Non-Platinum Metal Complexes as Potential Anti-Triple Negative Breast Cancer Agents


Abstract
:1. Introduction
2. Anticancer Effects of Non-Platinum MBC (NPMBC)
2.1. Suppression of Cancer Cell Viability in Association with the Induction of Cell Death
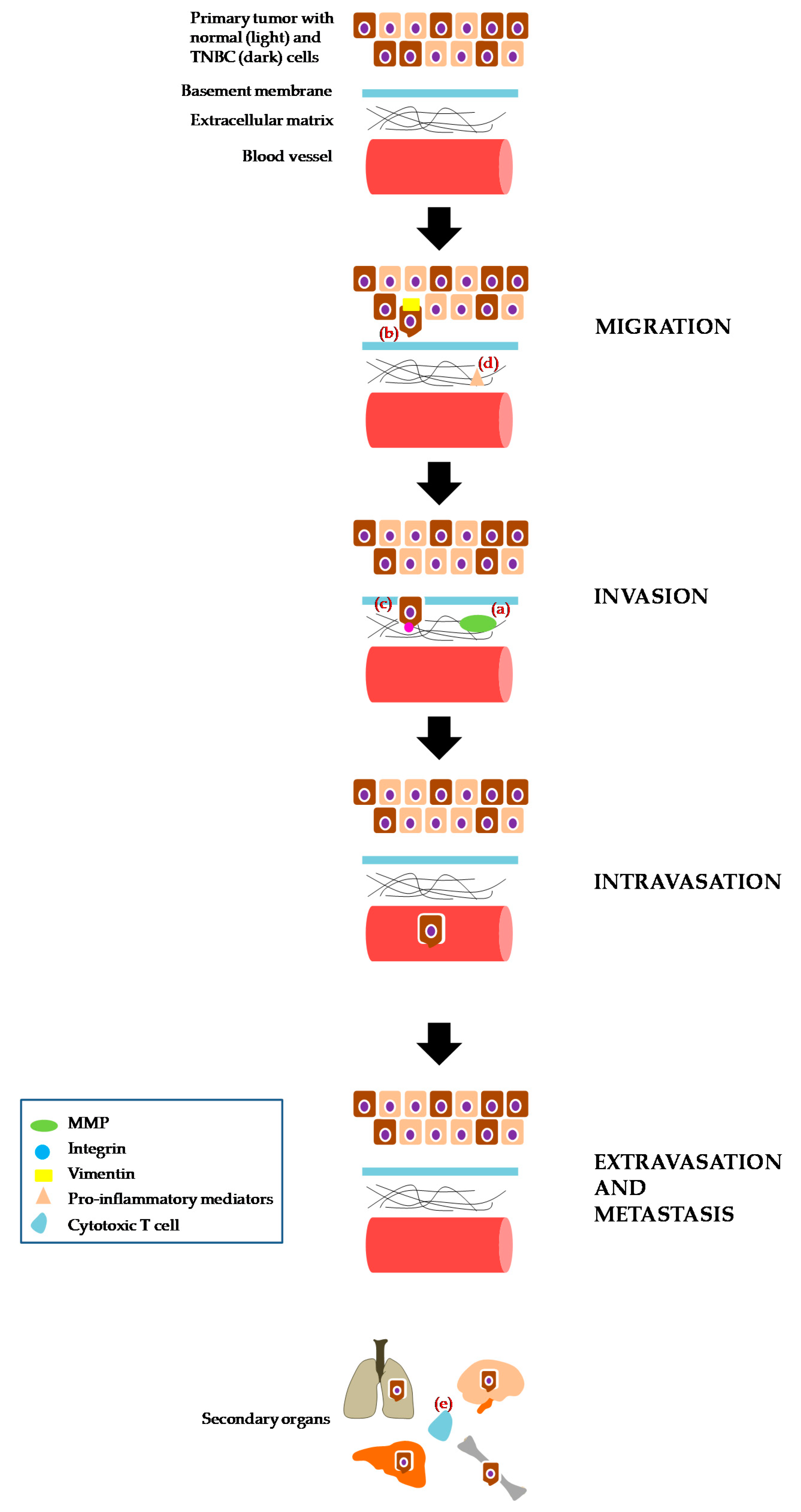
2.2. Migration, Invasion and/or Metastasis
3. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- World Health Organization. Breast Cancer. Available online: http://www.who.int/cancer/prevention/diagnosis-screening/breast-cancer/en/ (accessed on 27 July 2018).
- Howell, A.; Anderson, A.S.; Clarke, R.B.; Duffy, S.W.; Evans, D.G.; Garcia-Closas, M.; Gescher, A.J.; Key, T.J.; Saxton, J.M.; Harvie, M.N. Risk Determination and Prevention of Breast Cancer. Breast Cancer Res. 2014, 16, 446. [Google Scholar] [CrossRef] [PubMed]
- Ricks-Santi, L.; McDonald, J.T.; Gold, B.; Dean, M.; Thompson, N.; Abbas, M.; Wilson, B.; Kanaan, Y.; Naab, T.J.; Dunston, G. Next Generation Sequencing Reveals High Prevalence of BRCA1 and BRCA2 Variants of Unknown Significance in Early-Onset Breast Cancer in African American Women. Ethn. Dis. 2017, 27, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Cheng, H.; Bai, Z.; Li, J. Breast Cancer Cell Line Classification and Its Relevance with Breast Tumor Subtyping. J. Cancer 2017, 8, 3131–3141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaha, D.C. Significance of Immunohistochemistry in Breast Cancer. World J. Clin. Oncol. 2014, 5, 382–392. [Google Scholar] [CrossRef] [PubMed]
- Aysola, K.; Desai, A.; Welch, C.; Xu, J.; Qin, Y.; Reddy, V.; Matthews, R.; Owens, C.; Okoli, J.; Beech, D.J.; et al. Triple Negative Breast Cancer—An Overview. Hered. Genet. 2013, 2013. [Google Scholar] [CrossRef]
- Sharma, P. Update on the Treatment of Early-Stage Triple-Negative Breast Cancer. Curr. Treat. Options Oncol. 2018, 19, 22. [Google Scholar] [CrossRef] [PubMed]
- Miller, E.; Lee, H.J.; Lulla, A.; Hernandez, L.; Gokare, P.; Lim, B. Current Treatment of Early Breast Cancer: Adjuvant and Neoadjuvant Therapy. F1000Res 2014, 3, 198. [Google Scholar] [CrossRef] [PubMed]
- Hongthong, K.; Ratanaphan, A. BRCA1-Associated Triple-Negative Breast Cancer and Potential Treatment for Ruthenium-Based Compounds. Curr. Cancer Drug Targets 2016, 16, 606–617. [Google Scholar] [CrossRef] [PubMed]
- Chavez, K.J.; Garimella, S.V.; Lipkowitz, S. Triple Negative Breast Cancer Cell Lines: One Tool in the Search for Better Treatment of Triple Negative Breast Cancer. Breast Dis. 2010, 32, 35–48. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, B.D.; Bauer, J.A.; Chen, X.; Sanders, M.E.; Chakravarthy, A.B.; Shyr, Y.; Pietenpol, J.A. Identification of Human Triple-Negative Breast Cancer Subtypes and Preclinical Models for Selection of Targeted Therapies. J. Clin. Investig. 2011, 121, 2750–2767. [Google Scholar] [CrossRef] [PubMed]
- Neve, R.M.; Chin, K.; Fridlyand, J.; Yeh, J.; Baehner, F.L.; Fevr, T.; Clark, L.; Bayani, N.; Coppe, J.P.; Tong, F.; et al. A Collection of Breast Cancer Cell Lines for the Study of Functionally Distinct Cancer Subtypes. Cancer Cell 2006, 10, 515–527. [Google Scholar] [CrossRef] [PubMed]
- Jiang, G.; Zhang, S.; Yazdanparast, A.; Li, M.; Pawar, A.V.; Liu, Y.; Inavolu, S.M.; Cheng, L. Comprehensive Comparison of Molecular Portraits Between Cell Lines and Tumors in Breast Cancer. BMC Genomics 2016, 17, 525. [Google Scholar] [CrossRef] [PubMed]
- Webb, J.A.; Ou, Y.C.; Faley, S.; Paul, E.P.; Hittinger, J.P.; Cutright, C.C.; Lin, E.C.; Bellan, L.M.; Bardhan, R. Theranostic Gold Nanoantennas for Simultaneous Multiplexed Raman Imaging of Immunomarkers and Photothermal Therapy. ACS Omega 2017, 2, 3583–3594. [Google Scholar] [CrossRef] [PubMed]
- Tao, Y.; Li, M.; Auguste, D.T. Pattern-Based Sensing of Triple Negative Breast Cancer Cells with Dual-Ligand Cofunctionalized Gold Nanoclusters. Biomaterials 2017, 116, 21–33. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Kim, H.C.; Wolfram, J.; Mu, C.; Zhang, W.; Liu, H.; Xie, Y.; Mai, J.; Zhang, H.; Li, Z.; et al. A Liposome Encapsulated Ruthenium Polypyridine Complex as a Theranostic Platform for Triple-Negative Breast Cancer. Nano Lett. 2017, 17, 2913–2920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Pang, B.; Luehmann, H.; Detering, L.; Yang, X.; Sultan, D.; Harpstrite, S.; Sharma, V.; Cutler, C.S.; Xia, Y.; et al. Gold Nanoparticles Doped with (199) Au Atoms and Their Use for Targeted Cancer Imaging by SPECT. Adv. Healthc. Mater. 2016, 5, 928–935. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Kim, H.S.; Jin, T.; Yi, A.; Moon, W.K. Ultrasound-Guided Photoacoustic Imaging for the Selective Detection of EGFR-Expressing Breast Cancer and Lymph Node Metastases. Biomed. Opt. Express 2016, 7, 1920–1931. [Google Scholar] [CrossRef] [PubMed]
- Abulrob, A.; Corluka, S.; Blasiak, B.; Gino Fallone, B.; Ponjevic, D.; Matyas, J.; Tomanek, B. LyP-1 Conjugated Nanoparticles for Magnetic Resonance Imaging of Triple Negative Breast Cancer. Mol. Imaging Biol. 2018, 20, 428–435. [Google Scholar] [CrossRef] [PubMed]
- Guo, P.; Huang, J.; Wang, L.; Jia, D.; Yang, J.; Dillon, D.A.; Zurakowski, D.; Mao, H.; Moses, M.A.; Auguste, D.T. ICAM-1 as a Molecular Target for Triple Negative Breast Cancer. Proc. Natl. Acad. Sci. USA 2014, 111, 14710–14715. [Google Scholar] [CrossRef] [PubMed]
- Pang, B.; Zhao, Y.; Luehmann, H.; Yang, X.; Detering, L.; You, M.; Zhang, C.; Zhang, L.; Li, Z.Y.; Ren, Q.; et al. 64Cu-Doped PdCu@Au Tripods: A Multifunctional Nanomaterial for Positron Emission Tomography and Image-Guided Photothermal Cancer Treatment. ACS Nano 2016, 10, 3121–3131. [Google Scholar] [CrossRef] [PubMed]
- Ndagi, U.; Mhlongo, N.; Soliman, M.E. Metal Complexes in Cancer Therapy—An Update from Drug Design Perspective. Drug Des. Dev. Ther. 2017, 11, 599–616. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Lippard, S.J. Cellular Processing of Platinum Anticancer Drugs. Nat. Rev. Drug Discov. 2005, 4, 307–320. [Google Scholar] [CrossRef] [PubMed]
- Birkbak, N.J.; Wang, Z.C.; Kim, J.Y.; Eklund, A.C.; Li, Q.; Tian, R.; Bowman-Colin, C.; Li, Y.; Greene-Colozzi, A.; Iglehart, J.D.; et al. Telomeric Allelic Imbalance Indicates Defective DNA Repair and Sensitivity to DNA-Damaging Agents. Cancer Discov. 2012, 2, 366–375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silver, D.P.; Richardson, A.L.; Eklund, A.C.; Wang, Z.C.; Szallasi, Z.; Li, Q.; Juul, N.; Leong, C.O.; Calogrias, D.; Buraimoh, A.; et al. Efficacy of Neoadjuvant Cisplatin in Triple-Negative Breast Cancer. J. Clin. Oncol. 2010, 28, 1145–1153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciarimboli, G. Membrane Transporters as Mediators of Cisplatin Side-Effects. Anticancer Res. 2014, 34, 547–550. [Google Scholar] [CrossRef] [PubMed]
- Dam, D.H.; Culver, K.S.; Odom, T.W. Grafting Aptamers onto Gold Nanostars Increases in vitro Efficacy in a Wide Range of Cancer Cell Types. Mol. Pharm. 2014, 11, 580–587. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Tian, Y.; Tian, W.; Sun, J.; Zhao, S.; Liu, Y.; Wang, C.; Tang, Y.; Ma, X.; Teng, Z.; et al. Selectively Sensitizing Malignant Cells to Photothermal Therapy Using a CD44-Targeting Heat Shock Protein 72 Depletion Nanosystem. ACS Nano 2016, 10, 8578–8590. [Google Scholar] [CrossRef] [PubMed]
- Ou, Y.C.; Webb, J.A.; Faley, S.; Shae, D.; Talbert, E.M.; Lin, S.; Cutright, C.C.; Wilson, J.T.; Bellan, L.M.; Bardhan, R. Gold Nanoantenna-Mediated Photothermal Drug Delivery from Thermosensitive Liposomes in Breast Cancer. ACS Omega 2016, 1, 234–243. [Google Scholar] [CrossRef] [PubMed]
- Santiago, T.; DeVaux, R.S.; Kurzatkowska, K.; Espinal, R.; Herschkowitz, J.I.; Hepel, M. Surface-Enhanced Raman Scattering Investigation of Targeted Delivery and Controlled Release of Gemcitabine. Int. J. Nanomed. 2017, 12, 7763–7776. [Google Scholar] [CrossRef] [PubMed]
- Saw, W.S.; Ujihara, M.; Chong, W.Y.; Voon, S.H.; Imae, T.; Kiew, L.V.; Lee, H.B.; Sim, K.S.; Chung, L.Y. Size-Dependent Effect of Cystine/Citric Acid-Capped Confeito-Like Gold Nanoparticles on Cellular Uptake and Photothermal Cancer Therapy. Colloids Surf. B Biointerfaces 2018, 161, 365–374. [Google Scholar] [CrossRef] [PubMed]
- Ayala-Orozco, C.; Urban, C.; Bishnoi, S.; Urban, A.; Charron, H.; Mitchell, T.; Shea, M.; Nanda, S.; Schiff, R.; Halas, N.; et al. Sub-100 nm Gold Nanomatryoshkas Improve Photo-Thermal Therapy Efficacy in Large and Highly Aggressive Triple Negative Breast Tumors. J. Control. Release 2014, 191, 90–97. [Google Scholar] [CrossRef] [PubMed]
- Kong, T.; Zeng, J.; Wang, X.; Yang, X.; Yang, J.; McQuarrie, S.; McEwan, A.; Roa, W.; Chen, J.; Xing, J.Z. Enhancement of Radiation Cytotoxicity in Breast-Cancer Cells by Localized Attachment of Gold Nanoparticles. Small 2008, 4, 1537–1543. [Google Scholar] [CrossRef] [PubMed]
- Dube, E.; Oluwole, D.O.; Nwaji, N.; Nyokong, T. Glycosylated Zinc Phthalocyanine-Gold Nanoparticle Conjugates for Photodynamic Therapy: Effect of Nanoparticle Shape. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2018, 203, 85–95. [Google Scholar] [CrossRef] [PubMed]
- Dozie-Nwachukwu, S.O.; Obayemi, J.D.; Danyuo, Y.; Anuku, N.; Odusanya, O.S.; Malatesta, K.; Soboyejo, W.O. A Comparative Study of the Adhesion of Biosynthesized Gold and Conjugated Gold/Prodigiosin Nanoparticles to Triple Negative Breast Cancer Cells. J. Mater. Sci. Mater. Med. 2017, 28, 143. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Jiang, Y.; Li, X.; Hu, L. Thioglucose-Bound Gold Nanoparticles Increase the Radiosensitivity of a Triple-Negative Breast Cancer Cell Line (MDA-MB-231). Breast Cancer 2015, 22, 413–420. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Kim, H.S.; Jin, T.; Woo, J.; Piao, Y.J.; Moon, W.K. Near-Infrared Photothermal Therapy Using Anti-EGFR-Gold Nanorod Conjugates for Triple Negative Breast Cancer. Oncotarget 2017, 8, 86566–86575. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Kim, H.S.; Jin, T.; Moon, W.K. Near-Infrared Photothermal Therapy Using EGFR-Targeted Gold Nanoparticles Increases Autophagic Cell Death in Breast Cancer. J. Photochem. Photobiol. B. 2017, 170, 58–64. [Google Scholar] [CrossRef] [PubMed]
- Malik, M.T.; O’Toole, M.G.; Casson, L.K.; Thomas, S.D.; Bardi, G.T.; Reyes-Reyes, E.M.; Ng, C.K.; Kang, K.A.; Bates, P.J. AS1411-Conjugated Gold Nanospheres and Their Potential for Breast Cancer Therapy. Oncotarget 2015, 6, 22270–22281. [Google Scholar] [CrossRef] [PubMed]
- Haynes, B.; Zhang, Y.; Liu, F.; Li, J.; Petit, S.; Kothayer, H.; Bao, X.; Westwell, A.D.; Mao, G.; Shekhar, M.P.V. Gold Nanoparticle Conjugated Rad6 Inhibitor Induces Cell Death in Triple Negative Breast Cancer Cells by Inducing Mitochondrial Dysfunction and PARP-1 Hyperactivation: Synthesis and Characterization. Nanomedicine 2016, 12, 745–757. [Google Scholar] [CrossRef] [PubMed]
- Conde, J.; Oliva, N.; Artzi, N. Implantable Hydrogel Embedded Dark-Gold Nanoswitch as a Theranostic Probe to Sense and Overcome Cancer Multidrug Resistance. Proc. Natl. Acad. Sci. USA 2015, 112, E1278–E1287. [Google Scholar] [CrossRef] [PubMed]
- Ayala-Orozco, C.; Urban, C.; Knight, M.W.; Urban, A.S.; Neumann, O.; Bishnoi, S.W.; Mukherjee, S.; Goodman, A.M.; Charron, H.; Mitchell, T.; et al. Au Nanomatryoshkas as Efficient Near-Infrared Photothermal Transducers for Cancer Treatment: Benchmarking Against Nanoshells. ACS Nano 2014, 8, 6372–6381. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Liu, T.; Xie, Y.; Sun, Z.; Liu, H.; Lin, J.; Liu, C.; Mao, Z.W.; Nie, S. Chitosan Layered Gold Nanorods as Synergistic Therapeutics for Photothermal Ablation and Gene Silencing in Triple-Negative Breast Cancer. Acta Biomater. 2015, 25, 194–204. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, S.; Konar, S.; Prasad, P.N.; Rajput, S.; Kumar, B.N.P.; Rao, R.R.; Pathak, A.; Fisher, P.B.; Mandal, M. Micellear Gold Nanoparticles as Delivery Vehicles for Dual Tyrosine Kinase Inhibitor ZD6474 for Metastatic Breast Cancer Treatment. Langmuir 2017, 33, 7649–7659. [Google Scholar] [CrossRef] [PubMed]
- Atkinson, R.L.; Zhang, M.; Diagaradjane, P.; Peddibhotla, S.; Contreras, A.; Hilsenbeck, S.G.; Woodward, W.A.; Krishnan, S.; Chang, J.C.; Rosen, J.M. Thermal Enhancement with Optically Activated Gold Nanoshells Sensitizes Breast Cancer Stem Cells to Radiation Therapy. Sci. Transl. Med. 2010, 2. [Google Scholar] [CrossRef] [PubMed]
- Zhao, N.; Yang, Z.; Li, B.; Meng, J.; Shi, Z.; Li, P.; Fu, S. RGD-Conjugated Mesoporous Silica-Encapsulated Gold Nanorods Enhance the Sensitization of Triple-Negative Breast Cancer to Megavoltage Radiation Therapy. Int. J. Nanomed. 2016, 11, 5595–5610. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Chatterjee, D.K.; Lee, M.H.; Krishnan, S. Gold Nanoparticles in Breast Cancer Treatment: Promise and Potential Pitfalls. Cancer Lett. 2014, 347, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Klassen, N.V.; Kedrov, V.V.; Ossipyan, Y.A.; Shmurak, S.Z.; Shmyt Ko, I.M.; Krivko, O.A.; Kudrenko, E.A.; Kurlov, V.N.; Kobelev, N.P.; Kiselev, A.P.; et al. Nanoscintillators for Microscopic Diagnostics of Biological and Medical Objects and Medical Therapy. IEEE Trans. Nanobiosci. 2009, 8, 20–32. [Google Scholar] [CrossRef] [PubMed]
- Fan, L.; Tian, M.; Liu, Y.; Deng, Y.; Liao, Z.; Xu, J. Salicylate •Phenanthroline Copper (II) Complex Induces Apoptosis in Triple-Negative Breast Cancer Cells. Oncotarget 2017, 8, 29823–29832. [Google Scholar] [CrossRef] [PubMed]
- Afrasiabi, Z.; Stovall, P.; Finley, K.; Choudhury, A.; Barnes, C.; Ahmad, A.; Sarkar, F.; Vyas, A.; Padhye, S. Targeting Triple Negative Breast Cancer Cells by N3-Substituted 9,10-Phenanthrenequinone Thiosemicarbazones and Their Metal Complexes. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2013, 114, 114–119. [Google Scholar] [CrossRef] [PubMed]
- Ahir, M.; Bhattacharya, S.; Karmakar, S.; Mukhopadhyay, A.; Mukherjee, S.; Ghosh, S.; Chattopadhyay, S.; Patra, P.; Adhikary, A. Tailored-CuO-Nanowire Decorated with Folic Acid Mediated Coupling of the Mitochondrial-ROS Generation and miR425-PTEN Axis in Furnishing Potent Anti-Cancer Activity in Human Triple Negative Breast Carcinoma Cells. Biomaterials 2016, 76, 115–132. [Google Scholar] [CrossRef] [PubMed]
- Frik, M.; Martínez, A.; Elie, B.T.; Gonzalo, O.; Ramírez de Mingo, D.; Sanaú, M.; Sánchez-Delgado, R.; Sadhukha, T.; Prabha, S.; Ramos, J.W.; et al. In Vitro and in Vivo Evaluation of Water-Soluble Iminophosphorane Ruthenium(II) Compounds. A Potential Chemotherapeutic Agent for Triple Negative Breast Cancer. J. Med. Chem. 2014, 57, 9995–10012. [Google Scholar] [CrossRef] [PubMed]
- Irace, C.; Misso, G.; Capuozzo, A.; Piccolo, M.; Riccardi, C.; Luchini, A.; Caraglia, M.; Paduano, L.; Montesarchio, D.; Santamaria, R. Antiproliferative Effects of Ruthenium-Based Nucleolipidic Nanoaggregates in Human Models of Breast Cancer in Vitro: Insights into Their Mode of Action. Sci. Rep. 2017, 7, 45236. [Google Scholar] [CrossRef] [PubMed]
- Nhukeaw, T.; Temboot, P.; Hansongnern, K.; Ratanaphan, A. Cellular Responses of BRCA1-Defective and Triple-Negative Breast Cancer Cells and in Vitro BRCA1 Interactions Induced by Metallo-Intercalator Ruthenium(II) Complexes Containing Chloro-Substituted Phenylazopyridine. BMC Cancer 2014, 14, 73. [Google Scholar] [CrossRef] [PubMed]
- Montani, M.; Pazmay, G.V.B.; Hysi, A.; Lupidi, G.; Pettinari, R.; Gambini, V.; Tilio, M.; Marchetti, F.; Pettinari, C.; Ferraro, S.; et al. The Water Soluble Ruthenium(II) Organometallic Compound [Ru(p-cymene)(bis(3,5 dimethylpyrazol-1-yl)methane)Cl]Cl Suppresses Triple Negative Breast Cancer Growth by Inhibiting Tumor Infiltration of Regulatory T Cells. Pharmacol. Res. 2016, 107, 282–290. [Google Scholar] [CrossRef] [PubMed]
- Beauperin, M.; Top, S.; Richard, M.A.; Plażuk, D.; Pigeon, P.; Toma, S.; Poláčková, V.; Jaouen, G. The Length of the Bridging Chain in Ansa-Metallocenes Influences Their Antiproliferative Activity Against Triple Negative Breast Cancer Cells (TNBC). Dalton Trans. 2016, 45, 13126–13134. [Google Scholar] [CrossRef] [PubMed]
- Biancalana, L.; Zacchini, S.; Ferri, N.; Lupo, M.G.; Pampaloni, G.; Marchetti, F. Tuning the Cytotoxicity of Ruthenium(Ii) Para-Cymene Complexes by Mono-Substitution at a Triphenylphosphine/Phenoxydiphenylphosphine Ligand. Dalton Trans. 2017, 46, 16589–16604. [Google Scholar] [CrossRef] [PubMed]
- Becceneri, A.B.; Popolin, C.P.; Plutin, A.M.; Maistro, E.L.; Castellano, E.E.; Batista, A.A.; Cominetti, M.R. The Trans-[Ru(PPh3)2(N,N-dimethyl-N’-thiophenylthioureato-k2O,S)(bipy)]PF6 Complex Has Pro-Apoptotic Effects on Triple Negative Breast Cancer Cells and Presents Low Toxicity in Vivo. J. Inorg Biochem. 2018, 186, 70–84. [Google Scholar] [CrossRef] [PubMed]
- Popolin, C.P.; Reis, J.P.B.; Becceneri, A.B.; Graminha, A.E.; Almeida, M.A.P.; Corrêa, R.S.; Colina-Vegas, L.A.; Ellena, J.; Batista, A.A.; Cominetti, M.R. Cytotoxicity and Anti-Tumor Effects of New Ruthenium Complexes on Triple Negative Breast Cancer Cells. PLoS ONE 2017, 12, e0183275. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Burnett, J.; Zhang, F.; Zhang, J.; Paholak, H.; Sun, D. Highly Crystallized Iron Oxide Nanoparticles as Effective and Biodegradable Mediators for Photothermal Cancer Therapy. J. Mater. Chem. B 2014, 2, 757–765. [Google Scholar] [CrossRef]
- Paholak, H.J.; Stevers, N.O.; Chen, H.; Burnett, J.P.; He, M.; Korkaya, H.; McDermott, S.P.; Deol, Y.; Clouthier, S.G.; Luther, T.; et al. Elimination of Epithelial-Like and Mesenchymal-Like Breast Cancer Stem Cells to Inhibit Metastasis Following Nanoparticle-Mediated Photothermal Therapy. Biomaterials 2016, 104, 145–157. [Google Scholar] [CrossRef] [PubMed]
- Manigandan, A.; Handi, V.; Sundaramoorthy, N.S.; Dhandapani, R.; Radhakrishnan, J.; Sethuraman, S.; Subramanian, A. Responsive Nanomicellar Theranostic Cages for Metastatic Breast Cancer. Bioconjug. Chem. 2018, 29, 275–286. [Google Scholar] [CrossRef] [PubMed]
- Xie, W.; Gao, Q.; Guo, Z.; Wang, D.; Gao, F.; Wang, X.; Wei, Y.; Zhao, L. Injectable and Self-Healing Thermosensitive Magnetic Hydrogel for Asynchronous Control Release of Doxorubicin and Docetaxel to Treat Triple-Negative Breast Cancer. ACS Appl. Mater. Interfaces 2017, 9, 33660–33673. [Google Scholar] [CrossRef] [PubMed]
- Harmon, T.; Harbuzariu, A.; Lanier, V.; Lipsey, C.C.; Kirlin, W.; Yang, L.; Gonzalez-Perez, R.R. Nanoparticle-Linked Antagonist for Leptin Signaling Inhibition in Breast Cancer. World J. Clin. Oncol. 2017, 8, 54–66. [Google Scholar] [CrossRef] [PubMed]
- Vyas, D.; Lopez-Hisijos, N.; Gandhi, S.; El-Dakdouki, M.; Basson, M.D.; Walsh, M.F.; Huang, X.; Vyas, A.K.; Chaturvedi, L.S. Doxorubicin-Hyaluronan Conjugated Super-Paramagnetic Iron Oxide Nanoparticles (DOX-HA-SPION) Enhanced Cytoplasmic Uptake of Doxorubicin and Modulated Apoptosis, IL-6 Release and NF-KappaB Activity in Human MDA-MB-231 Breast Cancer Cells. J. Nanosci. Nanotechnol. 2015, 15, 6413–6422. [Google Scholar] [CrossRef] [PubMed]
- Swanner, J.; Mims, J.; Carroll, D.L.; Akman, S.A.; Furdui, C.M.; Torti, S.V.; Singh, R.N. Differential Cytotoxic and Radiosensitizing Effects of Silver Nanoparticles on Triple-Negative Breast Cancer and Non-Triple-Negative Breast Cells. Int. J. Nanomed. 2015, 10, 3937–3953. [Google Scholar] [CrossRef]
- Juarez-Moreno, K.; Gonzalez, E.B.; Girón-Vazquez, N.; Chávez-Santoscoy, R.A.; Mota-Morales, J.D.; Perez-Mozqueda, L.L.; Garcia-Garcia, M.R.; Pestryakov, A.; Bogdanchikova, N. Comparison of Cytotoxicity and Genotoxicity Effects of Silver Nanoparticles on Human Cervix and Breast Cancer Cell Lines. Hum. Exp. Toxicol. 2017, 36, 931–948. [Google Scholar] [CrossRef] [PubMed]
- Hearn, J.M.; Hughes, G.M.; Romero-Canelón, I.; Munro, A.F.; Rubio-Ruiz, B.; Liu, Z.; Carragher, N.O.; Sadler, P.J. Pharmaco-Genomic Investigations of Organo-Iridium Anticancer Complexes Reveal Novel Mechanism of Action. Metallomics 2018, 10, 93–107. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.J.; Zhong, H.J.; Ko, C.N.; Wong, S.Y.; Vellaisamy, K.; Ye, M.; Ma, D.L.; Leung, C.H. Identification of a Rhodium(III) Complex as a Wee1 Inhibitor Against TP53-Mutated Triple-Negative Breast Cancer Cells. Chem. Commun. 2018, 54, 2463–2466. [Google Scholar] [CrossRef] [PubMed]
- Mooney, R.; Roma, L.; Zhao, D.; van Haute, D.; Garcia, E.; Kim, S.U.; Annala, A.J.; Aboody, K.S.; Berlin, J.M. Neural Stem Cell-Mediated Intratumoral Delivery of Gold Nanorods Improves Photothermal Therapy. ACS Nano 2014, 8, 12450–12460. [Google Scholar] [CrossRef] [PubMed]
- Lacroix, M.; Leclercq, G. Relevance of Breast Cancer Cell Lines as Models for Breast Tumours: An Update. Breast Cancer Res. Treat. 2004, 83, 249–289. [Google Scholar] [CrossRef] [PubMed]
- Holliday, D.L.; Speirs, V. Choosing the Right Cell Line for Breast Cancer Research. Breast Cancer Res. 2011, 13, 215. [Google Scholar] [CrossRef] [PubMed]
- Rock, K.L.; Kono, H. The Inflammatory Response to Cell Death. Annu. Rev. Pathol. 2008, 3, 99–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mammucari, C.; Rizzuto, R. Signaling Pathways in Mitochondrial Dysfunction and Aging. Mech. Ageing Dev. 2010, 131, 536–543. [Google Scholar] [CrossRef] [PubMed]
- Sohur, U.S.; Dixit, M.N.; Chen, C.L.; Byrom, M.W.; Kerr, L.A. Rel/NF-KappaB Represses Bcl-2 Transcription in Pro-B Lymphocytes. Gene Expr. 1999, 8, 219–229. [Google Scholar] [PubMed]
- Selvakannan, P.R.; Mandal, S.; Pasricha, R.; Adyanthaya, S.D.; Sastry, M. One-Step Synthesis of Hydrophobized Gold Nanoparticles of Controllable Size by the Reduction of Aqueous Chloroaurate Ions by Hexadecylaniline at the Liquid-Liquid Interface. Chem. Commun. 2002, 13, 1334–1335. [Google Scholar] [CrossRef]
- Okitsu, K.; Yue, A.; Tanabe, S.; Matsumoto, H.; Yobiko, Y. Formation of Colloidal Gold Nanoparticles in an Ultrasonic Field: Control of Rate of Gold (III) Reduction and Size of Formed Gold Particles. Langmuir 2001, 17, 7717–7720. [Google Scholar] [CrossRef]
- Korbekandi, H.; Ashari, Z.; Iravani, S.; Abbasi, S. Optimization of Biological Synthesis of Silver Nanoparticles Using Fusarium Oxysporum. Iran J. Pharm. Res. 2013, 12, 289–298. [Google Scholar] [PubMed]
- Iravani, S.; Korbekandi, H.; Mirmohammadi, S.V.; Zolfaghari, B. Synthesis of Silver Nanoparticles: Chemical, Physical and Biological Methods. Res. Pharm. Sci. 2014, 9, 385–406. [Google Scholar] [PubMed]
- Lesniak, A.; Kilinc, D.; Rashdan, S.A.; von Kriegsheim, A.; Ashall, B.; Zerulla, D.; Kolch, W.; Lee, G.U. In Vitro Study of the Interaction of Heregulin-Functionalized Magnetic-Optical Nanorods with MCF7 and MDA-MB-231 Cells. Faraday Discuss. 2014, 175, 189–201. [Google Scholar] [CrossRef] [PubMed]
- Greish, K. Enhanced Permeability and Retention (EPR) Effect for Anticancer Nanomedicine Drug Targeting. Methods Mol. Biol. 2010, 624, 25–37. [Google Scholar] [CrossRef] [PubMed]
- Matsumura, Y.; Maeda, H. A New Concept for Macromolecular Therapeutics in Cancer Hemotherapy: Mechanism of Tumoritropic Accumulation of Proteins and the Antitumor Agent Smancs. Cancer Res. 1986, 46, 6387–6392. [Google Scholar] [PubMed]
- Maeda, H.; Wu, J.; Sawa, T.; Matsumura, Y.; Hori, K. Tumor Vascular Permeability and the EPR Effect in Macromolecular Therapeutics: A Review. J. Control. Release 2000, 65, 271–284. [Google Scholar] [CrossRef]
- Dam, D.H.; Culver, K.S.; Kandela, I.; Lee, R.C.; Chandra, K.; Lee, H.; Mantis, C.; Ugolkov, A.; Mazar, A.P.; Odom, T.W. Biodistribution and in Vivo Toxicity of Aptamer-Loaded Gold Nanostars. Nanomedicine 2015, 11, 671–679. [Google Scholar] [CrossRef] [PubMed]
- Cioce, M.; Gherardi, S.; Viglietto, G.; Strano, S.; Blandino, G.; Muti, P.; Ciliberto, G. Mammosphere-Forming Cells from Breast Cancer Cell Lines as a Tool for the Identification of CSC-Like- and Early Progenitor-Targeting Drugs. Cell Cycle 2010, 9, 2878–2887. [Google Scholar] [CrossRef] [PubMed]
- Rosen, J.M.; Jordan, C.T. The Increasing Complexity of the Cancer Stem Cell Paradigm. Science 2009, 324, 1670–1673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bravo-Cordero, J.J.; Hodgson, L.; Condeelis, J. Directed Cell Invasion and Migration during Metastasis. Curr. Opin. Cell Biol. 2012, 24, 277–283. [Google Scholar] [CrossRef] [PubMed]
- Anampa, J.; Sparano, J.A. New Agents for the Management of Resistant Metastatic Breast Cancer. Expert Opin. Pharmacother. 2017, 18, 1815–1831. [Google Scholar] [CrossRef] [PubMed]
- Toraya-Brown, S.; Sheen, M.R.; Zhang, P.; Chen, L.; Baird, J.R.; Demidenko, E.; Turk, M.J.; Hoopes, P.J.; Conejo-Garcia, J.R.; Fiering, S. Local Hyperthermia Treatment of Tumors Induces CD8(+) T Cell-Mediated Resistance Against Distal and Secondary Tumors. Nanomedicine 2014, 10, 1273–1285. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, B.B.; Shishodia, S.; Sandur, S.K.; Pandey, M.K.; Sethi, G. Inflammation and Cancer: How Hot Is the Link? Biochem. Pharmacol. 2006, 72, 1605–1621. [Google Scholar] [CrossRef] [PubMed]
- Salgado, R.; Junius, S.; Benoy, I.; van Dam, P.; Vermeulen, P.; van Marck, E.; Huget, P.; Dirix, L.Y. Circulating Interleukin-6 Predicts Survival in Patients with Metastatic Breast Cancer. Int. J. Cancer 2003, 103, 642–646. [Google Scholar] [CrossRef] [PubMed]
- Berberoglu, U.; Yildirim, E.; Celen, O. Serum Levels of Tumor Necrosis Factor Alpha Correlate with Response to Neoadjuvant Chemotherapy in Locally Advanced Breast Cancer. Int. J. Biol. Markers 2004, 19, 130–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartmann, L.C.; Keeney, G.L.; Lingle, W.L.; Christianson, T.J.; Varghese, B.; Hillman, D.; Oberg, A.L.; Low, P.S. Folate Receptor Overexpression Is Associated with Poor Outcome in Breast Cancer. Int. J. Cancer 2007, 121, 938–942. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Subtype | TNBC Cell Lines | Others Immunohistochemical Markers besides ER, PR and HER2 a | Tumor of Origin |
|---|---|---|---|
| A | BT-20 | WNT3 oncogene +, WNT7B oncogene + | Invasive ductal carcinoma |
| CAL148 | CK 7 +, CK 8 +, CK 17 − CK 18 +, CK 19 +, desmin −, endothel −, EpCAM +, GFAP −, neurofilament −, vimentin − | Adenocarcinoma | |
| DU4475 | CK 7 −, CK 8 +, CK 17 −, CK 18 +, CK 19 −, desmin −, endothel −, EpCAM +, GFAP −, neurofilament −, vimentin − | Invasive ductal carcinoma | |
| EMG3 | CK 5 +, CK 14 +, CK 18 +, CK 19 −, EMA +, P63+, SMA −, ESA +, EGFR + | Invasive ductal carcinoma | |
| HCC1143 | p53 +, EGP2 +, CK 19 + | Ductal carcinoma | |
| HCC1187 | EGP2 +, CK 19 +, p53 +++ | Ductal carcinoma | |
| HCC1599 | EGP2 +, CK 19 +, p53 − | Ductal carcinoma | |
| HCC1806 | p53 −, EGP2 +, CK 19 +, Ob-R + | Squamous carcinoma | |
| HCC1937 | p53 −, EGP2 +, CK 19 + | Ductal carcinoma | |
| HCC2157 | EGP2 +, CK 19 +, p53 + | Ductal carcinoma | |
| HCC3153 | N.A. | Ductal carcinoma | |
| HCC70 | p53 +++, EGP2 + CK 19 + | Ductal carcinoma | |
| HMT3522 | Vimentin −, PLK1 + | Benign tumor | |
| KPL-3C | Keratins −, Vimentin − | Invasive ductal carcinoma | |
| MA11 | e-cadherin + | Invasive lobular carcinoma | |
| MDA-MB-436 | Tubulin +, actin +, ICAM +++ | Adenocarcinoma | |
| MDA-MB-468 | EGFR +, TGF alpha +, Ki67 high | Adenocarcinoma | |
| MFM223 | CK 7 +, CK 8 +, CK 17 −, CK 18 +, CK 19 +, desmin −, endothel −, EpCAM +, GFAP −, neurofilament −, vimentin − | Carcinoma | |
| SUM229PE | CK 8 +, CK 18 +, CK 19 + | Ductal carcinoma | |
| B | A17 | Vimentin +, CK 14 +, N-cadherin +, COX2 +++ | Carcinoma |
| BCM-2665A | ALDH + | Invasive ductal carcinoma | |
| BT-549 | Ki67 low, e-cadherin low, claudin-3 low, claudinin-4 low, claudinin-7 low | Invasive ductal carcinoma | |
| CAL-120 | CK 7 −, CK 8 +, CK 17 −, CK 18 +, CK 19 +, desmin −, endothel −, EpCAM +, GFAP −, neurofilament −, vimentin + | Adenocarcinoma | |
| CAL-51 | CK 7 −, CK 8 +, CK 17 −, CK 18 +, CK 19 +, desmin −, endothel −, EpCAM +, GFAP −, neurofilament −, vimentin + | Adenocarcinoma | |
| CAL-85-1 | CK 7 +, CK 8 +, CK 17 +, CK 18 +, CK 19 +, desmin −, endothel −, EpCAM +, GFAP −, neurofilament −, vimentin + | Adenocarcinoma | |
| HCC1395 | p53 +, EGP2 +, CK 19 + | Ductal carcinoma | |
| HCC1739 | N.A. | Ductal carcinoma | |
| HCC38 | p53 +, EGP2 +, CK 19 + | Ductal carcinoma | |
| HDQ-P1 | CK 7 +, CK 8 +, CK 17 +, CK 18 +, CK 19 +, desmin −, endothel −, EpCAM +, GFAP −, neurofilament −, vimentin + | Invasive ductal carcinoma | |
| Hs578T | Ki67 low, e-cadherin low, claudin-3 low, claudinin-4 low, claudinin-7 low, EGFR + | Invasive ductal carcinoma | |
| MDA-MB-157 | WNT7B oncogene +, ICAM +++ | Medullary carcinoma | |
| MDA-MB-231 | EGFR +, TGF alpha +, CD44 +++, LHRH +++, WNT7B oncogene +, ICAM-1 +++, Ki67 low, e-cadherin low, claudin-3 low, claudinin-4 low, claudinin-7 low, alpha-V beta-3 integrin +, Ob-R + | Adenocarcinoma | |
| OCUB-M | e-cadherin +, Laminin + | Carcinoma | |
| SUM1315 | EGFR + | Carcinoma | |
| SUM149 | CK 8 +, CK 18 +, CK 19 + | Carcinoma | |
| SUM159 | claudin low, CK 8 +, CK 18 +, CK 19 + | Carcinoma |
| In Vitro | ||||||
| Cell Line | NPMBC | Assay | Mode of Action/Pathway Involved | Effect Observed | Doses Assayed | Reference |
| MDA-MB-231 | Multibranched gold nanoantennas | Calcein/PI staining assay | --- | Induction of cell death | 170 μg/mL | [14] |
| MDA-MB-231 SUM159 MDA-MB-468 BT-549 | Liposome encapsulated ruthenium polypyridine complex | MTT assay | --- | Reduced cell viability | <4 µM | [16] |
| MDA-MB-231 | Liposome encapsulated ruthenium polypyridine complex | Western blot Immunofluorescence | γ-H2AX ATM | Increased levels of γ-H2AX Degradation and dephosphorylation of ATM | 3 µM | [16] |
| MDA-MB-231 | Gold nanostars loaded with AS1411 | Resazurin cell viability assay | --- | 40% cell viability | 0.3 nM | [27] |
| MDA-MB-231 | CD44-targeting HSP72 depletion nanosystem based on plasmonic gold nanoparticles | MTT assay | --- | 20–85% cell death | 50 mg/L | [28] |
| MDA-MB-231 | Multibranched gold nanoantennas | MTT assay | --- | ~40% cell viability | 170 μg/mL | [29] |
| MDA-MB-231 | Gold nanoparticles | MTT assay | --- | 15–75% survival | 0.1–100 µM | [31] |
| MDA-MB-231 | Thioglucose-bound gold nanoparticles | MTT assay | --- | Inhibited cell growth | 20 nM | [36] |
| MDA-MB-231 | Thioglucose-bound gold nanoparticles | Clonogenic cell survival assay | --- | Enhanced radiosensitivity | 20 nM | [36] |
| MDA-MB-231 MDA-MB-468 | Anti-EGFR-gold nanorod conjugates | MTT assay | --- | Inhibited cell growth | 1.84 µg/mL | [37] |
| MDA-MB-231 | Anti-EGFR-gold nanorod conjugates | Western blot | Ki-67 EGFR Akt ERK1/2 mTOR FAK | Decreased Ki-67 and EGFR expression Suppression of phosphorylation of FAK, ERK1/2, mTOR and Akt | 1.84 µg/mL | [37] |
| MDA-MB-231 | Anti-EGFR-gold nanorod conjugates | CCK-8 assay | --- | 32.2% cell viability | 120 pM | [38] |
| MDA-MB-231 | Gold nanospheres conjugated with AS1411 | MTT assay | --- | Inhibited cell viability | <100 nM | [39] |
| MDA-MB-231 SUM1315 HCC1937 MDA-MB-468 | Gold-nanoparticles conjugated with Rad6 inhibitor | MTT assay Trypan blue staining assay | --- | Decreased cell viability (SUM1315 and MDA-MB-231) No change in viability of HCC1937 and MDA-MB-468 | 0.5 µM for SUM1315; 8.2 µM for MDA-MB-231 | [40] |
| MDA-MB-231 | Gold nanoparticles loaded with 5-fluorouracil | MTT assay | --- | Decreased cell viability | 0.5 nM | [41] |
| MDA-MB-231 | Chitosan layered gold nanorods | MTS-based colorimetric assay | --- | ~65% inhibition of cell proliferation | 100 nM | [43] |
| MDA-MB-231 MDA-MB-468 | Micellar gold nanoparticles conjugated with ZD6474 | MTT assay | --- | 80.51% (MDA-MB-231) and ~92% (MDA-MB-468) inhibition of cell proliferation | 1–10 µM | [44] |
| MDA-MB-231 BT-20 | Copper salicylate phenanthroline complexes | MTS-based colorimetric assay | --- | Inhibition of cell growth | 5–25 µM | [49] |
| MDA-MB-231 | Copper complexes of phenanthrenequinone thiosemicarbazone | MTT assay | --- | Decreased cell viability | 2.3–4.8 µM | [50] |
| MDA-MB-231 | Nickel complexes of phenanthrenequinone thiosemicarbazone | MTT assay | --- | Decreased cell viability | >10 µM | [50] |
| MDA-MB-231 | Palladium complexes of phenanthrenequinone thiosemicarbazone | MTT assay | --- | Decreased cell viability | >10 µM | [50] |
| MDA-MB-231 | Copper oxide nanowire conjugated with folic acid | Flow cytometry | --- | 60% cell death | 2.5 µg/mL | [51] |
| MDA-MB-231 | Water-soluble iminophosphorane ruthenium(II) compounds | MTT assay | --- | Decreased cell viability | 2.61–75.4 µM | [52] |
| MDA-MB-231 MDA-MB-468 MDA-MB-436 | Ruthenium-based nucleolipidic nanoaggregates | MTT assay Trypan blue | --- | Reduced cell proliferation | 12.1–14.7 µM | [53] |
| MDA-MB-231 HCC1937 | Ruthenium complexes | Real-time cell growth profiling MTT assay | --- | Inhibited cell viability | 13.2 and 14.1 µM for MDA-MB-231; 1.8 and 9.9 µM for HCC1937 | [54] |
| A17 MDA-MB-231 | Ruthenium complexes | MTT assay | --- | Inhibited cell viability | 230.66 µM for A17; 409.89 µM for MDA-MB-231 | [55] |
| MDA-MB-231 | Ansa-ferrocenes ruthenocenophanes | MTT assay | --- | Decreased cell viability | 0.09–4.53 µM | [56] |
| MDA-MB-231 | Ruthenium(II) paracyme complexes | Sulforhodamine B assay | --- | Decreased cell viability | 9.2–29.1 µM | [57] |
| MDA-MB-231 | Ruthenium complexes | MTT assay | --- | Decreased cell viability | 8.81–21.92 µM | [58] |
| MDA-MB-231 | Ruthenium complexes | Colony formation | --- | 100% inhibition of the colony formation | 2 µM | [58] |
| MDA-MB-231 | Ruthenium-based compounds | MTT assay | --- | Decreased cell viability | 31.16 and 52.74 µM | [59] |
| MDA-MB-231 | Ruthenium-based compounds | Colony formation | --- | 100% inhibition of the colony formation | 20 µM | [59] |
| SUM159 | Iron oxide nanoparticles | Alamar blue cell viability assay Flow cytometry | --- | 5–80% cell viability | 10 µM | [61] |
| MDA-MB-231 | Supermagnetic iron oxide nanoparticles conjugated with doxorubicin | MTS-based colorimetric assay | --- | 31%-cell viability | 520 µM | [62] |
| MDA-MB-231 | Chitosan hydrogel cross-linked with telechelic difunctional poly(ethylene glycol) modified iron oxide magnetic nanoparticles | CCK-8 assay | --- | 5.6% cell viability | 10.8–35.8 µM | [63] |
| MDA-MB-231 HCC1806 | Iron oxide nanoparticles linked antagonist for leptin | MTT assay | --- | Prevention of leptin-induced cell proliferation | 0.0036 pmol/L | [64] |
| MDA-MB-231 HCC1806 | Iron oxide nanoparticles linked antagonist for leptin | Immunoblotting analysis | Cyclin D1 | Prevention of leptin-induced cyclin D1 expression | 0.0036 pmol/L | [64] |
| MDA-MB-231 HCC1806 | Iron oxide nanoparticles linked antagonist for leptin | Cell cycle analysis | --- | Prevention of leptin-induced cell cycle progression | 0.0018–0.0036 pmol/L for MDA-MB-231 and 0.0018–0.036 pmol/L for HCC1806 | [64] |
| MDA-MB-231 HCC 1806 | Iron oxide nanoparticles linked antagonist for leptin | FITC-annexin V/PI staining assay | --- | Decreased cell viability | 0.0036 pmol/L | [64] |
| MDA-MB-231 | Doxorubicin-hyaluronan conjugated iron oxide nanoparticles | Confocal microscopy | Vimentin e-cadherin | Decreased vimentin expression No change in e-cadherin expression | 2 µM | [65] |
| MDA-MB-231 BT549 SUM-159 | Silver nanoparticles | MTT assay | --- | Inhibited cell viability | ≥10 µg/mL | [66] |
| MDA-MB-231 BT549 SUM-159 | Silver nanoparticles | Clonogenic assay | --- | 100% inhibition of clonogenic growth | ≥10 µg/mL | [66] |
| MDA-MB-231 | Silver nanoparticles | MTT assay | --- | Decreased cell viability | 6.72 µg/mL at 12 h and 2.62 µg/mL at 24 h | [67] |
| MDA-MB-468 OCUB-M | Organo-iridium complexes Organo-osmium complexes | MTT assay | --- | Decreased cell viability | <1 µM | [68] |
| MDA-MB-468 MDA-MB-231 | Rhodium(III) complex | MTT assay | --- | Inhibited cell proliferation | 0.35 and 0.55 µM | [69] |
| MDA-MB-231 | Rhodium(III) complex | Western blot | Wee 1 Cdc2 ERK Akt Cyclin A2 γ-H2AX | Decreased phosphorylation of ERK, Akt, Cdc2 Decreased expression of cyclin A2 Increased γ-H2AX levels Inhibition of Wee1 activity | 1–3 μM | [69] |
| In Vivo | ||||||
| Cell Line | NPMBC | Animal (Model)/Analysis from Animal Tissue | Mode of Action/Pathway Involved | Effect Observed | Doses Assayed (Route of Administration) | Reference |
| MDA-MB-231 | Liposome encapsulated ruthenium polypyridine complex | Athymic nude mice (orthotopic)/Immunofluorescen imaging | Ki-67 | Decreased Ki-67 level | 5 mg ruthenium/kg (i.v.) | [16] |
| MDA-MB-231 | Liposome encapsulated ruthenium polypyridine complex | Athymic nude mice (orthotopic) | --- | Suppression of tumor growth (tumor weights: 0.342 with nanoparticles vs. 0.992 with control) | 5 mg ruthenium/kg (i.v.) | [16] |
| MDA-MB-231 | CD44-targeting HSP72 depletion nanosystem based on plasmonic gold nanoparticles | Nude mice (xenograft) | --- | Smaller tumor volume than control | 10 mg/kg (i.v.) | [28] |
| MDA-MB-231 | Gold nanomatryoshkas and nanoshells | Sprague-Dawley athymic nude mice (xenograft) | --- | Improved survival (15.8 and 11.3 days with nanomatryoshkas and nanoshells respectively) Slower tumor growth with nanomatryoshkas | 300 µg of gold (i.v.) | [32] |
| MDA-MB-231 | Anti-EGFR-gold nanorod conjugates | BALB/c nude mice (xenograft) | --- | Smaller tumor volumen than control (~<50 vs. 250 mm3 respectively) | 0.5 pmol/g mouse (i.v.) | [37] |
| MDA-MB-231 | Anti-EGFR-gold nanorod conjugates | BALB/c nude mice (xenograft)/Immunohistochemistry | Ki-67 EGFR | Decreased Ki-67 and EGFR levels | 0.5 pmol/g mouse (i.v.) | [37] |
| MDA-MB-231 | Gold nanospheres conjugated with AS1411 | Nude mice Fox1nu (xenograft) | --- | Decreased tumor grow rate and tumor regression (tumor size at 12 days: 100 with nanospheres vs. 1000 mm3 with control) | 1 mg/kg/day (i.p.) | [39] |
| MDA-MB-231 | Hydrogel embedded with gold nanoparticles | SCID hairless congenic mice (orthotopic) | --- | Efficient and sustained inhibition of tumor growth (~90% tumor size reduction) | 10 nM of gold nanoparticles (i.h.) | [41] |
| MDA-MB-231 | Gold nanomatryoshkas and nanoshells | Sprague-Dawley athymic nude mice (xenograft) | --- | Improved survival (83% and 33% at 60 days with nanomatryoshkas and nanoshells respectively) Complete tumor regression at 60 days post-treatment with relapse rates between 16.7% (nanomatryoshkas) and 50% (nanoshells) | 300 µg of gold (i.v.) | [42] |
| MDA-MB-231 | Micellar gold nanoparticles conjugated with ZD6474 | Athymic BALB/c (nu+/nu+) mice (xenograft) | --- | Decrease in tumor volume (63% of control) | 30 mg/kg of ZDD6474 (i.v.) | [44] |
| BCM-2665A | Gold nanoshells | SCID/beige mice (xenograft) | --- | Smaller tumor volume compared to control (400 vs. 800 mm3 respectively) | ~8 × 108 nanoshells/g body weight (i.v.) | [45] |
| MDA-MB-231 | RGD-conjugated mesoporous silica-encapsulated gold nanorods | Nude mice (orthotopic) | --- | Smaller tumor volume than control (569 vs. 1302 mm3 respectively) | ∼50 μg of gold/g of body weight (i.v.) | [46] |
| MDA-MB-231 | Copper salicylate phenanthroline complexes | Nude mice (xenograft) | --- | Smaller tumor volume than control (20 vs. 35 mm3 respectively) | 0.5 mg/mL (i.p.) | [49] |
| MDA-MB-231 | Copper salicylate phenanthroline complexes | Nude mice (xenograft)/Immunohistochemical staining | Ki-67 | Reduced Ki-67 expression | 0.5 mg/mL (i.p.) | [49] |
| MDA-MB-231 | Water-soluble iminophosphorane ruthenium(II) compounds | NOD.CB17-Prkdc SCID/J mice (xenograft) | --- | Significant inhibition of tumor growth and smaller tumor size than control (56% decrease vs. 200% increase in tumor volume respectively) | 5 mg/kg/day (i.p.) | [52] |
| A17 | Ruthenium complexes | FVB/NCrl mice (xenograft) | --- | Decreased tumor grow rate and smaller tumor size than control (<3 vs. 7 mm) | 210 mg/kg/day (i.p.) | [55] |
| SUM159 | Highly crystallized iron oxide nanoparticles | BALB/c immune-competent mice (xenograft) | --- | Complete tumor regression without relapses | 20 mg iron/kg body weight (i.v.) | [60] |
| SUM159 | Iron oxide nanoparticles | NOD/SCID immune-compromised mice (orthotopic) | --- | Smaller tumor size than control | 20 mg iron/kg body weight (i.v.) | [61] |
| 4T1 | Supermagnetic iron oxide nanoparticles conjugated with doxorubicin | BALB/c mice (orthotopic) | --- | Smaller tumor size than control (7 vs. 74 mm3 respectively) | 0.25 mg iron/100 mm3 (i.t.) | [62] |
| MDA-MB-231 | Chitosan hydrogel cross-linked with telechelic difunctional poly(ethylene glycol) modified iron oxide magnetic nanoparticles | BALB/c mice (xenograft) | --- | Smaller tumor size than control (relative tumor volume ~0.75 vs. 4.5 v/v respectively) | 18.7 mg/kg (i.t.) | [63] |
| MDA-MB-231 | Silver nanoparticles | nu/nu athymic mice (xenograft) | --- | Decreased tumor grow rate and smaller tumor size than control (250 vs. 550 mm3 respectively) | 0.2 µg/mm3 tumor volume (i.t.) | [66] |
| MDA-MB-231 | Gold nanorod-loaded neural stem cells | Athymic nude mice (xenograft) | --- | Complete tumor ablation, improved survival and reduced tumor recurrence compared to control | 12.5 µg/(i.t.) | [70] |
| In Vitro | ||||||
| Cell Line | NPMBC | Assay | Mode of Action/Pathway Involved | Effect Observed | Doses Assayed | Reference |
| MDA-MB-231 | Liposome encapsulated ruthenium polypyridine complex | Western blot | PARP Caspase 3 Caspase 8 Caspase 9 | Increased cleaved PARP, caspase 3, caspase 8 and caspase 9 levels | 5 µM | [16] |
| MDA-MB-231 | Liposome encapsulated ruthenium polypyridine complex | Flow cytometry (cell cycle arrest) | --- | 75.9% apoptotic cells 54% cells in the G2/M phase | 5 µM | [16] |
| MDA-MB-231 | Gold nanostars | Fluorescent assay qPCR | Caspase 3/7 Bcl-2 | Increased caspase activity Downregulation of Bcl-2 expression | 0.3 nM | [27] |
| MDA-MB-231 | Multibranched gold nanoantennas | Annexin V/calcein/PI staining assay | --- | Induction of apoptotic cell death | 170 μg/mL | [29] |
| MDA-MB-231 | Gold nanoparticles conjugated gemcitabine | Annexin V/7-AAD assay | --- | Induction of apoptosis | 0.5, 1, 5, 25 nM | [30] |
| MDA-MB-231 | Anti-EGFR-gold nanorod conjugates | FITC-annexin V/PI staining assay | --- | ~25–40% apoptotic cells | 1.84 µg/mL | [37] |
| MDA-MB-231 | Anti-EGFR-gold nanorod conjugates | Western blot | Caspase 3 | Increased cleaved caspase 3 protein | 1.84 µg/mL | [37] |
| MDA-MB-231 | Gold nanospheres | FITC-annexin V/PI staining assay | --- | 88% apoptotic cells | 200 nM | [39] |
| MDA-MB-231 SUM1315 HCC1937 MDA-MB-468 | Gold-nanoparticles conjugated with Rad6 inhibitor | Acridine orange/ethidium bromide staining | --- | SUM1315 and MDA-MB-231 cells in early and late stages of apoptosis No morphological alterations consistent with apoptosis were observed in HCC1937 or MDA-MB-468 cells | 1–5 μM | [40] |
| SUM1315 HCC1937 | Gold-nanoparticles conjugated with Rad6 inhibitor | Mitochondrial membrane potential (JC-1) | --- | Loss in the mitochondrial function of SUM1315 cells Mitochondrial function of HCC1937 cells was unaffected | 1 µM | [40] |
| SUM1315 HCC1937 | Gold-nanoparticles conjugated with Rad6 inhibitor | Western blot | PARP-1 | Activation of PARP-1 | 1–5 μM | [40] |
| MDA-MB-468 | Micellar gold nanoparticles conjugated with ZD6474 | FITC-annexin V/PI staining assay | --- | 28.2% apoptotic cells | 5 µM | [44] |
| MDA-MB-231 | RGD-conjugated mesoporous silica-encapsulated gold nanorods | FITC-annexin V/PI staining assay | --- | Increased apoptotic cells | 50 µg/mL | [46] |
| MDA-MB-231 | RGD-conjugated mesoporous silica-encapsulated gold nanorods | DHE fluorescence method | ROS | Increased cellular ROS levels | 50 µg/mL | [46] |
| MDA-MB-231 BT-20 | Copper salicylate phenanthroline complexes | Flow cytometry (Annexin V/PI) | --- | >80% apoptotic cells | 25 µM | [49] |
| MDA-MB-231 BT-20 | Copper salicylate phenanthroline complexes | Western blot | Bcl-2 Bcl-xL Survivin PARP | Decreased Bcl-2, Bcl-xL and surviving expression Increased cleaved PARP expression | 25 μM | [49] |
| MDA-MB-231 | Copper oxide nanowire conjugated with folic acid | FITC-annexin V/PI staining assay | --- | Induction of apoptosis | 2.5 µg/mL | [51] |
| MDA-MB-231 | Copper oxide nanowire conjugated with folic acid | SEM | --- | Formation of apoptotic bodies and membrane blebbing | 2.5 µg/mL | [51] |
| MDA-MB-231 | Copper oxide nanowire conjugated with folic acid | Flow cytometry | --- | Induction of ROS | 2.5 µg/mL | [51] |
| MDA-MB-231 | Copper oxide nanowire conjugated with folic acid | Mitochondrial membrane potential (JC-1) Immunofluorescence Western blotting analysis | ROS Cyt C | Loss in the mitochondrial membrane potential Increased expression of Cyt C in the cytosol | 2.5 µg/mL | [51] |
| MDA-MB-231 | Copper oxide nanowire conjugated with folic acid | Western blotting analysis | Caspase 3 Caspase 9 | Increased cleaved caspase 9 and caspase 3 expression | 2.5 µg/mL | [51] |
| MDA-MB-231 | Copper oxide nanowire conjugated with folic acid | Western blotting analysis Immuno-fluorescence staining | NF-κB p65 protein | Inhibition of nuclear transport of the p65 subunit of NF-κB protein | 2.5 µg/mL | [51] |
| MDA-MB-231 | Copper oxide nanowire conjugated with folic acid | qRT-PCR Western blot | miR425 PTEN Akt | Down-regulation of miR-425 and Akt, and up-regulation of PTEN | 2.5 µg/mL | [51] |
| MDA-MB-231 | Ruthenium-based nucleolipidic nanoaggregates | Phase-contrast microscopy | --- | Cell shrinkage and loss of cell-cell contact | 14.7 µM | [53] |
| MDA-MB-231 | Ruthenium-based nucleolipidic nanoaggregates | FITC-annexin V/PI staining assay | --- | ~80% of cells in late apoptosis phase | 14.7 µM | [53] |
| MDA-MB-231 | Ruthenium-based nucleolipidic nanoaggregates | DNA fragmentation assay | --- | Internucleosomal DNA laddering | 14.7 µM | [53] |
| MDA-MB-231 | Ruthenium-based nucleolipidic nanoaggregates | Western blot | Bax Bcl-2 Caspase 3 Caspase 9 Caspase 8 | Up-regulated Bax expression Down-regulated Bcl-2 expression Activation of caspase 9, caspase 8 and caspase 3 | 14.7 µM | [53] |
| MDA-MB-231 HCC1937 | Ruthenium complexes | RT-PCR | P21 p53 BRCA1 | Upregulated p53, P21 and BRCA1 expression | 13.2 and 14.1 µM for MDA-MB-231; 1.8 and 9.9 µM for HCC1937 | [54] |
| A17 MDA-MB-231 | Ruthenium complexes | Immunohistochemistry Western blot | Caspase 3 | Increased number of apoptotic cells (cleaved caspase 3 positive cells) | 750 μM for A17; 1 mM for MDA-MB-231 | [55] |
| MDA-MB-231 | Ruthenium complexes | DAPI staining | --- | Nuclear chromatin condensation with formation of apoptotic bodies | 2, 4 and 8 µM | [58] |
| MDA-MB-231 | Ruthenium complexes | Flow cytometry (PE annexin-V) | --- | 43% apoptotic cells | 2 µM | [58] |
| MDA-MB-231 | Ruthenium complexes | qRT-PCR | Bax Caspase 3 Bcl-2 | Increased Bax and caspase 3 expression Decreased Bcl-2 expression | 4, 8 and 16 µM | [58] |
| MDA-MB-231 | Ruthenium complexes | Western blotting | Bax Caspase 3 Bcl-2 | Increased Bax expression Decreased Bcl-2 expression No change in caspase-3 expression | 0.5, 1, 2, 4, 8 and 16 µM | [58] |
| MDA-MB-231 | Ruthenium complexes | Comet assay | --- | DNA damage | 4 and 8 µM | [58] |
| MDA-MB-231 | Ruthenium-based compounds | DAPI staining | --- | Nuclear fragmentation | 60 and 70 µM | [59] |
| MDA-MB-231 | Ruthenium-based compounds | Flow cytometry (PE annexin-V) | --- | 74.4% apoptotic cells | 20 µM | [59] |
| MDA-MB-231 | Ruthenium-based compounds | qRT-PCR | Bax Caspase 3 Bcl-2 | Increased expression of Bax and caspase 3. No change in gene expression of Bcl-2 | 40 µM | [59] |
| MDA-MB-231 | Ruthenium-based compounds | Western blotting | Caspases 3 and 9 | Increased caspase 3 and 9 expression | 2.5 µM | [59] |
| MDA-MB-231 | Ruthenium-based compounds | Western blotting | Bcl-2 | No change in Bcl-2 expression | 10 µM | [59] |
| MDA-MB-231 | Supermagnetic iron oxide nanoparticles conjugated with doxorubicin | Mitochondrial membrane potential (JC-1) | --- | Increased depolarization of mitochondrial membrane potential | 520 µM | [62] |
| MDA-MB-231 HCC 1806 | Iron oxide nanoparticles linked antagonist for leptin | Immunoblotting analysis | STAT3 | Prevention of leptin-induced STAT3 expression only in HCC1806 cells | 0.0036 pmol/L | [64] |
| MDA-MB-231 | Doxorubicin-hyaluronan conjugated iron oxide nanoparticles | FITC-annexin V/PI staining assay | --- | 12% apoptotic cells (early and late stages of apoptosis) | 2 µM | [65] |
| MDA-MB-231 | Doxorubicin-hyaluronan conjugated iron oxide nanoparticles | Steady-Glo luciferase reagent assay | NF-κB | Reduced NF-κB transcriptional activity (~40%) | 2 µM | [65] |
| MDA-MB-231 | Silver nanoparticles | FITC-annexin V/PI staining assay | --- | Cells in early and late stages of apoptosis | 1.25–10 µg/mL | [67] |
| MDA-MB-231 | Silver nanoparticles | Comet assay | --- | DNA damage | 5 and 10 µg/mL | [67] |
| MDA-MB-231 | Silver nanoparticles | Flow cytometry Confocal microscopy Fluorescence microscopy | ROS | Increase ROS levels | 2.5, 5 and 10 µg/mL | [67] |
| MDA-MB-231 | Rhodium(III) complex | Western blot | Caspase 3 Caspase 7 Caspase 8 Bcl-2 PARP | Increased PARP, and cleaved caspases-3, -7, and -8 expression Decreased Bcl-2 expression | 1 and 3 µM | [69] |
| In Vivo | ||||||
| Cell Line | NPMBC | Animal (Model))/Analysis from Animal Tissue | Mode of Action/Pathway Involved | Effect Observed | Doses Assayed (Route of Administration) | Reference |
| MDA-MB-231 | Liposome encapsulated ruthenium polypyridine complex | Athymic nude mice (orthotopic))/Western blot, immunohistochemistry | PARP Caspase 3 Caspase 8 Caspase 9 TUNEL | Increased TUNEL cleaved PARP, caspase 3, caspase 8 and caspase 9 levels | 5 mg ruthenium/kg (i.v.) | [16] |
| MDA-MB-231 | CD44-targeting HSP72 depletion nanosystem based on plasmonic gold nanoparticles | Immunohistochemistry | TUNEL | Massive apoptotic cells | 10 mg/kg (i.v.) | [28] |
| MDA-MB-231 | Anti-EGFR-gold nanorod conjugates | BALB/c nude mice (xenograft))/ Immunohistochemistry | TUNEL Caspase 3 | Increased TUNEL and caspase 3 levels | 0.5 pmol/g mouse (i.v.) | [37] |
| MDA-MB-231 | Copper salicylate phenanthroline complexes | Nude mice (xenograft))/ Immunohistochemical staining | Bcl-2 Bcl-xL Survivin | Reduced Bcl-2, survivin and Bcl-xL expression | 0.5 mg/mL (i.p.) | [49] |
| 4T1 | Copper oxide nanowire conjugated with folic acid | Immunohistochemical analysis of tumors of BALB/c mice (orthotopic)) | --- | Suppression of tumor growth (lower tumor weight and volume) | 2.5 mg/kg body weight (i.p.) | [51] |
| 4T1 | Copper oxide nanowire conjugated with folic acid | Immunohistochemical analysis of tumors of BALB/c mice (orthotopic))/Immunohistochemical analysis and western blot | PTEN miR425 | Increased PTEN expression and decreased miR-425 expression | 2.5 mg/kg body weight (i.p.) | [51] |
| A17 | Ruthenium complexes | FVB/NCrl mice (xenograft))/Immunohistochemistry analysis | Caspase 3 | Increased number of apoptotic cells in tumors (cleaved caspase-3 positive cells) | 210 mg/kg/day (i.p.) | [55] |
| In Vitro | ||||||
| Cell Line | NPMBC | Assay | Mode of Action/Pathway Involved | Effect Observed | Doses Assayed | Reference |
| MDA-MB-231 | Anti-EGFR-gold nanorod conjugates | Immunofluorescence staining | LC3 | Increased LC3 expression | 120 pM | [38] |
| MDA-MB-231 | Anti-EGFR-gold nanorod conjugates | TEM | --- | Induction of double- or multiple-membrane vesicles | 120 pM | [38] |
| MDA-MB-231 | Anti-EGFR-gold nanorod conjugates | Western blot | Beclin-1 Atg5 p62 LC3 Akt mTOR | Increased Beclin-1, Atg5, LC3-II, and p62 expression Decreased phosphorylation of mTOR and Akt | 120 pM | [38] |
| MDA-MB-231 SUM1315 | Gold-nanoparticles conjugated with Rad6 inhibitor | Western blot | LC3-I LC3-II p62 | Increased conversion of LC3-I to LC3-II in SUM1315 cells Decreased p62 expression in SUM1315 cells No changes in LC3-I/II ratios or expression of p62 protein in MDA-MB-231 cells | 1–5 μM | [40] |
| MDA-MB-231 | Ruthenium-based nucleolipidic nanoaggregates | Phase-contrast microscopy Monodansylcadaverine assay | --- | Increased formation of autophagic vacuoles | 12.1–14.7 µM | [53] |
| In Vivo | ||||||
| Cell Line | NPMBC | Animal (Model)/Analysis from Animal Tissue | Mode of Action/Pathway Involved | Effect Observed | Doses Assayed (Route of Administration) | Reference |
| MDA-MB-231 | Anti-EGFR-gold nanorod conjugates | BALB/c nude mice (xenograft) | --- | Significant decrease in tumor volume compared to control (120 vs. 250 mm3) | 0.5 pmol/g mouse (i.v.) | [38] |
| MDA-MB-231 | Anti-EGFR-gold nanorod conjugates | BALB/c nude mice (xenograft)/Histological analysis | Beclin-1 LC3 | Enhanced beclin-1 and LC3 expression | 0.5 pmol/g mouse (i.v.) | [38] |
| In Vitro | ||||||
| Cell Line | NPMBC | Assay | Mode of Action/Pathway Involved | Effect Observed | Doses Assayed | Reference |
| MDA-MB-231 | Chitosan layered gold nanorods | Transwell assay | --- | Inhibition of cell migration | 50 nM | [43] |
| MDA-MB-231 | Micellar gold nanoparticles conjugated with ZD6474 | Boyden chamber assay | --- | ~90% inhibition of cell invasion and migration | 1 µM | [44] |
| MDA-MB-231 | Copper oxide nanowire conjugated with folic acid | Wound healing assay | --- | 85% inhibition of cell migration | 1.25 µg/mL | [51] |
| MDA-MB-231 | Copper oxide nanowire conjugated with folic acid | Transwell migration assay | --- | 80% inhibition of cell migration | 1.25 µg/mL | [51] |
| MDA-MB-231 | Copper oxide nanowire conjugated with folic acid | Immunocytochemistry | FAK PTEN | Decreased FAK expression Up-regulated PTEN expression | 1.25 µg/mL | [51] |
| MDA-MB-231 | Ruthenium-based complexes | Transwell assay | --- | 90.8% inhibition of cell invasion | 4 µM | [58] |
| MDA-MB-231 | Ruthenium-based complexes | Wound healing assay | --- | 79% inhibition of cell migration | 4 µM | [58] |
| MDA-MB-231 | Ruthenium-based complexes | Adhesion assay | Type I collagen Fibronectin Laminin Vitronectin | 35–70% inhibition of cancer cell adhesion to extracellular matrix proteins | 8 µ | [58] |
| MDA-MB-231 | Ruthenium-based complexes | Wound healing assay | --- | Inhibition of cell migration | 5, 10 and 20 µM | [59] |
| MDA-MB-231 | Ruthenium-based complexes | Transwell assay | --- | 80% inhibition of cell invasion | 20 µM | [59] |
| MDA-MB-231 | Ruthenium-based complexes | Zymography assay | MMP-9 | 60% of inhibition of MMP-9 expression | 20 µM | [59] |
| MDA-MB-231 | Ruthenium-based complexes | Adhesion assay | Type I collagen Fibronectin Laminin Vitronectin | 50–65% inhibition of cancer cell adhesion to type I collagen, fibronectin, laminin, vitronectin | 40 µM | [59] |
| MDA-MB-231 4T1 | Supermagnetic iron oxide nanoparticles conjugated with doxorubicin | Wound healing assay | --- | Minimal closure of the scratch | 520 µM | [62] |
| MDA-MB-231 | Doxorubicin-hyaluronan conjugated iron oxide nanoparticles | Confocal microscopy | Vimentin e-cadherin | Decreased vimentin expression No change in e-cadherin expression | 2 µM | [65] |
| MDA-MB-231 | Doxorubicin-hyaluronan conjugated iron oxide nanoparticles | ELISA | IL-6 IL-10 | Decreased IL-6 secretion No change in secretion of IL-10 | 2 µM | [65] |
| MDA-MB-231 | Doxorubicin-hyaluronan conjugated iron oxide nanoparticles | Steady-Glo luciferase reagent assay | NF-κB | Reduced NF-κB transcriptional activity (~40%) | 2 µM | [65] |
| In Vivo | ||||||
| Cell Line | NPMBC | Animal (Model)/Analysis from Animal Tissue | Mode of Action/Pathway Involved | Effect Observed | Doses Assayed (Route of Administration) | Reference |
| T41 | Iron oxide nanoparticles | BALB/c immune-competent mice (xenograft) | --- | Lower lung metastatic nodules than control (13 vs. 42) | 20 mg iron/kg body weight (i.v.) | [61] |
| SUM159 | Iron oxide nanoparticles | NOD/SCID immune-compromised mice (orthotopic) | --- | Lower incidence of lymph node metastasis than control (25% vs. 62.5%) | 20 mg iron/kg body weight (i.v.) | [61] |
| 4T1 | Supermagnetic iron oxide nanoparticles conjugated with doxorubicin | BALB/c mice (orthotopic) | αvβ3 | Lower metastatic colonies than control (~<1 vs. ~10 at lung respectively; ~1 vs. ~3 at liver respectively) which could be related with the inhibition of αvβ3 | 0.25 mg iron/100 mm3 (i.t.) | [62] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Domínguez-Martís, E.M.; Mosteiro-Miguéns, D.G.; Vigo-Gendre, L.; López-Ares, D.; Freire-Garabal, M.; Núñez-Iglesias, M.J.; Novío, S. Non-Platinum Metal Complexes as Potential Anti-Triple Negative Breast Cancer Agents. Crystals 2018, 8, 369. https://doi.org/10.3390/cryst8100369
Domínguez-Martís EM, Mosteiro-Miguéns DG, Vigo-Gendre L, López-Ares D, Freire-Garabal M, Núñez-Iglesias MJ, Novío S. Non-Platinum Metal Complexes as Potential Anti-Triple Negative Breast Cancer Agents. Crystals. 2018; 8(10):369. https://doi.org/10.3390/cryst8100369
Chicago/Turabian StyleDomínguez-Martís, Eva María, Diego Gabriel Mosteiro-Miguéns, Lucía Vigo-Gendre, David López-Ares, Manuel Freire-Garabal, María Jesús Núñez-Iglesias, and Silvia Novío. 2018. "Non-Platinum Metal Complexes as Potential Anti-Triple Negative Breast Cancer Agents" Crystals 8, no. 10: 369. https://doi.org/10.3390/cryst8100369