently used NHP species in the regulatory testing of biologics and classical pharmaceutics to expand our panel of iPSC lines from NHP species with high relevance in preclinical testing and translational research.
1. Introduction
Due to their origin, in vitro expansion potential, and capacity to differentiate into any cell of the adult body, induced pluripotent stem cells (iPSCs) hold great potential for the treatment of degenerative diseases. The generation of advanced therapy medicinal products (ATMPs) derived from iPSCs has been the focus of study of many research groups since the development of iPSC technologies in 2006 [1,2]. Nowadays, using iPSCs, it is possible to generate a large number of different functional cell types in vitro, e.g., neurons and cardiomyocytes, which can be transplanted into patients. Many reports have been published showing promising results regarding the treatment of diseases like myocardial infarction, Parkinson’s disease, or macular degeneration [3]. Over the last few years, the field has broadly diversified, and, considering the diverse upcoming novel therapeutic strategies for the treatment of diverse diseases, it is necessary to adapt and optimize the panel of translational animal models that allow the efficient translation of these novel therapeutic approaches to the clinics.
For the preclinical testing of regenerative therapies, research mainly relies on large animal models such as pigs and non-human primates (NHPs), depending on the specific questions each study aims to address [4,5,6,7]. In particular, NHPs emerged as invaluable models to study the potential of stem cell-based regenerative therapies, due to their similarities in physiology, immunology, and genetics to humans [6,7].
To generate robust preclinical safety and efficacy data for ATMPs, ideally, both xenogenic and autologous/allogenic transplantation strategies of ATMPs should be tested. In xenogenic transplantation studies in NHPs, human iPSC-derived tissue-specific cells are usually transplanted into the NHP. These studies mainly address the characteristics and the safety (tumorigenicity) of the actual ATMPs that will be used later in patients. In autologous/allogenic transplantation studies, species-specific iPSC-derived ATMPs are transplanted into the NHP. This approach addresses survival, immunological aspects, safety, and functionality [7].
To perform translational allogenic studies, it is expedient to generate NHP-iPSCs of quality and developmental potential that are homologous to their human counterparts. However, NHP-PSC technologies have lagged behind human or mouse stem cell technologies [7]. Nevertheless, several reports over the last few years have expanded the panel of NHP-iPSCs generated under feeder-free and transgene-free conditions suitable for preclinical testing [8,9,10]. This expanding panel of species includes the baboon, rhesus macaque, cynomolgus macaque, and marmoset [7,11].
In a previous study [9], we reported a robust protocol to generate transgene- and feeder-free iPSCs from human, rhesus macaque, and baboon cells using episomes and chemically defined cell culture conditions. These cells have demonstrated their usefulness not only for the validation of cell replacement therapies but are also currently used in several projects addressing primate developmental and evolutionary questions. Therefore, in the current work, we have successfully applied our previously published universal primate reprogramming and maintenance conditions to cynomolgus macaque cells (Macaca fascicularis). This is of major relevance as this NHP species is broadly and predominantly used in pharmaceutical research, development, and toxicity testing [12,13,14,15]. We have generated and characterized four novel cynomolgus monkey iPSC lines, the differentiation capacities of which were exemplarily tested by directed differentiation into two clinically relevant cell types, i.e., neurons and cardiomyocytes. The cell lines generated from the best-established NHP species in regulatory safety testing represent a significant expansion of the panel of NHP-iPSC lines available for the translation of preclinical findings to the clinics.
2. Materials and Methods
2.1. General Statement Addressing the Methods Used in This Study
This communication complements our previous study [9], expanding the versatility of the protocol described previously regarding an additional NHP species with major relevance in regulatory testing. Therefore, the methods for the generation, keeping, and differentiation of the Cyno-iPSCs are the same as those previously published. Detailed descriptions of the methods have been published recently [9,16].
2.2. Animals
Samples from two adult healthy and untreated male cynomolgus macaques (Macaca fascicularis) were used for fibroblast isolation. The tissue samples were collected in accordance with §4 (3) of the German Animal Welfare Act (killing for scientific purposes) at Charles River Laboratories during necropsy. The primary purpose was the removal of monkey hearts, lungs, and testes, which was approved by the responsible animal welfare officer. The treatment of the animals and their euthanasia was carried out in accordance with all legal guidelines and requirements.
2.3. Isolation and Cultivation of Macaque Primary Fibroblasts
Biopsies from the skin, gingiva, and foreskin were collected in ice-cold PBS (phosphate-buffered saline) with antibiotics (1%, v/v, penicillin/streptomycin, Gibco, Grand Island, NY, USA). Fibroblasts were extracted from tissue according to the method used in [17]. In brief, the tissue was minced using scissors and scalpels and digested in Dulbecco’s modified Eagle’s medium (DMEM) (Gibco), supplemented with 10 mg mL−1 collagenase type IV (Gibco), for 1–2 h at 37 °C in a Falcon tube rotator at 40 rpm. After digestion, the cell suspension was centrifuged (300× g; 5 min; room temperature). After centrifugation, the supernatant was removed and the cell pellet resuspended in Rh15 medium (DMEM, Gibco, 15%, v/v; fetal bovine serum, Gibco, 1%, v/v; penicillin/streptomycin, Gibco, 0.25 µg mL−1; amphotericin B, Sigma-Aldrich (Taufkirchen, Germany), 1%, v/v; minimum essential medium (MEM) non-essential amino acids solution, Gibco; 2 mM GlutaMAX, Gibco). Gingiva-derived fibroblasts were kept with a double amount of antibiotics (2%, v/v; penicillin/streptomycin, Gibco, 0.25 µg mL−1; amphotericin B, Sigma-Aldrich, 2%, v/v) for 4 days. Fibroblasts were expanded in Rh15 medium, using StemPro Accutase for dissociation (Thermo Fisher, Waltham, MA, USA).
2.4. Reprogramming NHP and Human Skin Fibroblasts and iPSC Maintenance
Reprogramming of the fibroblasts was performed by the nucleofection of primary fibroblast (6 × 105 cells per transfection) with episomal vectors pCXLE-hOCT3/4-shp53-F (Addgene #27077), pCXLE-hSK (#27078) and pCXLE-hUL (#27080) (2 µg of each plasmid per transfection, 6 µg total plasmid DNA) [18] using the 4D-Nucleofector (P2 Primary Cell Solution) (Lonza, Basel, Switzerland). The transfected fibroblasts were cultured for 1 day in Rh15 medium supplemented with 10 ng/mL bFGF, 5 µM pro-survival compound (ROCKi; Merck, Darmstadt, Germany), and penicillin-streptomycin. On day 1, after transfection, the medium was supplemented with 10 ng/mL bFGF and 0.5 mM sodium butyrate (Sigma-Aldrich). After 7 days, the cells were transferred to Geltrex™ (0.16 mg/mL; Thermo Fisher)-coated tissue culture plates. On day 8, the Rh15 medium was substituted by Essential 8 (E8) medium (Thermo Fisher) supplemented with 0.5 mM sodium butyrate until day 11. From day 12, the cells were cultured in E8 medium only. After 30 to 40 days, the iPSC colonies were picked manually and transferred to freshly coated plates.
Putative iPSC lines were cultured in a universal primate pluripotent stem cell (UPPS) medium [9], StemMACS iPS Brew XF (Miltenyi Biotec, Bergisch Gladbach, Germany), with 1 µM inhibitor of Wnt response-1 (IWR-1, Sigma-Aldrich) and 0.5 µM of the GSK-3α/β inhibitor Chir99021 (Merck) and were then passaged in clumps using Versene solution (Thermo Fisher).
2.5. In Vitro Differentiation
Cyno-iPSCs were differentiated using the embryoid body (EB) formation assay [9,16]. In brief, cells were dissociated using collagenase type IV into cell clumps that were cultured in suspension in UPPS medium for 24 h. One day later, the medium was changed for differentiation medium (79 mL IMDM (Thermo Fisher), 1 mL 100× NEAA, 20 mL FBS, 450 µM 1-thioglycerol (Sigma-Aldrich)). On day 8, the EBs in suspension were plated onto plates containing gelatin-coated coverslips (0.1% gelatin) (Fisher Scientific, Schwerte, Germany) to allow them to attach. The attached EBs were maintained in culture until day 25, when they were fixed and analyzed by immunocytochemistry.
2.6. Cardiac Differentiation
The cardiac differentiation protocol has been adapted with minor modifications adopted from [19]. Cyno-iPSCs (iPSC#1.1, 1.2, and 2.1) were seeded in 12-well plates, with 90,000 cells/well, and cultured for two days. Then, the cells were exposed to mesodermal induction medium (RPMI 1640, B27 supplement without insulin (ThermoFisher), 200 µM L-ascorbic acid 2-phosphate (Sigma-Aldrich), 1 mM sodium pyruvate (ThermoFisher), 1 µM Chir99021 (Tocris, Bristol, UK), 9 ng/mL activin A (Miltenyi Biotec), and 5 ng/mL BMP4 (Miltenyi Biotec). The medium was changed after 24 h. On day 3, the medium was replaced by cardiac induction medium (RPMI 1640, B27 supplement without insulin, 1 mM sodium pyruvate, 200 µM L-ascorbic acid 2-phosphate, and 5 µM IWR-1). The medium was changed on day 5. On day 7, after the beginning of differentiation, the medium was changed to cardiomyocyte culture medium (RPMI 1640, B27 with insulin, and 200 µM L-ascorbic acid 2-phosphate). The cells were cultured further until day 11 and were then passaged using 1 mL/well TrypLE™ Express (ThermoFisher) for 10 min at 37 °C, centrifuged at 300× g for 5 min, and plated into Geltrex™-coated 6-well plates. After passaging, the iPSC-cardiomyocyte cultures were exposed for 7 days to selection by adding a culture medium containing lactate instead of glucose (RPMI 1640 without glucose (Thermo Fisher), 0.2 mg/mL L-ascorbic acid 2-phosphate, 4 mM lactate/HEPES solution, and 0.5 mg/mL recombinant human albumin) in order to further enrich the population in iPSC-cardiomyocytes [9]. Then, the medium was changed back to the cardiomyocyte culture medium and the cells were fed every second day for the next 7 days. Finally, the cells were processed for further analysis.
2.7. Neural Differentiation
The iPSCs (iPSC#1.1 and iPSC#2.1) were differentiated into neurons, generally based on the method of Qi et al. [4], following a protocol that was published recently [20]. In brief, iPSC cultures with 60–80% confluence were exposed for the first 7 days to neural induction medium (DMEM/F12 (Thermo Fisher Scientific), 10% KnockOut serum (KOS; Thermo Fisher Scientific), 1% non-essential amino acids (NEAA, Thermo Fisher Scientific), 200 µM L-ascorbic acid (L-AA, Sigma-Aldrich), 2 µM SB431542 (Peprotech, Hamburg, Germany), 3 µM Chir99021 (Sigma-Aldrich), and 1.5 µM dorsomorphin (Peprotech)). After induction, the cells were passaged (0.25% Trypsin/EDTA (Thermo Fisher Scientific)) and transferred to Geltrex-coated plates. At day 7, the medium was changed to neuralization medium containing DMEM/F12, 100 µM L-AA, 1% NEAA, 1× N2 supplement (Thermo Fisher Scientific), 1× B27 supplement (Thermo Fisher Scientific), 10 ng/mL bFGF (Peprotech), and 10 ng/mL EGF (Peprotech). After the neuralization phase, the cells were passaged once on poly-L-Lysin/laminin-coated plates (1 µg/mL poly-L-Lysin in water (Sigma-Aldrich) for 30 min at 37 °C) (2 µg/mL laminin in PBS (Sigma-Aldrich; #11243217001) for 8 h at RT) and maintained for 7 days in neuronal differentiation medium I, (DMEM/F12, 100 µM L-AA, 1% NEAA, 1× N2 supplement, 1× B27 supplement, and 300 ng/mL cAMP (Peprotech). Finally, for the last 7 days, the cells were maintained in differentiation medium II (DMEM/F12, 100 µM L-AA, 1% NEAA, 1× N2 supplement, 1X B27 supplement, 300 ng/mL cAMP, 10 ng/mL BDNF (Peprotech), and 10 ng/mL NT-3 (Peprotech)).
2.8. Immunostaining
Cells were fixed with paraformaldehyde (4% (w/v) Merck) in PBS (Thermo Fisher) for 20 min at room temperature and subsequently washed at least three times with PBS. Then, the cells were incubated for 30 min in 1% bovine serum albumin (BSA; Thermo Fisher) supplemented with 0.1% Triton X-100 (Sigma-Aldrich). After blocking/permeabilization, the cells were incubated at 4 °C overnight with primary antibody solutions (Table 1). Subsequently, secondary antibody (Table 1) incubation was performed for 1 h at 37 °C, followed by incubation with 1 µg/mL DAPI (4′,6-diamidino-2-phenylindole) (Sigma-Aldrich) for 5 min at room temperature. Finally, the coverslips were mounted using Fluoromount–G (Thermo Fisher) and imaged with an epifluorescence microscope (Zeiss, Oberkochen, Germany).
2.9. Flow Cytometry
Eleven days after the beginning of differentiation, the Cyno-iPSC-derived cardiomyocytes were dissociated with 1 mL/well TrypLE™ Express (ThermoFisher) for 10 min at 37 °C.
The single cells were fixed for 10 min at room temperature with 4% (w/v) paraformaldehyde. Subsequently, blocking/permeabilization was performed with 1% BSA supplemented with 0.1% Triton X-100 in PBS at 4 °C overnight. Afterwards, the cells were incubated with conjugated α-actinin (Vio® R667) antibody solution (Table 1) at 37 °C for 1 h. Before analysis, the cells were washed with PBS and resuspended in 200 µL flow cytometry buffer (0.5% BSA, 2 mM EDTA (Carl Roth, Karlsruhe, Germany)). Flow cytometric analyses were performed with a SH800S Cell Sorter using a 100 µm chip (Sony Biotechnology, Weybridge, UK).
2.10. Nucleic Acid (DNA) Isolation and Polymerase Chain Reaction
Genomic DNA from the cell pellet samples was isolated using the DNeasy Blood & Tissue Kit (Qiagen, Hilden, Germany). The loss of episomal plasmids was demonstrated by a polymerase chain reaction (PCR) using plasmid-specific oligonucleotides (Sigma-Aldrich) (shown in Table 2) and polymerase with Standard Taq Buffer (New England Biolabs, Frankfurt, Germany). The different oligonucleotides and the PCR conditions were designed and validated for our previous study [9,16].
2.11. Protein Extraction and Western Blot Analysis
Protein from cultured cells (iPSCs) was isolated using RIPA lysis buffer (supplemented with Halt™ Protease Inhibitor Cocktail (100×), Thermo Fisher). The protein concentration in each sample was determined using the PierceTM BCA Protein Assay Kit (Thermo Fisher). For Western blot analysis, 10 µg of protein lysate (containing 1× DTT and 1× Laemmli sample buffer) was loaded onto a house-made 7.5% SDS-PAGE for protein separation. Following separation, the proteins were transferred onto an Immobilon®-P transfer membrane (Carl Roth, Karlsruhe, Germany). The membrane was then rinsed with TBS-T (1× TBS with 0.1% Tween-20) and blocked overnight at +4 °C using a solution of 5% powdered milk (Carl Roth) and TBS-T. Primary antibody incubation (Table 1) was carried out overnight at 4 °C. After washing with TBS-T, the membranes were incubated with a secondary HRP-conjugated antibody (Affinipure Goat Anti-rabbit IgG(H + L) and Anti-mouse (H + L) from Proteintech, no. SA00001-1 and SA00001-2). The Spectra™ Multicolor Broad Range Protein Ladder (Thermo Fisher) was used as a size reference. Signal detection was achieved using the chemoluminescent Pierce™ ECL Western Blotting Substrate (Thermo Fisher) and the ChemiDoc Western blot imaging system (Bio-Rad, Hercules, CA, USA).
3. Results
The aim of this study was to fine-tune the protocols to (1) generate iPSCs from an additional NHP species and (2) differentiate the iPSCs into clinically relevant cells under conditions that also work for human iPSCs [9]. We previously published a robust protocol to generate and characterize human, baboon, and rhesus macaque iPSCs. The present communication systematically tests the exact conditions in a different macaque species, the cynomolgus monkey, which represents the preferred NHP species in regulatory toxicity studies.
3.1. NHP and Human Fibroblast Reprogramming
We used tissue biopsies from two adult male macaques, hereinafter named Cyno#1 and Cyno#2, including the gingiva, foreskin, and skin. We successfully generated primary fibroblast lines from all tissues obtained from the two animals (exemplarily shown for Cyno#1 in Supplementary Figure S1a). For reprogramming, we selected an early passage of the fibroblast lines with a high proliferation rate and characteristic morphology. For one of the macaques, foreskin fibroblasts were selected (Cyno#1) for reprogramming, and for the other, the fibroblasts were derived from skin (Cyno#2). To induce pluripotency in the somatic cells, we used a set of episomal vectors for the transient expression of reprogramming factors [18]. Approximately 20 days after transfection, the first putative colonies became evident, and at day 30, colonies with a medium size were manually picked. As an early assessment of the pluripotency, alkaline phosphatase staining was performed in some of the primary plates on day 20. Alkaline phosphatase activity was detected in all cell clusters that showed primary colony morphology (Supplementary Figure S1b).
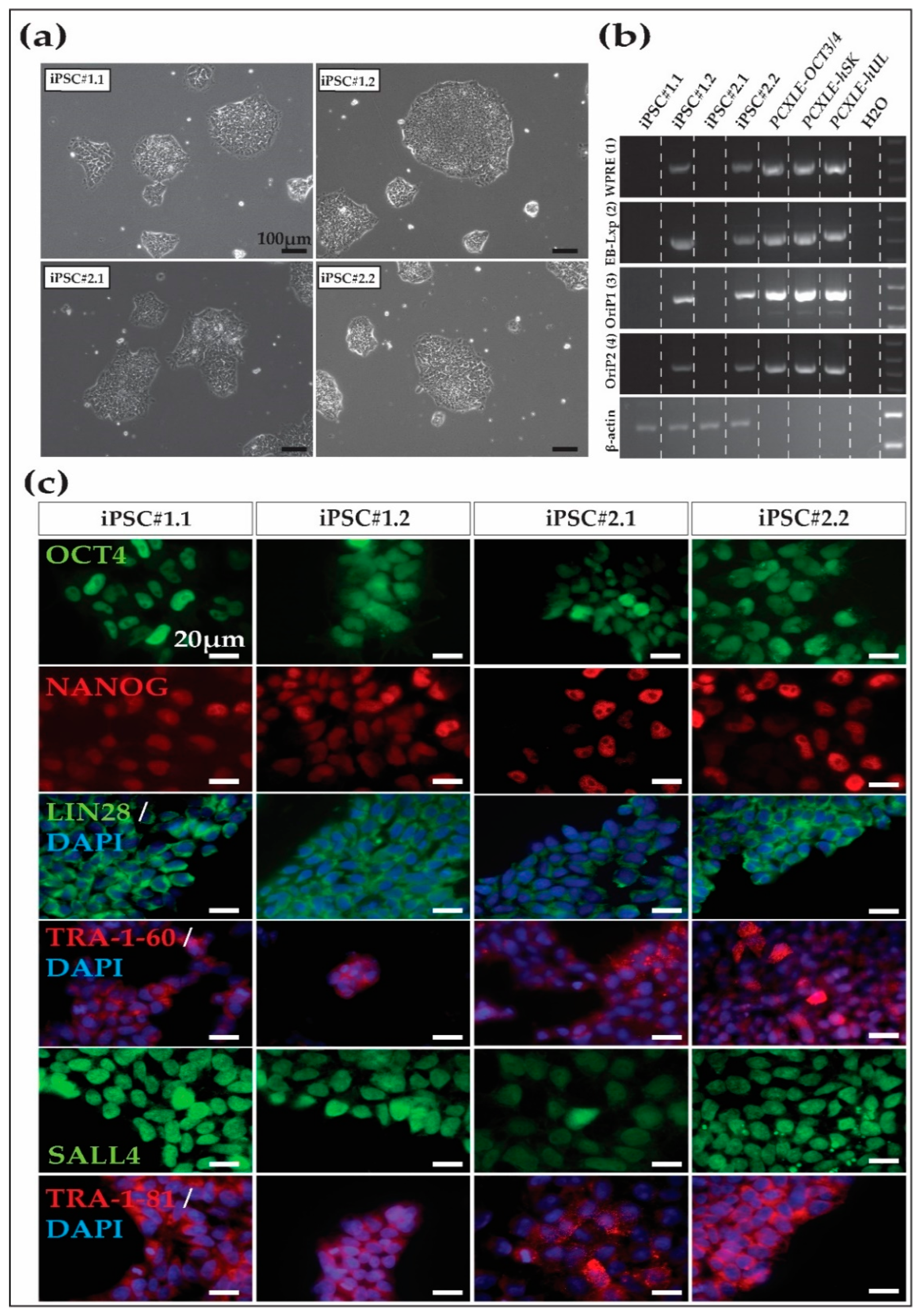
After the initial expansion of several putative iPSC lines until approximately passage five, two iPSC lines from each macaque were selected according to morphology for further passage, expansion, and characterization. The four iPSC lines (Cyno_iPSC#1.1 and Cyno_iPSC#1.2, from Cyno#1) (Cyno_iPSC#2.1 and Cyno_iPSC#2.2 from Cyno#2) showed the expected iPSC morphology, including compact colonies and sharp borders (Figure 1a). After ~15 passages, we evaluated the presence of the episomal vectors in the four iPSC lines by PCR. Two out of four cell lines showed no evidence of the transgenes at this passage (Figure 1b).
3.2. Cynomolgus Macaque iPSC Characterization
After derivation and expansion, the putative iPSC lines were characterized. Immunofluorescence analysis of the iPSCs revealed the expression of key pluripotency markers, including nuclear OCT4A, NANOG, SALL4, cytoplasmic LIN28, and the surface markers TRA-1-60 and TRA-1-81 (Figure 1c and Supplementary Figure S2 show nuclear marker colocalization with the nucleus). NANOG and SALL4 are not encoded by the reprogramming vectors.
To further characterize the cell lines, we analyzed the developmental potency of the cells to confirm their pluripotent state using an embryoid body formation assay. The cells of the four cell lines aggregated under defined conditions (Figure 2). Additionally, after exposing the cell aggregates to a differentiation medium for more than 20 days, representative cell types of the three embryonic germ layers were detected in the EB outgrowths. Representative SMA (alpha-smooth muscle actin and mesoderm), beta III tubulin (ectoderm), and AFP (alpha-fetoprotein and endoderm)-positive cells were found in all cell lines (Figure 2).
3.3. NHP-iPSC-Derived Cardiomyocyte and Neurons
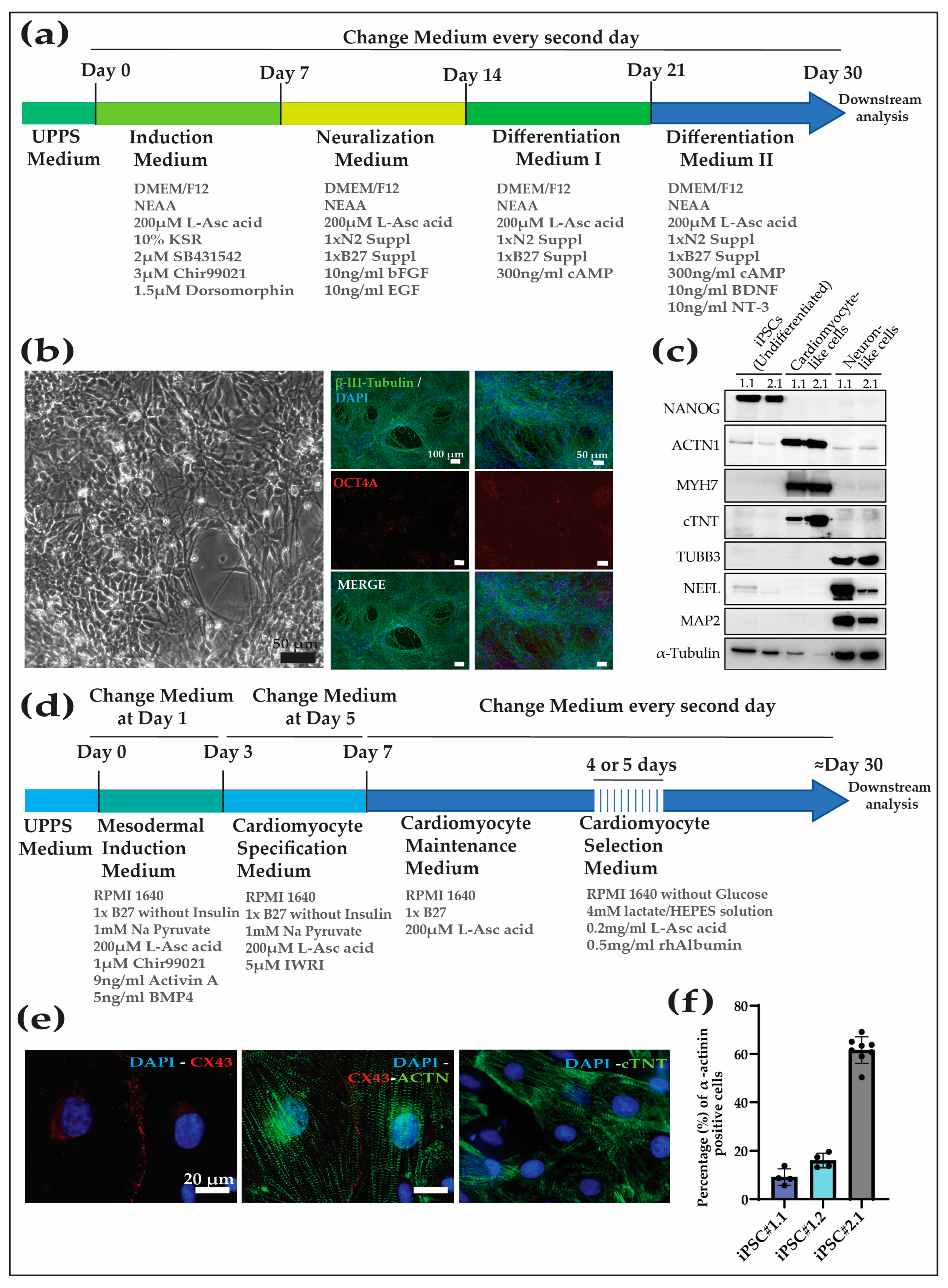
Different protocols are used to differentiate human iPSCs into tissue-specific cell types. However, one of the major difficulties in the field of preclinical testing is the translation of protocols of directed differentiation for human iPSCs to NHP-iPSCs. Previously, we have shown that rhesus macaque (Macaca mulatta) and baboon (Papio anubis) cells were refractory to directed differentiation following the protocols established for human cells [9]. After testing several approaches, we developed a protocol that was effective in human, rhesus macaque, and baboon cells. Here, we tested two different protocols in order to show (1) the translatability of those protocols to other macaque species and (2) further confirm the pluripotent state of the Cyno-iPSCs. As cardiovascular and neurodegenerative disorders have the highest clinical relevance, we decided to differentiate the Cyno-iPSCs exemplarily into neurons and cardiomyocytes.
For the neuronal differentiation protocol, we used our recently published method [20] (Figure 3a). The differentiation protocol efficiently directed the cells to the neural lineage (tested for Cyno_1.1 and 2.1), and, after approximately 30 days of differentiation, the cell population was highly enriched in cells showing neuronal morphology, the downregulation of pluripotency factors (e.g., OCT4A), and expressed neuron-specific markers like beta-III-tubulin (Figure 3b,c).
For the cardiomyocyte differentiation protocol, we followed a protocol based on that used in [19] (Figure 3d). The Cyno-iPSCs (Cyno_iPSC#1.1, 1.2, and 2.1) differentiated into contracting cells after about ten days. After differentiation, the percentage of cardiomyocytes in the cell population was evaluated by analyzing the percentage of alpha actinin-positive cells by FACS. We observed significant heterogeneity in the efficiency of the different cell lines to differentiate into iPSC-CM, as shown for Cyno_1.1, Cyno_1.2, and Cyno_2.1, with 12%, 16%, and 58%, respectively (the gating strategy and individual values are shown in Supplementary Figure S3). The differences in the propensity of the cell lines to differentiate into cardiomyocytes could be overcome after metabolic selection using lactate instead of glucose as the energy source. After 30 days in culture, the generated cardiomyocytes expressed cardiac-specific markers, as shown by staining and Western blot analysis for sarcomeric markers, and connexin 43, which is important for cell–cell communication between cardiomyocytes (Figure 3c,e,f).
4. Discussion
NHP are invaluable animal models to preclinically test cell replacement therapies. To generate predictive studies, it is necessary to count on a broad panel of iPSCs from the most relevant species in translational research. Old-world monkey species, like the rhesus macaque or baboon, are well established in this context; however, in the last few years, some key new-world monkey species, e.g., the marmoset, are gaining relevance. Additionally, to reinforce the translatability of such studies, the NHP-iPSCs need to meet the high standards of human iPSCs for generation and maintenance. Therefore, it is important to generate reprogramming protocols that are translatable between different primate species, including humans.
In this communication, we further expand the panel of primate species in which somatic cells can be reprogrammed following our previously established conditions using episomes and a chemically defined medium. Following this workflow, we report the generation of novel cynomolgus monkey iPSCs. This macaque species is of particular interest as it is the most widely used NHP species in regulatory testing and pharmaceutical research and development [12,13,14,15,21].
The generated novel iPSC lines present human iPSC-like morphology and cell behavior, including the potential to be expanded and maintained undifferentiated in UPPS medium for a high number of passages. Two of the novel cell lines are transgene-free, increasing their potential to be used by the scientific community and pharmaceutical industry in preclinical and regulatory studies in vivo and also in vitro, supporting and complementing in vivo studies. The four iPSC cell lines that have been characterized show the reactivation of the pluripotency network, as shown by the reactivation of pluripotency markers such as OCT4, NANOG, LIN28, SALL4, TRA-1-60, and TRA-1-81 revealed in the protein and transcript levels.
Additionally, the novel iPSC lines’ differentiation potential was first evaluated by spontaneous differentiation by performing an embryoid body assay. Upon differentiation, the four different iPSC lines were able to generate representative cells from the three embryonic layers. Additionally, we validated two differentiation protocols to direct cynomolgus macaque iPSCs toward enriched populations of clinically relevant cell types, i.e., cardiomyocytes and neurons. The cardiomyocyte protocol that was previously validated for humans, baboons, and rhesus macaques can also robustly generate Cyno-iPSC-derived cardiomyocyte-like cells. These cells spontaneously contract and express cell type-specific markers. The variable efficiencies in differentiation can be overcome later on by the metabolic selection of the cultures, generating highly enriched cardiomyocyte-like cell populations (Figure 3d,f). The neuronal differentiation protocol, which was previously validated only in human iPSCs, can generate neuronal-like cells that present a characteristic morphology and express cell type-specific markers.
5. Conclusions
In this study, we generated novel iPSC lines from an NHP species with high biomedical relevance. Additionally, we evaluated the capacity of the novel iPSC lines to be differentiated towards the neural and cardiac fates, proving this cell’s capacity to be considered for the allogeneic transplantation of ATMP in preclinical studies using cynomolgus macaques.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/cells13060558/s1, Figure S1: Cynomolgus macaque fibroblasts can be reprogrammed using episomal vectors; Figure S2: Pluripotency marker expression shown by immunofluorescence. Figure S3: Fluorescence-activated cell sorting (FACS) analysis of macaque iPSC-derived cardiomyocyte population purity before metabolic selection using α-actinin antibody.
Author Contributions
Conceptualization, I.R.-P. and R.B.; methodology, Y.T., N.E., I.R.-P., D.R., E.G.-R. and D.R.; formal analysis, Y.T., N.E. and I.R.-P.; resources, R.B.; data curation, Y.T., N.E. and I.R.-P.; writing—original draft preparation, I.R.-P., D.R. and E.G.-R.; writing—review and editing, R.B.; supervision, R.B.; funding acquisition, R.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research was partially funded by the Deutsches Zentrum für Herzkreislaufforschung (DZHK), grant number 81Z3300232 (to R.B.).
Institutional Review Board Statement
The study was conducted in accordance with the German Animal Welfare Act. The organ collection protocol was approved by the Institutional Animal Care and Use Committee (IACUC) of Charles River Laboratories Germany GmbH, Göttingen (protocol code 160-2019-005; date of approval 8 July 2019).
Informed Consent Statement
Not applicable.
Data Availability Statement
The cell lines generated in this study will be available upon request to the Research Platform Degenerative Diseases, German Primate Center-Leibniz Institute for Primate Research (to R.B.).
Acknowledgments
We thank Nicole Umland, Ulrike Gödecke, Angelina Berenson, and Simone Kalinowski for their excellent technical support of the work and Kerstin Zaft for administrative support. This work was supported by the German Centre for Cardiovascular Research (DZHK) and the German Primate Center, which is financed as a Leibniz Institute by the Bundesrepublik Deutschland and the Bundesländer (Federal states). The plasmids for reprogramming were provided by Addgene.
Conflicts of Interest
The authors Y.T., N.E., I.R.-P. and R.B., employed by the German Primate Center-Leibniz Institute for Primate Research, and the authors D.R. and E.G.-R., employed by Charles River Laboratories declare no conflict of interest.
References
- Takahashi, K.; Yamanaka, S. Induction of Pluripotent Stem Cells from Mouse Embryonic and Adult Fibroblast Cultures by Defined Factors. Cell 2006, 126, 663–676. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Tanabe, K.; Ohnuki, M.; Narita, M.; Ichisaka, T.; Tomoda, K. Induction of Pluripotent Stem Cells from Adult Human Fibroblasts by Defined Factors. Cell 2007, 131, 861–872. [Google Scholar] [CrossRef] [PubMed]
- Deinsberger, J.; Reisinger, D.; Weber, B. Global trends in clinical trials involving pluripotent stem cells: A systematic multi-database analysis. NPJ Regen. Med. 2020, 5, 15. [Google Scholar] [CrossRef] [PubMed]
- Qi, Y.; Zhang, X.-J.; Renier, N.; Wu, Z.; Atkin, T.; Sun, Z.; Ozair, M.Z.; Tchieu, J.; Zimmer, B.; Fattahi, F.; et al. Combined small-molecule inhibition accelerates the derivation of functional, early-born, cortical neurons from human pluripotent stem cells. Nat. Biotechnol. 2017, 35, 154–163. [Google Scholar] [CrossRef] [PubMed]
- Johnsen, D.O.; Johnson, D.K.; Whitney, R.A. Nonhuman Primates in Biomedical Research, 2nd ed.; Elsevier: Amsterdam, The Netherlands, 2012. [Google Scholar]
- Cox, L.A.; Olivier, M.; Spradling-Reeves, K.; Karere, G.M.; Comuzzie, A.G.; Vandeberg, J.L. Nonhuman Primates and Translational Research—Cardiovascular Disease. ILAR J. 2017, 58, 235–250. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Polo, I.; Behr, R. Non-human primate pluripotent stem cells for the preclinical testing of regenerative therapies. Neural Regen. Res. 2022, 17, 1867–1874. [Google Scholar] [CrossRef] [PubMed]
- Geuder, J.; Wange, L.E.; Janjic, A.; Radmer, J.; Janssen, P.; Bagnoli, J.W.; Müller, S.; Kaul, A.; Ohnuki, M.; Enard, W. A non-invasive method to generate induced pluripotent stem cells from primate urine. Sci. Rep. 2021, 11, 3516. [Google Scholar] [CrossRef] [PubMed]
- Stauske, M.; Polo, I.R.; Haas, W.; Knorr, D.Y.; Borchert, T.; Streckfuss-Bömeke, K.; Dressel, R.; Bartels, I.; Tiburcy, M.; Zimmermann, W.-H.; et al. Non-Human Primate iPSC Generation, Cultivation, Cardiac Differentiation under Chemically Defined Conditions. Cells 2020, 9, 1349. [Google Scholar] [CrossRef] [PubMed]
- Vermilyea, S.C.; Guthrie, S.; Meyer, M.; Smuga-Otto, K.; Braun, K.; Howden, S.; Thomson, J.A.; Zhang, S.-C.; Emborg, M.E.; Golos, T.G. Induced Pluripotent Stem Cell-Derived Dopaminergic Neurons from Adult Common Marmoset Fibroblasts. Stem Cells Dev. 2017, 26, 1225–1235. [Google Scholar] [CrossRef] [PubMed]
- Anwised, P.; Moorawong, R.; Samruan, W.; Somredngan, S.; Srisutush, J.; Laowtammathron, C.; Aksoy, I.; Parnpai, R.; Savatier, P. An expedition in the jungle of pluripotent stem cells of non-human primates. Stem Cell Rep. 2023, 18, 2016–2037. [Google Scholar] [CrossRef] [PubMed]
- Shen, H.; Yang, Z.; Rodrigues, A.D. Cynomolgus Monkey as an Emerging Animal Model to Study Drug Transporters: In Vitro, In Vivo, In Vitro-to-In Vivo Translation. Drug Metab. Dispos. 2022, 50, 299–319. [Google Scholar] [CrossRef] [PubMed]
- Cappon, G.D.; Bailey, G.P.; Buschmann, J.; Feuston, M.H.; Fisher, J.E.; Hew, K.W.; Hoberman, A.M.; Ooshima, Y.; Stump, D.G.; Hurtt, M.E. Juvenile animal toxicity study designs to support pediatric drug development. Birth Defects Res. B Dev. Reprod. Toxicol. 2009, 86, 463–469. [Google Scholar] [CrossRef] [PubMed]
- Bailey, G.P.; Wise, L.D.; Buschmann, J.; Hurtt, M.; Fisher, J.E. Pre- and postnatal developmental toxicity study design for pharmaceuticals. Birth Defects Res. B Dev. Reprod. Toxicol. 2009, 86, 437–445. [Google Scholar] [CrossRef] [PubMed]
- Iwasaki, K.; Uno, Y.; Utoh, M.; Yamazaki, H. Importance of cynomolgus monkeys in development of monoclonal antibody drugs. Drug Metab. Pharmacokinet. 2019, 34, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Polo, I.; Stauske, M.; Behr, R. Generation and cultivation of transgene-free macaque and baboon iPSCs under chemically defined conditions. In iPS CELLS; Turksen, K., Nagy, A., Eds.; Springer: New York, NY, USA, 2021; pp. 1–5. [Google Scholar]
- Rodriguez-Polo, I.; Stauske, M.; Becker, A.; Bartels, I.; Dressel, R.; Behr, R. Baboon induced pluripotent stem cell generation by piggyBac transposition of reprogramming factors. Primate Biol. 2019, 6, 75–86. [Google Scholar] [CrossRef] [PubMed]
- Okita, K.; Matsumura, Y.; Sato, Y.; Okada, A.; Morizane, A.; Okamoto, S.; Hong, H.; Nakagawa, M.; Tanabe, K.; Tezuka, K.-I.; et al. A more efficient method to generate integration-free human iPS cells. Nat. Methods 2011, 8, 409–412. [Google Scholar] [CrossRef] [PubMed]
- Tiburcy, M.; Hudson, J.E.; Balfanz, P.; Schlick, S.; Meyer, T.; Liao, M.-L.C.; Levent, E.; Raad, F.; Zeidler, S.; Wingender, E.; et al. Defined engineered human myocardium with advanced maturation for applications in heart failure modeling and repair. Circulation 2017, 135, 1832–1847. [Google Scholar] [CrossRef] [PubMed]
- Knorr, D.Y.; Polo, I.R.; Pies, H.S.; Schwedhelm-Domeyer, N.; Pauls, S.; Behr, R.; Heinrich, R. The cytokine receptor CRLF3 is a human neuroprotective EV-3 (Epo) receptor. Front. Mol. Neurosci. 2023, 16, 1154509. [Google Scholar] [CrossRef] [PubMed]
- Chellman, G.J.; Bussiere, J.L.; Makori, N.; Martin, P.L.; Ooshima, Y.; Weinbauer, G.F. Developmental and reproductive toxicology studies in nonhuman primates. Birth Defects Res. B Dev. Reprod. Toxicol. 2009, 86, 446–462. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Cynomolgus macaque iPSC morphology and characterization. (a) Morphology of the four iPSC lines generated in this study and maintained in universal primate pluripotent stem cell (UPPS) medium. iPSC#1.1 and iPSC#1.2 were from Cyno#1, while iPSC#2.1 and iPSC#2.2 were from Cyno#2. Cynomolgus macaque iPSC colonies present human primed iPSC-like morphology, including clear borders and compact colonies consisting of cells that show a high nucleus-to-cytoplasm ratio. Scale bars 100 µm. (b) Two out of four iPSC cell lines are transgene-free at passage ~15, as shown by PCR using primers specific for different regions of the episomal reprogramming plasmids. (c) Pluripotency marker expression, as shown by immunofluorescence. Note that NANOG and SALL4 are not encoded by the reprogramming vectors. Scale bars 20 µm.
Figure 1.
Cynomolgus macaque iPSC morphology and characterization. (a) Morphology of the four iPSC lines generated in this study and maintained in universal primate pluripotent stem cell (UPPS) medium. iPSC#1.1 and iPSC#1.2 were from Cyno#1, while iPSC#2.1 and iPSC#2.2 were from Cyno#2. Cynomolgus macaque iPSC colonies present human primed iPSC-like morphology, including clear borders and compact colonies consisting of cells that show a high nucleus-to-cytoplasm ratio. Scale bars 100 µm. (b) Two out of four iPSC cell lines are transgene-free at passage ~15, as shown by PCR using primers specific for different regions of the episomal reprogramming plasmids. (c) Pluripotency marker expression, as shown by immunofluorescence. Note that NANOG and SALL4 are not encoded by the reprogramming vectors. Scale bars 20 µm.

Figure 2.
Embryoid body formation assay. The four macaque cell lines form cell aggregates and differentiate into representative cell types of the three embryonic germ layers. Staining was conducted for the markers β-tubulin III (ectoderm), alpha-smooth muscle actin (SMA, mesoderm), and α-fetoprotein (AFP, endoderm) in embryoid body outgrowths generated with DPZ_Cyno#1.1, DPZ_Cyno#1.2, DPZ_Cyno#2.1, and DPZ_Cyno#2.2. Scale bars 100 µm and 20 µm.
Figure 2.
Embryoid body formation assay. The four macaque cell lines form cell aggregates and differentiate into representative cell types of the three embryonic germ layers. Staining was conducted for the markers β-tubulin III (ectoderm), alpha-smooth muscle actin (SMA, mesoderm), and α-fetoprotein (AFP, endoderm) in embryoid body outgrowths generated with DPZ_Cyno#1.1, DPZ_Cyno#1.2, DPZ_Cyno#2.1, and DPZ_Cyno#2.2. Scale bars 100 µm and 20 µm.

Figure 3.
Neuron and cardiomyocyte differentiation protocols were applied to the Cyno-iPSCs. (a) Directed neuronal differentiation of Cyno-iPSCs. (b) Bright-field and immunofluorescence images of neuronal cells. The Cyno-iPSC-derived neurons expressed beta-III-tubulin and showed the characteristic morphology (shown for iPSC#2.1). (c) Protein abundance analysis was conducted on undifferentiated iPSCs and their differentiated counterparts, including cardiomyocyte- and neuron-like cells (Cyno_iPSC#1.1 and 2.1). The analysis encompassed one pluripotency marker (Nanog) and three markers specific to cardiomyocytes (ACTN1, MYH7, and cTNT), as well as three neuronal-specific markers (TUBB3, NEFL, and MAP2). Alfa-tubulin expression is used as a housekeeping marker. (d) Cardiac differentiation of Cyno-iPSCs. (e) Cyno-iPSC cardiomyocytes express cardiac-specific proteins, as shown by immunofluorescence staining (shown for Cyno_iPSC#2.1). The immunofluorescence of cardiac proteins shows the typical sarcomeric structures in the Cyno-iPSC-derived cardiomyocytes: sarcomeric α-actinin, cardiac troponin T (cTNT), and connexin 43 (Cx43). Scale bars, 20 µm. (f) Additionally, the efficiency of differentiation was evaluated by assessing the percentage of alfa-actinin-positive cells in the post-differentiation, pre-selection cell populations (Cyno_iPSC#1.1, #1.2, and #2.1) by fluorescence-activated cell sorting (FACS).
Figure 3.
Neuron and cardiomyocyte differentiation protocols were applied to the Cyno-iPSCs. (a) Directed neuronal differentiation of Cyno-iPSCs. (b) Bright-field and immunofluorescence images of neuronal cells. The Cyno-iPSC-derived neurons expressed beta-III-tubulin and showed the characteristic morphology (shown for iPSC#2.1). (c) Protein abundance analysis was conducted on undifferentiated iPSCs and their differentiated counterparts, including cardiomyocyte- and neuron-like cells (Cyno_iPSC#1.1 and 2.1). The analysis encompassed one pluripotency marker (Nanog) and three markers specific to cardiomyocytes (ACTN1, MYH7, and cTNT), as well as three neuronal-specific markers (TUBB3, NEFL, and MAP2). Alfa-tubulin expression is used as a housekeeping marker. (d) Cardiac differentiation of Cyno-iPSCs. (e) Cyno-iPSC cardiomyocytes express cardiac-specific proteins, as shown by immunofluorescence staining (shown for Cyno_iPSC#2.1). The immunofluorescence of cardiac proteins shows the typical sarcomeric structures in the Cyno-iPSC-derived cardiomyocytes: sarcomeric α-actinin, cardiac troponin T (cTNT), and connexin 43 (Cx43). Scale bars, 20 µm. (f) Additionally, the efficiency of differentiation was evaluated by assessing the percentage of alfa-actinin-positive cells in the post-differentiation, pre-selection cell populations (Cyno_iPSC#1.1, #1.2, and #2.1) by fluorescence-activated cell sorting (FACS).


{kind=link}
{kind=link}
{kind=link}
Table 1.
List of primary and secondary antibodies.
| Name | Company | Catalog # | Dilution | |
|---|---|---|---|---|
| Primary Antibodies | α-feto protein | Dako (Glostrup, Denmark) | A0008 | 1:100 |
| α-actinin | Sigma-Aldrich | A7811 | 1:1000 | |
| β-tubulin III | Sigma-Aldrich | T8660 | 1:1000 | |
| Connexin 43 (Cx43) | Abcam (Cambridge, UK) | ab11370 | 1:1000 | |
| α-actinin * | Miltenyi Biotec | 130-128-591 | 1:50 | |
| Cardiac troponin T (cTNT) | Miltenyi Biotec | 130-106-687 | 1:10 | |
| LIN28 | R&D systems (Minneapolis, MN, USA) | AF3757 | 1:300 | |
| NANOG | Cell Signalling (Danvers, MA, USA) | 4903 | 1:400 | |
| OCT4A | Cell Signalling | C52G3 | 1:1600 | |
| TRA-1-81 | Abcam | ab16289 | 1:200 | |
| SALL4 | Abcam | ab57577 | 1:200 | |
| Smooth muscle actin | Sigma-Aldrich | A2547 | 1:1000 | |
| TRA-1-60 | Abcam | ab16288 | 1:200 | |
| α-actinin # | Sigma-Aldrich | A7811 | 1:2000 | |
| α-tubulin # | Cell Signalling | mAb#3873 | 1:5000 | |
| MYH7 # | Sigma-Aldrich | HPA001239 | 1:1000 | |
| Nanog # | Cell Signalling | D73G4 | 1:1000 | |
| NEFL # | Sigma-Aldrich | N4142 | 1:2000 | |
| MAP2 # | Sigma-Aldrich | HPA012828 | 1:2000 | |
| B III tubulin # | Biolegend (San Diego, CA, USA) | Biolegend | 1:2000 | |
| cTNT # | Miltenyi Biotec | 130-106-687 | 1:1000 | |
| Secondary antibodies | Alexa555-goat-α-mouse IgG | Thermo Fisher | A21424 | 1:1000 |
| Alexa488-goat-α-mouse IgG | Thermo Fisher | A11029 | 1:1000 | |
| Alexa488-goat-α-mouse IgG/IgM | Thermo Fisher | A10680 | 1:1000 | |
| Alexa488-donkey-α-goat IgG | Thermo Fisher | A11055 | 1:1000 | |
| Alexa488-donkey-α-rabbit IgG | Thermo Fisher | A21206 | 1:1000 |
(*) Antibodies used for flow cytometry analysis; (#) antibodies used for Western blot analysis.
Table 2.
Oligonucleotides used in this study. The table includes the name, sequence, amplicon size (Amp) in base pairs, annealing temperature (T, in °C), and cycles used during DNA amplification. Primers used for the detection of the episomal plasmids (1–4), and β-actin primers used as positive control.
Table 2.
Oligonucleotides used in this study. The table includes the name, sequence, amplicon size (Amp) in base pairs, annealing temperature (T, in °C), and cycles used during DNA amplification. Primers used for the detection of the episomal plasmids (1–4), and β-actin primers used as positive control.
| Name | Sequence | Amp | T | C |
|---|---|---|---|---|
| WPRE (1) | for: 5′-GCT ATT GCT TCC CGT ATG GC-3′ | 470 | 54 | 32 |
| rev: 5′-CAA AGG GAG ATC CGA CTC GT-3′ | ||||
| EBNA-LoxP (2) | for: 5′-AAG AGG AGG GGT CCC GAG A-3′ | 555 | 61 | 32 |
| rev: 5′-GCC AAT GCA ACT TGG ACG TT-3′ | ||||
| OriP1 (3) | for: 5′-GGT TCA CTA CCC TCG TGG AAT-3′ | 592 | 57 | 32 |
| rev: 5′-CGG GGC AGT GCA TGT AAT-3′ | ||||
| OriP2 (4) | for: 5′-GGT GAC TGT GTG CAG CTT TG-3′ | 416 | 54 | 32 |
| rev: 5′-GGA GCT GAG TGA CGT GAC AA-3′ | ||||
| β-actin | for: 5′-GAC CTG ACT GAC TAC CTC ATG-3′ | 380 | 61 | 32 |
| rev: 5′-GGT AGT TTC GTG GAT GCC ACA-3′ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Tereshchenko, Y.; Esiyok, N.; Garea-Rodríguez, E.; Repetto, D.; Behr, R.; Rodríguez-Polo, I. Transgene-Free Cynomolgus Monkey iPSCs Generated under Chemically Defined Conditions. Cells 2024, 13, 558. https://doi.org/10.3390/cells13060558
AMA Style
Tereshchenko Y, Esiyok N, Garea-Rodríguez E, Repetto D, Behr R, Rodríguez-Polo I. Transgene-Free Cynomolgus Monkey iPSCs Generated under Chemically Defined Conditions. Cells. 2024; 13(6):558. https://doi.org/10.3390/cells13060558
Chicago/Turabian StyleTereshchenko, Yuliia, Nesil Esiyok, Enrique Garea-Rodríguez, Daniele Repetto, Rüdiger Behr, and Ignacio Rodríguez-Polo. 2024. "Transgene-Free Cynomolgus Monkey iPSCs Generated under Chemically Defined Conditions" Cells 13, no. 6: 558. https://doi.org/10.3390/cells13060558
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.