Combined Cross-Linked Enzyme Aggregates as Biocatalysts


1
School of Pharmacy, United Pharmaceutical Institute of Jiangsu University and Shandong Tianzhilvye Biotechnology Co. Ltd., Jiangsu University, Zhenjiang 212013, China
2
College of Petroleum and Chemical Engineering, Qinzhou University, Qinzhou 535011, China
*
Authors to whom correspondence should be addressed.
Catalysts 2018, 8(10), 460; https://doi.org/10.3390/catal8100460
Submission received: 17 September 2018
/
Revised: 11 October 2018
/
Accepted: 14 October 2018
/
Published: 17 October 2018
(This article belongs to the Special Issue Recent Advances in Biocatalysis and Metabolic Engineering for Biomanufacturing)
Abstract
:Enzymes are efficient biocatalysts providing an important tool in many industrial biocatalytic processes. Currently, the immobilized enzymes prepared by the cross-linked enzyme aggregates (CLEAs) have drawn much attention due to their simple preparation and high catalytic efficiency. Combined cross-linked enzyme aggregates (combi-CLEAs) including multiple enzymes have significant advantages for practical applications. In this review, the conditions or factors for the preparation of combi-CLEAs such as the proportion of enzymes, the type of cross-linker, and coupling temperature were discussed based on the reaction mechanism. The recent applications of combi-CLEAs were also reviewed.
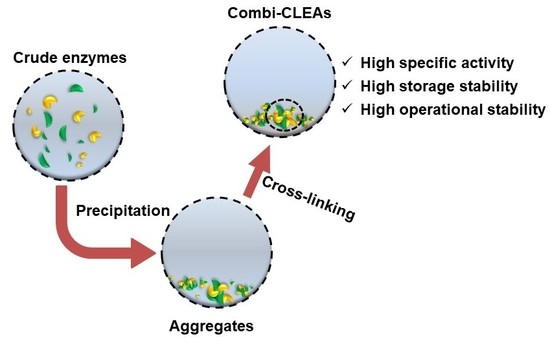
1. Introduction
With a high catalytic efficiency, chemo-, region-, and stereoselectivities, enzyme-mediated biocatalytic reactions have a wide range of applications in the fermentation, chemical, food industry and environmental management [1,2,3,4,5]. These reactions usually involve one enzyme or multi-enzymes as catalysts. Compared with single enzymes, multi-enzymes can produce more valuable products although the composition and preparation are complicated. A lot of research has employed soluble enzymes to catalyze reactions [6,7]. The poor operational stability and difficult enzyme separation hampered the development of multiple enzymes as biocatalysts [8]. These limitations could be overcome by membrane reactors [9]. However, the applications of such membrane bioreactors are relatively expensive because of membrane fouling [10], high energy consumption [11], and difficulty to separate macromolecular substrate and enzymes from the reaction mixture. To lower the cost of biocatalysts, the most practical option is enzyme immobilization [12,13].
Enzyme immobilization offers a considerable prospect of reusability and increases the enzyme stability, such as the improvement of organic solvents resistance, pH tolerance, and thermal stability [14,15,16,17]. Immobilization of enzymes onto solid carriers is an effective method for stabilizing enzymes because of the superior physical stability and easier recovery [18]. Till today, many carriers including nanomaterials [19,20,21,22,23,24,25,26,27,28,29], magnetic materials [30,31,32,33,34,35], and graphene carriers [36,37] are widely used for drug delivery, food preparation, and the immobilization of enzymes. Most of these carriers are metal particles [38,39,40,41,42], and some of them as are biological macromolecules [43,44,45,46,47]. Unfortunately, extra carrier leads to the dilution of activity of immobilized enzymes because of the introducing of a large portion of the non-catalytic mass. This will result in lower space-time yields, volumetric activity, and productivity in the enzymatic reactions [48,49]. Furthermore, the preparation and modification of carrier significantly increase additional costs of the biocatalysts. To overcome these shortcomings, carrier-free techniques for immobilization of enzymes have received much interest since they were developed.
Carrier-free immobilized enzymes were developed since the middle of the 20th century [48]. As the most typical carrier-free immobilized enzyme, cross-linked enzyme crystal (CLEC) is chemically cross-linked between enzyme crystals [50]. It was firstly reported by Quiocho et al. in 1964 [51]. In their research, carboxypeptidase A crystals were obtained from pure enzyme protein by adjusting the pH of carboxypeptidase A containing 1 M NaCl solution and dialyzing to reduce the salt concentration. CLEC was found to have advantages such as the controllable particle size, high recycling rate and favorable stability, and enhanced tolerance to organic reagent and extreme pH. Because of these advantages, it has been successfully used in chromatography, drug release, chiral synthesis, and other fields. However, it is difficult to prepare CLECs in the industrial scale because of the critical conditions for protein crystallization. These conditions include suitable ionic species and strength, proper seed loading, cooling rate, and temperature [52,53]. The crystal size and shape were affected directly by these factors, and they will determinate the activity of CLEC [54,55,56]. These conditions are hardly controlled simultaneously in industrial production, and the preparation of high purity protein is another burden for production of CLEC. To overcome CLEC’s drawbacks, cross-linked enzyme aggregate (CLEA) was then developed. This technique does not require highly purified enzymes but it could be performed starting from crude enzyme preparations.
2. Combi-CLEA
2.1. Advantages of CLEAs and Combi-CLEAs
Developed from CLEC, CLEA also has all its advantages like a high specific activity, high resistance to pH organic solvent, improved thermal stability. Additionally, it also has a good stability under operation and storage conditions which CLEC process could be achieved by CLEA because there is no dilution caused by the insertion of solid carriers [57,58]. The CLEA displays high resistance to organic solvents, extreme pH, and high temperatures. This is largely due to the fact that immobilization decreases the enzyme flexibility and suppresses enzymes towards unfolding tertiary structures necessary for catalytic activity [59,60,61]. Thus, CLEAs would not lose much activity after several re-utilizations. For example, the thermal stability of CLEAs subtilisin was more improved than that of its soluble counterpart and the immobilized enzyme kept 45% of the catalytic activity after 10 reuses [62]. The investigation of sucrose phosphorylase CLEAs found that the optimum temperature of the immobilized enzyme was increased by 17 °C and had a broader pH range [63]. Epoxide hydrolases CLEAs could protect enzymes from hydrophobic organic solvents as the activity of CLEAs was 21.5% higher than that of the free enzymes in n-hexane [64]. The storage stability of immobilized enzymes was also improved; CLEAs kept about 67% and the free enzymes remained less than 35% of the initial activity. These properties of CLEAs will undoubtedly promote the development of novel processes of industrial applications. Compared with CLEC, the advantages of CLEAs are that the highly pure enzyme is not required in the process of immobilization and CLEA can be prepared directly from a crude enzyme. CLEAs of hydroxynitrile lyase were prepared from crude enzyme precipitation resulting in an obvious improvement of the enzyme stability in acidic conditions [53]. The general procedure for CLEA preparation includes precipitation and cross-linking. Firstly, adding salts or water-miscible organic solvents or nonionic polymers to precipitate the enzyme to obtain physical aggregates from aqueous solutions [54,65]. Physical aggregates keep most of the enzyme activity because the interaction between the molecules of aggregates is non-covalent, which makes the protein form insoluble structures and will not destroy their tertiary structure [59,61]. Precipitation is also a purification process of enzymes. Thus, the CLEA combines purification and immobilization into one operational unit. In principle, CLEAs can be prepared from the crude enzyme directly [66,67]. The simple preparation process of CLEAs indicates CLEAs are much economical and practical in the industry. Cross-linking with bifunctional reagents will make the immobilized enzyme aggregates much more stable due to the fixation of molecular conformation. Therefore, the immobilized enzyme could be used for multiple cycles while maintaining most of its initial activity.
Normally, biocatalytic reactions in vivo involve more than one enzyme and thus keep a high efficiency of the life cycle. To achieve such a goal of the multiple-enzymes catalysis, combined cross-linked enzyme aggregates (combi-CLEAs) were developed based on the CLEAs. Combi-CLEAs include two or more immobilized enzymes that can catalyze cascade or parallel reactions in one reaction system. For example, combi-CLEAs of xylanase, cellulase and β-1,3-glucanase was used to achieve one-pot bioconversion of lignocellulosic biomass to fermentable sugars [8]. To make the combi-CLEAs highly efficient, the reaction conditions such as enzymes ratio, the pH of the reaction medium, and the temperature of preparation should be finely optimized [68,69].
2.2. Factors Influencing CLEAs and Combi-CLEAs Preparation
In order to prepare highly efficient and stable industrial biocatalysts by combi-CLEAs, their preparative conditions are important in affecting catalytic properties that include the activity, stability, and kinetic parameters [69,70]. Many factors such as the ratio of enzymes, the type of precipitants, the crosslinkers, the cross-linking time, and the pH of the reaction system influence the preparation of combi-CLEAs are discussed below (Table 1).
2.2.1. Proportion of Enzymes
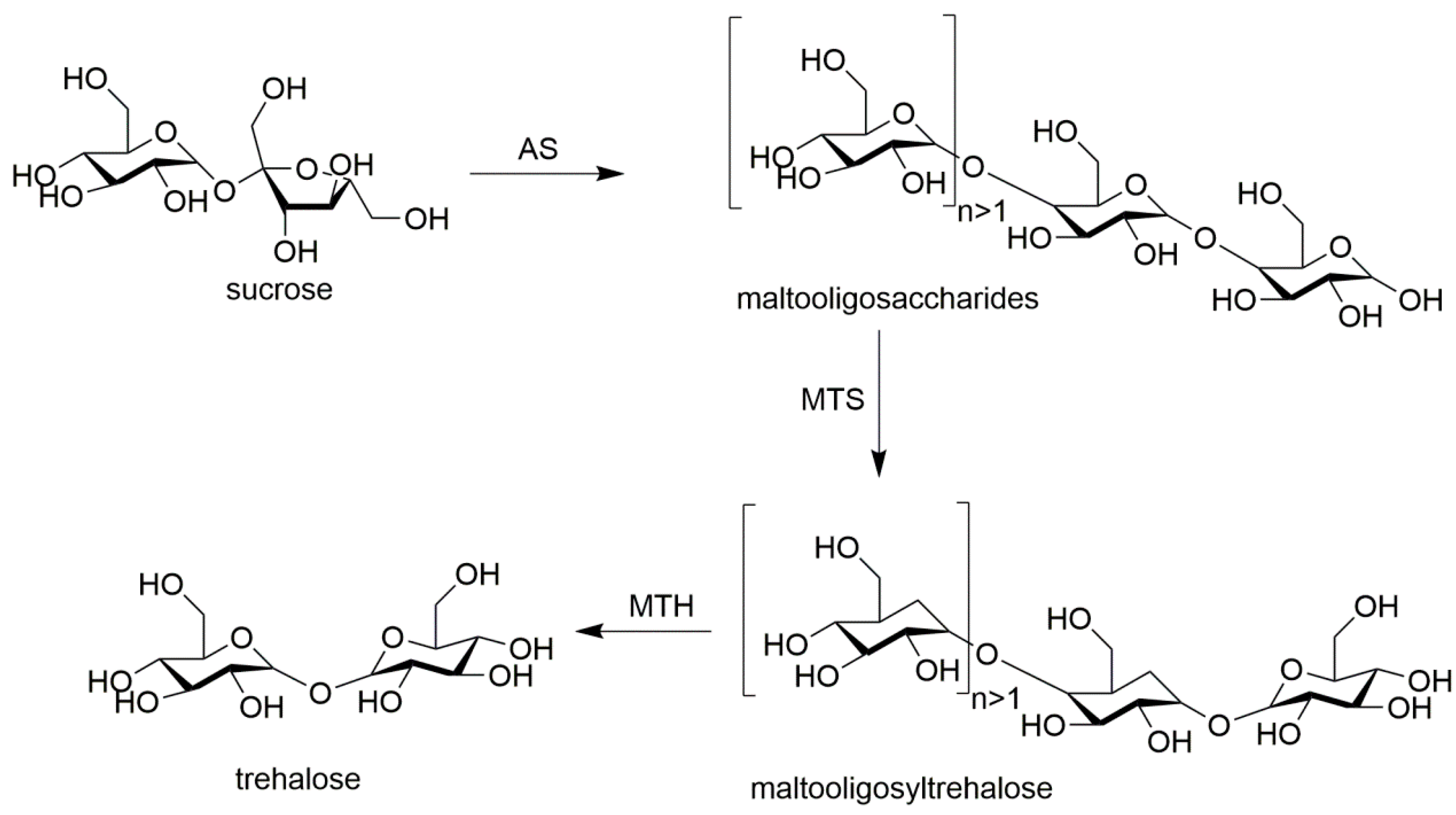
The proportion of every enzyme involved in the fabrication of combi-CLEAs affects the catalytic efficiency. Thus, it is necessary to determine the optimal ratio of enzymes because each enzyme has its own catalytic rate under the reaction conditions. For example, in glucose oxidase (GOD) and versatile peroxidase (VP) CLEAs the glucose can be catalyzed by GOD to D-glucono-δ-lactone and hydrogen peroxide which is the substrate of VP. Excessive GOD catalyzes the production of superfluous hydrogen peroxide that would cause the inactivation of the VP, and a low concentration of GOD would limit the apparent rate of the combi-CLEAs. When the ratio of VP and GOD is 10:7, the maximal apparent rate of combi-CLEAs was observed, which was about 2-fold higher than that without optimization. The reason might be that the produced hydrogen peroxide could be consumed completely by VP [71]. Perfectly setting the right ratio of enzymes in the preparation process of combi-CLEAs is inevitable fundamental in order to make the catalysts economical, have a relative high productivity, with high reaction rates, and more stable in practical applications. Combi-CLEAs of amylosucrase (AS), maltooligosyltrehalose synthase (MTS), and maltooligosyltrehalose trehalohydrolase (MTH) were prepared to produce trehalose from sucrose. AS catalyzes sucrose to form maltooligosaccharides that are substrates for MTS and MTH. The ratio of MTS and MTH was investigated and it was found that the yield of trehalose was similar in the range from 14:1 to 1:14 of MTS:MTH. The results indicated that MTS and MTH were not rate-limiting enzymes in the cascade reactions. The same quantity of MTS and MTH was arbitrarily used in further optimization reactions. The ratios of AS:(MTS and MTH) from 0.25:1 to 8:1 were further investigated. When the ratio was set as 8:1, the trehalose yield was 13 times higher than that of 1:1 [72]. It could be concluded that AS catalyzed the formation of sufficient maltooligosaccharides and long-glucan polymers led to more efficient MTS and MTH catalyzed reactions.
2.2.2. The Precipitants
Adding neutral salts or water-miscible organic solvents or nonionic polymers to free enzymes could induce the physical aggregation and precipitation of protein molecules [73]. These additives could change the hydration state of the enzyme molecule or change the dielectric constant of the solution. Meanwhile, the supramolecular structure is formed in aggregates by non-covalent bonds, the original tertiary structure of enzymes is not destroyed, and the structure may collapse in an aqueous medium. Every enzyme has a unique primary sequence and quaternary structure, so the optimal precipitation condition varies from one enzyme to another. Different precipitant induces distinct conformations of enzyme aggregates which affect the catalytic properties of CLEAs. In the screening of precipitants, the precipitation efficiency of the enzyme is usually determined, then the physical aggregates are re-dissolved in an appropriate buffer and evaluated with their activities [65]. To choose suitable precipitation agents will benefit the preparation of combi-CLEAs. Manganese peroxidase was precipitated in the form of CLEAs using acetone, ammonium sulfate, ethanol, 2-propanol, and tert-butanol, followed by glutaraldehyde cross-linking for 3 h at 4 °C. The results showed that the activity recovery and aggregation yield of acetone were at a maximum, reached 31.26 and 73.46%. The CLEAs recovered 47.57% of the initial activity following cross-linking with glutaraldehyde [74]. The aggregation yield and activity recovery of combi-CLEAs of xylanase and mannanase treated by acetone were both 1.2-folds than those of treated by ammonium sulfate. After cross-linking with 125 mM glutaraldehyde, combi-CLEAs precipitated with 80% acetone retained a higher activity than that precipitated with 80% ammonium sulfate [14]. Four ice-cold organic solvents (acetone, acetonitrile, ethanol, and 2-propanol) were used as the precipitant agents for the preparation of peroxidase CLEAs. The results showed that the activity recoveries of CLEAs were less than 10% when ethanol and 2-propanol were served as precipitants. When acetone was utilized as the precipitant, a 28% activity recovery and 78% aggregation yield of CLEAs were obtained. After incubating at 70 °C for 15 min, the CLEAs precipitated with acetone remained about 37% activity while the free counterpart was found totally inactivated [75]. Compared with acetone and t-butanol, dimethoxyethane was found to be the best precipitant for the preparation of combi-CLEAs including lipase, α-amylase, and phospholipase A2. The preparation parameters included 5 mL of pre-cooling dimethoxyethane, 20 mM of glutaraldehyde, 4 h of cross-linking time and a 4 °C cross-linking temperature. The obtained combi-CLEAs could keep most initial activities after 3 cycles [76]. Generally, the best precipitant for enzymes could be variable, and consequently, the selection of precipitation parameters is a critical step in the preparation of CLEAs with a high recovery of enzyme activity. Additionally, a suitable precipitant should be inexpensive and commercially available. More importantly, it should be aqueous soluble and not react with enzymes and buffers.
2.2.3. The Cross-Linker
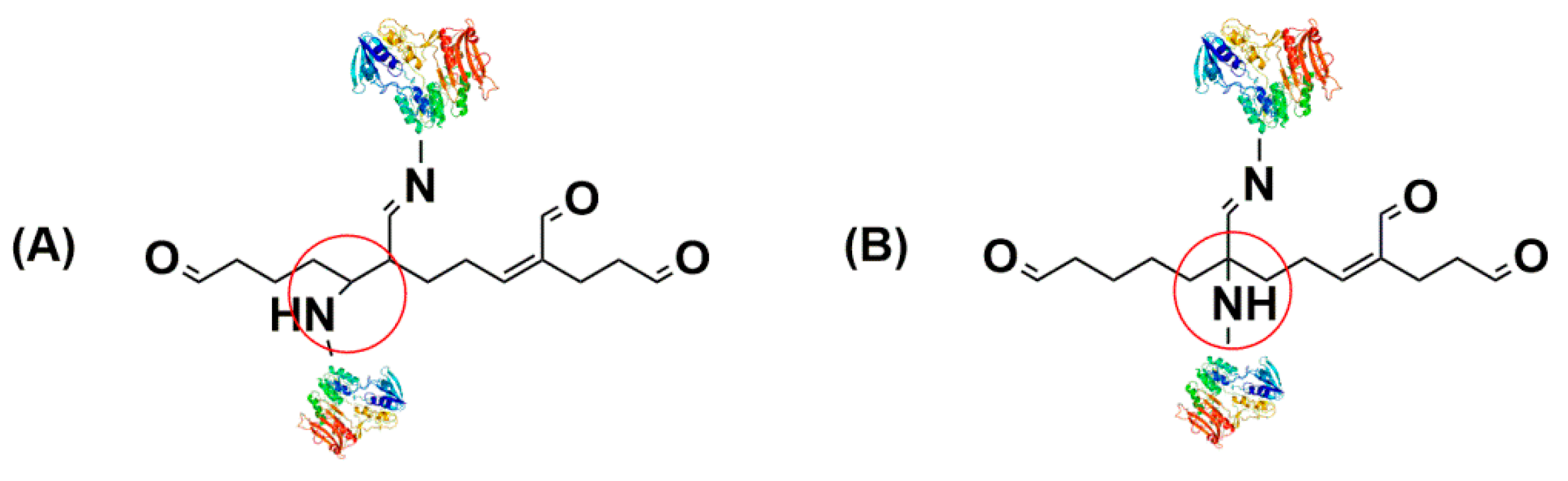
The cross-linker is a bifunctional agent that can covalently link the amino acid residues of enzyme surfaces [77,78]. Glutaraldehyde has been used as a cross-linker for years in the preparation of CLEAs since it is inexpensive and readily available in commercial quantities [79]. CLEAs were prepared by reacting ε-amino groups of lysine residues on the surface of neighboring enzyme molecules with glutaraldehyde to form inter- and intramolecular aldol condensates [80]. The end products obtained under alkaline or acidic conditions were different. Under alkaline conditions, the end product formed a Schiff base with an amino group from one protein molecule and a C–N bond by Michael addition to the β-carbon in the amino group from another protein molecule. Under acidic conditions, the end product formed a Schiff base with an amino group from one protein molecule and a C–N bond by anti-Markownikoff addition to the α-carbon in another amino group from a neighboring protein (Figure 1) [81]. However, there it was reported that alcohol dehydrogenase and nitrilase lost most activities after cross-linking with glutaraldehyde [82]. Glutaraldehyde has a relatively small size, and it could occupy the binding sites of substrates or even block the entry of macromolecular substrates into the catalytic center which could inactivate the enzymes [80]. To prevent this, cross-linkers with large sizes like dextran polyaldehyde were explored. When β-mannanase aggregate was cross-linked with 0.01 mL of 20% glutaraldehyde or 0.1 mL of 3% dextran polyaldehyde for 16 h at 4 °C, it was found that the CLEAs prepared with linear dextran polyaldehyde showed a higher activity to macromolecular substrates, which was 16 times higher than that prepared with glutaraldehyde. It could be explained that such CLEAs contained a porous structure with low steric hindrance [83]. CLEAs of lipase were prepared with the p-benzoquinone as cross-linker and it was observed that this biocatalyst retained 75.18% of their initial activity. Additionally, after heat treatment for 96 h at 50 °C, the residual activity of CLEAs prepared using p-benzoquinone was 5.01-fold higher than that of the CLEAs cross-linked with glutaraldehyde [84]. In all the above-mentioned reports, the reaction mechanism is based on the formation of the covalent bonds between the aldehyde group or ketone of cross-linkers and lysine residues of enzymes to form a Schiff base. Another reaction model was developed with new cross-linkers. As well as lysine amino groups, free carboxyl groups from aspartic and glutamic acids are also potentially reactive and could be used to increase the number of possible cross-linking sites. The free carboxyl groups could be cross-linked with amino-containing polymers to form CLEAs. Several interesting cross-linking agents containing amino groups have been successfully developed. Polyethylenimines was used for the production of a stable Candida rugose lipase CLEAs by the cross-linking of carboxyl groups activated with carbodiimide. Compared with the amino-CLEAs of lipase cross-linked with glutaraldehyde, carboxyl-CLEAs cross-linked with polyethylenimines had a higher activity and thermostability [85]. Laccase CLEAs were prepared by cross-linking the carboxyl groups activated with carbodiimide and the amino groups of chitosan to form amide bonds at 20 °C for 4 h. The CLEAs retained 65% of their initial activity while the free laccase was completely denatured after 12 h of thermal denaturation. The three-dimensional structure of laccase molecules was strengthened by the covalent linking enzyme with chitosan, preventing the unfolding of laccase under heat stress conditions [86]. The improved thermos-stability would make the biocatalyst suitable for applications in industrial processes carried out at high temperatures. The enzymes should be stable in the pH range which chitosan could be dissolved when using chitosan as the linker. Otherwise, the enzyme would be inactivated during the cross-linking reaction. The selection of the cross-linking agent should be based on the number of positive or negative amino acids on the enzyme surface and the location of the catalytic center. Amine-rich precipitators (such as polyethylenimines) should be selected if the negative residues (aspartic or glutamic acid) are more abundant than the positive residues on the protein surface. Conversely, cross-linking agents with aldehyde groups (such as glutaraldehyde) or ketone groups (such as p-benzoquinone) should be chosen to prepare CLEAs with more positive residues on the surface. Otherwise, macromolecular cross-linking agents should be adopted because small size cross-linking molecules might occupy the reactive center and interrupt the normal enzymatic function when the catalytic center located at the protein surface.
The ratio of cross-linker and protein also should be considered because the cross-linker influences the activity and morphology of the resulting CLEAs. As a result, a suitable quantity of cross-linking agent is essential for CLEA. According to the previous reports, the residual activity of CLEA has a maximum value while changing concentrations of cross-linker [87,88,89,90]. Therefore, an appropriate cross-linking agent should be quantitatively used according to the active residues on the enzyme surface. Additionally, a series of proportions of enzyme and cross-linkers should be investigated to obtain the maximum value of enzyme activity retention. As reported in the literature, insufficient bonds were formed at a lower concentration of the crosslinker. On the contrary, a high concentration of cross-linker will be harmful to the CLEA because it can damage the flexibility of the enzyme and result in a change of rigidity. The rigidification of enzymes could prevent the substrate from reaching the active sites and increase the internal mass transfer limitations, consequently decrease the catalytic activity of CLEAs [91,92,93]. Since enzymes might be susceptible to high concentrations of glutaraldehyde, adding bovine serum albumin (BSA), which owns a lot of amine groups, could avoid the excessive cross-linking of enzymes [84,94]. CLEAs of halohydrin dehalogenase were obtained by slowly adding 0.75% (v/v) glutaraldehyde for 6 h under stirring. The activity retention of CLEAs (91.2%) was highest when the cross-linking was carried out with 0.75% (v/v) of glutaraldehyde. When the concentration of glutaraldehyde was increased, the enzyme activity of CLEAs decreased, which might be attributed to mass transfer limitations caused by excessive cross-linking [94].
2.2.4. Effect of Temperature on the Cross-Linking
CLEAs’ preparation depends on the accessibility of the cross-linker with protein residues, and the reaction rate also depends on the reaction temperature [95,96]. At lower temperatures, cross-linking reaction with low reaction rate requires longer reaction time while it will finish in a shorter time at a higher temperature. At higher temperatures, the possibility of enzyme irreversible denaturation tends to increase, but at low temperatures, the enzyme is stable and maintain good catalytic activity [97]. There are several reports that the CLEAs preparation carried out in 4 °C could maintain higher specific activity [74,75,98,99]. It is mainly because enzymes undergo partial unfolding and the heat disrupts the intramolecular bonds in the tertiary and quaternary structure [100].
3. Applications of Combined CLEAs
3.1. Amylosucrase, Maltooligosyltrehalose Synthase, and Maltooligosyltrehalose
Amylosucrase (AS), maltooligosyltrehalose synthase (MTS), and maltooligosyltrehalose trehalohydrolase (MTH) were co-immobilized as combi-CLEAs to one-pot bioconversion of sucrose to trehalose [72] (shown in Figure 2). Physical aggregates obtained by adding 3.6 mL of cold acetone to a 3.6 mL of a mixture solution containing 4 mg of AS, 0.25 mg of MTS, 0.25 mg of MTH, and 9 mg of BSA for 30 min at 4 °C. Then the physical aggregates reacted with 10 mM of glutaraldehyde at 4 °C for 4 h, and the combi-CLEAs were harvested by centrifugation. In this multiple-enzymes catalyzed cascade system, AS is responsible for the bioconversion of sucrose to maltooligosaccharides which are substrates of MTS and transformed into maltooligosyltrehalose. The produced maltooligosyltrehalose was then cleaved by MTH to get trehalose and a shorter maltooligosaccharide. Their experiments showed that combi-CLEA could be used at high substrate concentrations (up to 400 mM). It is found that the activity of combi-CLEAs was well maintained after five cycles. The combi-CLEAs catalyzed multi-step bioconversions into a single reaction system, which brought many advantages such as the low cost of the substrate, handling simplicity, and reusability.
3.2. Hydroxynitrile Lyase and Nitrilase
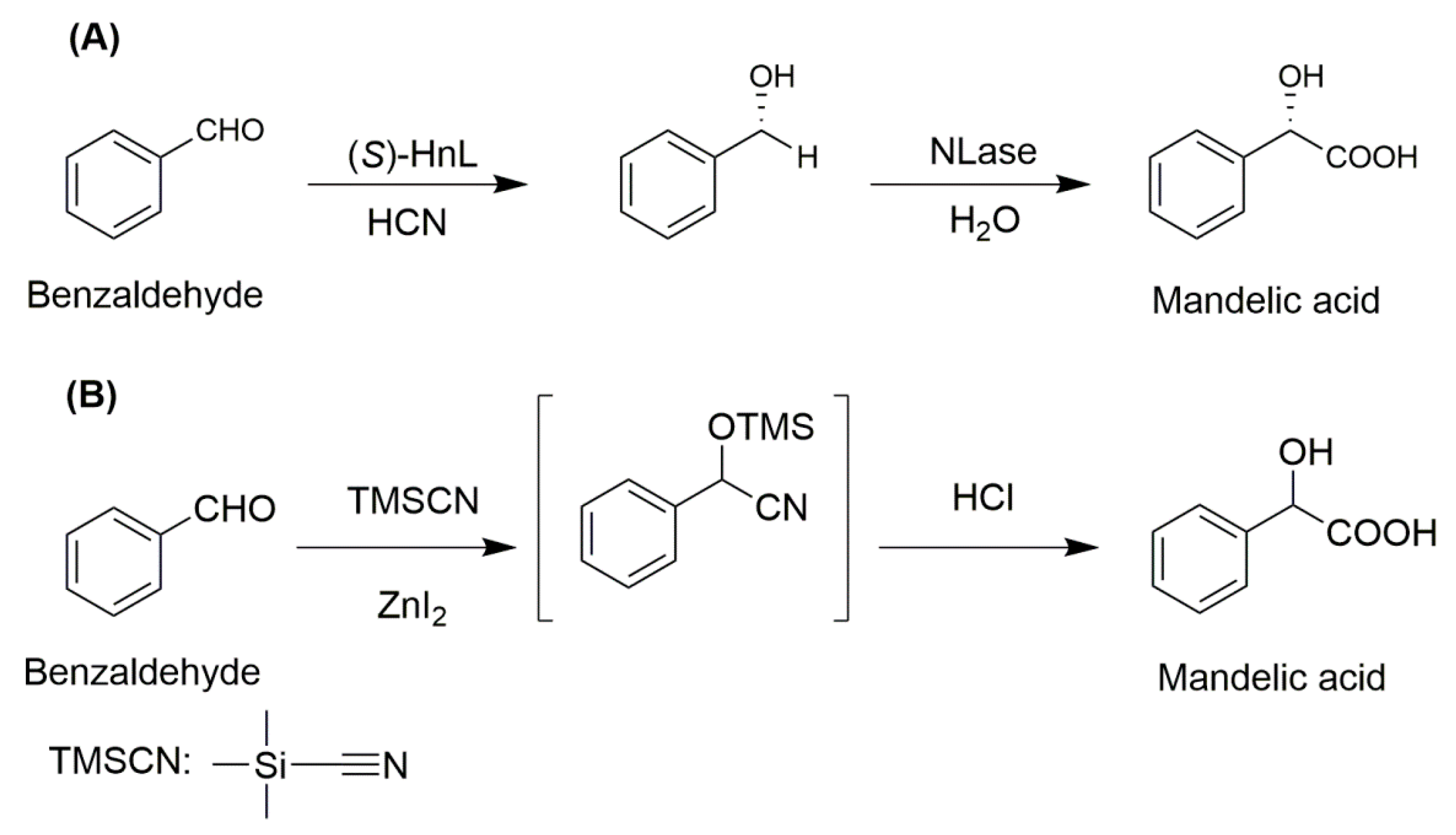
Combi-CLEAs of hydroxynitrile lyase (HnL) and enantioselective nitrilase (NLase) were prepared to synthesize enantiomerically pure (S)-mandelic acid [100]. HnL catalyzed the conversion of aldehyde into the corresponding nitrile which was a substrate of NLase. The enantiomeric excess value of (S)-mandelic acid synthetized by the mixture of HnL-CLEAs and NLase-CLEAs was 94%. Combi-CLEA resulted in a further improvement and 98% enantiomerically pure (S)-mandelic acid was obtained (Figure 3A). It could be explained that the nitrile intermediate was immediately hydrolyzed in the combi-CLEA particles, which suppressed nitrile diffusion into the water phase and possible racemization. Therefore, it is concluded that combi-CLEAs could improve its stereoselectivity. On the contrary, the chemical process for the production of tmandelic acid includes tedious four-steps. Firstly, a mixture comprising benzaldehyde, trimethylsilyl cyanide, and ZnI2 was stirred for 24 h, followed by chemical hydrolysis in the presence of concentrated hydrochloric acid for 24 h. Then the mixture was boiled to remove water and hydrochloric acid. Finally, the racemic mandelic acid was extracted and recrystallized from benzene (Figure 3B) [108]. Compared with the chemical method, the combi-CLEAs mediated synthesis of mandelic acid has significant advantages such as having fewer unit operations, a smaller reactor volume and solvent, less waste generation, good stereo-selectivity, and it is less time-consuming [11,102]. Additionally, there is no need to isolate intermediates which brings many potential economic and environmental advantages. Furthermore, in combi-CLEAs catalyzed cascade reactions, the equilibrium of the reaction could be driven to the target product, thereby improving the catalytic yield [103,104].
3.3. Amylase, Glucoamylase, and Pullulanase
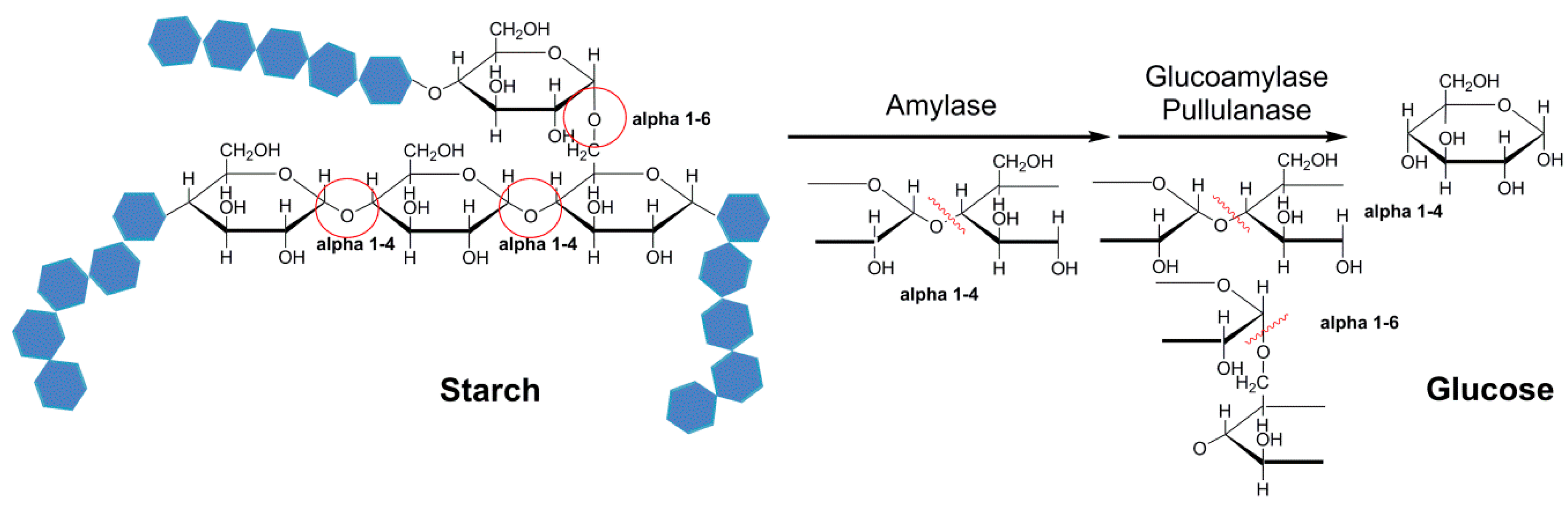
Combi-CLEAs of amylase, glucoamylase, and pullulanase were obtained for one-pot hydrolysis of starch (Figure 4) [101]. In this report, amylase (45 U), glucoamylase (45 U), and pullulanase (15 U) were precipitated with 10 mL of saturated ammonium sulfate for 0.5 h at 4 °C and cross-linked with 40 mM of glutaraldehyde for 4.5 h at 30 °C. The biocatalyst was harvested by centrifugation. The starch conversion of 100% was obtained by combi-CLEAs, whereas a 60% and 40% conversion were obtained by using CLEAs mixture and mixed free enzymes, respectively. The observed different starch conversion attributed to a lower thermal stability of free enzymes and enzymes in separated CLEAs at the reaction temperature compared to those in combi-CLEAs. The reason for a higher rate of starch conversion was that increased proximity of enzymes and reduced the diffusion limitation of the substrate from one enzyme to another by using combi-CLEAs. Moreover, combi-CLEAs showed the highest thermal stability at 55 °C and 75 °C. As immobilized enzymes, the combi-CLEAs have a more evident protection from thermal denaturation and require much more energy to break down the active conformation [38,109].
3.4. l-Arabinosidase and d-Glycosidase
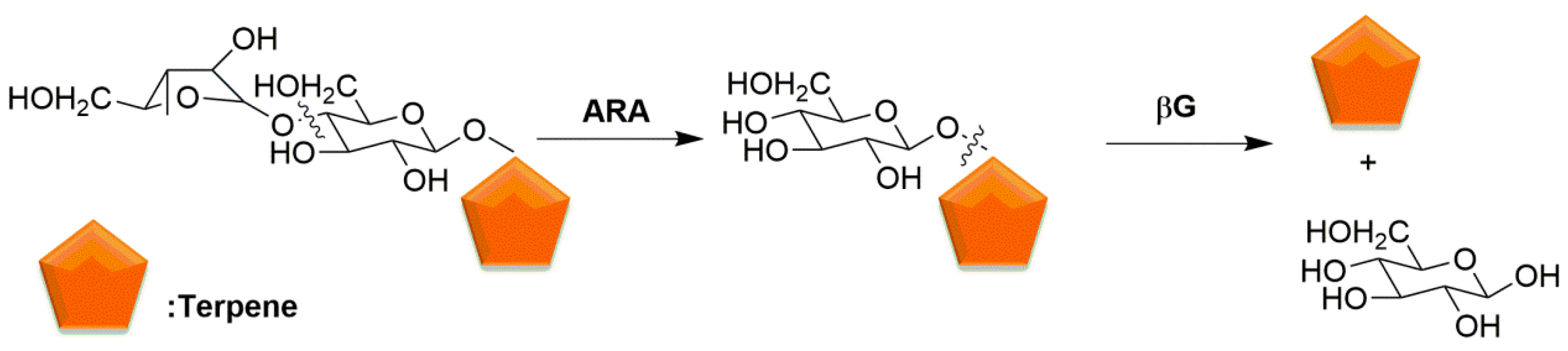
α-l-arabinosidase (ARA) and β-d-glucosidase (βG) were co-immobilized to form combi-CLEAs [102]. Ammonium sulfate (40 mL) was added slowly to the mixture solution (10 mL) containing 20 mg of enzymes and 6.6 mg of BSA under stirring at 4 °C for 30 min. Then 1.06 mg glutaraldehyde was added slowly at 4 °C under stirring for 1 h. The biocatalyst was obtained by centrifugation from suspension. In the hydrolysis of diglycoside, the corresponding sugar and glucoside are released by ARA and then the glucoside is hydrolyzed by βG, liberating the aromatic compound. ARA catalyzes the dissociation of the monoterpenyl β-d-glycoside from its corresponding residual sugar. Then βG catalyzes substrate to release monoterpenes. The released volatile terpenes could enhance wine aroma (Figure 5) [110]. The results showed that the half-lives of βG and ARA in combi-CLEAs were 33.7 and 8.8 times higher than those of the soluble enzymes, respectively. The immobilized biocatalysts were more stable than the soluble enzymes and the higher stability of combi-CLEAs was explained by inter and intramolecular covalent cross-linking [36,111].
3.5. Aminopeptidase N and X-Prolyl-Dipeptidyl Aminopeptidase
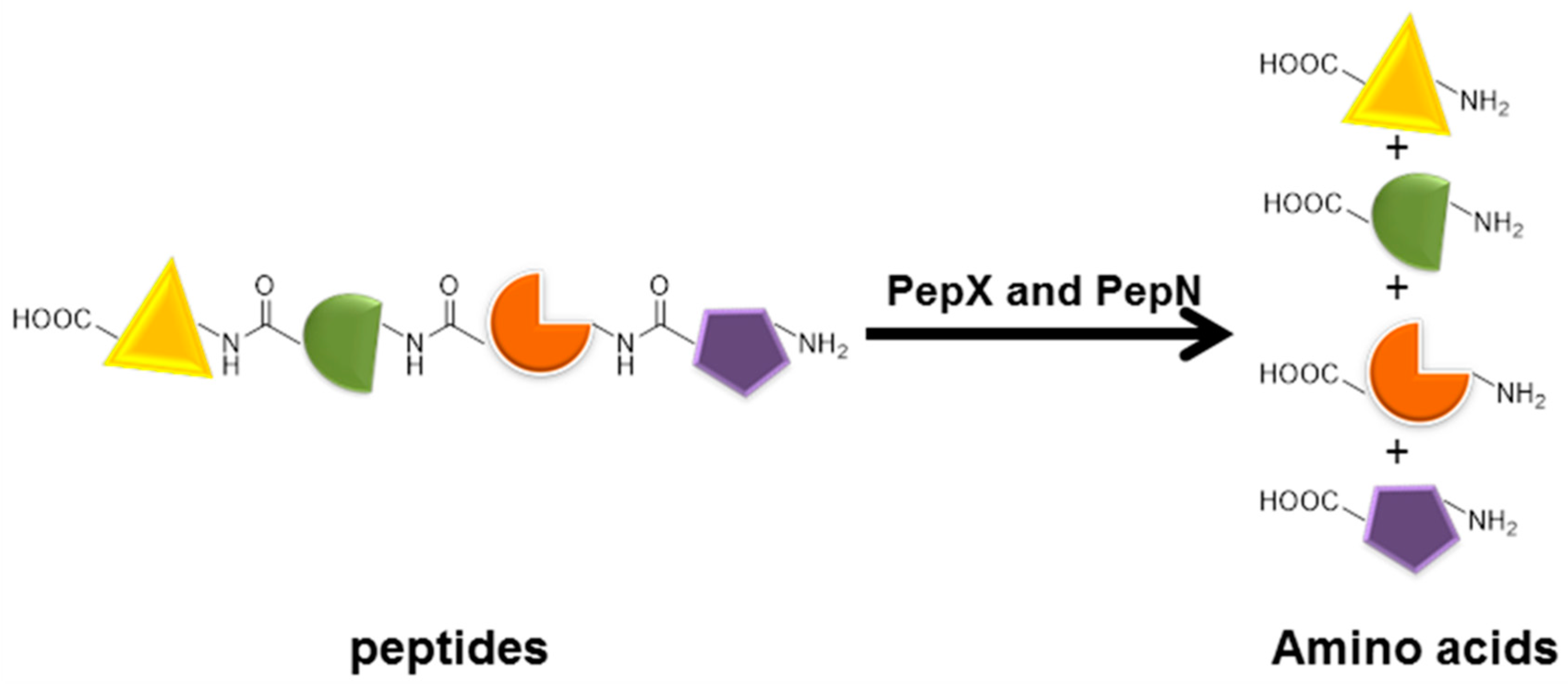
Proline-specific X-prolyl-dipeptidyl aminopeptidase (PepX and aminopeptidase N (PepN) are used in the food industry for cheese-making, baking, and meat tenderization. The enzymatic hydrolysis of proteins can result in a bitter taste. A significant reduction in casein hydrolysate bitterness can be achieved by using PepX combined PepN. A total of 12.5 μg of PepX and 12.5 μg of (PepN) were co-immobilized through precipitation with pre-cooling 4 M ammonium sulfate for 15 min and cross-linked with 50 mM glutaraldehyde. The suspension was centrifuged to separate the combi-CLEAs [103]. The combi-CLEAs were applied to a pre-hydrolyzed casein solution (Figure 6). It could be concluded that, compared with the free PepX and PepN, the relative degree of hydrolysis of the combi-CLEAs of PepX and PepN was increased by approximately 52%. The hydrolysis of food proteins can result in an improved digestibility, modification of sensory quality such as texture or taste, improvement of antioxidant capability or reduction in allergenic compounds. This proved that the combi-CLEAs might have the potential for application in protein hydrolysis.
3.6. Lipase and Protease
Protease and lipase are generally used to hydrolyze proteins and long-chain fatty acid esters, respectively. A novel combi-CLEAs comprising the enzymes lipase and protease from viscera were prepared by Mahmod et al. [104]. The optimum conditions for preparing combi-CLEAs included 65 mM glutaraldehyde, 55% (w/v) ammonium sulfate, and 0.113 mM BSA. Reactions catalyzed by these combi-CLEAs could facilitate the removal of different kinds of food stains and biodiesel production. The stain removal percentage was improved 67.78% when the combi-CLEAs was added to a commercial detergent. In addition, the combi-CLEAs were used to catalyze biodiesel production from vegetable oil with a percentage conversion of 51.7%. The study had presented that combi-CLEAs with a high activity could expand application in the washing process, as well as in catalyzing biodiesel production from vegetable oil.
3.7. Eductase and Glucose Dehydrogenase
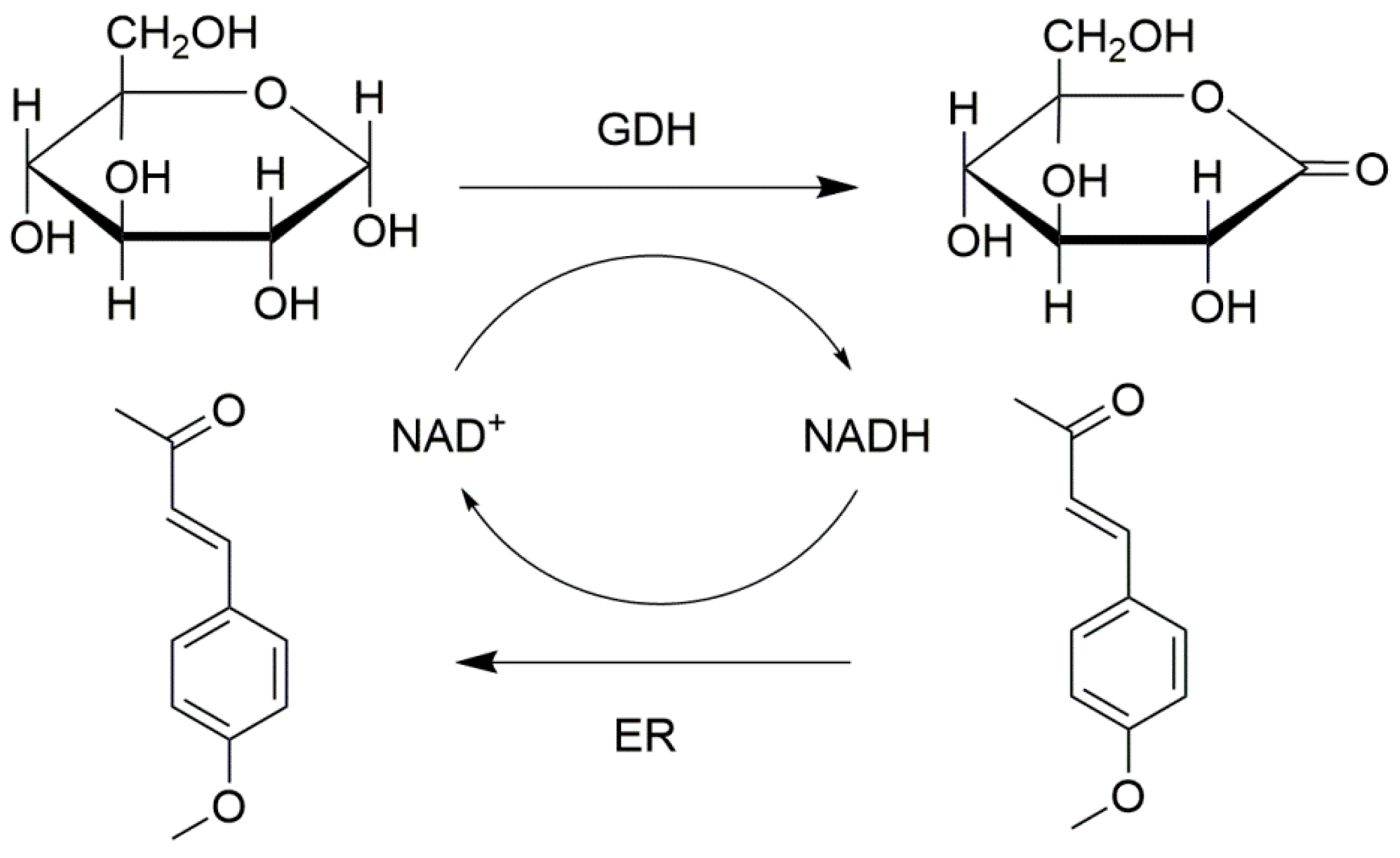
Eductases (ERs) (11 U) and glucose dehydrogenase (GDH) (50 U) were employed to form combi-CLEAs by 1 h of precipitation with 4.0 M ammonium sulfate and 3 h cross-linking with 15% (v/v) glutaraldehyde or oxidized dextran [105]. Then the combi-CLEAs of ER and GDH were collected by centrifugation. The temperature of the whole process was controlled at 4 °C. In this combi-CLEA catalyzed system, ER reduce the C=C bond by using NADH as the cofactor, and GDH can in situ regenerate NADH and make the reaction efficient (Figure 7). Cofactor regeneration is an important issue for the biochemical or pharmaceutical process, and the stability of the biocatalysts is essential for the biotransformation. In order to confirm the thermal stability of the combi-CLEAs, the immobilized biocatalyst and free enzymes were investigated at 50 °C for 8 h. The ERs in combi-CLEAs could keep 65.2% of its initial activity, whereas free ER and GDH had approximately only maintained 9.2% and 19.4% of their initial activity, respectively. The reasons for the improved thermostability of immobilized enzymes might be the suitable microenvironment and the steric constrained structure created by the immobilization of the enzyme molecules. In addition, the activity of ERs in combi-CLEAs could maintain 110% of their initial activity after 14 cycles. The co-immobilized combi-CLEAs were successfully constructed and the stability of the enzymes was improved, which could be implemented in the cofactor regeneration.
3.8. Peroxidase and Glucose Oxidase
Combi-CLEAs of Versatile peroxidase (VP) and glucose oxidase (GOD) were obtained by Taboada et al. [71]. In their report, the detailed conditions were 0.5 mg of VP, 0.35 mg of GOD, 900 μL of 70% polyethylene glycol, 72 mM glutaraldehyde and cross-linking for 21.5 h at 30 °C. The obtained biocatalysts were collected by centrifugation. The combi-CLEAs provided an integrated system in which GOD oxidized glucose in situ produced hydrogen peroxide required by VP. The activity of VP is easily affected by the concentration of hydrogen peroxide. However, the immobilization could increase the stability of VP against hydrogen peroxide. The combi-CLEAs was more stable than free VP, and their results showed that the residual activities of combi-CLEAs and free VP were 50% and 10% at 20 mM hydrogen peroxide, respectively. The improved stability of the immobilized enzymes could be concluded that the technique of cross-linking enzyme can protect the enzyme structure. In batch experiments, four endocrine disrupting chemicals (bisphenol A, nonylphenol, triclosan, 17α-ethinylestradiol, and the hormone 17β-estradiol) were eliminated by either combi-CLEAs or the free enzymes.
3.9. Glucose Oxidase and Horseradish Peroxidase
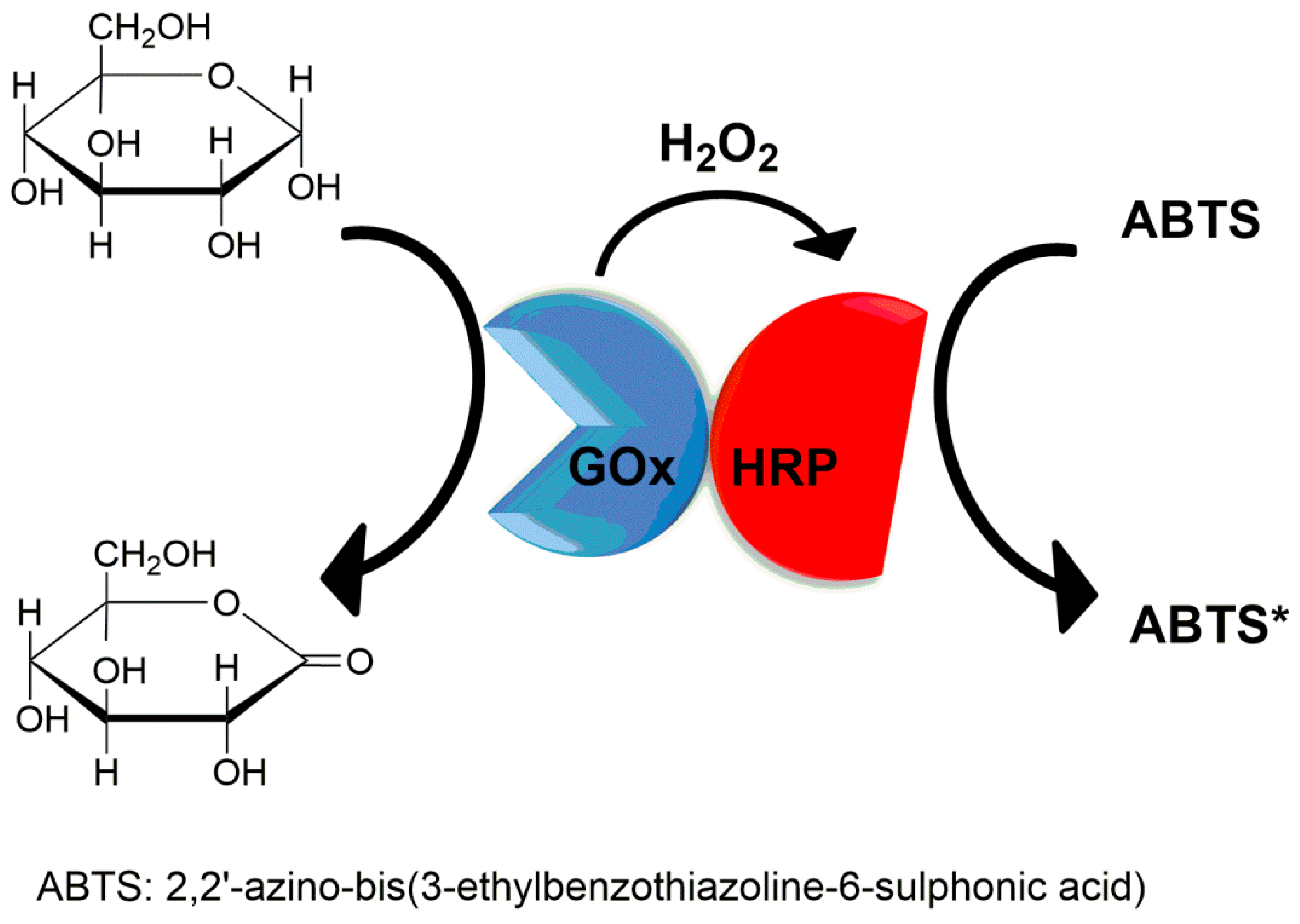
Glucose oxidase (GOx) and horseradish peroxidase (HRP) were combined into CLEAs [106]. This preparation was performed in a self-made millifluidic reactor that consisted of an inner capillary (ID 0.3 mm, OD 0.4 mm) and an outer capillary (ID 0.8 mm, OD 1.0 mm). The inner capillary was fixed by a T-shape connector at the center of the outer capillary. The distance from the confluence point to the outlet was 20 mm. The enzyme mixture flowed through inner capillary while acetonitrile containing 0.5 mM of glutaraldehyde was in the outer capillary. When the weight ratio of GOx/HRP was 150 and the flow rate of the inner and outer capillary was 20 μL/min, the combi-CLEAs could retain 96.5% of free enzyme activity. Glucose is the substrate of GOx and the product hydrogen peroxide is the substrate of HRP (Figure 8). The apparent Km value of combi-CLEA (12.4 ± 0.03 mM) was lower than that of the free enzyme (19.3 ± 0.09 mM). The apparent Vmax value of combi-CLEA (15.8 ± 0.05 μM/min) was closed to that of the free enzyme (15.6 ± 0.09 μM/min). Moreover, the catalytic efficiency (kcat/Km) of combi-CLEA was 1.47 times higher than the free enzyme. These results indicated that combi-CLEA led to a slightly higher reaction rate than free enzymes. The increased reaction rate could be attributed to the in situ fast consumption of hydrogen peroxide by HRP inside the combi-CLEAs.
3.10. Alcohol Dehydrogenase and Glucose Dehydrogenase
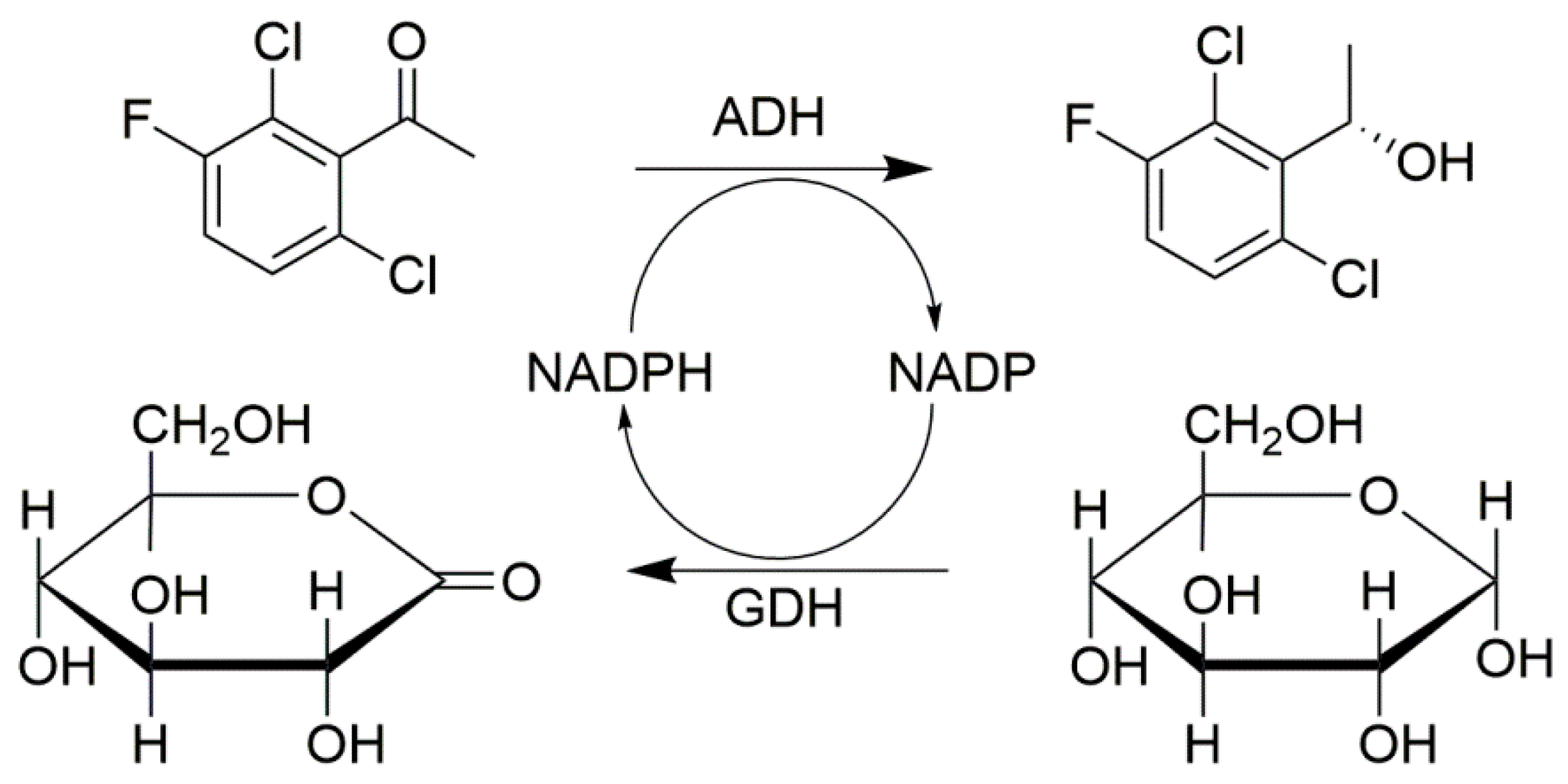
Another example for the cofactor regeneration system is combi-CLEAs of alcohol dehydrogenase (ADH) and glucose dehydrogenase (GDH) [89]. In this work, 90% acetone was added to the crude ADH and GDH solution for 30 min at 4 °C and then cross-linked with 2.5% glutaraldehyde (glutaraldehyde:total protein at 0.5:1) for 1.5 h at ambient temperature. The combi-CLEAs were collected by centrifugation. The obtained immobilized catalysts catalyze the reduction of 1-(2,6-dichloro-3-fluorophenyl) acetophenone and they retained 81.90% (ADH) and 40.29% (GDH) of the initial activity, respectively. The optimum pH was found to be 7 for free enzymes and combi-CLEAs. Meanwhile, combi-CLEAs showed a higher stability under acidic conditions for cofactor regeneration. Since the cofactor recycling will lead to the continuous better acidification of the reaction system, this result especially benefits the reductive coupling reactions of ADH and GDH. This allows the use of these biocatalysts more efficiently in a broader pH region compared to free ADH/GDH (Figure 9).
3.11. Ketoreductase and Glucose Dehydrogenase
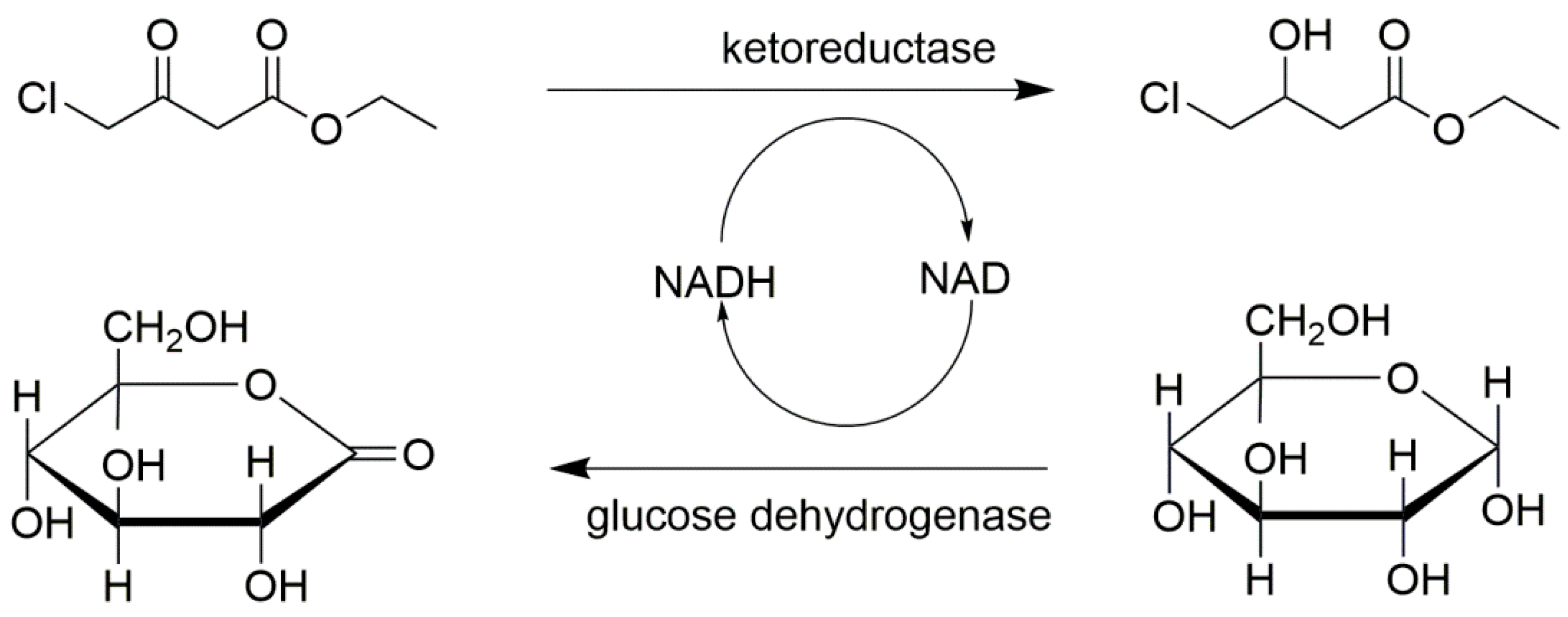
Combi-CLEAs composed of ketoreductase and D-glucose dehydrogenase were prepared according to the work of Ning et al. [107]. They added 1,2-Dimethoxyethane prechilled at 4 °C into the enzyme mixture to a final concentration of 90% (v/v) at 4 °C for 20 min, and the CLEAs were obtained by cross-linking with 0.2% (w/v) glutaraldehyde at 20 °C for 1 h. The whole process was under shaking at 200 rpm. These combi-CLEAs had been demonstrated to be a robust regeneration system for pyridine nucleotide cofactor (Figure 10). The results indicated it is an effective cofactor regeneration system and the optimal substrate concentration in a biphasic system of combi-CLEAs was 300 mM. The improvement of affinity for substrate suggesting that combi-CLEAs had a positive effect on volumetric productivity.
4. Conclusions and Prospects
Combi-CLEA includes multiple enzymes and it is a promising immobilization method without carriers that combined purification and immobilization into one step and do not require highly pure enzymes. However, enzymes from different sources require the optimization of the precipitation and cross-linking parameters. Although the precipitant and cross-linker are the priorities for the preparation of combi-CLEA, all the parameters should be considered carefully because of the interactions among them. Generally, combi-CLEAs can improve enzyme stability and apparent catalytic rate. These will definitely benefit the biotransformation in industrial applications, especially in the cofactor regeneration systems. When combi-CLEAs are used for cofactor regeneration, some advantages such as a high selectivity, high efficiency, and low diffusional limitation are obvious. Due to most oxidoreductases require stoichiometric amounts of expensive cofactors such as NAD+, the Combi-CLEAs of oxidoreductases or can ensure the reuse of cofactors to reduce the cost. It is a robust regeneration system which could be prepared in a simple, rapid, and economical way.
With the development of biomolecular engineering, tailor-made enzymes with specific modifications are produced in a bench scale. Chemical or biological modifications of enzyme surface residues with the specific incorporation of unnatural or functional amino acids will make the preparation of combi-CLEA easier. Some unnatural amino acids with azide (e.g., l-azidohomoalanine, p-azidophenylalanine, and 5-azidopentanoic acid) and alkyne (e.g., l-homopropargylglycine) could be used for bioorthogonal reactions with “click” linking, so the cross-linking reaction could be achieved easily by coupling both the residue-specific and site-specific incorporation of unnatural amino acids into a single protein or different proteins. As a consequence, these enzymes could be conjugated directly to form combi-CLEAs without a cross-linker. If so, combi-CLEAs will be applied to a wide field of cascade processes of unprecedented complexity, efficiency, and elegance.
Author Contributions
M.-Q.X., and Y.-W.Z. prepared the manuscript. M.-Q.X., S.-S.W., and L.-N.L. collected the literature. J.G. and Y.-W.Z. aided in writing and revised the manuscript.
Funding
This research was funded by Guangxi Natural Science Foundation, grant number 2016GXNSFCA380011. The APC was funded by “Seagull Plan” from Qinzhou city.
Conflicts of Interest
All authors declare no conflict of interest.
References
- Wen, Y.; Xu, L.; Chen, F.; Gao, J.; Li, J.; Hu, L.; Li, J. Discovery of a novel inhibitor of NAD(P)+-dependent malic enzyme (ME2) by high-throughput screening. Acta Pharmacol. Sin. 2014, 35, 674–684. [Google Scholar] [CrossRef] [PubMed]
- Feng, W.; Zhao, T.; Zhou, Y.; Li, F.; Zou, Y.; Bai, S.; Wang, W.; Yang, L.; Wu, X. Optimization of enzyme-assisted extraction and characterization of collagen from Chinese sturgeon (Acipenser sturio Linnaeus) skin. Pharmacogn. Mag. 2013, 9, 32–37. [Google Scholar] [CrossRef]
- Yang, H.; Shen, Y.; Xu, Y.; Maqueda, A.S.; Zheng, J.; Wu, Q.; Tam, J.P. A novel strategy for the discrimination of gelatinous Chinese medicines based on enzymatic digestion followed by nano-flow liquid chromatography in tandem with orbitrap mass spectrum detection. Int. J. Nanomed. 2015, 10, 4947–4955. [Google Scholar] [CrossRef] [PubMed]
- Magne, V.; Amounas, M.; Innocent, C.; Dejean, E.; Seta, P. Enzyme textile for removal of urea with coupling process: Enzymatic reaction and electrodialysis. Desalination 2018, 144, 163–166. [Google Scholar] [CrossRef]
- Liu, L.; Zhang, R.; Deng, Y.; Zhang, Y.; Xiao, J.; Huang, F.; Wei, W.; Zhang, M. Fermentation and complex enzyme hydrolysis enhance total phenolics and antioxidant activity of aqueous solution from rice bran pretreated by steaming with α-amylase. Food Chem. 2017, 221, 636–643. [Google Scholar] [CrossRef] [PubMed]
- Shah, S.; Agera, R.; Sharma, P.; Sunder, A.V.; Bajwa, H.; James, H.M.; Gaikaiwari, R.P.; Wangikar, P.P. Development of biotransformation process for asymmetric reduction with novel anti-Prelog NADH-dependent alcohol dehydrogenases. Process Biochem. 2018. [Google Scholar] [CrossRef]
- Zhang, L.; Singh, R.; Sivakumar, D.; Guo, Z.; Li, J.; Chen, F.; He, Y.; Guan, X.; Kang, Y.C.; Lee, J.K. An artificial synthetic pathway for acetoin, 2,3-butanediol, and 2-butanol production from ethanol using cell free multi-enzyme catalysis. Green Chem. 2017, 221, 636–643. [Google Scholar] [CrossRef]
- Touahar, I.E.; Haroune, L.; Ba, S.; Bellenger, J.-P.; Cabana, H. Characterization of combined cross-linked enzyme aggregates from laccase, versatile peroxidase and glucose oxidase, and their utilization for the elimination of pharmaceuticals. Sci. Total Environ. 2014, 481, 90–99. [Google Scholar] [CrossRef] [PubMed]
- Lloret, L.; Eibes, G.; Feijoo, G.; Moreira, M.T.; Lema, J.M. Degradation of estrogens by laccase from Myceliophthora thermophila in fed-batch and enzymatic membrane reactors. J. Hazard. Mater. 2012, 175–183. [Google Scholar] [CrossRef] [PubMed]
- Le-Clech, P.; Chen, V.; Fane, T.A.G. Fouling in membrane bioreactors used in wastewater treatment. J. Membr. Sci. 2006, 284, 17–53. [Google Scholar] [CrossRef]
- Sheldon, R.A. E factors, green chemistry and catalysis: An odyssey. Chem. Commun. 2008, 39, 3352–3365. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, M.Y.; Jiang, X.P.; Ling, X.M.; Xu, M.Q.; Zhu, Y.H.; Zhang, Y.W. Immobilization of glycerol dehydrogenase and NADH oxidase for enzymatic synthesis of 1,3-dihydroxyacetone with in situ cofactor regeneration. J. Chem. Technol. Biotechnol. 2018, 93, 2351–2358. [Google Scholar] [CrossRef]
- Shi, Y.; Liu, W.; Tao, Q.L.; Jiang, X.P.; Liu, C.H.; Zeng, S.; Zhang, Y.W. Immobilization of Lipase by Adsorption Onto Magnetic Nanoparticles in Organic Solvents. J. Nanosci. Nanotechnol. 2016, 16, 601–607. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, A.; Pletschke, B.I. Strategic optimization of xylanase–mannanase combi-CLEAs for synergistic and efficient hydrolysis of complex lignocellulosic substrates. J. Mol. Catal. B Enzym. 2015, 115, 140–150. [Google Scholar] [CrossRef]
- Stepankova, V.; Bidmanova, S.; Koudelakova, T.; Prokop, Z.; Chaloupkova, R.; Damborsky, J. Strategies for Stabilization of Enzymes in Organic Solvents. ACS Catal. 2013, 3, 2823–2836. [Google Scholar] [CrossRef]
- Guzik, U.; Hupertkocurek, K.; Wojcieszyńska, D. Immobilization as a strategy for improving enzyme properties-application to oxidoreductases. Molecules 2014, 19, 8995–9018. [Google Scholar] [CrossRef] [PubMed]
- Hwang, E.T.; Gu, M.B. Enzyme stabilization by nano/microsized hybrid materials. Eng. Life Sci. 2013, 13, 49–61. [Google Scholar] [CrossRef]
- Wang, F.; Chen, L.; Zhang, D.; Jiang, S.; Shi, K.; Huang, Y.; Li, R.; Xu, Q. Methazolamide-loaded solid lipid nanoparticles modified with low-molecular weight chitosan for the treatment of glaucoma: Vitro and vivo study. J. Drug Target. 2014, 22, 849–858. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Yuan, L.; Zhou, L.; Zhang, Z.; Cao, W.; Wu, Q. Effect of cell-penetrating peptide-coated nanostructured lipid carriers on the oral absorption of tripterine. Int. J. Nanomed. 2012, 7, 4581–4591. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Qi, W.; Su, R.; He, Z. Advances in carrier-bound and carrier-free immobilized nanobiocatalysts. Chem. Eng. Sci. 2015, 135, 21–32. [Google Scholar] [CrossRef]
- Xie, M.; Xu, Y.; Shen, H.; Shen, S.; Ge, Y.; Xie, J. Negative-charge-functionalized mesoporous silica nanoparticles as drug vehicles targeting hepatocellular carcinoma. Int. J. Pharm. 2014, 474, 223–231. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Shi, S.; Cao, J.; Ji, L.; He, Y.; Xi, J. Preparation and evaluation of a novel bioactive glass/lysozyme/PLGA composite microsphere. Drug Dev. Ind. Pharm. 2015, 41, 458–463. [Google Scholar] [CrossRef] [PubMed]
- Shi, F.; Zhao, Y.; Firempong, C.K.; Xu, X. Preparation, characterization and pharmacokinetic studies of linalool-loaded nanostructured lipid carriers. Pharm. Biol. 2016, 54, 2320–2328. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Wang, J.-J.; Chen, X.-L.; Du, L.; Li, F. Quercetin-loaded freeze-dried nanomicelles: Improving absorption and anti-glioma efficiency in vitro and in vivo. J. Control. Release 2016, 235, 276–290. [Google Scholar] [CrossRef] [PubMed]
- Zhu, T.; Tao, Z.; Jia, L.; Luo, Y.-F.; Xu, J.; Chen, R.-H.; Ge, Z.-J.; Ma, T.-L.; Chen, H. Multifunctional nanocomposite based on halloysite nanotubes for efficient luminescent bioimaging and magnetic resonance imaging. Int. J. Nanomed. 2016, 11, 4765–4776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rui, M.; Xin, Y.; Li, R.; Ge, Y.; Feng, C.; Xu, X. Targeted Biomimetic Nanoparticles for Synergistic Combination Chemotherapy of Paclitaxel and Doxorubicin. Mol. Pharm. 2017, 14, 107–123. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Firempong, C.K.; Wang, Y.; Xu, W.; Wang, M.; Cao, X.; Zhu, Y.; Tong, S.; Yu, J.; Xu, X. Ergosterol-loaded poly(lactide-co-glycolide) nanoparticles with enhanced in vitro antitumor activity and oral bioavailability. Acta Pharmacol. Sin. 2017, 37, 834–844. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.-S.; Ngai, S.-C.; Goh, B.-H.; Chan, K.-G.; Lee, L.-H.; Chuah, L.-H. Anticancer Activities of Surfactin and Potential Application of Nanotechnology Assisted Surfactin Delivery. Front. Pharmacol. 2017, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, X.; Deng, W.W.; Fu, M.; Wang, L.; Tong, S.S.; Wei, Y.W.; Xu, Y.; Su, W.Y.; Xu, X.M.; Yu, J.N. In vitro release and in vitro–in vivo correlation for silybin meglumine incorporated into hollow-type mesoporous silica nanoparticles. Int. J. Nanomed. 2012, 753. [Google Scholar] [CrossRef]
- Peng, W.; Jiang, X.; Zhu, Y.; Omari-Siaw, E.; Deng, W.; Yu, J.; Xu, X.; Zhang, W. Oral delivery of capsaicin using MPEG-PCL nanoparticles. Acta Pharmacol. Sin. 2015, 36, 139–148. [Google Scholar] [CrossRef] [PubMed]
- Shen, S.; Wu, L.; Liu, J.; Xie, M.; Shen, H.; Qi, X.; Yan, Y.; Ge, Y.; Jin, Y. Core–shell structured Fe3O4@TiO2-doxorubicin nanoparticles for targeted chemo-sonodynamic therapy of cancer. Int. J. Pharm. 2015, 486, 380–388. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Zhou, F.; Zhang, X.-Y.; Li, Y.; Wang, X.-Y.; Xu, X.-M.; Zhang, Y.-W. Preparation of Magnetic Fe3O4@SiO2 Nanoparticles for Immobilization of Lipase. J. Nanosci. Nanotechnol. 2014, 14, 3068–3072. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wang, X.-Y.; Jiang, X.-P.; Ye, J.-J.; Zhang, Y.-W.; Zhang, X.-Y. Fabrication of graphene oxide decorated with Fe3O4@SiO2 for immobilization of cellulase. J. Nanopart. Res. 2015, 17, 8. [Google Scholar] [CrossRef]
- Wang, X.-Y.; Jiang, X.-P.; Li, Y.; Zeng, S.; Zhang, Y.-W. Preparation Fe3O4@chitosan magnetic particles for covalent immobilization of lipase from Thermomyces lanuginosus. Int. J. Biol. Macromol. 2015, 75, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Ling, X.-M.; Wang, X.-Y.; Ma, P.; Yang, Y.; Qin, J.-M.; Zhang, X.-J.; Zhang, Y.-W. Covalent Immobilization of Penicillin G Acylase onto Fe3O4@Chitosan Magnetic Nanoparticles. J. Microbiol. Biotechnol. 2016, 26, 829–836. [Google Scholar] [CrossRef] [PubMed]
- Tao, Q.-L.; Li, Y.; Shi, Y.; Liu, R.-J.; Zhang, Y.-W.; Guo, J. Application of Molecular Imprinted Magnetic Fe3O4@SiO2 Nanoparticles for Selective Immobilization of Cellulase. J. Nanosci. Nanotechnol. 2016, 16, 6055–6060. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.H.; Li, X.Q.; Jiang, X.P.; Zhuang, M.Y.; Zhang, J.X.; Bao, C.H.; Zhang, Y.W. Preparation of Functionalized Graphene Oxide Nanocomposites for Covalent Immobilization of NADH Oxidase. Nanosci. Nanotechnol. Lett. 2016, 8, 164–167. [Google Scholar] [CrossRef]
- Gao, J.; Lu, C.-L.; Wang, Y.; Wang, S.-S.; Shen, J.-J.; Zhang, J.-X.; Zhang, Y.-W. Rapid Immobilization of Cellulase onto Graphene Oxide with a Hydrophobic Spacer. Catalysts 2018, 8, 180. [Google Scholar] [CrossRef]
- Jiao, Z.; Chen, Y.; Wan, Y.; Zhang, H. Anticancer efficacy enhancement and attenuation of side effects of doxorubicin with titanium dioxide nanoparticles. Int. J. Nanomed. 2011, 6, 2321–2326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, F.; Hu, K.; Yu, H.; Zhou, L.; Song, L.; Zhang, Y.; Shan, X.; Liu, J.; Gu, N. A Functional Iron Oxide Nanoparticles Modified with PLA-PEG-DG as Tumor-Targeted MRI Contrast Agent. Pharm. Res. 2017, 34, 1683–1692. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L.; Geng, L.; Ge, L.; Yu, P.; Duan, X.; Chen, J.; Chang, Y. Effect of iron liposomes on anemia of inflammation. Int. J. Pharm. 2013, 454, 82–89. [Google Scholar] [CrossRef] [PubMed]
- Du, F.; Lou, J.; Jiang, R.; Fang, Z.; Zhao, X.; Niu, Y.; Zou, S.; Zhang, M.; Gong, A.; Wu, C. Hyaluronic acid-functionalized bismuth oxide nanoparticles for computed tomography imaging-guided radiotherapy of tumor. Int. J. Nanomed. 2017, 12, 5973–5992. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.S.; Morsy, R.; Elzawawy, N.A.; Fareed, M.F.; Bedaiwy, M.Y. Synthesized zinc peroxide nanoparticles (ZnO2-NPs): A novel antimicrobial, anti-elastase, anti-keratinase, and anti-inflammatory approach toward polymicrobial burn wounds. Int. J. Nanomed. 2017, 12, 6059–6073. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Yuan, L.; Congyan, L.; Zhang, Z.; Zhou, L.; Qu, D. Antitumor activity of tripterine via cell-penetrating peptide-coated nanostructured lipid carriers in a prostate cancer model. Int. J. Nanomed. 2013, 4339–4350. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Huang, J.; Jiang, S.; Liu, Z.; Gu, W.; Yu, H.; Li, Y. Porous starch based self-assembled nano-delivery system improves the oral absorption of lipophilic drug. Int. J. Pharm. 2013, 444, 162–168. [Google Scholar] [CrossRef] [PubMed]
- Rui, M.; Qu, Y.; Gao, T.; Ge, Y.; Feng, C.; Xu, X. Simultaneous delivery of anti-miR21 with doxorubicin prodrug by mimetic lipoprotein nanoparticles for synergistic effect against drug resistance in cancer cells. Int. J. Nanomed. 2017, 12, 217–237. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Dong, W.F.; Zhou, J.Y.; Xu, X.M.; Li, F.Q. Triggering effect of N-acetylglucosamine on retarded drug release from a lectin-anchored chitosan nanoparticles-in-microparticles system. Int. J. Pharm. 2013, 449, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.; Lv, L.; Sheldon, R.A. Immobilised enzymes: Carrier-bound or carrier-free? Curr. Opin. Biotechnol. 2003, 14, 387–394. [Google Scholar] [CrossRef]
- Tischer, W.; Kasche, V. Immobilized enzymes: Crystals or carriers? Trends Biotechnol. 1999, 17, 326–335. [Google Scholar] [CrossRef]
- Jegan Roy, J.; Emilia Abraham, T. Strategies in Making Cross-Linked Enzyme Crystals. Chem. Rev. 2004, 104, 3705–3722. [Google Scholar] [CrossRef] [PubMed]
- Quiocho, F.A.; Richards, F.M. Intermolecular cross linking of a protein in the crystallinestate: Carboxypeptidasw-A. Proc. Natl. Acad. Sci. USA 1964, 52, 833–839. [Google Scholar] [CrossRef] [PubMed]
- Kwon, J.S.; Nayhouse, M.; Christofides, P.D.; Orkoulas, G. Protein Crystal Shape and Size Control in Batch Crystallization: Comparing Model Predictive Control with Conventional Operating Policies. Ind. Eng. Chem. Res. 2014, 53, 5002–5014. [Google Scholar] [CrossRef]
- Liu, J.J.; Ma, C.Y.; Hu, Y.D.; Wang, X.Z. Effect of seed loading and cooling rate on crystal size and shape distributions in protein crystallization—A study using morphological population balance simulation. Comput. Chem. Eng. 2010, 34, 1945–1952. [Google Scholar] [CrossRef]
- Velascolozano, S.; Lópezgallego, F.; Mateosdíaz, J.C.; Favelatorres, E. Cross-linked enzyme aggregates (CLEA) in enzymeimprovement—A review. Biocatalysis 2016, 1, 166–177. [Google Scholar] [CrossRef]
- Clair, N.L.S.; Navia, M.A. Cross-linked enzyme crystals as robust biocatalysts. J. Am. Chem. Soc. 1992, 114, 7314–7316. [Google Scholar]
- Zelinski, T.; Waldmann, H. Cross-Linked Enzyme Crystals(CLECs): Efficient and Stable Biocatalysts for Preparative Organic Chemistry. Angew. Chem. Int. Ed. Engl. 1997, 36, 722–724. [Google Scholar] [CrossRef]
- Roessl, U.; Nahálka, J.; Nidetzky, B. Carrier-free immobilized enzymes for biocatalysis. Biotechnol. Lett. 2010, 32, 341–350. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Guo, Y.L.; Chen, D.W.; Peng, C.; Yan, Y.J. Conformation and Activity of Sol-Gels Encapsulated Cross-Linked Enzyme Aggregates of Lipase from Burkholderia cepacia. Adv. Mater. Res. 2011, 291–294, 614–620. [Google Scholar] [CrossRef]
- Goetze, D.; Foletto, E.F.; da Silva, H.B.; Silveira, V.C.C.; Dal Magro, L.; Rodrigues, R.C. Effect of feather meal as proteic feeder on combi-CLEAs preparation for grape juice clarification. Process Biochem. 2017, 62, 122–127. [Google Scholar] [CrossRef]
- Min, H.K.; Park, S.; Yong, H.K.; Won, K.; Sang, H.L. Immobilization of formate dehydrogenase from Candida boidinii through cross-linked enzyme aggregates. J. Mol. Catal. B Enzym. 2013, 97, 209–214. [Google Scholar] [CrossRef]
- Matijošytė, I.; Arends, I.W.C.E.; Vries, S.D.; Sheldon, R.A. Preparation and use of cross-linked enzyme aggregates (CLEAs) of laccases. J. Mol. Catal. B Enzym. 2010, 62, 142–148. [Google Scholar] [CrossRef]
- Sangeetha, K.; Abraham, T.E. Preparation and characterization of cross-linked enzyme aggregates (CLEA) of Subtilisin for controlled release applications. Int. J. Biol. Macromol. 2008, 43, 314–319. [Google Scholar] [CrossRef] [PubMed]
- Cerdobbel, A.; De, W.K.; Desmet, T.; Soetaert, W. Sucrose phosphorylase as cross-linked enzyme aggregate: Improved thermal stability for industrial applications. Biotechnol. J. 2010, 5, 1192–1197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, C.Y.; Li, X.F.; Lou, W.Y.; Zong, M.H. Cross-linked enzyme aggregates of Mung bean epoxide hydrolases: A highly active, stable and recyclable biocatalyst for asymmetric hydrolysis of epoxides. J. Biotechnol. 2013, 166, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Lanfranchi, E.; Grill, B.; Raghoebar, Z.; Pelt, S.V.; Sheldon, R.; Steiner, K.; Glieder, A.; Winkler, M. Production of hydroxynitrile lyase from D. tyermanii (DtHNL) in Komagataella phaffii and its immobilization as CLEA to generate a robust biocatalyst. Chembiochem 2017, 19, 312–316. [Google Scholar] [CrossRef] [PubMed]
- Mafra, A.C.O.; Kopp, W.; Beltrame, M.B.; Giordano, R.D.L.C.; Tardioli, P.W. Diffusion effects of bovine serum albumin on cross-linked aggregates of catalase. J. Mol. Catal. B Enzym. 2016, 133, 107–116. [Google Scholar] [CrossRef]
- Talekar, S.; Joshi, A.; Joshi, G.; Kamat, P.; Haripurkar, R.; Kambale, S. Parameters in preparation and characterization of cross linked enzyme aggregates (CLEAs). RSC Adv. 2013, 3, 12485–12511. [Google Scholar] [CrossRef]
- Periyasamy, K.; Santhalembi, L.; Mortha, G.; Aurousseau, M.; Subramanian, S. Carrier-free co-immobilization of xylanase, cellulase and β-1,3-glucanase as combined cross-linked enzyme aggregates (combi-CLEAs) for one-pot saccharification of sugarcane bagasse. RSC Adv. 2016, 6, 32849–32857. [Google Scholar] [CrossRef]
- Hanefeld, U.; Gardossi, L.; Magner, E. Understanding enzyme immobilisation. Chem. Soc. Rev. 2009, 38, 453–468. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Jiang, Q.; Sun, L.; Li, Q.; Zhou, L.; Chen, Q.; Li, S.; Yu, M.; Li, W. Magnetic Combined Cross-Linked Enzyme Aggregates of Ketoreductase and Alcohol Dehydrogenase: An Efficient and Stable Biocatalyst for Asymmetric Synthesis of (R)-3-Quinuclidinol with Regeneration of Coenzymes In Situ. Catalysts 2018, 8, 334. [Google Scholar] [CrossRef]
- Taboada-Puig, R.; Junghanns, C.; Demarche, P.; Moreira, M.T.; Feijoo, G.; Lema, J.M.; Agathos, S.N. Combined cross-linked enzyme aggregates from versatile peroxidase and glucose oxidase: Production, partial characterization and application for the elimination of endocrine disruptors. Bioresour. Technol. 2011, 102, 6593–6599. [Google Scholar] [CrossRef] [PubMed]
- Jung, D.-H.; Jung, J.-H.; Seo, D.-H.; Ha, S.-J.; Kweon, D.-K.; Park, C.-S. One-pot bioconversion of sucrose to trehalose using enzymatic sequential reactions in combined cross-linked enzyme aggregates. Bioresour. Technol. 2013, 130, 801–804. [Google Scholar] [CrossRef] [PubMed]
- Jian, D.C.; Shi, R.J. Optimization protocols and improved strategies of cross-linked enzyme aggregates technology: Current development and future challenges. Crit. Rev. Biotechnol. 2015, 35, 15–28. [Google Scholar] [CrossRef]
- Bilal, M.; Asgher, M.; Iqbal, H.M.N.; Hu, H.; Zhang, X. Bio-based degradation of emerging endocrine-disrupting and dye-based pollutants using cross-linked enzyme aggregates. Environ. Sci. Pollut. Res. 2017, 24, 7035–7041. [Google Scholar] [CrossRef] [PubMed]
- Tandjaoui, N.; Tassist, A.; Abouseoud, M.; Couvert, A.; Amrane, A. Preparation and characterization of cross-linked enzyme aggregates (CLEAs) of Brassica rapa peroxidase. Biocatal. Agric. Biotechnol. 2015, 4, 208–213. [Google Scholar] [CrossRef]
- Dalal, S.; Kapoor, M.; Gupta, M.N. Preparation and characterization of combi-CLEAs catalyzing multiple non-cascade reactions. J. Mol. Catal. B Enzym. 2007, 44, 128–132. [Google Scholar] [CrossRef]
- Andreazza, R.; Pieniz, S.; Okeke, B.; Camargo, F.A. Evaluation of copper resistant bacteria from vineyard soils and mining waste for copper biosorption. Braz. J. Microbiol. 2011, 42, 66–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pchelintsev, N.A.; Youshko, M.I.; Švedas, V.K. Quantitative characteristic of the catalytic properties and microstructure of cross-linked enzyme aggregates of penicillin acylase. J. Mol. Catal. B Enzym. 2009, 56, 202–207. [Google Scholar] [CrossRef]
- Richards, F.M.; Knowles, J.R. Glutaraldehyde as a protein cross-linkage reagent. J. Mol. Biol. 1968, 37, 231–233. [Google Scholar] [CrossRef]
- Dartiguenave, M.C.; Bertrand, M.J.; Waldron, K.C. Glutaraldehyde: Behavior in aqueous solution, reaction with proteins, and application to enzyme crosslinking. Biotechniques 2004, 37, 790–802. [Google Scholar] [CrossRef]
- Wine, Y.; Cohenhadar, N.; Freeman, A.; Frolow, F. Elucidation of the mechanism and end products of glutaraldehyde crosslinking reaction by X-ray structure analysis. Biotechnol. Bioeng. 2010, 98, 711–718. [Google Scholar] [CrossRef] [PubMed]
- Mateo, C.; Palomo, J.M.; Van Langen, L.M.; Van Rantwijk, F.; Sheldon, R.A. A new, mild cross-linking methodology to prepare cross-linked enzyme aggregates. Biotechnol. Bioeng. 2004, 86, 273–276. [Google Scholar] [CrossRef] [PubMed]
- Zhen, Q.; Wang, M.; Qi, W.; Su, R.; He, Z. Preparation of β-mannanase CLEAs using macromolecular cross-linkers. Catal. Sci. Technol. 2013, 3, 1937–1941. [Google Scholar] [CrossRef]
- Wang, A.; Zhang, F.; Chen, F.; Wang, M.; Li, H.; Zeng, Z.; Xie, T.; Chen, Z. A facile technique to prepare cross-linked enzyme aggregates using p-benzoquinone as cross-linking agent. Korean J. Chem. Eng. 2011, 28, 1090–1095. [Google Scholar] [CrossRef]
- Velasco-Lozano, S.; López-Gallego, F.; Vázquez-Duhalt, R.; Mateos-Díaz, J.C.; Guisán, J.M.; Favela-Torres, E. Carrier-free immobilization of lipase from Candida rugosa with polyethyleneimines by carboxyl-activated cross-linking. Biomacromolecules 2014, 15, 1896–1903. [Google Scholar] [CrossRef] [PubMed]
- Cabana, H.; Ahamed, A.; Leduc, R. Conjugation of laccase from the white rot fungus Trametes versicolor to chitosan and its utilization for the elimination of triclosan. Bioresour. Technol. 2011, 102, 1656–1662. [Google Scholar] [CrossRef] [PubMed]
- Kunjukunju, S.; Roy, A.; Shekhar, S.; Kumta, P.N. Cross-linked enzyme aggregates of alginate lyase: A systematic engineered approach to controlled degradation of alginate hydrogel. Int. J. Biol. Macromol. 2018, 115, 176–184. [Google Scholar] [CrossRef] [PubMed]
- Mehde, A.A.; Mehdi, W.A.; Özacar, M.; Özacar, Z.Z. Evaluation of different saccharides and chitin as eco-friendly additive to improve the magnetic cross-linked enzyme aggregates (CLEAs) activities. Int. J. Biol. Macromol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Liu, L.; Chen, D.; Wang, Y.; Zhang, J.; Shao, L. Co-expression of the recombined alcohol dehydrogenase and glucose dehydrogenase and cross-linked enzyme aggregates stabilization. Bioresour. Technol. 2016, 224, 531–535. [Google Scholar] [CrossRef] [PubMed]
- Torabizadeh, H.; Tavakoli, M.; Safari, M. Immobilization of thermostable α-amylase from Bacillus licheniformis by cross-linked enzyme aggregates method using calcium and sodium ions as additives. J. Mol. Catal. B Enzym. 2014, 108, 13–20. [Google Scholar] [CrossRef]
- Barbosa, O.; Ortiz, C.; Berenguer-Murcia, A.; Torres, R.; Rodrigues, R.C.; Fernandez-Lafuente, R. ChemInform Abstract: Glutaraldehyde in Bio-Catalysts Design: A Useful Crosslinker and a Versatile Tool in Enzyme Immobilization. Cheminform 2013, 4, 1583–1600. [Google Scholar] [CrossRef]
- Reshmi, R.; Sugunan, S. Improved biochemical characteristics of crosslinked β-glucosidase on nanoporous silica foams. J. Mol. Catal. B Enzym. 2013, 85, 111–118. [Google Scholar] [CrossRef]
- Šulek, F.; Fernández, D.P.; Knez, Ž; Habulin, M.; Sheldon, R.A. Immobilization of horseradish peroxidase as crosslinked enzyme aggregates (CLEAs). Process Biochem. 2011, 46, 765–769. [Google Scholar] [CrossRef]
- Liao, Q.; Du, X.; Jiang, W.; Tong, Y.; Zhao, Z.; Fang, R.; Feng, J.; Tang, L. Cross-linked enzyme aggregates (CLEAs) of halohydrin dehalogenase from Agrobacterium radiobacter AD1: Preparation, characterization and application as a biocatalyst. J. Biotechnol. 2018, 272–273, 48–55. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.I.; Kim, J.; Lee, J.; Jia, H.; Na, H.B.; Youn, J.K.; Kwak, J.H.; Dohnalkova, A.; Grate, J.W.; Wang, P. Crosslinked enzyme aggregates in hierarchically-ordered mesoporous silica: A simple and effective method for enzyme stabilization. Biotechnol. Bioeng. 2007, 96, 210–218. [Google Scholar] [CrossRef] [PubMed]
- Jung, D.; Paradiso, M.; Wallacher, D.; Brandt, A.; Hartmann, M. Formation of Cross-Linked Chloroperoxidase Aggregates in the Pores of Mesocellular Foams: Characterization by SANS and Catalytic Properties. Chemsuschem 2010, 2, 161–164. [Google Scholar] [CrossRef] [PubMed]
- Rajendhran, J.; Gunasekaran, P. Application of cross-linked enzyme aggregates of Bacillus badius penicillin G acylase for the production of 6-aminopenicillanic acid. Lett. Appl. Microbiol. 2007, 44, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Arsenault, A.; Cabana, H.; Jones, J.P. Laccase-Based CLEAs: Chitosan as a Novel Cross-Linking Agent. Enzyme Res. 2011, 2011, 376015. [Google Scholar] [CrossRef] [PubMed]
- Ji, Q.; Wang, B.; Tan, J.; Zhu, L.; Li, L. Immobilized multienzymatic systems for catalysis of cascade reactions. Process Biochem. 2016, 51, 1193–1203. [Google Scholar] [CrossRef]
- Mateo, C.; Chmura, A.; Rustler, S.; Rantwijk, F.V.; Stolz, A.; Sheldon, R.A. Synthesis of enantiomerically pure (S)-mandelic acid using an oxynitrilase–nitrilase bienzymatic cascade: A nitrilase surprisingly shows nitrile hydratase activity. Tetrahedron Asymmetry 2006, 17, 320–323. [Google Scholar] [CrossRef]
- Talekar, S.; Pandharbale, A.; Ladole, M.; Nadar, S.; Mulla, M.; Japhalekar, K.; Pattankude, K.; Arage, D. Carrier free co-immobilization of alpha amylase, glucoamylase and pullulanase as combined cross-linked enzyme aggregates (combi-CLEAs): A tri-enzyme biocatalyst with one pot starch hydrolytic activity. Bioresour. Technol. 2013, 147, 269–275. [Google Scholar] [CrossRef] [PubMed]
- Ahumada, K.; Urrutia, P.; Illanes, A.; Wilson, L. Production of combi-CLEAs of glycosidases utilized for aroma enhancement in wine. Food Bioprod. Process. 2015, 94, 555–560. [Google Scholar] [CrossRef]
- Stressler, T.; Ewert, J.; Eisele, T.; Fischer, L. Cross-linked enzyme aggregates (CLEAs) of PepX and PepN–production, partial characterization and application of combi-CLEAs for milk protein hydrolysis. Biocatal. Agric. Biotechnol. 2015, 4, 752–760. [Google Scholar] [CrossRef]
- Mahmod, S.S.; Yusof, F.; Shah, H.; Jami, M.S.; Khanahmadi, S. Development of an immobilized biocatalyst with lipase and protease activities as a multipurpose cross-linked enzyme aggregate (multi-CLEA). Process Biochem. 2015, 50, 2144–2157. [Google Scholar] [CrossRef]
- Li, H.; Xiao, W.; Xie, P.; Zheng, L. Co-immobilization of enoate reductase with a cofactor-recycling partner enzyme. Enzyme Microb. Technol. 2018, 109, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.T.; Yang, K.L. Combined cross-linked enzyme aggregates of horseradish peroxidase and glucose oxidase for catalyzing cascade chemical reactions. Enzyme Microb. Technol. 2017, 100, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Ning, C.; Su, E.; Tian, Y.; Wei, D. Combined cross-linked enzyme aggregates (combi-CLEAs) for efficient integration of a ketoreductase and a cofactor regeneration system. J. Biotechnol. 2014, 184, 7–10. [Google Scholar] [CrossRef] [PubMed]
- Sirimanne, S.R.; Patterson, D.G., Jr. A one-pot synthesis of (±)-(ring 13C6)-mandelic acid. J. Label. Compd. Radiopharm. 1993, 33, 725–731. [Google Scholar] [CrossRef]
- Gao, J.; Wang, A.R.; Jiang, X.P.; Zhang, J.X.; Zhang, Y.W. Preparation of Expoxy-Functionalized Magnetic Nanoparticles for Immobilization of Glycerol Dehydrogenase. J. Nanosci. Nanotechnol. 2018, 4852–4857. [Google Scholar] [CrossRef] [PubMed]
- Ahumada, K.; Martínez-Gil, A.; Moreno-Simunovic, Y.; Illanes, A.; Wilson, L. Aroma Release in Wine Using Co-Immobilized Enzyme Aggregates. Molecules 2016, 21, 1485. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.K.; Zhang, Y.W.; Nguyen, N.P.; Jeya, M.; Lee, J.K. Covalent immobilization of β-1,4-glucosidase from Agaricus arvensis onto functionalized silicon oxide nanoparticles. Appl. Microbiol. Biotechnol. 2010, 89, 337–344. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
The reaction of polymeric glutaraldehyde with lysine residues protein in alkaline conditions (A) and acidic conditions (B).
Figure 1.
The reaction of polymeric glutaraldehyde with lysine residues protein in alkaline conditions (A) and acidic conditions (B).

Figure 2.
The one-step bioconversion of sucrose to trehalose with combi-CLEAs of AS, MTS, and MTH.
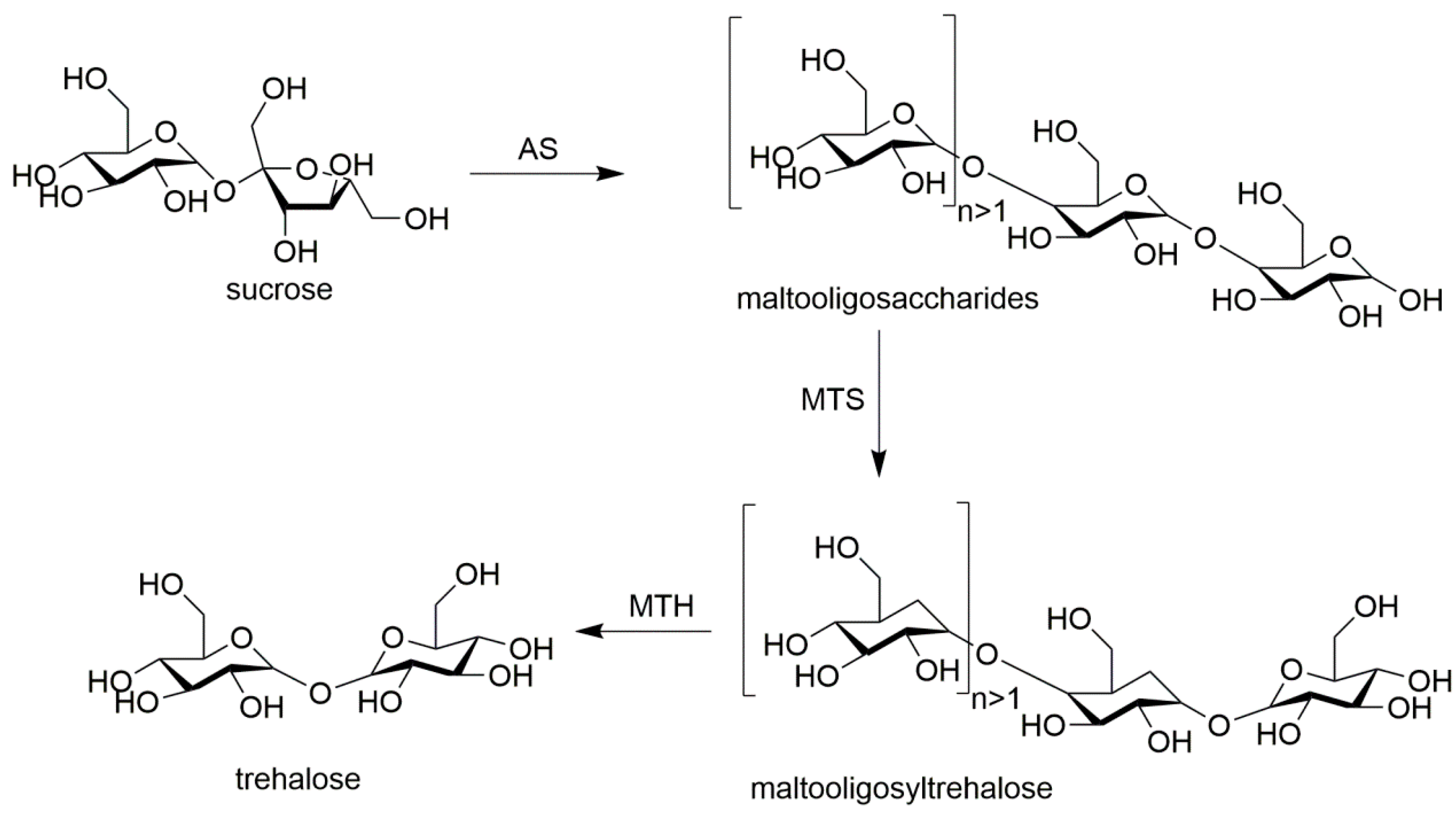
Figure 3.
The Biocatalytic (A) and Chemical (B) synthesis of mandelic acid from benzaldehyde.

Figure 4.
The combi-CLEAs of amylase, glucoamylase, and pullulanase for hydrolyzing of starch.

Figure 5.
The combi-CLEAs of L-arabinose and β--glucosidase involved in aroma release in wine.

Figure 6.
The combi-CLEAs of PepX and PepN.

Figure 7.
The combi-CLEAs of eductases and glucose dehydrogenase for cofactor regeneration system.
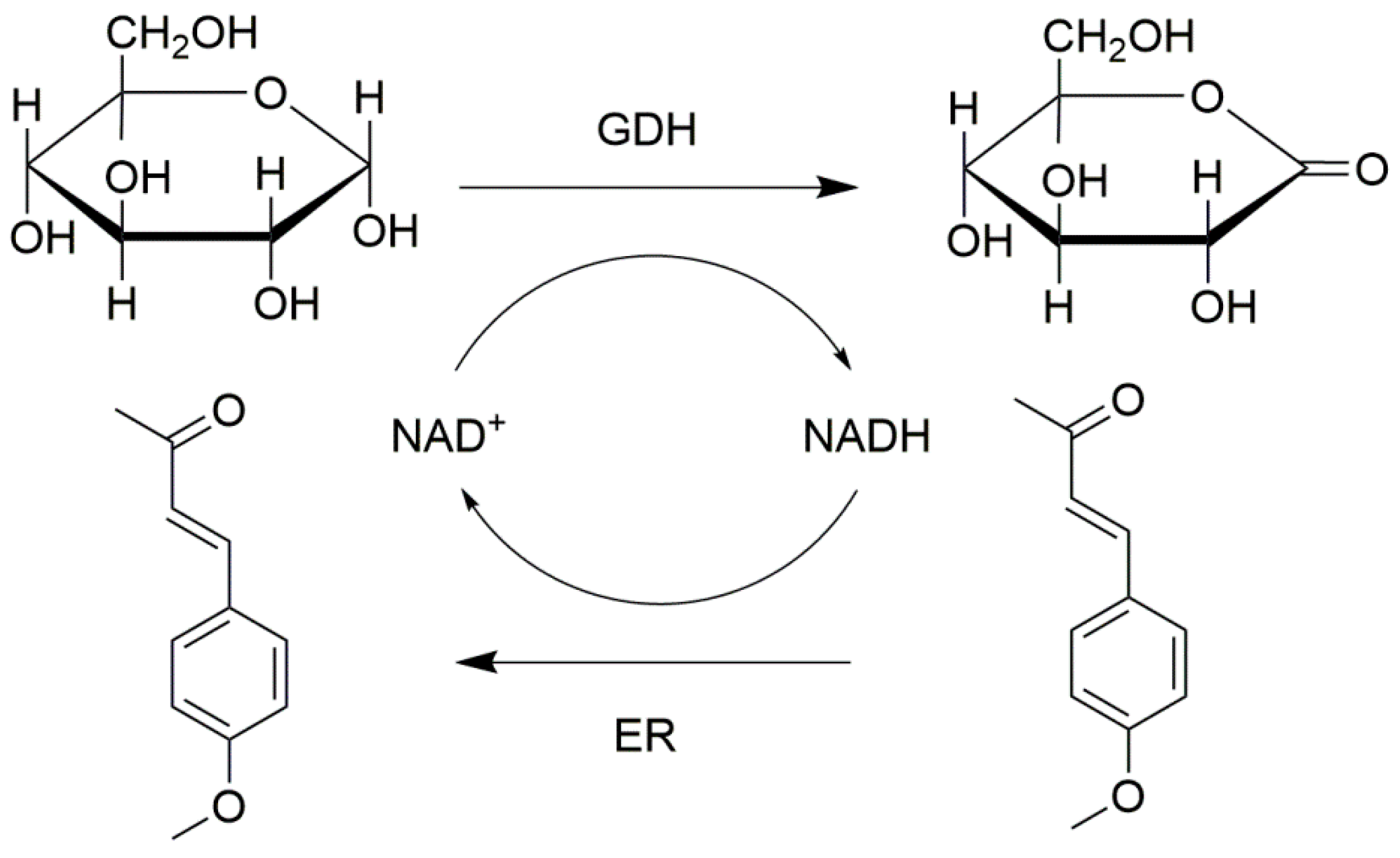
Figure 8.
The combi-CLEAs of glucose oxidase and horseradish peroxidase.

Figure 9.
Combi-CLEAs of ADH and GDH for cofactor regeneration system.

Figure 10.
The combi-CLEAs of ketoreductase and D-glucose dehydrogenase for the cofactor regeneration system.
Figure 10.
The combi-CLEAs of ketoreductase and D-glucose dehydrogenase for the cofactor regeneration system.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
The factors for preparing CLEAs.
| Enzymes | Factors | References | |||
|---|---|---|---|---|---|
| Enzymes Proportion | Precipitants | Cross-Linker | Cross-Linking Temperature | ||
| Glucose oxidase and versatile peroxidase | 10:7 (mass) | Polyethylene glycol | Glutaraldehyde | 30 °C | [71] |
| Amylosucrase, maltooligosyltrehalose synthase and maltooligosyltrehalose trehalohydrolase | 16:1:1 (mass) | Acetone | Glutaraldehyde | 4 °C | [72] |
| Xylanase and mannanase | 1:1 (mass) | Acetone | Glutaraldehyde | 37 °C | [14] |
| Lipase, α-amylase and phospholipase A2 | NA | Dimethoxyethane | Glutaraldehyde | 4 °C | [76] |
| Amylase, glucoamylase and pullulanase | 3:3:1 (activity) | Ammonium sulfate | Glutaraldehyde | 35 °C | [101] |
| α-l-arabinosidase and β-d-glucosidase | NA | Ammonium sulfate | Glutaraldehyde | 4 °C | [102] |
| X-prolyl-dipeptidyl aminopeptidase and general aminopeptidase N | 1:1 (mass) | Ammonium sulfate | Glutaraldehyde | Ice | [103] |
| Lipase and protease | NA | Ammonium sulfate | Glutaraldehyde | 4 °C | [104] |
| Eductases and glucose dehydrogenase | 1.1:5 (activity) | Ammonium sulfate | Oxidized dextran | 4 °C | [105] |
| Glucose oxidase and horseradish peroxidase | 150:1 (mass) | Acetonitrile | Glutaraldehyde | NA | [106] |
| Ketoreductase and D-glucose dehydrogenase | 1:1(mass) | 1,2-Dimethoxyethane | Glutaraldehyde | 20 °C | [107] |
NA: Not available.
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Xu, M.-Q.; Wang, S.-S.; Li, L.-N.; Gao, J.; Zhang, Y.-W. Combined Cross-Linked Enzyme Aggregates as Biocatalysts. Catalysts 2018, 8, 460. https://doi.org/10.3390/catal8100460
AMA Style
Xu M-Q, Wang S-S, Li L-N, Gao J, Zhang Y-W. Combined Cross-Linked Enzyme Aggregates as Biocatalysts. Catalysts. 2018; 8(10):460. https://doi.org/10.3390/catal8100460
Chicago/Turabian StyleXu, Meng-Qiu, Shuang-Shuang Wang, Li-Na Li, Jian Gao, and Ye-Wang Zhang. 2018. "Combined Cross-Linked Enzyme Aggregates as Biocatalysts" Catalysts 8, no. 10: 460. https://doi.org/10.3390/catal8100460
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.