

LC-DAD–ESI-MS/MS and NMR Analysis of Conifer Wood Specialized Metabolites

1
Department of Pharmacognosy and Molecular Basis of Phytotherapy, Medical University of Warsaw, 02-097 Warsaw, Poland
2
Doctoral School, Medical University of Warsaw, 02-091 Warsaw, Poland
3
Institut des Biomolécules Max Mousseron, Université de Montpellier, CNRS, ENSCM, 34293 Montpellier, France
4
Structural Studies Department, Centre of Molecular and Macromolecular Studies, Polish Academy of Sciences, 90-001 Łódź, Poland
*
Authors to whom correspondence should be addressed.
Cells 2022, 11(20), 3332; https://doi.org/10.3390/cells11203332
Submission received: 1 September 2022
/
Revised: 13 October 2022
/
Accepted: 20 October 2022
/
Published: 21 October 2022
(This article belongs to the Special Issue Profiling of Secondary Metabolites with Mass Spectroscopy-Based Methods)
Abstract
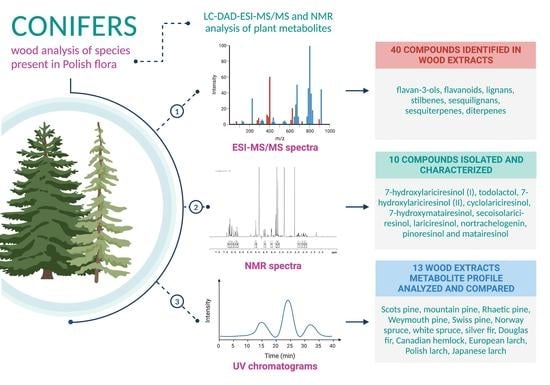
:Many species from the Pinaceae family have been recognized as a rich source of lignans, flavonoids, and other polyphenolics. The great common occurrence of conifers in Europe, as well as their use in the wood industry, makes both plant material and industrial waste material easily accessible and inexpensive. This is a promising prognosis for both discovery of new active compounds as well as for finding new applications for wood and its industry waste products. This study aimed to analyze and phytochemically profile 13 wood extracts of the Pinaceae family species, endemic or introduced in Polish flora, using the LC-DAD–ESI-MS/MS method and compare their respective metabolite profiles. Branch wood methanolic extracts were phytochemically profiled. Lignans, stilbenes, flavonoids, diterpenes, procyanidins, and other compounds were detected, with a considerable variety of chemical content among distinct species. Norway spruce (Picea abies (L.) H.Karst.) branch wood was the most abundant source of stilbenes, European larch (Larix decidua Mill.) mostly contained flavonoids, while silver fir (Abies alba Mill.) was rich in lignans. Furthermore, 10 lignans were isolated from the studied material. Our findings confirm that wood industry waste materials, such as conifer branches, can be a potent source of different phytochemicals, with the plant matrix being relatively simple, facilitating future isolation of target compounds.
1. Introduction
The Pinaceae family encompasses 11 genera and about 230 species, making it the largest family of conifers as well as the Gymnospermae. Genera include Abies (47 spp.), Cathaya (1 sp.), Cedrus (3 spp.), Keteleeria (3 spp.), Larix (11 spp.), Nothotsuga (1 sp.), Picea (38 spp.), Pinus (113 spp.), Pseudolarix (1 sp.), Pseudotsuga (4 spp.) and Tsuga (9 spp.). The members of this family are monoecious, resinous trees distributed widely in the Northern Hemisphere, with just one equatorial crossover species [1]. Although trees from the Pinaceae family occupy an area which is 68.4% of all forests in Poland [2], species diversity is not very large, with only a few species: Pinus sylvestris L. (Scots pine), Pinus mugo Turra (mountain pine), Pinus ×rhaetica Brügger (Rhaetic pine) and Pinus cembra L. (Swiss pine), Picea abies L. (Norway spruce), Larix decidua Mill. (European larch) and Larix polonica Rac. (Polish larch), Abies alba Mill. (silver fir) and few introduced species, of which only Pseudotsuga menziesii Mirb. (Douglas fir), Pinus strobus L. (Weymouth pine), Picea glauca (Moench) Voss (white spruce), Larix kaempferi Lamb. (Japanese larch) and Tsuga canadensis Carrière (Canadian hemlock) can be found in significant numbers [3]. Most of the genera present in Poland are economically important for the timber industry, which is why they have been preferred in plantings since the 19th century [2].
With recent technological advances in the timber industry, logs and other wood products brought to sawmills are almost totally processed, leaving little to no waste. However, this is still not always the case. According to experts, the low technological level of a large part of Polish industrial plants, the lack of financial resources for proper waste management, the state’s unclear ecological policy, as well as a low level of public awareness are the reasons for the generation of substantial amounts of waste [4]. By-products from the timber industry, such as wood chips, sawdust, bark, and others, form 38.6% of the total wood flow in Europe [5]. Their high quality and solid structure could bring them many applications, yet they are mainly used to generate energy (as a biofuel) and in wood panel production [5,6]. As more and more emphasis is placed on limiting the increase in global average temperature (i.e., global warming), with the Paris Climate Agreement, and the European Union’s 2030 and 2050 climate strategies, a new, more responsible and efficient approach to wood by-products is well founded.
One of such wood by-products are wood knots, which are branch bases inside a stem. They are not particularly of interest to the timber industry, as they are considered defects of the wood (fibers in this portion of wood are perpendicular to the stem), they are much harder to chip than normal wood, and their presence in wood pulp may lead to the formation of paper sheets with inferior strength, light absorption, and surface properties [7]. Therefore, it is not surprising that knotwood is considered a by-product of many timber industry processes, with little applications or value to-date; most of it is burned to produce energy [7]. Knotwood, as well as branchwood (which contains many knots on its own), due to their high lignan content have great potential for industrial applications. Not only can we find much higher amounts of common dietary lignans—some of them with proven dietary anti-diabetic, anti-inflammatory, antioxidant and phytoestrogenic properties [8,9,10,11]—but also some much rarer compounds, that is, oligolignans [12], which have not yet been thoroughly studied. Only recently, a single lignan molecule 7-hydroxymatairesinol (HMRlignan) from Picea abies wood knots was registered as a dietary supplement [13]. Although many lignans, such as secolariciresinol or sesamin, have been previously found in high concentrations in oilseeds, 7-hydroxymatairesinol has been detected in the human diet quite recently, using high-resolution detection methods [14]. Both 7-hydroxymatairesinol and Picea abies wood knots extract were found to improve glucose metabolism, decrease insulin resistance, and indirectly inhibit adipogenesis [15].
Extracts from Larix laricina K. Koch inner bark have been used by the Cree people of Northern Quebec to treat symptoms related to type 2 diabetes mellitus, and is mentioned in their traditional pharmacopeia. Recently both in vitro [16,17] and in vivo experiments [18] have confirmed the bark’s anti-diabetic effects, namely hypoglycemic, antioxidant and increasing insulin sensitivity. Although the exact compound responsible for this action is not yet known, lignans such as lariciresinol and its derivatives have been isolated using bioassay-guided fractionation from this extract [19].
Many species from the Pinaceae family have been recognized as a rich source, not only of lignans, but also of flavonoids and other polyphenolics. Phenolic standardized extracts from the bark of different species, such as Pinus pinaster Aiton (Pycnogenol, Flavangenol, Oligopin), and Abies alba Mill. (Abigenol), have been thoroughly studied and implemented in pharmaceutical use [20,21,22,23].
The bark of conifer trees, another wood industry by-product, is considered one of the major sources of stilbenes—a rare group of polyphenols of which resveratrol is perhaps the most studied [24]. These metabolites are not only considered phytoalexins (due to their antimicrobial activities) but also possess various pharmacological properties, such as anti-diabetic, anti-inflammatory, antioxidant, and cardioprotective [25,26,27]. They are not limited to the bark, as they have been also found in the wood of pines and spruces [28,29].
Although there have been studies screening the composition of some conifer wood, there are none on the composition of endemic and introduced conifers in Poland. Due to potential differences caused by geographical location, studying the wood of most common Polish conifers may bring some insight into their polyphenolic composition as well as their activity and further possibility of use in the food or pharmaceutical industry.
This study aimed to analyze and phytochemically profile wood extracts of the Pinaceae family species, endemic or introduced in Polish flora, using the LC-DAD–ESI-MS/MS (liquid chromatography with diode array detector coupled with electrospray ionization tandem mass spectrometry) method and compare their respective metabolite profiles. We also aimed to extract some of the more uncommon compounds to elucidate their structure under NMR (nuclear magnetic resonance) and use them as reference compounds in MS/MS (tandem mass spectrometry) analysis.
2. Materials and Methods
2.1. Chemicals and General Experimental Procedures
Methanol, chloroform, dichloromethane, ethyl acetate, formic acid, and n-hexane were purchased from POCh (Gliwice, Poland). Acetonitrile was purchased from Merck (Darmstadt, Germany). Water for HPLC experiments was prepared using the Milli-Q Plus system (Millipore, Billerica, MA, USA) (18.2 MΩ cm). All solvents used for chromatography were of gradient grade. Cyclolariciresinol, 7-hydroxylariciresinol (I), 7-hydroxylariciresinol (II), 7-hydroxymatairesinol, lariciresinol, matairesinol, nortrachelogenin, pinoresinol, secoisolariciresinol, and todolactol were isolated in our laboratory. Myricetin, dihydroquercetin (taxifolin), and epi-catechin were purchased from Serva (Heidelberg, Germany). Abietic acid and catechin were purchased from Sigma-Aldrich Chemie GmbH (Steinheim, Germany).
2.2. Plant Material and Extract Preparation
Branch wood of Pinus cembra L., Pinus mugo Turra, Pinus strobus L., Pinus ×rhaetica Brügger, Abies alba Mill., Picea abies L., Picea glauca (Moench) Voss, Pseudotsuga menziesii Mirb., Tsuga canadensis Carrière, and Larix kaempferi Lamb. was collected from the Polish Academy of Sciences Botanical Garden. Branch wood of Larix decidua Mill. and Larix polonica Rac. was collected from the University of Warsaw Botanical Garden. Pinus sylvestris L. branch wood was collected from Chojnów Landscape Park. The plant material was authenticated according to Flora Europaea by botanical gardens botanists. Voucher specimens have been deposited in the Plant Collection, Department of Pharmacognosy and Molecular Basis of Phytotherapy, Medical University of Warsaw, Poland. Plant material was dried at room temperature and shredded. Afterward, each sample was weighed (5.0 g), defatted with 50 mL n-hexane, and extracted with 50 mL of 90% (v/v) aqueous methanol under reflux for 2 h. The extract was then filtered through a 0.45 μm membrane, reduced in a rotary evaporator and lyophilized, resulting in dried methanolic extracts.
2.3. Isolation of Lignans
Pure lignans were isolated from the branch wood of Abies alba Mill. (660 g) and Pinus sylvestris L. (250 g). Briefly, the plant material was degreased with 1 L of n-hexane and extracted 3 times in a SONIC-5 ultrasonic bath (POLSONIC, Poznań, Poland) with 100% methanol at 60 °C for 2 h and evaporated to dryness on a Rotavator rotary evaporator R-100 (Buchi, Flawil, Switzerland) yielding 43.7 g and 11.5 g of dry silver fir and Scots pine extracts, respectively, which was phytochemically characterized.
The crude extract of fir was subjected to silica gel column chromatography (65 × 5 cm) and eluted with a CHCL3-MeOH gradient (100:0 → 90:10) of 11 steps, 0.5 L each, to obtain 100 fractions of 55 mL, which were pooled into 10 main fractions (F1–F10) based on their TLC (60G F254 silica gel, mobile phase dichloromethane and methanol (93:7, v/v), reagent 1% vanillin solution in concentrated sulfuric acid and heat treatment at 105 °C) and LC-DAD–ESI-MS/MS profiles.
Fraction F6 (5.98 g) was rechromatographed on a silica gel column (40 × 3 cm) with CHCL3-MeOH gradient (95:5 → 85:15) of 10 steps, 250 mL each, to obtain 5 fractions (F6_1–F6_5) of 500 mL each. Using preparative HPLC, lignans were isolated from fraction F6_1 (90 mg): compounds 22 (4.5 mg) and 27 (21.4 mg), from fractions F6_2 (230 mg), F6_3 (2660 mg), F6_4 (440 mg) and F6_5 (480 mg): compounds 16 (49.4 mg), 20 (157.8 mg) and 23 (1238 mg), and from directly from fraction F8 (2610 mg): compounds 9 (15.7 mg), 12 (54.5 mg) and 20 (24.1 mg).
The crude extract of Scots pine was suspended in water and extracted 3 times with ethyl acetate. Ethyl acetate fraction (8.4 g) was then subjected to Sephadex LH-20 (Pharmacia, Stockholm, Sweden) chromatography (40 × 3 cm) and eluted with methanol to obtain 34 fractions of 8 mL, which were pooled into 5 main fractions (F1–F5) based on their TLC and HPLC profiles. From fraction F2 (110 mg), compounds 29 (9 mg), 30 (13 mg), and 32 (13 mg) were isolated using preparative HPLC and identified using NMR methods.
These compounds were used as reference compounds for ESI-MS/MS analysis.
2.4. Preparative Chromatography
Preparative HPLC was performed with a Shimadzu LC20-AP instrument (Shimadzu, Japan) using a Zorbax SB-C18 column (150 mm × 21.2 mm, 5 μm) (Agilent, Santa Clara, CA, USA) at a flow rate of 20.0 mL/min and detection at λ1 = 254 nm, λ2 = 280 nm. The mobile phase consisted of 0.1% formic acid in water (A) and 0.1% formic acid in acetonitrile (B) using the following gradient: 0–60 min, 15–100% B.
2.5. General NMR Procedures
1H, 13C, and 2D NMR spectra (ROESY, COSY, HSQC, HMBC) were obtained on a Bruker Avance III 500 NMR spectrometer (Bruker BioSpin, Rheinstetten, Germany), operating at 500 and 126 MHz, respectively, using standard pulse programs and 5 mm NMR tubes. All measurements were performed at 295 K. Spectra were recorded in methanol-d4 or in CDCl3 (Armar AG, Döttingen, Switzerland). In each case, spectra were calibrated at residual solvent resonances, at 3.31 ppm for 1H and 49.15 ppm for 13C (methanol-d4, compounds 9, 12, 16, 20, 22, 23, 27, 30, 32) or 7.24 ppm for 1H and 77.23 ppm for 13C (CDCl3-d6, compound 29).
2.6. Phytochemical Characterization by LC-DAD–ESI-MS/MS Method
LC-DAD–ESI-MS/MS analysis was performed on a UHPLC-3000 RS system (Dionex, Dreieich, Germany) with DAD detection (Dionex, Dreieich, Germany) and an AmaZon SL ion trap mass spectrometer with an ESI interface (Bruker Daltonik GmbH, Bremen, Germany). Separation was performed on a Zorbax SB-C18 column (150 mm × 2.1 mm, 1.9 μm) (Agilent, Santa Clara, CA, USA). The mobile phase consisted of 0.1% formic acid in water (A) and 0.1% formic acid in acetonitrile (B) using the following gradient: 0–60 min, 15–100% B, then 10 min of equilibration. Samples for LC-DAD–ESI-MS/MS analysis were prepared by dissolving dried extracts in 0.1% formic acid in methanol at the concentration of 10 mg/mL. Standards were prepared in the same way at the concentration of 1 mg/mL). The flow rate was 0.2 mL/min, injection volume was 5 μL, column temperature was set at 25 °C. The LC eluate was introduced into the ESI interface without splitting, and compounds were analyzed in both positive and negative ion mode with the following settings: nebulizer pressure of 40 psi, drying gas flow rate of 9 L/min, nitrogen gas temperature of 300 °C, and a capillary voltage of 4.5 kV. The mass scan ranged from 100 to 2200 m/z. Low-energy collision-induced dissociation (less than 100 eV) was used to obtain MS/MS spectra, with collision energies chosen automatically based on precursor ion masses by the integrated SmartFrag mode.
UV spectra were recorded in the range 190–400 nm. Compounds were identified by comparing their retention time and UV-visible and mass spectra with those obtained from reference compounds and/or tentatively identified by comparison with literature information.
3. Results
Through comprehensive LC-DAD–ESI-MS/MS analysis of Pinus sylvestris L., Pinus cembra L., Pinus mugo Turra, Pinus strobus L., Pinus ×rhaetica Brügger, Abies alba Mill., Picea abies L., Picea glauca (Moench) Voss, Pseudotsuga menziesii Mirb., Tsuga canadensis Carrière, Larix decidua Mill., Larix polonica Rac., and Larix kaempferi Lamb. branch wood methanolic extracts, 40 compounds were identified or partially identified based on the elution order, UV maxima, pseudomolecular and fragmentation ions in both positive and negative ion modes and comparison to literature data (Table 1). Presence of some flavan-3-ols, flavonoids and abietic acid was confirmed by comparison with commercial standards. In the case of lignans, most of the compounds were isolated and their structures were confirmed by NMR analysis. Then, their presence in selected wood extracts was confirmed by LC-DAD–ESI-MS/MS analysis by comparison with pure compounds.
The investigated plant material contained specialized metabolites belonging to flavan-3-ols, flavonoids, lignans, sesquilignans, stilbenes, sesquiterpenes, and diterpenes. The presence of identified compounds in each studied species is presented in (Table 2). The UV chromatograms of each wood extract are provided in Supplementary Materials (Figures S1–S13).
3.1. Flavan-3-ols
Eight compounds belonging to the group of flavan-3-ols were identified in selected wood extracts. Compounds 5 (tr = 4.5 min) and compound 7 (tr = 6.6 min) exhibited a pseudomolecular ion at m/z 289 and had identical fragmentation patterns. Based on their elution order and comparison to reference standards, these compounds were identified as catechin and epi-catechin, respectively. Whilst catechin was present in all studied plant samples, we were not able to detect epi-catechin in P. sylvestris, P. mugo, P. ×rhaetica, and both spruces (Table 2).
Four compounds, 2 (tr = 3.3 min), 3 (tr = 3.6 min), 6 (tr = 5.2 min) and 15 (tr = 11.6 min), exhibited a pseudomolecular [M−H]− ion at m/z 577, which fragmented into m/z 559 [M−H−18]—loss of water, m/z 451 [M−H−126]− from heterocyclic ring fissure fragment, m/z 407 [M−H−170]−—product of retro-Diels–Alder reaction followed by the loss of water and m/z 289, which corresponds to (epi)catechin. Based on fragmentation patterns, they were all characterized as type B dimeric procyanidins [41]. Previously, procyanidins B1-B4 have been isolated from the bark of Larix gmelinii (Rupr.) Kuzen. [42] and Pseudotsuga menziesii Mirb. [31]. In each plant sample, at least two dimeric procyanidins B were present. Larch wood was the richest in procyanidins, containing all four compounds in comparison to all studied genera (Table 2).
Additionally, one more procyanidin was detected in all wood extracts—compound 4 (tr = 4.2 min). The main pseudomolecular ion was at m/z 865 and fragmented into m/z 695, m/z 577, m/z 407 and m/z 289. Based on the fragmentation pattern [43], this compound was assigned as type B trimeric procyanidin (procyanidin C).
3.2. Flavonoids
A total of 10 flavonoids were found in studied wood extracts. Most of them were present in aglycone form, with just one glycoside form detected. Compound 18 (tr = 12.4 min) was the most abundant flavonoid in these extracts, as it was not present only in A. alba and T. canadensis (Table 2), and had the highest peak of all flavonoid aglycones. Compound 18 had its UV maximum at 280 nm and exhibited a pseudomolecular ion at m/z 303, which fragmented into m/z 285 and m/z 177. The compound was then identified by comparison with the reference standard and comparison to the literature data [44,45,46] as taxifolin (syn. dihydroquercetin). Another major flavonoid compound 13 (tr = 10.2 min) had its UV maximum at 287 nm and its pseudomolecular ion at m/z 465 and gave a primary fragment ion at m/z 303 [M−H−162]− corresponding to the cleavage of hexose (probably glucose). Thus, compound 13 was assigned as taxifolin hexoside, which was consistent with the fragmentation pattern reported previously [44].
Four more flavanonols were detected in Pinaceae wood, with their UV maxima at around 290 nm. Compound 8 (tr = 7.7 min) exhibited a pseudomolecular ion at m/z 319 and fragment ions at m/z 301 [M−H−18]− corresponding to the loss of water, at m/z 193 and m/z 125. By comparing fragmentation pattern with literature data [47], compound 8 was assigned as dihydromyricetin (syn. ampelopsin, ampeloptin). It was found in samples from P. mugo, P. rhaetica, L. decidua, and L. polonica (Table 2). Compound 11 (tr = 8.4 min) had its pseudomolecular ion at m/z 287, which fragmented into m/z 259 [M−H−28]−, corresponding to the loss of CO, m/z 243 [M−H−44]−, corresponding to the loss of CO2 and m/z 181. The fragmentation pattern matched to the literature data of dihydrokaempferol (syn. aromadendrin) [47]. Dihydrokaempferol was only found in two larch species, namely European and Polish larch (Table 2). Compound 33 (tr = 22.0 min) exhibited a pseudomolecular ion at m/z 271 and fragment ion at m/z 253 [M−H−18]− corresponding to the loss of water. It was tentatively identified, comparing to literature data [48], as pinobanksin, and compound 36 (tr = 30.5 min), which exhibited a similar fragmentation pattern, was assigned as pinobanksin 3-O-acetate, also consistent with the data found in the literature [49]. The presence of pinobanksin was not genus-specific, while pinobanksin 3-O-acetate was only found in P. strobus wood (Table 2).
We have found presence of two flavanones, compound 24 (tr = 15.4 min) and 35 (tr = 30.3 min), which had their UV maxima at around 290 nm. Compound 24 presented a similar fragmentation pattern to that of compound 11. Based on the elution profile and referring to the literature [50], it was tentatively assigned as eriodictyol. It was present in all larch species, Douglas fir, mountain pine, Swiss pine, and Weymouth pine. Pseudomolecular ion of compounds 35 was at m/z 255 and fragmented into m/z 213 and m/z 211. By comparison with the literature data [51] it was assigned as pinocembrin. We have found its presence in most pine species (apart from P. cembra) and in Douglas fir (Table 2).
We report presence of two flavonols in studied wood extracts, which were characterized by their UV maxima at around 370 nm. Compounds 25 (tr = 15.7 min) and 31 (tr = 19.8 min) had typical fragmentation patterns of myricetin and quercetin, respectively [52], which was confirmed by comparison with reference standards. Quercetin and myricetin were found in larches and Douglas fir. Additionally, Weymouth pine contained quercetin, while myricetin was also present in mountain pine (Table 2).
3.3. Lignans
Lignans have been identified as major constituents in fir and pine species. However, the spectral data for this class of compound is rather limited, thus, MS/MS analysis of this group of plant metabolites had to be supported by the isolation of studied compounds and elucidation of their structures by NMR. Ten lignans have been isolated using chromatographic methods, namely 7-hydroxylariciresinol (I) (9), todolactol (12), 7-hydroxylariciresinol (II) (16), cyclolariciresinol (20), 7-hydroxymatairesinol (22), secoisolariciresinol (23), lariciresinol (27), nortrachelogenin (29), pinoresinol (30) and matairesinol (32), with their structures confirmed through NMR analysis and comparison with literature data [53,54,55,56]. Isolated compounds were then used as reference standards in phytochemical profiling of conifer wood extracts.
The 1D NMR and ESI-MS/MS spectra along with NMR chemical shifts of all isolated item are presented in Supplementary Materials (Figures S94−S103; Table S1).
Based on the LC-DAD–ESI-MS/MS analysis, three compounds, 9 (tr = 7.8 min), 12 (tr = 9.9min), and 16 (tr = 11.6 min), had their pseudomolecular ion at m/z 375 [M−H]− and UV maxima at around 225 and 280 nm. There have been previously reported three lignan isomers in conifer wood, that could give pseudomolecular ion at m/z 375: liovil, todolactol and 7-hydroxylariciresinol [57]. Unfortunately, no ESI-MS/MS fragmentation pattern data could be found for liovil and 7-hydrolariciresinol., with just one source for todolactol [57]. Through NMR analysis, we were able to assign compounds 9 and 16 as 7-hydroxylariciresinol diastereoisomers, respectively. Fragmentation of m/z 375 ion corresponding to compound 12 gave m/z 327, m/z 191, and m/z 176 which was consistent with the fragmentation pattern proposed for todolactol [57], and in agreement with fragmentation of our isolated standard. The occurrence of lignans in different conifer species was as follows: 7-hydroxylariciresinol (I) was found in silver fir and both spruces, todolactol was additionally present in European and Japanese larch, Douglas fir and Canadian hemlock. The lignan 7-hydroxylariciresinol (II) was present in at least one species of each genus except spruces (Table 2). LC-DAD–ESI-MS/MS analysis did not detect the signal that could correspond to liovil.
Compounds 30 (tr = 19.1 min) and 32 (tr = 21.6 min) exhibited pseudomolecular ions at m/z 357 [M−H]− and similar fragmentation patterns. Compound 32 gave a characteristic primary product ion for dibenzylbutyrolactone lignans [M−H−44]− at m/z 313, which corresponds to the loss of CO2 from the lactone ring [57], and that was not observed for compound 30. Compound 30 produced a fragment ion at m/z 342 [M−H−15]− corresponding to the loss of methyl radical, and at m/z 151 (guaiacyl) and m/z 136, which were the products of α,β-cleavage in the side chain. This fragmentation pattern was characteristic for furofuran lignans [57]. According to the fragmentation, elution order in RP-LC, and comparison with reference standards, compound 30 was identified as pinoresinol, while compound 32 was identified as matairesinol. Both compounds were also isolated, and their structure was confirmed through NMR analysis. Pinoresinol was only present in Scots pine wood and matairesinol could be found in nearly every plant material except silver fir and Douglas fir (Table 2).
Compounds 22 (tr = 15.0 min) and 29 (tr = 18.3 min) exhibited their pseudomolecular ions at m/z 373 [M−H]−. They both formed a fragment ion at m/z 355 [M−H−18]− corresponding to the loss of water. For compound 22, a fragment ion at m/z 311 [M−H−62]− could be found, corresponding to the loss of water and CO2 from the lactone ring (again suggesting the compound belonged to the dibenzylbutyrolactone group). On the other hand, compound 29 did not produce a fragment ion at m/z 311; instead, an ion at m/z 327 [M−H−46]− which corresponds to the loss of water and CO was observed,. Both fragmentation patterns and differences among them have been previously observed [58]. According to the fragmentation, elution order in RP-LC, and comparison with reference standards, compound 22 was identified as 7-hydroxymatairesinol and compound 29 was identified as nortrachelogenin (syn. 8′-hydroxymatairesinol). They were also isolated and their structure was confirmed using NMR methods. Nortrachelogenin and 7-hydroxymatairesinol were observed in most analyzed conifer wood, only excluding mountain and Rhaetic pine for the former, and additionally excluding Scots pine for the latter. Compound 21 (tr = 14.0 min) had a pseudomolecular ion at m/z 419 [M−H+HCOOH]− which fragmented into m/z 373 [M−H]− corresponding to the loss of formate adduct. Based on literature data, it was tentatively identified as α-conidendric acid [57]. It was found in silver fir, mountain pine, Weymouth pine, all larch species, and Canadian hemlock (Table 2).
Compound 27 (tr = 16.1 min) had its pseudomolecular ion at m/z 405 which fragmented to m/z 359 [M−H]−, corresponding to the presence of formyl adduct. We also observed another compound, 20 (tr = 12.7 min), exhibiting its pseudomolecular ion at m/z 359 [M−H]−. Differences in further fragmentation could be observed for these compounds. For peak 27 we could observe a fragment ion at m/z 329 [M−H−30]− resulting from the loss of formaldehyde. Compound 20 exhibited fragment ions at m/z 344 [M−H−15]−, corresponding to the loss of methyl radical, at m/z 313 [M−H−46]−, formed by the loss of methoxyl and methyl radicals, and at m/z 189, which was a product of β-cleavage in the side chain. Both fragmentation patterns have been previously observed [57] for lariciresinol (27) and cyclolariciresinol (syn. isolariciresinol) (20), respectively, which was then confirmed by isolating these compounds and elucidating their structure through NMR analysis. Cyclolariciresinol was only found in silver fir, while lariciresinol was present in all larch species, silver fir, Douglas fir, Weymouth pine, Canadian hemlock, and white spruce (Table 2).
Apart from cyclolariciresinol (20), one other butanediol lignan has been found. For compound 23 (tr = 15.3 min) a pseudomolecular ion could be observed at m/z 361 [M−H]− and fragment ions at m/z 346 [M−H−15]−, corresponding to the loss of methyl radical, at m/z 313 [M−H−48]−, formed by the loss of formaldehyde and water in the diol structure and at m/z 299 [M−H−48−15]−, corresponding to both rearrangements. This structure was compared to fragmentation data described in the literature [57], isolated, and identified as secoisolariciresinol. This was the dominant metabolite of silver fir and was also found in all larch and spruce species, mountain and Weymouth pine, Douglas fir, and Canadian hemlock (Table 2).
3.4. Sesquilignans
Two sesquilignans were found in conifer wood. Compound 26 (tr = 15.7 min) exhibited a pseudomolecular ion at m/z 557 [M−H]− and fragment ions at m/z 539 [M−H−18]− corresponding to the loss of water, m/z 525 [M−H−32]−, m/z 521 [M−H−36]−, m/z 509 [M−H−48]−, formed by the loss of formaldehyde and water in the diol structure, m/z 415 and m/z 361 [M−H−196]−, corresponding to the cleavage of guaiacylglyceryl moiety. The fragmentation pattern was in accordance with the data for oligolignans isolated from Norway spruce and Scots pine knots [12]. Based on fragmentation, this compound was tentatively identified as secoisolariciresinol 4-O-guaiacylglyceryl ether. It was found in silver and Douglas fir, as well as all larch species and mountain pine (Table 2).
Compound 28 (tr = 16.7 min) had its pseudomolecular ion at m/z 555 [M−H]− and fragment ions at m/z 525 [M−H−20]−, m/z 507 [M−H−48]− from the loss of formaldehyde and water, m/z 359 [M−H−196]−, corresponding to the cleavage of guaiacylglyceryl moiety, and at m/z 329 [M−H−196−30]−, resulting from the loss of formaldehyde. This compound was tentatively identified as lariciresinol 4-O-guaiacylglyceryl ether in accordance to previous MS/MS data [12]. It was only found in silver fir wood (Table 2).
3.5. Stilbenes
The Pinaceae family, is known to be a rich source of stilbenes [28,29,41]. In our study we found six stilbenes in conifer wood, mostly in spruce and pine species. They had characteristic UV maxima either at around 320 nm (glycosidic form) or 300 nm (aglycone form). Compounds 10 (tr = 8.1 min) and 14 (tr = 10.8 min) exhibited two pseudomolecular ions at m/z 811 [2M−H]− and m/z 405 [M−H]− with similar fragmentation patterns. Primary ion at m/z 405 fragmented into m/z 243 [M−H−162]−, corresponding to the cleavage of hexose, and showed fragments at m/z 225 [M−H−18]−, corresponding to the loss of water, m/z 201 [M−H−42]−, corresponding to the losses of C2H2O, characteristic for stilbenoids, m/z 173 [M−H−42−28]−, and m/z 159 [M−H−42−42]−. Based on elution order and comparison with literature data [28,59,60,61], compounds 10 and 14 were identified as trans-astringin (piceatannol 3-O-glucoside) and cis-astringin, respectively. Trans-astringin was only observed in spruce species, while cis-astringin was also present in larch species (Table 2).
Compounds 17 (tr = 11.7 min) and 19 (tr = 12.5 min) were only found in spruces. The main pseudomolecular ion for compound 17 was at m/z 389 [M−H]− and gave one fragment ion at m/z 227 [M−H−162]−, corresponding to the cleavage of hexose, and showed fragments of aglycon at m/z 185 [M−H−42]− and 183 [M−H−42−H2]−, corresponding to the losses of C2H2O, characteristic for stilbenoids, m/z 157 [M−H−42−28]− and m/z 143 [M−H−42−42]− [61]. It was tentatively identified as piceid (resveratrol 3-O-glucoside) in comparison with literature data [60]. Compound 19 exhibited a pseudomolecular ion at m/z 465 [M−H+HCOOH]− which showed fragment ions at m/z 419 [M−H]−, corresponding to the loss of formyl adduct, at m/z 257 [M−H−162]−, corresponding to the cleavage of hexose, at m/z 242 [M−H−162−15]− and at m/z 241 [M−H−162−16]− [28]. It was tentatively identified as isorhapontin (isorhapontigenin 3-O-glucoside).
Compounds 34 (tr = 26.3 min) and 37 (tr = 33.6 min) were characterized as stilbenes based on their primary pseudomolecular ion and UV maxima at around 300 nm. Comparing mass of [M−H]− ion at m/z 211 [M−H−]− and m/z at 225 [M−H]− with literature reports on stilbenes isolated previously from conifer wood and their elution order [53], these compounds were tentatively identified: 34 as pinosylvin and 37 as pinosylvin monomethyl. Moreover, compared to MS/MS data from the literature, pinosylvin (34) demonstrated a fragment ion at 169 m/z [M−H−42]− corresponding to the losses of C2H2O, characteristic for stilbenoids [59,62]. Pinosylvin monomethyl ether (37) pseudomolecular ion gave a fragment ion at 210 m/z [M−H−15]−, corresponding to the loss of methyl radical. Their occurrence was limited to the genus Pinus, with only Scots pine and mountain pine containing all the above (Table 2).
3.6. Sesquiterpenoids and Diterpenoids
We were able to identify one sesquiterpenoid and two diterpenes in conifer wood— all of them in positive ion mode. Compound 38 (tr = 41.0 min) exhibited a pseudomolecular ion at m/z 265 [M+H]+ and fragmented into m/z 251, m/z 233, m/z 205, m/z 187 m/z 176 and m/z 83. It was tentatively identified as dehydrojuvabione by comparison with MS/MS data found in literature [63]. Compounds 39 (tr = 52.0 min) and 40 (tr = 52.3 min) both exhibited a pseudomolecular ion at m/z 303 [M+H]+ with similar fragmentation patterns: m/z 285, m/z 257 and m/z 123. Based on UV maxima, 251 nm for compound 39, and 241 nm for compound 40, their elution order, comparing this data to the literature [64,65] and the reference standard, we were able to identify compound 39 as neoabietic acid and compound 40 as abietic acid. Dehydrojuvabione was only found in A. alba while both abietane-type resin acids were present in all studied conifer species wood (Table 2).
4. Discussion
Conifer wood and waste materials from its processing in the paper and wood industry (such as knots, bark and branches) have been identified as a good source of many polyphenols. Willfor and his team characterized wood knots of spruces, pines and firs, isolating many lignans in the process [34,39,66,67]. More recently, Gabaston et al. not only characterized wood knots of Pinus pinaster Aiton using LC-MS/MS, but also studied their antifungal activity against grapevine pathogens [53]. Another fine work offered LC-MS/MS analysis of stilbenes in Picea jezoensis Carrière bark [28]. To the best of our knowledge, most of the wood studied here was never characterized before using LC-MS/MS techniques. This study was also the first attempt at phytochemical profiling of wood from P. ×rhaetica and L. polonica, as well as the first to compare the composition of Pinaceae species growing in Poland.
Referring to previous research, our study confirms that conifers from the Pinaceae family are rich in otherwise scarce compounds, such as lignans and stilbenes, offering a large variety of these structures.
Lignans are a group of natural polyphenols (and one of the most lipophilic), located in plant cell walls. According to the recent nomenclature: lignans dimers of two coniferyl, sinapyl, 4-hydroxycinnamyl alcohol or similar alcohol monomers. Some authors restrict the term lignan only to those molecules coupled by the central carbon of the sidechain (i.e., 8,8′ or β, β’ dimers) [68]. They were first identified in conifer wood, which is why they often obtain their names from those species, e.g., pinoresinol from Pinus nigra Aiton and lariciresinol from Larix decidua Mill. [69]. Although lignans have been detected in numerous plants, with many being part of human diet, their content is usually very low. Apart from conifer wood knots (where they can accumulate in amounts up to 30% (w/w)), flaxseed and sesame seeds are considered rich but challenging sources of lignans. In this context, studied wood from the Pinaceae family offers a much easier-to-extract source of these polyphenols, with much fewer steps required to obtain pure compounds (directly in aglycone form with much larger structure variety). Wood from all studied species contained detectable amounts of lignans, with silver fir being the richest source of these polyphenols. Although A. alba offered large quantities of lignans, especially secoisolariciresinol and lariciresinol, P. sylvestris was far richer than all other species in nortrachelogenin, pinoresinol, and matairesinol. This is particularly interesting, as Scots pine is the most abundant and economically most important tree in Europe. Another important notion from our study is the detection of a lignan (namely matairesinol) in P. cembra, which was previously described as the only pine not containing lignans [7].
Through our analysis, we did not detect few lignans that have been previously found in some of the studied species. Willfor et al. reported presence of matairesinol, pinoresinol, and nortrachelogein in A. alba wood knots [34], but we did not observe any signal at expected retention times. In the same study, presence of secoisolariciresinol monomethyl ether and dimethyl ether was established. We cannot rule out the presence of these compounds in our samples, as pseudomolecular ions [M−H]− with expected mass for these lignans were observed. Unfortunately, no fragmentation pattern for them could be found in the literature, nor could we isolate them from the plant material. Similar problems occurred with analysis of sesquilignans, for which only few fragmentation patterns could be found, thus limiting our ability to identify them through simple LC-MS/MS analysis. Noticing that many lignans and sesquilignans did not have ESI fragmentation patterns reported previously, our study fills this knowledge gap for at least some of these compounds.
Species from the Pinaceae family are considered as one of the richest in stilbenes content. These rare polyphenols consist of two aromatic rings linked by a C2 ethylene sidechain, formed through cyclization of 3 malonyl-CoA with cinnamoyl-CoA, coumaroyl-CoA or caffeoyl-CoA, followed by hydroxylation of aromatic rings [70]. It has been previously reported that stilbenes are abundant constituents of P. abies and P. jezoensis bark, with their levels in wood being much lower [28,32]. Hovelstad et al. reported no presence of stilbenes in P. abies heartwood and knotwood; however, their analysis was limited to pinosylvins [29]. Our study offered analysis of stilbenes from different groups and included species not screened for stilbenes before. We have shown presence of stilbenes in all studied pines, spruces, and larches wood extracts, though their structure and nature varied among species. Stilbenes present in pine wood belonged to the pinosylvins, which were more lipophilic than those found in spruce and larch wood, which mostly occurred in glycosidic form. In the case of spruces, the UV signal for trans-astringin was much stronger than that of most abundant lignan reported in these species, i.e., 7-hydroxymatairesinol, which could imply higher content of stilbenes than lignans in spruce wood.
As was the case with lignans, fragmentation patterns for stilbenes are rarely reported in the literature, thus making LC-DAD–ESI-MS/MS analysis of these compounds difficult. This study offers MS2 fragmentations for some of these compounds, yet we see the need for further exploration of ESI fragmentation. Additional focus should be directed at differentiation of cis/trans isomers and the site of glucose substitution. In the case of our study, no differences were observed for such compounds and identification had to be based on elution order.
Our analysis also offered an insight into flavonoid, flavan-3-ol, and diterpene composition of conifer wood. These compounds were present in all studied samples, with differences concerning structures and their exact content. Larches and Douglas fir could be considered the richest in flavonoids and flavan-3-ols.
5. Conclusions
Overall, results obtained from this study show that conifer wood is a significant source of different phytochemicals, with many rare structures present. Silver fir wood could be a rich source of lignans, pine wood offer both lignans and stilbenes aglycones, while spruce wood had stilbenes glycosides as dominant metabolites. Larch and Douglas fir wood can be considered abundant sources of flavonoids and flavan-3-ols.
Furthermore, isolation of these metabolites from the plant matrix can be easily achieved. As these plant metabolites have already proven numerous pharmacological and dietary activities, they may find applications in pharmaceutical, cosmetic and food industries. Additionally, LC-ESI-MS has been established as quite a successful method for conifer wood metabolites detection and identification, allowing for simultaneous identification of phytoconstituents from different groups.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/cells11203332/s1, Figures S1–S13: LC-DAD chromatograms of studied wood extracts; Figures S14–S93: ESI-MS/MS and UV spectra of compounds 1–42; Figures S94–S103: 1H NMR spectrum of compounds 9, 12, 16, 20, 22, 23, 27, 29, 30 and 32; Table S1: 1H chemical shifts in ppm for compounds 9, 12, 16, 20, 22, 23, 27, 30 and 32 in methanol-d4 and 29 in chloroform-d (500 MHz, 295 K); Table S2: Canonical SMILES of identified compounds.
Author Contributions
Conceptualization, A.P. and A.K.K.; methodology, A.K.K.; formal analysis, A.P. and M.K.D.; investigation, A.P. and M.K.D.; resources, A.P.; data curation, A.P.; writing—original draft preparation, A.P.; writing—review and editing, A.K.K.; visualization, A.P.; supervision, A.K.K.; project administration, A.P.; funding acquisition, A.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Science Centre, Poland, 2021/41/N/NZ7/00313. For the purpose of Open Access, the authors have applied a CC-BY public copyright license to any Author Accepted Manuscript (AAM) version arising from this submission. The APC was funded by the Medical University of Warsaw.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
The plant material was donated by the Polish Academy of Sciences Botanical Garden, Powsin, Poland, and the University of Warsaw Botanical Garden, Warsaw, Poland. The modernization of the Bruker 500 MHz spectrometer, which was used for NMR measurements carried out in this work, was financed by European Union funds under the Regional Operational Program of the Lodz Voievodship, RPLD.01.01.00–10–0008/18.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Farjon, A. A Handbook of the World’s Conifers; Brill: Leiden, The Netherlands, 2010. [Google Scholar]
- Zajączkowski, G.J.M.; Jabłoński, T.; Szmidla, H.; Kowalska, A.; Małachowska, J.; Piwnicki, J. Raport o stanie lasów w Polsce 2020; Państwowe Gospodarstwo Leśne Lasy Państwowe: Warsaw, Poland, 2021. [Google Scholar]
- Farjon, A.; Filer, D. An Atlas of the World’s Conifers: An Analysis of their Distribution, Biogeography, Diversity and Conservation Status; Brill: Leiden, The Netherlands, 2013. [Google Scholar]
- Rosik-Dulewska, C. Podstawy Gospodarki Odpadami; Wydawnictwo Naukowe PWN: Warsaw, Poland, 2015. [Google Scholar]
- Mantau, U. Wood flow analysis: Quantification of resource potentials, cascades and carbon effects. Biomass Bioenergy 2015, 79, 28–38. [Google Scholar] [CrossRef]
- Hassan, M.K.; Villa, A.; Kuittinen, S.; Jänis, J.; Pappinen, A. An assessment of side-stream generation from Finnish forest industry. J. Mater. Cycles Waste Manag. 2019, 21, 265–280. [Google Scholar] [CrossRef]
- Kumar, R.; Tsvetkov, D.E.; Varshney, V.K.; Nifantiev, N.E. Chemical constituents from temperate and subtropical trees with reference to knotwood. Ind. Crops Prod. 2020, 145, 112077. [Google Scholar] [CrossRef]
- Lunder, M.; Roskar, I.; Hosek, J.; Strukelj, B. Silver Fir (Abies alba) Extracts Inhibit Enzymes Involved in Blood Glucose Management and Protect against Oxidative Stress in High Glucose Environment. Plant. Foods Hum. Nutr 2019, 74, 47–53. [Google Scholar] [CrossRef]
- Huang, X.X.; Bai, M.; Zhou, L.; Lou, L.L.; Liu, Q.B.; Zhang, Y.; Li, L.Z.; Song, S.J. Food Byproducts as a New and Cheap Source of Bioactive Compounds: Lignans with Antioxidant and Anti-inflammatory Properties from Crataegus pinnatifida Seeds. J. Agric. Food Chem. 2015, 63, 7252–7260. [Google Scholar] [CrossRef]
- Sammartino, A.; Tommaselli, G.A.; Gargano, V.; di Carlo, C.; Attianese, W.; Nappi, C. Short-term effects of a combination of isoflavones, lignans and Cimicifuga racemosa on climacteric-related symptoms in postmenopausal women: A double-blind, randomized, placebo-controlled trial. Gynecol. Endocrinol. 2006, 22, 646–650. [Google Scholar] [CrossRef]
- Vo, Q.V.; Nam, P.C.; Bay, M.V.; Thong, N.M.; Cuong, N.D.; Mechler, A. Density functional theory study of the role of benzylic hydrogen atoms in the antioxidant properties of lignans. Sci. Rep. 2018, 8, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Willför, S.; Reunanen, M.; Eklund, P.; Sjöholm, R.; Kronberg, L.; Fardim, P.; Pietarinen, S.; Holmbom, B. Oligolignans in Norway spruce and Scots pine knots and Norway spruce stemwood. Wood Res. Technol. Holzforschung 2004, 58, 345–354. [Google Scholar] [CrossRef]
- Cosentino, M.; Marino, F.; Maio, R.C.; Delle Canne, M.G.; Luzzani, M.; Paracchini, S.; Lecchini, S. Immunomodulatory activity of the lignan 7-hydroxymatairesinol potassium acetate (HMR/lignan) extracted from the heartwood of Norway spruce (Picea abies). Int. Immunopharmacol. 2010, 10, 339–343. [Google Scholar] [CrossRef]
- Smeds, A.I.; Eklund, P.C.; Sjöholm, R.E.; Willför, S.M.; Nishibe, S.; Deyama, T.; Holmbom, B.R. Quantification of a broad spectrum of lignans in cereals, oilseeds, and nuts. J. Agric. Food Chem. 2007, 55, 1337–1346. [Google Scholar] [CrossRef]
- Biasiotto, G.; Zanella, I.; Predolini, F.; Archetti, I.; Cadei, M.; Monti, E.; Luzzani, M.; Pacchetti, B.; Mozzoni, P.; Andreoli, R.; et al. 7-Hydroxymatairesinol improves body weight, fat and sugar metabolism in C57BJ/6 mice on a high-fat diet. Br. J. Nutr. 2018, 120, 751–762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nistor Baldea, L.A.; Martineau, L.C.; Benhaddou-Andaloussi, A.; Arnason, J.T.; Levy, E.; Haddad, P.S. Inhibition of intestinal glucose absorption by anti-diabetic medicinal plants derived from the James Bay Cree traditional pharmacopeia. J. Ethnopharmacol. 2010, 132, 473–482. [Google Scholar] [CrossRef] [PubMed]
- Spoor, D.C.; Martineau, L.C.; Leduc, C.; Benhaddou-Andaloussi, A.; Meddah, B.; Harris, C.; Burt, A.; Fraser, M.H.; Coonishish, J.; Joly, E.; et al. Selected plant species from the Cree pharmacopoeia of northern Quebec possess anti-diabetic potential. Can. J. Physiol. Pharmacol. 2006, 84, 847–858. [Google Scholar] [CrossRef] [PubMed]
- Harbilas, D.; Vallerand, D.; Brault, A.; Saleem, A.; Arnason, J.T.; Musallam, L.; Haddad, P.S. Larix laricina, an Antidiabetic Alternative Treatment from the Cree of Northern Quebec Pharmacopoeia, Decreases Glycemia and Improves Insulin Sensitivity In Vivo. Evid Based Complement. Alternat. Med. 2012, 2012, 296432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shang, N.; Guerrero-Analco, J.A.; Musallam, L.; Saleem, A.; Muhammad, A.; Walshe-Roussel, B.; Cuerrier, A.; Arnason, J.T.; Haddad, P.S. Adipogenic constituents from the bark of Larix laricina du Roi (K. Koch; Pinaceae), an important medicinal plant used traditionally by the Cree of Eeyou Istchee (Quebec, Canada) for the treatment of type 2 diabetes symptoms. J. Ethnopharmacol. 2012, 141, 1051–1057. [Google Scholar] [CrossRef]
- D’Andrea, G. Pycnogenol: A blend of procyanidins with multifaceted therapeutic applications? Fitoterapia 2010, 81, 724–736. [Google Scholar] [CrossRef]
- Shimada, T.; Tokuhara, D.; Tsubata, M.; Kamiya, T.; Kamiya-Sameshima, M.; Nagamine, R.; Takagaki, K.; Sai, Y.; Miyamoto, K.; Aburada, M. Flavangenol (pine bark extract) and its major component procyanidin B1 enhance fatty acid oxidation in fat-loaded models. Eur. J. Pharmacol. 2012, 677, 147–153. [Google Scholar] [CrossRef]
- Poussard, S.; Pires-Alves, A.; Diallo, R.; Dupuy, J.W.; Dargelos, E. A natural antioxidant pine bark extract, Oligopin®, regulates the stress chaperone HSPB1 in human skeletal muscle cells: A proteomics approach. Phytother. Res. 2013, 27, 1529–1535. [Google Scholar] [CrossRef]
- Benković, E.T.; Grohar, T.; Žigon, D.; Švajger, U.; Janeš, D.; Kreft, S.; Štrukelj, B. Chemical composition of the silver fir (Abies alba) bark extract Abigenol® and its antioxidant activity. Ind. Crops Prod. 2014, 52, 23–28. [Google Scholar] [CrossRef]
- Teka, T.; Zhang, L.; Ge, X.; Li, Y.; Han, L.; Yan, X. Stilbenes: Source plants, chemistry, biosynthesis, pharmacology, application and problems related to their clinical Application-A comprehensive review. Phytochemistry 2022, 197, 113128. [Google Scholar] [CrossRef]
- Hsieh, T.-C.; Wu, J.M. Unraveling and Trailblazing Cardioprotection by Resveratrol. In Resveratrol: State-of-the-Art Science and Health Applications; World Scientific: Singapore, 2019; pp. 1–28. [Google Scholar]
- Kim, H.; Seo, K.-H.; Yokoyama, W. Chemistry of Pterostilbene and Its Metabolic Effects. J. Agric. Food Chem. 2020, 68, 12836–12841. [Google Scholar] [CrossRef] [PubMed]
- Cavallini, G.; Straniero, S.; Donati, A.; Bergamini, E. Resveratrol requires red wine polyphenols for optimum antioxidant activity. J. Nutr. Health Aging 2016, 20, 540–545. [Google Scholar] [CrossRef] [PubMed]
- Suprun, A.R.; Dubrovina, A.S.; Aleynova, O.A.; Kiselev, K.V. The Bark of the Spruce Picea jezoensis Is a Rich Source of Stilbenes. Metabolites 2021, 11, 714. [Google Scholar] [CrossRef] [PubMed]
- Hovelstad, H.; Leirset, I.; Oyaas, K.; Fiksdahl, A. Screening Analyses of Pinosylvin Stilbenes, Resin Acids and Lignans in Norwegian Conifers. Molecules 2006, 11, 103–114. [Google Scholar] [CrossRef] [Green Version]
- Strack, D.; Heilemann, J.; Wray, V.; Dirks, H. Structures and accumulation patterns of soluble and insoluble phenolics from norway spruce needles. Phytochemistry 1989, 28, 2071–2078. [Google Scholar] [CrossRef]
- Yeap Foo, L.; Karchesy, J.J. Procyanidin dimers and trimers from Douglas fir inner bark. Phytochemistry 1989, 28, 1743–1747. [Google Scholar] [CrossRef]
- Sut, S.; Baldan, V.; Faggian, M.; Ferrarese, I.; Maccari, E.; Teobaldo, E.; De Zordi, N.; Bertoni, P.; Peron, G.; Dall’Acqua, S. The Bark of Picea abies L., a Waste from Sawmill, as a Source of Valuable Compounds: Phytochemical Investigations and Isolation of a Novel Pimarane and a Stilbene Derivative. Plants 2021, 10, 2106. [Google Scholar] [CrossRef]
- Pietarinen, S.P.; Willför, S.M.; Ahotupa, M.O.; Hemming, J.E.; Holmbom, B.R. Knotwood and bark extracts: Strong antioxidants from waste materials. J. Wood Sci. 2006, 52, 436–444. [Google Scholar] [CrossRef]
- Willför, S.; Nisula, L.; Hemming, J.; Reunanen, M.; Holmbom, B. Bioactive phenolic substances in industrially important tree species. Part 2: Knots and stemwood of fir species. Holzforschung 2004, 58, 650–659. [Google Scholar] [CrossRef]
- Dellus, V.; Mila, I.; Scalbert, A.; Menard, C.; Michon, V.; Herve du Penhoat, C.L.M. Douglas-fir polyphenols and heartwood formation. Phytochemistry 1997, 45, 1573–1578. [Google Scholar] [CrossRef]
- Ivanova, S.Z.; Babkin, V.A. Polyphenolic compounds from Larix gmelinii phloem. Chem. Nat. Compd. 2011, 47, 124–125. [Google Scholar] [CrossRef]
- Wu, L.; Li, Y.-L.; Li, S.-M.; Yang, X.-W.; Xia, J.-H.; Zhou, L.; Zhang, W.-D. Systematic Phytochemical Investigation of Abies spectabilis. Chem. Pharm. Bull. 2010, 58, 1646–1649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yashunsky, D.V.; Men’shov, V.M.; Tsvetkov, D.E.; Tsvetkov, Y.E.; Bel’ko, A.A.; Vasiyarov, G.G.; Titova, E.V.; Pimenov, A.V.; Onuchin, A.A.; Dokichev, V.A.; et al. Analysis of content of (–)-secoisolariciresinol and related polyphenols in different morphological parts and anatomical structures of larch wood from Siberia. Russ. Chem. Bull. 2014, 63, 2571–2576. [Google Scholar] [CrossRef]
- Willför, S.; Hemming, J.; Reunanen, M.; Holmbom, B. Phenolic and Lipophilic Extractives in Scots Pine Knots and Stemwood. Wood Res. Technol. Holzforschung 2003, 57, 359–372. [Google Scholar] [CrossRef]
- Fang, J.-M.; Su, W.-C.; Cheng, Y.-S. Flavonoids and stilbenes from armand pine. Phytochemistry 1988, 27, 1395–1397. [Google Scholar] [CrossRef]
- Jaiswal, R.; Jayasinghe, L.; Kuhnert, N. Identification and characterization of proanthocyanidins of 16 members of the Rhododendron genus (Ericaceae) by tandem LC-MS. J. Mass Spectrom. 2012, 47, 502–515. [Google Scholar] [CrossRef]
- Shen, Z.; Haslam, E.; Falshaw, C.P.; Begley, M.J. Procyanidins and polyphenols of Larix gmelini bark. Phytochemistry 1986, 25, 2629–2635. [Google Scholar] [CrossRef]
- Mbakidi-Ngouaby, H.; Pinault, E.; Gloaguen, V.; Costa, G.; Sol, V.; Millot, M.; Mambu, L. Profiling and seasonal variation of chemical constituents from Pseudotsuga menziesii wood. Industrial Crops Prod. 2018, 117, 34–49. [Google Scholar] [CrossRef]
- Razgonova, M.; Zakharenko, A.; Pikula, K.; Manakov, Y.; Ercisli, S.; Derbush, I.; Kislin, E.; Seryodkin, I.; Sabitov, A.; Kalenik, T.; et al. LC-MS/MS Screening of Phenolic Compounds in Wild and Cultivated Grapes Vitis amurensis Rupr. Molecules 2021, 26, 3650. [Google Scholar] [CrossRef]
- Abdelghffar, E.A.; Obaid, W.A.; Alamoudi, M.O.; Mohammedsaleh, Z.M.; Annaz, H.; Abdelfattah, M.A.O.; Sobeh, M. Thymus fontanesii attenuates CCl4-induced oxidative stress and inflammation in mild liver fibrosis. Biomed. Pharmacother. 2022, 148, 112738. [Google Scholar] [CrossRef]
- Tsimogiannis, D.; Samiotaki, M.; Panayotou, G.; Oreopoulou, V. Characterization of Flavonoid Subgroups and Hydroxy Substitution by HPLC-MS/MS. Molecules 2007, 12, 593–606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bibi Sadeer, N.; Sinan, K.I.; Cziáky, Z.; Jekő, J.; Zengin, G.; Jeewon, R.; Abdallah, H.H.; AlDhaheri, Y.; Eid, A.H.; Mahomoodally, M.F. Towards the Pharmacological Validation and Phytochemical Profiling of the Decoction and Maceration of Bruguiera gymnorhiza (L.) Lam.—A Traditionally Used Medicinal Halophyte. Molecules 2022, 27, 2000. [Google Scholar] [CrossRef] [PubMed]
- Koulis, G.A.; Tsagkaris, A.S.; Aalizadeh, R.; Dasenaki, M.E.; Panagopoulou, E.I.; Drivelos, S.; Halagarda, M.; Georgiou, C.A.; Proestos, C.; Thomaidis, N.S. Honey Phenolic Compound Profiling and Authenticity Assessment Using HRMS Targeted and Untargeted Metabolomics. Molecules 2021, 26, 2769. [Google Scholar] [CrossRef] [PubMed]
- Parisi, V.; Vassallo, A.; Pisano, C.; Signorino, G.; Cardile, F.; Sorrentino, M.; Colelli, F.; Fucci, A.; D’Andrea, E.L.; De Tommasi, N.; et al. A Herbal Mixture from Propolis, Pomegranate, and Grape Pomace Endowed with Anti-Inflammatory Activity in an In Vivo Rheumatoid Arthritis Model. Molecules 2020, 25, 2255. [Google Scholar] [CrossRef]
- Wu, H.; Cao, Y.; Qu, Y.; Li, T.; Wang, J.; Yang, Y.; Zhang, C.; Sun, Y. Integrating UPLC-QE-Orbitrap-MS technology and network pharmacological method to reveal the mechanism of Bailemian capsule to relieve insomnia. Nat. Prod. Res. 2022, 36, 2554–2558. [Google Scholar] [CrossRef]
- Mena, P.; Calani, L.; Dall’Asta, C.; Galaverna, G.; García-Viguera, C.; Bruni, R.; Crozier, A.; Del Rio, D. Rapid and Comprehensive Evaluation of (Poly)phenolic Compounds in Pomegranate (Punica granatum L.) Juice by UHPLC-MSn. Molecules 2012, 17, 14821–14840. [Google Scholar] [CrossRef] [Green Version]
- Mercolini, L.; Protti, M.; Saracino, M.A.; Mandrone, M.; Antognoni, F.; Poli, F. Analytical Profiling of Bioactive Phenolic Compounds in Argan (Argania spinosa) Leaves by Combined Microextraction by Packed Sorbent (MEPS) and LC-DAD-MS/MS. Phytochem. Anal. 2016, 27, 41–49. [Google Scholar] [CrossRef]
- Gabaston, J.; Richard, T.; Cluzet, S.; Palos Pinto, A.; Dufour, M.-C.; Corio-Costet, M.-F.; Mérillon, J.-M. Pinus pinaster Knot: A Source of Polyphenols against Plasmopara viticola. J. Agric. Food Chem. 2017, 65, 8884–8891. [Google Scholar] [CrossRef]
- Pompermaier, L.; Heiss, E.H.; Alilou, M.; Mayr, F.; Monizi, M.; Lautenschlaeger, T.; Schuster, D.; Schwaiger, S.; Stuppner, H. Dihydrochalcone Glucosides from the Subaerial Parts of Thonningia sanguinea and Their in Vitro PTP1B Inhibitory Activities. J. Nat. Prod. 2018, 81, 2091–2100. [Google Scholar] [CrossRef]
- Ahmed, A.; Li, W.; Zhang, J.S.; Sam, P.H.; Zou, Y.H.; Tang, G.H.; Yin, S. A new bisabolane sesquiterpenoid and a new abietane diterpenoid from Cephalotaxus sinensis. Nat. Prod. Res. 2018, 32, 175–181. [Google Scholar] [CrossRef]
- Sefkow, M. Enantioselective Synthesis of (−)-Wikstromol Using a New Approach via Malic Acid. J. Org. Chem. 2001, 66, 2343–2349. [Google Scholar] [CrossRef] [PubMed]
- Eklund, P.C.; Backman, M.J.; Kronberg, L.A.; Smeds, A.I.; Sjoholm, R.E. Identification of lignans by liquid chromatography-electrospray ionization ion-trap mass spectrometry. J. Mass Spectrom. 2008, 43, 97–107. [Google Scholar] [CrossRef] [PubMed]
- Voronin, K.S.; Fenin, A.A.; Zhevlakova, A.K.; Zavadskii, S.P.; Selivanova, I.A. Polyphenolic Profile of Larch Knotwood. Pharm. Chem. J. 2021, 55, 781–786. [Google Scholar] [CrossRef]
- Stella, L.; De Rosso, M.; Panighel, A.; Vedova, A.D.; Flamini, R.; Traldi, P. Collisionally induced fragmentation of [M-H](-) species of resveratrol and piceatannol investigated by deuterium labelling and accurate mass measurements. Rapid Commun. Mass Spectrom. 2008, 22, 3867–3872. [Google Scholar] [CrossRef]
- Flamini, R.; De Rosso, M.; De Marchi, F.; Dalla Vedova, A.; Panighel, A.; Gardiman, M.; Maoz, I.; Bavaresco, L. An innovative approach to grape metabolomics: Stilbene profiling by suspect screening analysis. Metabolomics 2013, 9, 1243–1253. [Google Scholar] [CrossRef]
- Buiarelli, F.; Coccioli, F.; Jasionowska, R.; Merolle, M.; Terracciano, A. Analysis of some stilbenes in Italian wines by liquid chromatography/tandem mass spectrometry. Rapid Commun. Mass Spectrom. 2007, 21, 2955–2964. [Google Scholar] [CrossRef]
- Yeo, S.C.M.; Luo, W.; Wu, J.; Ho, P.C.; Lin, H.-S. Quantification of pinosylvin in rat plasma by liquid chromatography–tandem mass spectrometry: Application to a pre-clinical pharmacokinetic study. J. Chromatogr. B 2013, 931, 68–74. [Google Scholar] [CrossRef]
- Manville, J.F. Juvabione and its Analogs. Juvabione and Δ4′-Dehydrojuvabione Isolated from the Whole Wood of Abiesbalsamea, have the R,R Stereoconfigurations, not the R,S. Can. J. Chem. 1975, 53, 1579–1585. [Google Scholar] [CrossRef] [Green Version]
- Brunswick, P.; Blajkevitch, O.; Chow, L.; MacInnis, C.; van Aggelen, G.; Kim, M.; Shang, D. Trace analysis of resin acids in surface waters by direct injection liquid chromatography time of flight mass spectrometry and triple quadrupole mass spectrometry. J. Chromatogr. A 2021, 1656, 462558. [Google Scholar] [CrossRef]
- Schymanski, E.L.; Ruttkies, C.; Krauss, M.; Brouard, C.; Kind, T.; Dührkop, K.; Allen, F.; Vaniya, A.; Verdegem, D.; Böcker, S.; et al. Critical Assessment of Small Molecule Identification 2016: Automated methods. J. Cheminformatics 2017, 9, 22. [Google Scholar] [CrossRef]
- Willför, S.; Nisula, L.; Hemming, J.; Reunanen, M.; Holmbom, B. Bioactive phenolic substances in industrially important tree species. Part 1: Knots and stemwood of different spruce species. Wood Res. Technol. Holzforschung 2004, 58, 335–344. [Google Scholar] [CrossRef]
- Willför, S.; Hemming, J.; Reunanen, M.; Eckerman, C.; Holmbom, B. Lignans and Lipophilic Extractives in Norway Spruce Knots and Stemwood. Wood Res. Technol. Holzforschung 2003, 57, 27–36. [Google Scholar] [CrossRef]
- Rodriguez-Garcia, C.; Sanchez-Quesada, C.; Toledo, E.; Delgado-Rodriguez, M.; Gaforio, J.J. Naturally Lignan-Rich Foods: A Dietary Tool for Health Promotion? Molecules 2019, 24, 917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holmbom, B.; Eckerman, C.; Eklund, P.; Hemming, J.; Nisula, L.; Reunanen, M.; Sjöholm, R.; Sundberg, A.; Sundberg, K.; Willför, S. Knots in trees–A new rich source of lignans. Phytochem. Rev. 2003, 2, 331–340. [Google Scholar] [CrossRef]
- Kiselev, K.V.; Grigorchuk, V.P.; Ogneva, Z.V.; Suprun, A.R.; Dubrovina, A.S. Stilbene biosynthesis in the needles of spruce Picea jezoensis. Phytochemistry 2016, 131, 57–67. [Google Scholar] [CrossRef]

{kind=link}
Table 1.
Retention time, UV, and ESI-MS/MS data of the compounds identified in Pinaceae species wood using LC-DAD–ESI-MS/MS analysis.
Table 1.
Retention time, UV, and ESI-MS/MS data of the compounds identified in Pinaceae species wood using LC-DAD–ESI-MS/MS analysis.
| Compounds | Retention Time [min] | UVmax [nm] | ESI 1 | MS1 [m/z] | MS2 [m/z] | Group | Presence in Pinaceae spp. |
|---|---|---|---|---|---|---|---|
| 1.gallocatechin | 3.0 | 199, 277 | [M−H]− | 305 | 219, 179, 137 | flavan-3-ol | [30] |
| 2.dimeric procyanidin B (I) | 3.3 | 230, 282 | [M−H]− | 577 | 559, 451, 407, 289 | flavan-3-ol | [31] |
| 3.dimeric procyanidin B (II) | 3.6 | 207, 238, 280 | [M−H]− | 577 | 559, 451, 407, 289 | flavan-3-ol | [31] |
| 4.trimeric procyanidin B | 4.2 | 199, 281 | [M−H]− | 865 | 695, 577, 407, 289 | flavan-3-ol | [32] |
| 5.catechin 2 | 4.5 | 232, 278 | [M−H]− | 289 | 245, 203, 179 | flavan-3-ol | [33] |
| 6.dimeric procyanidin B (III) | 5.2 | 238, 283 | [M−H]− | 577 | 559, 451, 407, 289 | flavan-3-ol | [31] |
| 7.epi-catechin 2 | 6.6 | 279 | [M−H]− | 289 | 245, 203, 179 | flavan-3-ol | [32] |
| 8.dihydromyricetin | 7.7 | 213, 225, 290 | [M−H]− | 319 | 301, 193, 125 | flavonoid | [33] |
| 9.7-hydroxylariciresinol (I) 3 | 7.8 | 199, 225, 279 | [M−H]− | 375 | 357, 345, 327, 297 | lignan | [34] |
| 10.trans-astringin | 8.1 | 192, 218, 324 | [M−H]− | 405 | 243, 225, 201, 173, 159 | stilbene | [33] |
| 11.dihydrokaempferol | 8.4 | 290 | [M−H]− | 287 | 259, 243, 181 | flavonoid | [34] |
| 12.todolactol 2 | 9.9 | 196, 225, 280 | [M−H]− | 375 | 327, 191, 176 | lignan | [33] |
| 13.taxifolin glucoside | 10.2 | 196, 287 | [M−H]− | 465 | 447, 437, 303, 285, 259 | flavonoid | [35] |
| 14.cis-astringin | 10.8 | 213, 319 | [M−H]− | 405 | 243, 225, 201, 173, 159 | stilbene | [28] |
| 15.dimeric procyanidin B (IV) | 11.6 | 243, 280 | [M−H]− | 577 | 559, 451, 407, 289 | flavan-3-ol | [31] |
| 16.7-hydroxylariciresinol (II) 3 | 11.6 | 198, 227, 279 | [M−H+HCOOH]− | 421 | 375, 357, 345, 325 | lignan | [34] |
| 17.piceid | 11.7 | 216, 319 | [M−H]− | 389 | 227, 185, 183, 157, 143 | stilbene | [33] |
| 18.taxifolin 2 | 12.4 | 280 | [M−H]− | 303 | 285, 177 | flavonoid | [34] |
| 19.isorhapontin | 12.5 | 192, 218, 325 | [M−H+HCOOH]− | 465 | 419, 257, 242, 241 | stilbene | [33] |
| 20.cyclolariciresinol 3 | 12.7 | 197, 283 | [M−H]− | 359 | 344, 313, 189 | lignan | [34] |
| 21.α-conidendric acid | 14.0 | 200, 225, 280 | [M−H+HCOOH]− | 419 | 373, 177 | lignan | [34] |
| 22.7-hydroxymatairesinol 3 | 15.0 | 197, 226, 280 | [M−H]− | 373 | 355, 340, 311, 293, 219 | lignan | [34] |
| 23.Secoisolariciresinol 3 | 15.3 | 232, 281 | [M−H]− | 361 | 346, 313, 299 | lignan | [34] |
| 24.eriodictyol | 15.4 | 194, 287 | [M−H]− | 287 | 259, 243, 151 | flavonoid | [36] |
| 25.myricetin 2 | 15.7 | 254, 373 | [M−H]− | 317 | 289, 179, 151, 137 | flavonoid | [37] |
| 26.secoisolariciresinol guaiacylglyceryl ether | 15.7 | 197, 280 | [M−H]− | 557 | 539, 525, 521, 509, 415, 361 | sesquilignan | [34] |
| 27.lariciresinol 3 | 16.1 | 197, 200, 202, 280 | [M−H+HCOOH]− | 405 | 359, 329 | lignan | [34] |
| 28.lariciresinol guaiacylglyceryl ether | 16.7 | 201, 225, 280 | [M−H]− | 555 | 525, 507, 359, 329, 315, 195, 165 | sesquilignan | [34] |
| 29.nortrachelogenin 3 | 18.3 | 199, 225, 280 | [M−H]− | 373 | 355, 327, 311, 249, 223, 191, 147 | lignan | [34] |
| 30.pinoresinol 3 | 19.1 | 201, 280 | [M−H]− | 357 | 342, 327, 311, 151, 136 | lignan | [34] |
| 31.quercetin 2 | 19.8 | 208, 368 | [M−H]− | 301 | 273, 179, 151 | flavonoid | [38] |
| 32.matairesinol 3 | 21.6 | 197, 281 | [M−H]− | 357 | 342, 313, 298, 281, 209, 191, 147 | lignan | [34] |
| 33.pinobanksin | 22.0 | 214, 284 | [M−H]− | 271 | 253 | flavonoid | [33] |
| 34.pinosylvin | 26.3 | 223, 299 | [M−H]− | 211 | 169 | stilbene | [39] |
| 35.pinocembrin | 30.3 | 214, 288 | [M−H]− | 255 | 213, 211 | flavonoid | [39] |
| 36.pinobanksin 3-O-acetate | 30.5 | 216, 292 | [M−H]− | 313 | 271, 253 | flavonoid | [40] |
| 37.pinosylvin monomethyl ether | 33.6 | 212, 223, 300 | [M−H]− | 225 | 210 | stilbene | [39] |
| 38.dehydrojuvabione | 41.0 | 224, 300 | [M+H]+ | 265 | 251, 233, 205, 187, 176 | sesquiterpene | [34] |
| 39.neoabietic acid | 52.0 | 251 | [M+H]+ | 303 | 257, 219, 179, 151, 123 | diterpene | [39] |
| 40.abietic acid 2 | 52.3 | 241 | [M+H]+ | 303 | 285, 257, 123 | diterpene | [39] |
1 Ionization mode (positive or negative); [M−H]−—deprotonated molecule; [M+H]+—protonated molecule; [M−H+HCOOH]−—formate adduct ion. 2 Identified by comparison with reference compounds. 3 Identified by comparison with isolated compound characterized by NMR.
Table 2.
Presence of compounds confirmed through LC-DAD–ESI-MS/MS analysis in Pinaceae species.
| Compounds | Abies alba | Pinus sylvestris | Pinus mugo | Pinus cembra | Pinus strobus | Pinus ×rhaetica | Larix decidua | Larix polonica | Larix kaempferi | Pseudotsuga menziesii | Tsuga canadensis | Picea abies | Picea glauca |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1.gallocatechin | + | + | + | + | + | + | + | + | + | − | − | − | − |
| 2.dimeric procyanidin B (I) | + | + | + | + | + | + | + | + | + | + | + | + | + |
| 3.dimeric procyanidin B (II) | + | + | + | + | + | + | + | + | + | + | + | + | + |
| 4.trimeric procyanidin B | + | + | + | + | + | + | + | + | + | + | + | + | + |
| 5.catechin | + | + | + | + | + | + | + | + | + | + | + | + | + |
| 6.dimeric procyanidin B (III) | − | − | − | − | − | − | + | + | + | + | − | + | − |
| 7.epi-catechin | + | − | − | + | + | − | + | + | + | + | + | − | − |
| 8.dihydromyricetin | − | − | + | − | − | + | + | + | − | − | − | − | − |
| 9.7-hydroxylariciresinol (I) | + | − | − | − | − | − | − | − | − | − | − | + | + |
| 10.trans-astringin | − | − | − | − | − | − | − | − | − | − | − | + | + |
| 11.dihydrokaempferol | − | − | − | − | − | − | + | + | − | − | − | − | − |
| 12.todolactol | + | − | − | − | − | − | + | − | + | + | + | + | + |
| 13.taxifolin hexoside | − | + | + | + | + | + | − | + | + | + | − | + | − |
| 14.cis-astringin | − | − | − | − | − | − | + | + | + | − | − | + | + |
| 15.dimeric procyanidin B (IV) | − | − | − | − | − | − | + | + | + | − | − | − | − |
| 16.7-hydroxylariciresinol (II) | + | − | + | − | + | − | + | − | + | + | + | − | − |
| 17.piceid | − | − | − | − | − | − | − | − | − | − | − | + | + |
| 18.taxifolin | − | + | + | + | + | + | + | + | + | + | − | + | + |
| 19.isorhapontin | − | − | − | − | − | − | − | − | − | − | − | + | + |
| 20.cyclolariciresinol | + | − | − | − | − | − | − | − | − | − | − | − | − |
| 21.α-conidendric acid | + | − | + | − | + | − | + | + | + | − | + | − | − |
| 22.7-hydroxymatairesinol | + | − | + | − | + | − | + | + | + | + | + | + | + |
| 23.secoisolariciresinol | + | − | + | − | + | − | + | + | + | + | + | + | + |
| 24.eriodictyol | − | − | + | + | + | − | + | + | + | + | − | − | − |
| 25.myricetin | − | − | + | − | − | − | + | + | + | + | − | − | − |
| 26.secoisolariciresinol guaiacylglyceryl ether | + | − | + | − | − | − | + | + | + | + | − | − | − |
| 27.lariciresinol | + | − | − | − | + | − | + | + | + | + | + | − | + |
| 28.lariciresinol guaiacylglyceryl ether | + | − | − | − | − | − | − | − | − | − | − | − | − |
| 29.nortrachelogenin | + | + | + | − | + | − | + | + | + | + | + | + | + |
| 30.pinoresinol | − | + | − | − | − | − | − | − | − | − | − | − | − |
| 31.quercetin | − | − | − | − | + | − | + | + | + | + | − | − | − |
| 32.matairesinol | − | + | + | + | + | + | + | + | + | − | + | + | + |
| 33.pinobanksin | − | − | + | + | + | − | + | + | + | + | − | − | + |
| 34.pinosylvin | − | + | + | − | + | − | − | − | − | − | − | − | − |
| 35.pinocembrin | − | + | + | − | + | + | − | − | − | + | − | − | − |
| 36.pinobanksin 3-O-acetate | − | − | − | − | + | − | − | − | − | − | − | − | − |
| 37.pinosylvin monomethyl ether | − | + | + | + | + | − | − | − | − | − | − | − | − |
| 38.dehydrojuvabione | + | − | − | − | − | − | − | − | − | − | − | − | − |
| 39.neoabietic acid | + | + | + | + | + | + | + | + | + | + | + | + | + |
| 40.abietic acid | + | + | + | + | + | + | + | + | + | + | + | + | + |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Patyra, A.; Dudek, M.K.; Kiss, A.K. LC-DAD–ESI-MS/MS and NMR Analysis of Conifer Wood Specialized Metabolites. Cells 2022, 11, 3332. https://doi.org/10.3390/cells11203332
AMA Style
Patyra A, Dudek MK, Kiss AK. LC-DAD–ESI-MS/MS and NMR Analysis of Conifer Wood Specialized Metabolites. Cells. 2022; 11(20):3332. https://doi.org/10.3390/cells11203332
Chicago/Turabian StylePatyra, Andrzej, Marta Katarzyna Dudek, and Anna Karolina Kiss. 2022. "LC-DAD–ESI-MS/MS and NMR Analysis of Conifer Wood Specialized Metabolites" Cells 11, no. 20: 3332. https://doi.org/10.3390/cells11203332
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.